Introduction
Epithelial ovarian cancer (EOC), which is commonly
diagnosed at an advanced disease stage due to the absence of
specific symptoms and the lack of early detection markers, is the
most lethal gynecological malignancy (1). Primary cytoreductive surgery and
platinum-based chemotherapy constitute the fundamental treatment
for EOC, with 70–80% of patients achieving complete remission
(2). Unfortunately, 60–70% of EOC
patients eventually undergo relapse, resulting in mortality that is
frequently ascribed to the development of chemoresistance (3,4). In
addition, the mechanism underlying chemoresistance in EOC remains
unclear.
Mesenchymal stem cells (MSCs), also called
multipotent mesenchymal stromal cells, comprise a heterogeneous
group of adult progenitor cells with differentiation capacities.
These cells may be easily derived and propagated from a variety of
sources, including bone marrow and adipose tissue (5). Accumulating evidence has demonstrated
that MSCs serve an essential role in tumor development and
progression (6–8). In addition, MSCs are receiving
attention as potential therapeutic agents with unique immunological
properties that allow them to survive in a xenogeneic environment
and exert a powerful potential for repairing injured tissues in
cancer and regenerative medicine (9).
The omentum, which is primarily composed of adipose
tissue and milky spots with islands of compact immune cells, is the
most common site for EOC metastatic spread and is invariably
involved in cases of advanced ovarian cancer (10). Interactions between ovarian cancer
and the omental microenvironment are an important driver of
clinical outcome in patients with EOC (11). Previous studies have demonstrated
that adipose tissue contains an abundance of mesenchymal progenitor
cells, which promote tumor progression (12–14).
Furthermore, it has been reported that the omentum is an
independent predictor of the response of ovarian cancer to
chemotherapy (15,16). In the present study, the effects of
primary omental adipose-derived mesenchymal stem cells (ADSCs) on
cisplatin-induced apoptosis in EOC cells were investigated, to
determine the contribution of ADSCs to chemoresistance.
Materials and methods
Cell lines
Human epithelial ovarian carcinoma cell lines ES2
(clear cell carcinoma) and SKOV3 (adenocarcinoma) were purchased
from the Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). Primary ADSCs were isolated from three healthy
adult female donors and were identified as described in a previous
study (17). All donors provided
written informed consent, and the present study was approved by the
Ethics Committee of Union Hospital, Tongji Medical College,
Huazhong University of Science and Technology (Wuhan, China),
approval no. 2014073. Primary ADSCs were cultured in Dulbecco's
modified Eagle's medium (DMEM)/F12 supplemented with 10% fetal
bovine serum (FBS) (both from Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA), 100 mg/ml penicillin and 100 mg/ml
streptomycin. The supernatants of ADSCs (passage 5 to 20) cultures
were harvested every 48–72 h, and the harvested conditioned medium
was centrifuged at 4°C at 300 × g for 5 min, filtered through a
0.22 µm syringe filter and stored at 4°C. ES2 and SKOV3 cells were
cultured in DMEM/F12 supplemented with 10% FBS or ADSC-derived
conditioned medium for 10 days. All cell lines were cultured at
37°C in a humidified atmosphere containing 5% CO2.
MTT assay
The effect of ADSC-derived conditioned medium on EOC
cells was assessed via the MTT assay. EOC cells were seeded in
96-well plates (Corning Life Sciences, Corning, NY, USA) at a
density of 5×103 cells/well. Following attachment, the
cells were treated with various concentrations of cisplatin (0, 10,
30, 60, 90 and 120 µM for ES2 cells; 0, 20, 40, 80, 120, 160 and
200 µM for SKOV3 cells) for 48 h followed by incubation with 20 µl
MTT reagent (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at a
final concentration of 0.5 mg/ml at 37°C for 4 h. The medium was
removed and 100 µl pure dimethyl sulfoxide was added to each well.
Wells without cells containing culture medium were used as blank
controls, and wells with cells treated with media without cisplatin
served as negative controls. The absorbance was measured at a
wavelength of 490 nm using a microplate reader (model iMark-10601;
Bio-Rad Laboratories, Inc., Hercules, CA, USA). Cell survival
curves were generated, from which the half maximal inhibitory
concentration (IC50) values of cisplatin in EOC cells
under different culture conditions were derived. Each assay was
performed in triplicate.
Hoechst 33342 staining
EOC cells (1×104 cells/well) were seeded
in 48-well plates (Corning Life Sciences), exposed to cisplatin (at
the IC50 value) at 37°C for 48 h and then fixed with
3.7% paraformaldehyde for 10 min at room temperature. Following
washing twice with cold PBS, the cells were stained for 5 min with
Hoechst 33342 (Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C
in the dark. Following washing, the cells were observed under a
fluorescence microscope (model IX70; Olympus Corporation, Tokyo,
Japan). The Hoechst reagent is taken up by nuclei, and the nuclei
of apoptotic cells exhibit bright blue fluorescence. Each
experiment was repeated three times separately.
Flow cytometric assay for
apoptosis
ES2 and SKOV3 cells (1×105 cells/well)
were seeded in 6-well plates (Corning Life Sciences), grown to
confluence and exposed to cisplatin at the IC50 value at
37°C for 48 h. The cells were then collected using trypsin without
EDTA, washed twice with PBS. Subsequently, FITC Annexin V Apoptosis
Detection kit (catalog no. 556547; BD Biosciences, San Jose, CA,
USA) was used to detect apoptosis. According to the manufacturer's
protocol, cells were resuspended in binding buffer (FITC Annexin V
Apoptosis Detection kit; BD Biosciences), and mixed with Annexin
V-fluorescein isothiocyanate and propidium iodide at room
temperature in the dark for 15 min. Subsequently, the cells were
analyzed by flow cytometry (FACSCalibur; BD Biosciences). Each
experiment was repeated at least three times separately, and
CellQuest™ analysis program software, version 5.1 (BD
Biosciences) was used to analyze the results.
Western blot analysis
Total protein from cultured cells was extracted
using a radioimmunoprecipitation assay buffer (Beyotime Institute
of Biotechnology, Haimen, China) containing a protease inhibitor
cocktail and quantified using a Bradford assay (Beyotime Institute
of Biotechnology) according to the manufacturer's protocol.
Proteins (30 µg) were separated by 10% SDS-PAGE and transferred
onto nitrocellulose membranes. The membranes were blocked with 5%
non-fat dry milk for 1 h at room temperature and incubated
overnight at 4°C with the following primary antibodies: Rabbit
anti-caspase-3 polyclonal antibody (cat no. 9665, 1:1,500 dilution;
Cell Signaling Technology, Inc., Danvers, MA, USA) and mouse
anti-β-actin polyclonal antibody (cat no. sc-130301, 1:500
dilution; Santa Cruz Biotechnology, Inc., Dallas, TX, USA).
Following washing with Tris-buffered saline containing 0.1% Tween,
the membranes were incubated with a horse radish
peroxidase-conjugated goat-anti-rabbit (cat no. sc 2004, 1:5,000
dilution) or goat-anti-mouse secondary antibody (cat no. sc-516102,
1:5,000 dilution) (both from Santa Cruz Biotechnology, Inc.) for 1
h at room temperature and visualized using an Enhanced
Chemiluminescence system (Beyotime Institute of Biotechnology). The
protein bands were quantified using Quantity One v4.62 software
(Bio-Rad Laboratories, Inc.). Each experiment was repeated at least
three times separately.
Atomic absorption spectroscopy for
detection of platinum in tumor cells
For the determination of the intracellular platinum
concentration, EOC cells were resuspended in PBS and divided into
two equal aliquots: One for protein content measurement and the
other for platinum determination. The protein content was measured
using the Bradford assay (Beyotime Institute of Biotechnology). For
platinum determination, cells were incubated with 0.2% nitric acid
at room temperature for 1 h. The intracellular platinum
concentrations were measured by flameless atomic absorption
spectrometry (AAS; Varian SpectrAA 240FS, Palo Alto, CA, USA) using
a standard curve covering a range of 60–240 µg/. The intracellular
platinum levels are expressed as ng of platinum per mg of protein.
All samples were analyzed three times in duplicate.
Statistical analysis
All data are expressed as the mean ± standard
deviation (n≥3). The statistical significance of differences
between two groups was analyzed using a two-tailed Students t-test
via SPSS statistical software package, version 16.0 (SPSS, Inc.,
Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
ADSCs decrease the sensitivity of EOC
cells to cisplatin
To investigate whether ADSCs modulate the
sensitivity of EOC cells to cisplatin, ES2 and SKOV3 cells were
pretreated with conditioned medium derived from ADSCs for 10 days
(named co-cultured ES2 and SKOV3 cells). The sensitivities of the
co-cultured cells and their parental cells to cisplatin were then
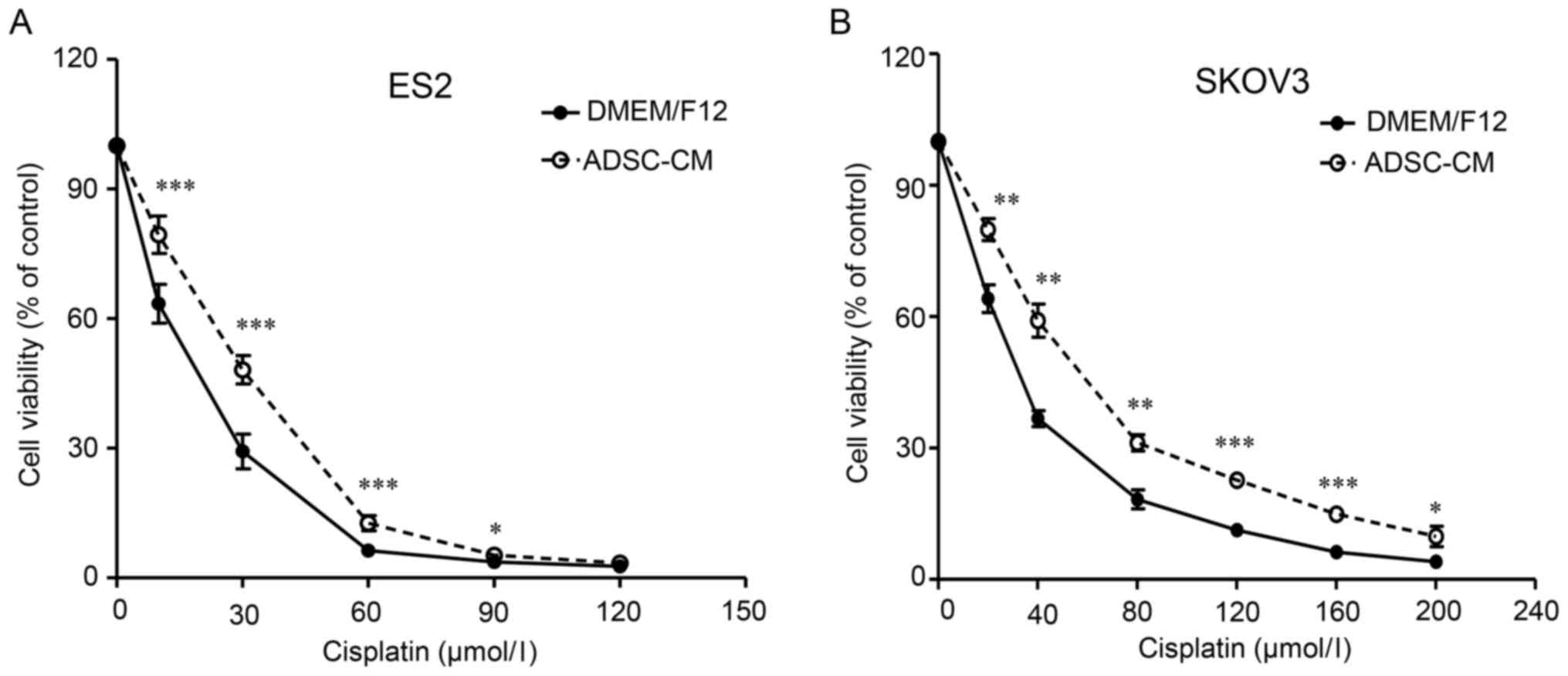
assessed using the MTT assay. As demonstrated in Fig. 1, the co-cultured SKOV3 and ES2
cells exhibited a right-shifted dose-survival curve and became more
resistant to cisplatin (Fig. 1).
The IC50 values for cisplatin of the co-cultured ES2 and
SKOV3 cells were 1.60- and 1.71-fold higher, respectively, than
that of the parental cells (Table
I). These findings suggested that ADSCs decrease the
sensitivity of EOC cells to cisplatin.
 | Table I.Sensitivity of ovarian cancer cell
lines to cisplatin. |
Table I.
Sensitivity of ovarian cancer cell
lines to cisplatin.
|
| Cisplatin |
|---|
|
|
|
|---|
| Cell lines | IC50
(µmol/l) | RI |
|---|
| ES2/DMEM/F12 | 14.82±0.98 | 1.00 |
| ES2/ADSC-CM |
23.70±0.97a | 1.60 |
| SKOV3/DMEM/F12 | 29.06±4.29 | 1.00 |
| SKOV3/ADSC-CM |
49.80±4.51a | 1.71 |
ADSCs protect EOC cells from
cisplatin-induced apoptosis
To further determine whether ADSCs protect EOC cells
against cisplatin-induced apoptosis, co-cultured ES2 and SKOV3
cells and parental cells were exposed to cisplatin for 48 h at
their IC50 doses based on the MTT assay. Hoechst 33342
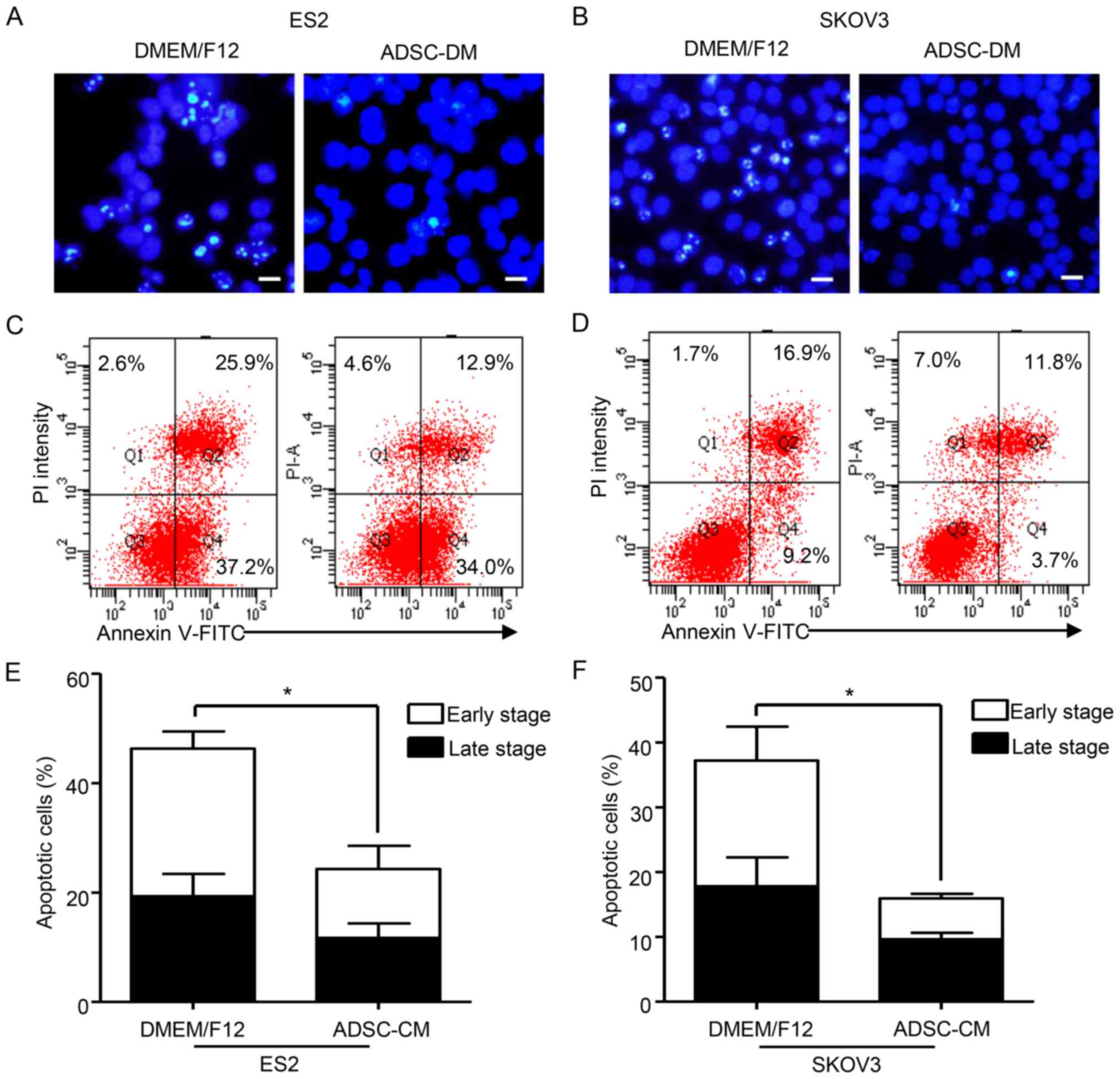
staining was then performed to analyze cisplatin-induced apoptosis.
According to fluorescence microscopy images, the co-cultured ES2
and SKOV3 cells revealed significantly lower nuclear fragmentation
compared with the parental cells (Fig.
2A and B). Consistently, Annexin V/propidium iodide dual
staining followed by flow cytometric analysis demonstrated that
ADSC-treated supernatants exhibited a significantly decreased
proportion of apoptotic ES2 and SKOV3 cells following exposure to
cisplatin. Compared with the parental cells, the proportion of
apoptotic cells among the co-cultured ES2 and SKOV3 cells decreased
by 1.9- and 2.33-fold, respectively (Fig. 2C-F). In addition, the proportion of
co-cultured ES2 and SKOV3 cells in the early phase of apoptosis was
2.14- and 3.01-fold lower, respectively, than the parental cells
(Fig. 2E and F). To further
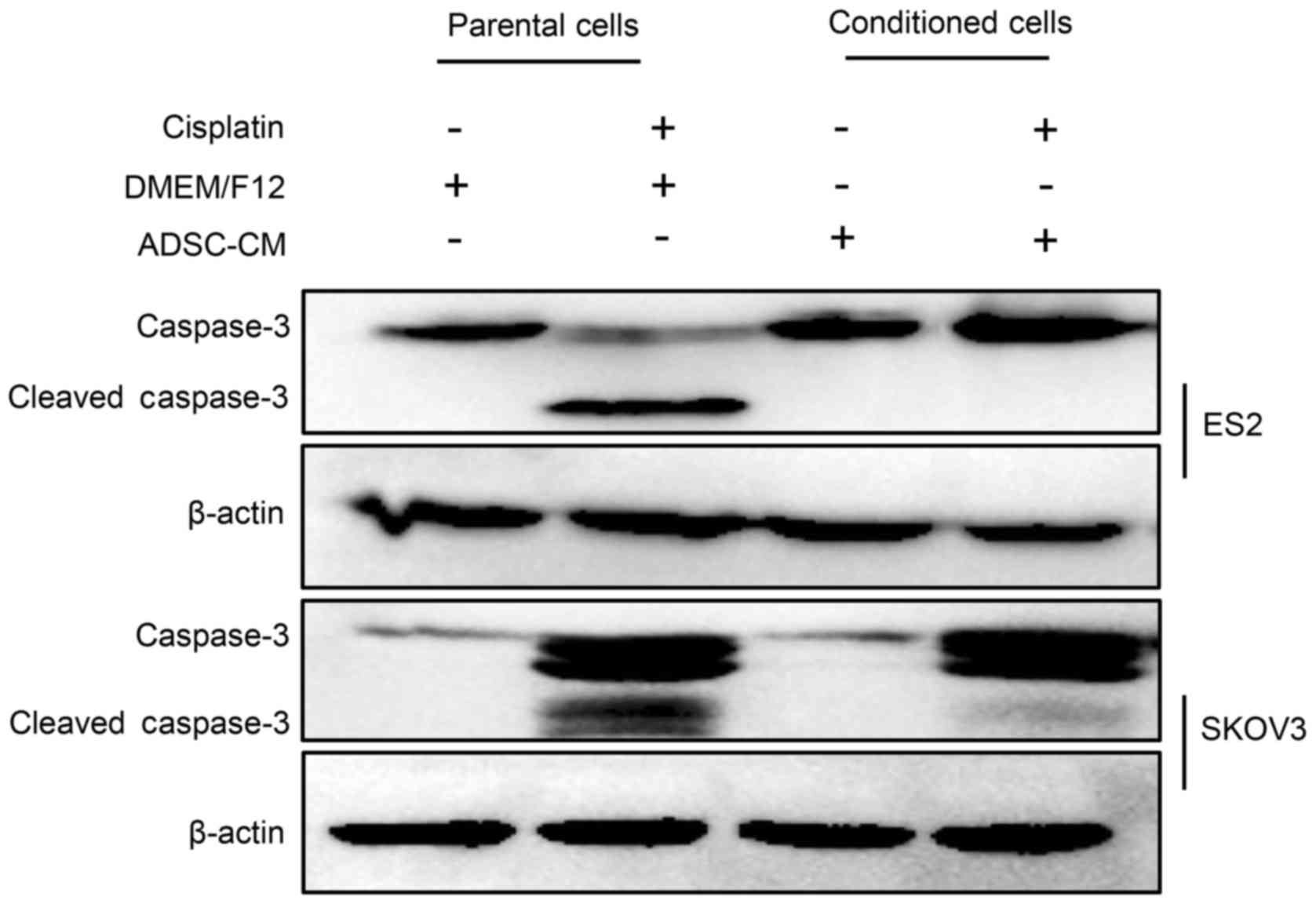
explore the molecular mechanisms underlying the ADSC-mediated
protection of EOC cells against cisplatin-induced apoptosis,
caspase-3 was evaluated by western blot analysis. As presented in
Fig. 3, after cisplatin treatment,
the expression levels of cleaved caspase-3 in conditioned cells
were significantly reduced in the co-cultured ES2 and SKOV3 cells
compared with the parental cells. These findings suggested that
ADSCs may reduce cisplatin-induced apoptosis in EOC cells by
inhibiting caspase-3 activity.
ADSCs decrease intracellular platinum
accumulation in EOC cells
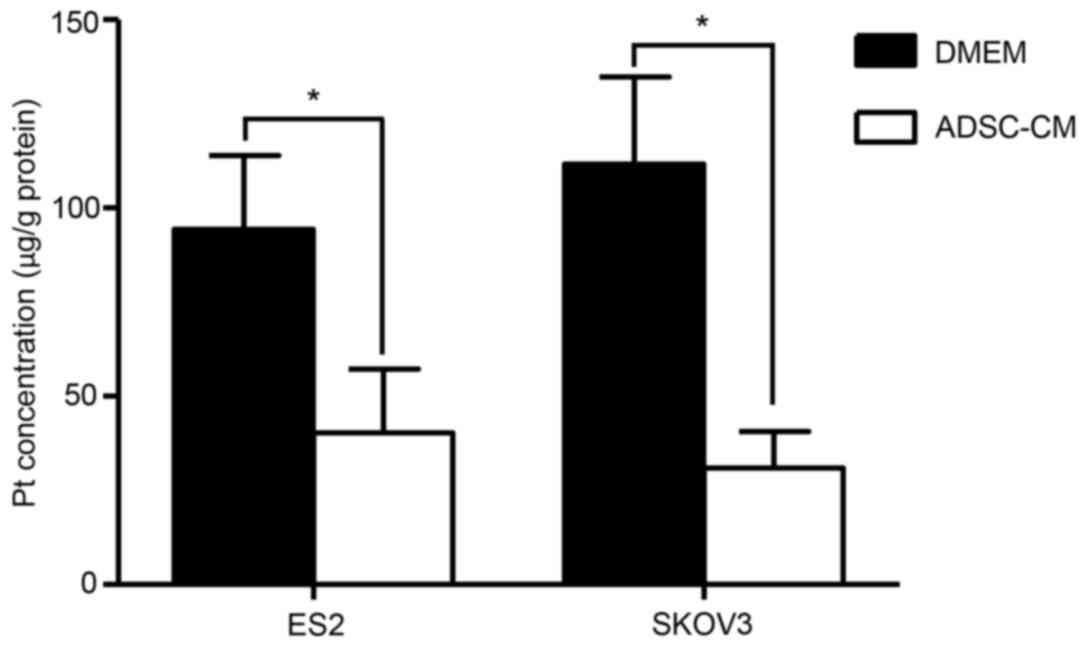
Given that decreased intracellular drug accumulation
is a critical mechanism of chemoresistance, the effect of ADSCs on
intracellular platinum accumulation in EOC cells was investigated
using atomic absorption spectroscopy assays. As presented in
Fig. 4, following treatment with
cisplatin for 48 h at their IC50 doses, platinum
concentrations in the co-cultured ES2 and SKOV3 cells were at least
2.34 and 3.60-fold lower than that the control cells, respectively
(Fig. 4). These results suggested
that ADSCs may decrease the intracellular platinum concentration in
EOC cells.
Discussion
Resistance to platinum-based chemotherapy remains of
primary concern in the treatment of EOC. In the present study, it
was demonstrated that ADSCs confer cisplatin resistance to EOC
cells via inhibition of cisplatin-induced apoptosis and decreased
intracellular platinum accumulation. These findings may provide a
novel insight into the mechanism of chemoresistance in ovarian
cancer.
The MSCs recruited to the tumor microenvironment
serve various roles in tumor growth and metastasis (18). The presence of environment-mediated
drug resistance (EMDR) suggests an adaptive, reciprocal signaling
connection between tumor cells and the surrounding
microenvironment, and MSCs are currently regarded as a primary
contributor to the development of drug resistance in tumor cells
(19). In the present study, ADSCs
were isolated from the human omentum and it was identified that
indirect co-culture with ADSCs significantly inhibited
cisplatin-induced apoptosis accompanied by a decreased level of
cleaved caspase-3. Similarly, a previous study reported that the
conditioned medium of bone marrow-derived MSCs may protect healthy
renal cells from apoptosis induced by cisplatin in vitro
(20). MSCs seem to provide non
cell-specific protection against cisplatin induced apoptosis in
both cancerous and non-cancerous cells. Research has also indicated
that exosomes released by human umbilical cord MSCs may protect
against cisplatin-induced renal oxidative stress and apoptosis
(21). These findings are
consistent with the results of the present study.
In the present study, the conditioned medium of
ADSCs was capable of supporting EOC cell survival in the presence
of cisplatin. It has been reported that MSCs associated with EMDR
secrete a series of cytokines and growth factors, including stromal
cell-derived factor-1, interleukin (IL)-6, nitric oxide, IL-3,
granulocyte-colony stimulating factor, macrophage-colony
stimulating factor and granulocyte-macrophage-colony stimulating
factor, which may protect tumor cells against chemotherapy drugs
(22). In addition, bone
marrow-derived MSCs secrete soluble neuregulin 1 to promote the
survival and inhibit the apoptosis of colorectal cancer cells
through human epidermal growth factor receptor
(HER)2/HER3-dependent phosphoinositide 3-kinase/protein kinase B
signaling (23), and MSCs secrete
several fatty acids that confer resistance to platinum chemotherapy
in breast cancer cells (24).
There are several reports to date regarding ADSC paracrine action
on tumor progression. For instance, ADSCs may secrete IL-6 to
promote the tumor-initiating capacity of breast and colon cancers
(25), and hypoxia-induced
secretion of IL-10 from ADSCs may promote tumor growth in Burkitt
lymphoma (26). However, future
studies are required to identify the key mediators linking omental
ADSCs and chemoresistance in EOC.
Cisplatin resistance is regulated by a cascade of
events that interfere with multiple steps involved in its cytotoxic
actions, from initial drug entry into cells to the final stages of
apoptosis. The findings of the present study suggested that ADSCs
regulate both platinum accumulation and platinum-induced apoptosis
in EOC cells, which in turn may lead to tumor cisplatin resistance.
The intracellular cisplatin accumulation is mainly regulated by
copper transporters, including copper transporter 1 and 2,
copper-transporting p-type adenosine triphosphatase 1 and 2
(27). Further investigation to
clarify whether ADSCs regulate the expression of these copper
transporters on EOC cells is needed. However, the results regarding
platinum accumulation are preliminary, as only AAS was used to
assess platinum concentrations. The effect of ADSCs on platinum
accumulation in EOC cells require verification by cisplatin uptake
and efflux assays using a platinum detection method with improved
accuracy, including inductively coupled plasma-mass spectrometry.
In addition, further studies are required to verify whether the
suppression of apoptosis mediated by ADSCs is associated with the
decrease in the intracellular platinum concentration.
In conclusion, ADSCs were identified to be a
contributor to cisplatin resistance, exhibiting an ability to
reduce caspase-3-dependent apoptosis and intracellular platinum
accumulation in EOC cells. These findings may provide evidence of
the link between the tumor microenvironment and
chemotherapy-associated clinical outcomes, and suggest that ADSCs
may serve as a therapeutic target for recurrent EOC. However, the
evidence provided regarding the effects of ADSCs on apoptosis and
platinum accumulation in ovarian cancer cells is preliminary, and
the underlying specific molecular mechanisms remain unclear. The
identification of key factors and pathways involved in the
interaction between ADSCs and tumor cells will help clarify the
role of ADSCs in cisplatin-resistance development in ovarian
cancer.
Acknowledgements
This work was supported by the National Natural
Science Foundation of China (grant no. 81472443).
References
|
1
|
Jayson GC, Kohn EC, Kitchener HC and
Ledermann JA: Ovarian cancer. Lancet. 384:1376–1388. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bookman MA, Brady MF, McGuire WP, Harper
PG, Alberts DS, Friedlander M, Colombo N, Fowler JM, Argenta PA, De
Geest K, et al: Evaluation of new platinum-based treatment regimens
in advanced-stage ovarian cancer: A Phase III Trial of the
Gynecologic Cancer Intergroup. J Clin Oncol. 27:1419–1425. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cannistra SA: Cancer of the ovary. N Engl
J Med. 351:2519–2529. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
McMeekin DS, Tillmanns T, Chaudry T, Gold
M, Johnson G, Walker J and Mannel R: Timing isn't everything: An
analysis of when to start salvage chemotherapy in ovarian cancer.
Gynecol Oncol. 95:157–164. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ilmer M, Vykoukal J, Boiles A Recio,
Coleman M and Alt E: Two sides of the same coin: Stem cells in
cancer and regenerative medicine. FASEB J. 28:2748–2761. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chan JK and Lam PY: Human mesenchymal stem
cells and their paracrine factors for the treatment of brain
tumors. Cancer Gene Ther. 20:539–543. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dittmer J and Leyh B: Paracrine effects of
stem cells in wound healing and cancer progression (Review). Int J
Oncol. 44:1789–1798. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ma S, Xie N, Li W, Yuan B, Shi Y and Wang
Y: Immunobiology of mesenchymal stem cells. Cell Death Differ.
21:216–225. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ghobadi F, Mehrabani D and Mehrabani G:
Regenerative potential of endometrial stem cells: A mini review.
World J Plast Surg. 4:3–8. 2015.PubMed/NCBI
|
|
10
|
Raja FA, Chopra N and Ledermann JA:
Optimal first-line treatment in ovarian cancer. Ann Oncol. 23 Suppl
10:x118–x127. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Weidle UH, Birzele F, Kollmorgen G and
Rueger R: Mechanisms and targets involved in dissemination of
ovarian cancer. Cancer Genomics Proteomics. 13:407–423. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Baer PC: Adipose-derived mesenchymal
stromal/stem cells: An update on their phenotype in vivo and in
vitro. World J Stem Cells. 6:256–265. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kang BJ, Lee SH, Kweon OK and Cho JY:
Differentiation of canine adipose tissue-derived mesenchymal stem
cells towards endothelial progenitor cells. Am J Vet Res.
75:685–691. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Minteer DM, Marra KG and Rubin JP: Adipose
stem cells: Biology, safety, regulation, and regenerative
potential. Clin Plast Surg. 42:169–179. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Arie AB, McNally L, Kapp DS and Teng NN:
The omentum and omentectomy in epithelial ovarian cancer: A
reappraisal: Part II-The role of omentectomy in the staging and
treatment of apparent early stage epithelial ovarian cancer.
Gynecol Oncol. 131:784–790. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ben Arie A, McNally L, Kapp DS and Teng
NN: The omentum and omentectomy in epithelial ovarian cancer: A
reappraisal. Part I-Omental function and history of omentectomy.
Gynecol Oncol. 131:780–783. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chu Y, Tang H, Guo Y, Guo J, Huang B, Fang
F, Cai J and Wang Z: Adipose-derived mesenchymal stem cells promote
cell proliferation and invasion of epithelial ovarian cancer. Exp
Cell Res. 337:16–27. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xue Z, Wu X, Chen X, Liu Y, Wang X, Wu K,
Nie Y and Fan D: Mesenchymal stem cells promote epithelial to
mesenchymal transition and metastasis in gastric cancer though
paracrine cues and close physical contact. J Cell Biochem.
116:618–627. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Meads MB, Gatenby RA and Dalton WS:
Environment-mediated drug resistance: A major contributor to
minimal residual disease. Nat Rev Cancer. 9:665–674. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qi S and Wu D: Bone marrow-derived
mesenchymal stem cells protect against cisplatin-induced acute
kidney injury in rats by inhibiting cell apoptosis. Int J Mol Med.
32:1262–1272. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou Y, Xu H, Xu W, Wang B, Wu H, Tao Y,
Zhang B, Wang M, Mao F, Yan Y, et al: Exosomes released by human
umbilical cord mesenchymal stem cells protect against
cisplatin-induced renal oxidative stress and apoptosis in vivo and
in vitro. Stem Cell Res Ther. 4:342013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Guan J and Chen J: Mesenchymal stem cells
in the tumor microenvironment. Biomed Rep. 1:517–521. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
De Boeck A, Pauwels P, Hensen K, Rummens
JL, Westbroek W, Hendrix A, Maynard D, Denys H, Lambein K, Braems
G, et al: Bone marrow-derived mesenchymal stem cells promote
colorectal cancer progression through paracrine neuregulin 1/HER3
signalling. Gut. 62:550–560. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Roodhart JM, Daenen LG, Stigter EC, Prins
HJ, Gerrits J, Houthuijzen JM, Gerritsen MG, Schipper HS, Backer
MJ, van Amersfoort M, et al: Mesenchymal stem cells induce
resistance to chemotherapy through the release of platinum-induced
fatty acids. Cancer Cell. 20:370–383. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wei HJ, Zeng R, Lu JH, Lai WF, Chen WH,
Liu HY, Chang YT and Deng WP: Adipose-derived stem cells promote
tumor initiation and accelerate tumor growth by interleukin-6
production. Oncotarget. 6:7713–7726. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu L, Wang X, Wang J, Liu D, Wang Y, Huang
Z and Tan H: Hypoxia-induced secretion of IL-10 from
adipose-derived mesenchymal stem cell promotes growth and cancer
stem cell properties of Burkitt lymphoma. Tumour Biol.
37:7835–7842. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Burger H, Loos WJ, Eechoute K, Verweij J,
Mathijssen RH and Wiemer EA: Drug transporters of platinum-based
anticancer agents and their clinical significance. Drug Resist
Updat. 14:22–34. 2011. View Article : Google Scholar : PubMed/NCBI
|