Introduction
Non-alcoholic fatty liver disease (NAFLD) is a
prevalent chronic liver disease, which threatens the health of both
adults and children. Furthermore, NAFLD is also regarded as a risk
factor for metabolism syndromes and is strongly associated with
type 2 diabetes mellitus, obesity and insulin resistance (1). However, the pathophysiology of NAFLD
remains unclear, and many strategies, such as diet control, weight
loss and exercise are difficult to maintain for a long time, and
available pharmacotherapy are still deficient (2). Thus, the development of novel
therapeutics and the identification of novel therapeutic targets
are required.
Notch signaling is a conserved signaling pathway
involved in cell commitment and development (3). Previous research has demonstrated
that Notch-1 can act as an oncogene and is upregulated in cases of
hepatocellular carcinoma (4).
Recently, Notch signaling has been recognized as a key regulator of
energy homeostasis, which is strongly associated with
glycometabolism and lipid accumulation (5). In addition, Notch signaling amplifies
signal transmissions responsible for fat production in cases of
obesity and diabetes. Furthermore, Notch-1 activation enhances
lipogenesis in the liver, resulting in NAFLD, and suppression of
Notch-1 signaling in the liver reduces hepatosteatosis (6). Thus, the Notch signaling pathway may
be a potential therapeutic target for the treatment of NAFLD and
insulin resistance.
Curcumin is the active ingredient of Curcuma
longa, which has various pharmacological activities, including
anti-inflammatory, anti-oxidant and anti-cancer activities
(7). Numerous studies have
demonstrated that administration of curcumin suppresses tumor
growth via inhibition of the Notch signaling pathway in hepatic
cancers (8–11), thus indicating the potential of
using curcumin to target the Notch signaling pathway. Furthermore,
curcumin has also been demonstrated to reduce serum lipids and
glucose levels in animal models (12), however, the underlying molecular
mechanism responsible for said effects remains unclear.
It has previously been revealed that curcumin
downregulates Notch signaling in hepatocellular carcinoma (13), and that this cancer type can also
develop as a result of hepatosteatosis and hepatocirrhosis.
Considering previous findings, the present study aimed to
investigate whether curcumin reduces the development of NAFLD, and
whether this is associated with inhibition of the Notch signaling
pathway. In the present study, rats were fed a high-fat diet and
were used to investigate the effects of curcumin on hepatic
steatosis and insulin resistance. Furthermore, Notch signaling and
associated metabolic targets were investigated to determine the
potential mechanisms involved.
Materials and methods
Chemicals and reagents
Curcumin was purchased from Sigma-Aldrich; Merck
KGaA (Darmstadt, Germany), dissolved in dimethyl sulfoxide, and
then olive oil was added to give a final concentration of 40 mg/ml.
Antibodies were purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA). Fluorescein isothiocyanate-conjugated goat
anti-rat antibody (1:3,000; cat. no. A0258) and immunoglobulin G
were purchased from Beyotime Institute of Biotechnology (Haimen,
China). Primers and other chemical reagents were purchased from
Sangon Biotech Co., Ltd. (Shanghai, China).
Animals and treatment
Male Sprague Dawley rats (n=40; 6-week-old) weighing
~200 g were obtained from the Shanghai Center of Experimental
Animals (Shanghai, China) and were housed in a standard environment
(20±2°C; 50±5% humidity; 12 h light/dark cycle, diet and filtered
water ad libitum). Following acclimatization for 1 week, 10 rats
were weaned to a normal chow diet (7% fat, 64.7% carbohydrate and
18.7% protein), and the other 30 began a high-fat diet (HFD; 43.6%
fat, 24.5% carbohydrate and 14.7% protein). The 30 rats were
randomly allocated into three groups (10 per group): HFD group, the
low dose group (HFD + curcumin 100 mg/kg/day), and the high dose
group (HFD + curcumin 200 mg/kg/day). Rats were given daily
intraperitoneal injections of either curcumin or vehicle (mixture
of dimethyl sulfoxide and olive oil, at a ratio of 1:20) daily for
8 weeks. Food consumption and body weight were measured weekly. The
ethics committee of the First Affiliated Hospital of Xiamen
University (Xiamen, China) granted ethical approval for the care
and use of animals in the present study.
Systemic glucose tolerance and insulin
sensitivity test [oral glucose tolerance test (OGTT) and
intra-peritoneal insulin tolerance test (IPITT)]
In preparation for the performance of glucose
tolerance tests, rats were fasted for 12 h and then administered an
intragastric injection of glucose at a dose of 2 g/kg/body weight
(bw). Blood glucose was then measured at 0, 30, 60, 90, 120 and 180
min time intervals using a One Touch Ultra blood glucose meter
(LifeScan, Inc., San Jose, CA, USA). In order to perform insulin
sensitivity tests, rats were administered an intraperitoneal
injection of insulin (0.75 U/kg/bw), which was then followed by 12
h of fasting. Blood glucose levels were then measured at the same
time intervals as detailed with regards to the OGTT. The areas
under the glucose curves were evaluated according to the
trapezoidal rule (14).
Biochemical analyses
Blood samples were collected from the rats and serum
was immediately centrifuged at 1,600 × g for 10 min at 4°C. Serum
levels of cholesterol (CHO), triglycerides (TG), high-density
lipoprotein (HDL) and low-density lipoprotein-c (LDL) were analyzed
using an automatic chemistry analyzer (Hitachi Ltd., Tokyo,
Japan).
Hepatic TG measurement
In order to quantitatively assess hepatic TG
content, a Triglyceride assay kit (Applygen Technologies, Inc.,
Beijing, China) was used. Briefly, 50 mg liver samples were
homogenized and extracted using a mixture of lipid extract
(chloroform/methanol, 2:1). Samples were then centrifuged and the
organic solvent layer was collected. Hepatic TG concentrations were
measured using the enzymatic hydrolysis method, according to the
manufacturer's protocol.
Hematoxylin and eosin (H&E)
staining
H&E staining was used to assess hepatic
steatosis. Briefly, liver tissues were fixed in 4% paraformaldehyde
at 4°C for 12 h and embedded in paraffin, prepared into 5-µm thick
sections, and stained with hematoxylin for 8 min, wash in running
tap water for 60 min, counterstain with eosin for 60 sec at room
temperature. Images were scored under a light microscope
(magnification, ×200; Nikon Corporation, Tokyo, Japan) according to
the following criteria (15):
Score 1 for 0–25% steatosis, score 2 for 26–50% steatosis, score 3
for 51–75% and score 4 for over 76% steatosis.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total mRNA was isolated from the liver using TRIzol
Reagent (Qiagen China Co., Ltd., Shanghai, China). RT was performed
using the BeyoRT™ II First Strand cDNA Synthesis Kit (Beyotime
Institute of Biotechnology) and then qPCR was performed using the
Light-Cycler 480 System (Roche Diagnostics, Basel, Switzerland)
with SYBR Green reagent (Takara Biotechnology, Co., Ltd., Dalian,
China). The amplification reactions were according to the following
thermal cycling conditions: 95°C for 10 min, 40 cycles of 95°C for
15 sec and 60°C for 30 sec. The quantification was expressed as the
ratio of target genes to GAPDH mRNA using the 2−ΔΔCq
method (16). RT-qPCR was
performed using the primers in Table
I.
 | Table I.Sequences of primers designed for
reverse transcription-quantitative polymerase chain reaction. |
Table I.
Sequences of primers designed for
reverse transcription-quantitative polymerase chain reaction.
| Gene | Forward primer | Reverse primer |
|---|
| CPT-1 |
GCTTTGAAGTTTTTGGTGAAATTGA |
GCTATGGTTTCATCACCTACCGT |
| PPAR-γ |
AAAGAAGCCGACACTAAACC |
CTTCCATTACGGAGAGATCC |
| PPAR-α |
TGTCACACAATGCAATCCGTTT |
TTCAGGTAGGCTTCGTGGATTC |
| SREBP-1 |
GGAGCCATGGATTGCACATT |
CCTGTCTCACCCCCAGCATA |
| FAS |
CACAGCATTCAGTCCTATCCACAGA |
CACAGCCAACCAGATGCTTCA |
| ACC |
CAATCCTCGGCACATGGAGA |
GCTCAGCCAACCGGATGTAGA |
| NF-κB |
TCTGGCGCAGAAGTTAGG |
CCAGAGACCTCATAGTTGT |
| TNF-α | TACAGGCTTGTCACTCGA
ATT | GATGGTGGGTATGGGTCAGAA
GGA |
| COX-2 |
ACTTGCTCACTTTGTTGAGTCATTC |
TTTGATTAGTACTGTAGGGGTTAATG |
| GAPDH |
ACAGCAACAGGGTGGTGGAC |
TTTGAGGGTGCAGCGAACTT |
Protein extraction and western blot
analysis
The protein in the liver samples was extracted and
homogenized using a radioimmunoprecipitation assay lysis buffer
with + phenylmethyl sulfonyl fluoride. Proteins (100 µg) were
separated by 10% SDS-PAGE and transferred to polyvinylidene
fluoride membranes (Roche Diagnostics), blocked using 10% skimmed
milk (Guangming Group, Shanghai, China). Membranes were incubated
overnight at 4°C with primary antibodies against Notch-1 (rabbit
polyclonal; 1:1,000; cat. no. D6F11; Cell Signaling Technology,
Inc., Danvers, MA, USA), HES1 (rabbit polyclonal; 1:800; cat. no.
D6P2U; Cell Signaling Technology, Inc.) and GAPDH (rabbit
polyclonal; 1:1000; cat. no. no. 5174; Cell Signaling Technology,
Inc.). The blots were then incubated at 37°C for 30 min with a
horseradish peroxidase-conjugated goat anti rabbit immunoglobulin G
secondary antibody (1:2,000; cat. no. A0208; Beyotime Institute of
Biotechnology). A UVP Bioimaging system (BioSpectrum 410; UVP Inc.,
Upland, CA, USA) and Gel-Pro-Analyzer software version 5.0 (Media
Cybernetics, Inc., Bethesda, MD, USA) were used for detection and
analysis.
Statistical analysis
Quantitative data were expressed as the mean ±
standard deviation. Statistical significance among the groups was
determined using one-way analysis of variance followed by Tukey's
multiple comparison test. P<0.05 was considered to indicate a
statistically significant difference.
Results
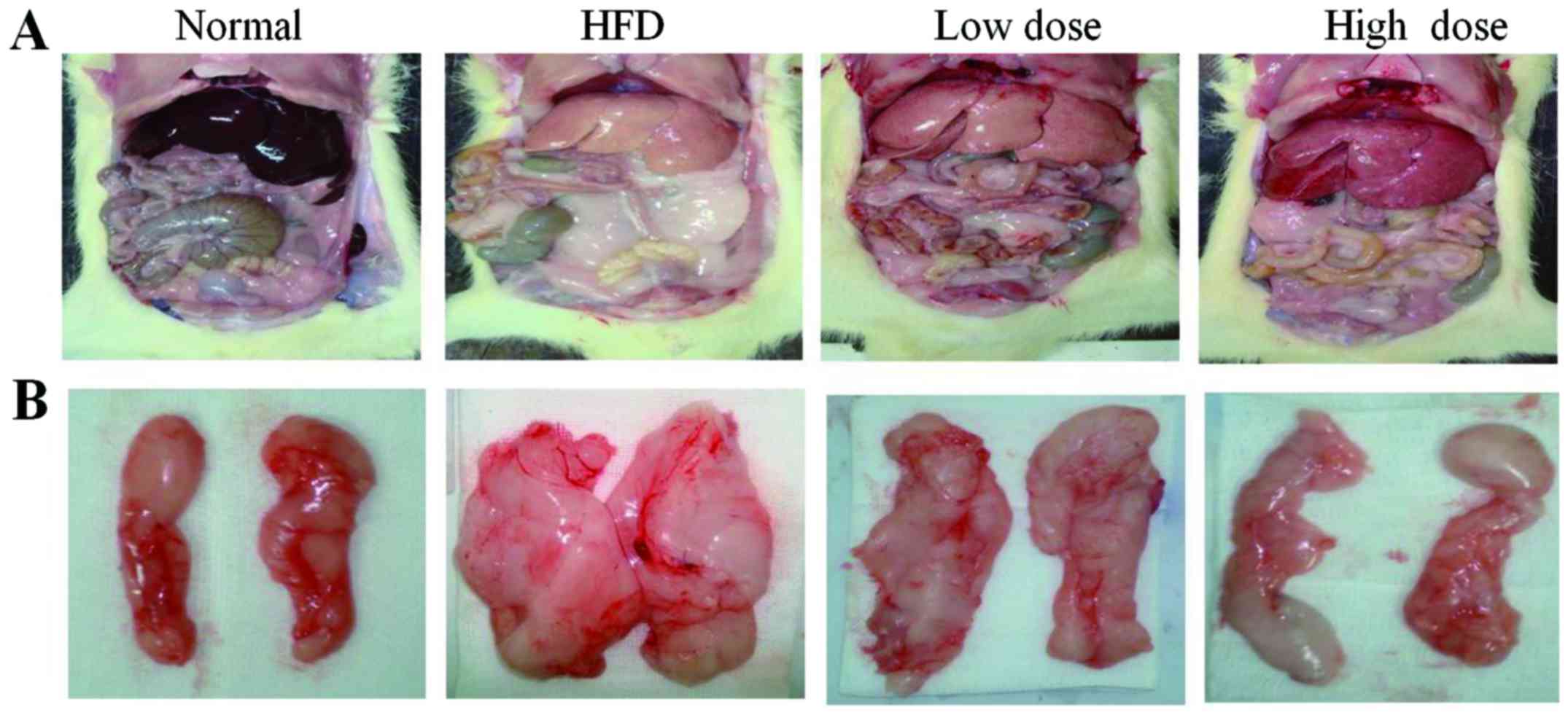
Administration of curcumin decreases
visceral fat and liver weight
With regards to liver morphology, the liver color
was grayer in the HFD group and less gray after curcumin treatment
(Fig. 1A). The visceral fat was
reduced in both the low dose and the high dose curcumin-treated
groups (Fig. 1B), and the liver
weight was also reduced following curcumin treatment in a
dose-dependent manner (Table II;
P<0.05). However, curcumin treatment did not reduce the total
body weight of the rats compared with the HFD group (Table II). In addition, the food
consumption of the rats did not change following treatment with
curcumin (data not shown). These results demonstrate that curcumin
treatment decreases abdominal obesity, but not via reduction of
food intake.
| Table II.Body weight of different groups at the
conclusion of the study. |
Table II.
Body weight of different groups at the
conclusion of the study.
| Parameter | Control | HFD | Low dose
curcumin | High dose
curcumin |
|---|
| Body weight (g) |
612.1±27.35 |
695.1±67.30a |
674.0±56.59 |
665.8±61.29 |
| Peri-epididymal fat
(g) |
11.1±3.13 |
18.5±6.23b |
13.8±5.74c |
11.8±4.15d |
| Peri-renal fat
(g) |
11.2±3.37 |
28.0±3.41a |
22.5±4.39c |
19.7±5.82c |
| Liver weight
(g) |
17.1±0.61 |
30.4±3.57b |
26.4±5.41 |
23.4±5.74c |
Curcumin improves glucose tolerance
and increases insulin sensitivity
OGTT and IPGTT were performed in order to
investigate the effects of curcumin on glucose homeostasis and
insulin sensitivity. As revealed by Fig. 2A, the HFD group experienced a
significant reduction in levels of glucose tolerance compared with
the normal control group. Curcumin treatment significantly reduced
blood glucose levels at 30 and 120 min compared with the HFD group
(P<0.05 and P<0.01). The area under the curve also showed
that glucose concentrations were lower in both the curcumin low
dose (non-significantly) and high dose groups. (compared to the HFD
groups (P<0.01) (Fig. 2B). In
comparison with the control group, HFD led to insulin resistance,
which manifested as a blunted response in the IPGTT (Fig. 2C). Furthermore, administration of
curcumin significantly decreased the blood glucose concentration at
60, 90 and 120 min compared with the HFD group (P<0.05 and
P<0.01; Fig. 2C). The area
under the curve showed that insulin caused a significant reduction
in glucose in curcumin high dose groups (P<0.01 vs. the HFD
group) (Fig. 2D). These results
suggest that curcumin treatment is effective in increasing glucose
tolerance and insulin sensitivity in HFD-induced obesity in
rats.
Curcumin decreases the concentration
of blood lipids
As presented in Table
III, the concentrations of serum CHO, TG and LDL were
significantly decreased in the curcumin-treated groups compared
with HFD rats (P<0.05 and P<0.01). Total HDL did not change
significantly among the groups.
| Table III.Characteristics of serum
chemistry. |
Table III.
Characteristics of serum
chemistry.
| Group | CHO (mmol/l) | TG (mmol/l) | LDL (mmol/l) | HDL (mmol/l) |
|---|
| Normal |
0.53±0.04 |
2.23±0.72 |
0.45±0.18 |
0.47±0.12 |
| HFD |
1.45±0.32b |
4.93±1.17a |
1.77±0.21a |
0.27±0.04b |
| Low dose |
1.24±0.44 |
3.14±1.10c |
1.46±0.29c |
0.25±0.06 |
| High dose |
1.12±0.26c |
2.41±0.72d |
1.28±0.13d |
0.28±0.06 |
Curcumin reduces hepatosteatosis
H&E staining was performed to determine the
effect of curcumin administration on hepatosteatosis. HFD-treated
rats had significantly increased TG levels compared with normal
diet-treated rats. However, treatment with curcumin, both at low
and high doses, reduced both TG levels and liver steatosis, which
was demonstrated by the decreased visualization of lipid droplets
following H&E staining of liver tissue sections (P<0.01;
Fig. 3A and C). Under higher
magnification, large cytoplasmic lipid droplets were observed
throughout the sections (>90%) in the HFD rat liver, but only
~75% fatty infiltration in the low dose and 55% in the high dose
curcumin groups was observed (Fig.
3B).
Curcumin regulates the expression of
genes involved in hepatic lipid metabolism and inflammation
In order to investigate the underlying mechanism of
curcumin in NAFLD, the expression of genes associated with hepatic
lipid metabolism, including those involved in fatty acid oxidation
[peroxisome proliferator-activated receptor-γ (PPAR-γ), peroxisome
proliferator-activated receptor-α (PPAR-α) and carnitine
palmitoyltransferase 1 (CPT-1)], lipogenesis [sterol regulatory
element-binding protein (SREBP-1c), acetyl-CoA carboxylase (ACC)
and fatty acid synthase (FASN)] and inflammation [nuclear factor-κB
(NF-κB), tumor necrosis factor-α (TNF-α) and
prostaglandin-endoperoxide synthase 2 (COX-2)] were investigated
using RT-qPCR. It was revealed that curcumin treatment enhanced
mRNA expression of PPAR-α, CPT-1 and PPAR-γ (Fig. 4A), whilst suppressing mRNA
expression of SREBP-1c, ACC and FASN in the liver compared with HFD
rats (Fig. 4B). Thus, this
suggested that curcumin administration improves hepatic lipid
metabolism in rats via promotion of fat degradation and suppression
of lipogenesis. Furthermore, the expression levels of
inflammation-associated genes NF-κB, TNF-α and COX-2 were reduced
by curcumin treatment compared with the HFD rats (Fig. 4C), therefore demonstrating the
anti-inflammatory effect of curcumin on rats.
 | Figure 4.Effects of curcumin on mRNA expression
in the livers of HFD rats. The results are presented as (A)
lipolysis-associated genes, (B) lipogenesis-associated genes and
(C) inflammation-associated genes. *P<0.01, **P<0.01 compared
with the control group; ∆P<0.05,
∆∆P<0.01 compared with the HFD group. HFD, high fat
diet; PPAR-α, peroxisome proliferator-activated receptor-α; CPT-1,
carnitine palmitoyltransferase 1; PPAR-γ, peroxisome
proliferator-activated receptor-γ; SREBP-1c, sterol regulatory
element-binding protein; FAS, fatty acid synthase; ACC, acetyl-CoA
carboxylase; NF-κB, nuclear factor-κB; TNF-α, tumor necrosis
factor-α; COX-2, prostaglandin-endoperoxide synthase 2. |
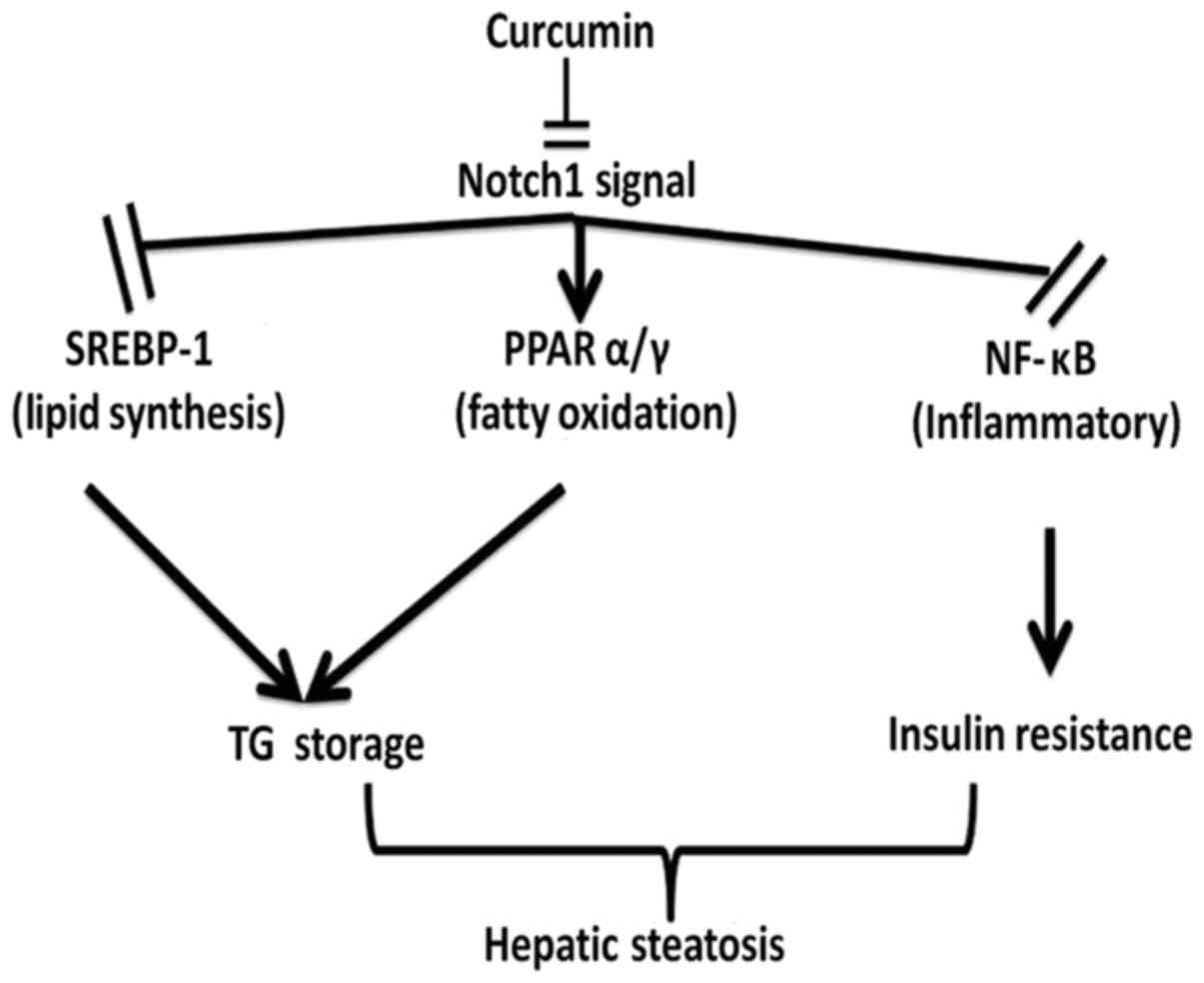
Curcumin reduces hepatic Notch
signaling protein expression
The protein expression of Notch-1 and Hes-1 (which
are the primary Notch-1 target genes that regulate cell fate
decision) were investigated using western blotting. It was revealed
that Notch-1 and Hes-1 were upregulated in the liver of HFD rats
compared with the normal controls, whereas curcumin treatment
reversed this effect in a dose-dependent manner (Fig. 5). Therefore, the results suggest
that curcumin may ameliorate fatty liver and insulin resistance via
downregulation of Notch signaling (Fig. 6).
Discussion
Several important results were demonstrated in the
present study: Curcumin suppresses serum CHO and TG levels, reduces
visceral fat accumulation and abdominal obesity, increases insulin
sensitivity and attenuates HFD-induced hepatic steatosis.
NAFLD is always coupled with insulin resistance and
other metabolic abnormalities (17), and liver fat synthesis and
lipolysis imbalance are associated with these pathological
processes. Various transcription factors are also involved in NAFLD
pathogenesis, including SREBP-1c, FAS, ACC and PPARs. SREBP-1c is a
key pathogenic factor in NAFLD, and is responsible for the
suppression of ACC and FAS expression, which are involved in fat
synthesis and TG aggregation (18). The PPAR family of nuclear receptors
is composed of three family members: PPAR-α, PPAR-β and PPAR-γ. The
activation of PPARs results in the proliferation of peroxisomes and
subsequent upregulation of genes, such as CPT-1, leptin and the
insulin receptor; and thereby attenuates adipogenesis and insulin
resistance (19).
In order to determine the therapeutic mechanism of
curcumin with regards to the amelioration of metabolism disturbance
and NAFLD, the expression levels of lipogenic and lipolytic genes
were investigated. The results of these analyses revealed that
curcumin upregulated the expression of PPAR-α, CPT-1 and PPAR-γ
lipolytic genes (P<0.05 or P<0.01); and downregulated the
expression of SREBP-1c, FAS and ACC lipogenic genes in HFD rats
(P<0.05). The expression levels of inflammation-associated genes
(NF-κB, COX-2 and TNF-α) were also downregulated compared with HFD
rats following administration of curcumin. Therefore, the results
of the present study suggest that curcumin suppresses the synthesis
of fatty acids, enhances lipolysis and decreases inflammation,
which all contribute to amelioration of fatty liver and insulin
resistance.
Notch signaling is an important transduction pathway
for cell fate decisions. Cell surface ligands bind to neighboring
receptors, which result in the formation of active notch
intracellular domains (NICD) (20). NICDs translocate from the membrane
to the nucleus, and then interact with the recombining binding
protein suppressor of hairless DNA-binding protein, in order to
promote the formation of an active complex. This formation leads to
the transcription of Hes-1 and hairy/enhancer-of-split related with
YRPW motif protein 2, which are Notch signaling pathway targets
that mediate transcriptional control of downstream genes (21).
The Notch pathway is always involved in the
pathological process underlying fatty liver, and the major Notch
signaling proteins, Notch-1 and Hes-1, are commonly implicated.
Notch-1 expression has previously been reported to be upregulated
in obese mice (22). Previous
studies have revealed that hepatic Notch signaling is upregulated
and positively correlates with insulin resistance and fatty liver
in humans and other mammals (23).
Furthermore, it has also been demonstrated that Notch
gain-of-function causes fatty liver, and is correlated with an
increasing rate of SREBP-1c-mediated lipogenesis (24). In addition, pharmacological
inhibition of the Notch pathway via a γ-secretase inhibitor has
been revealed to reduce obesity, increase glucose tolerance and
improve insulin sensitivity in diet-induced obesity in mice
(25). Furthermore, silencing of
Notch genes via RNA interference was demonstrated to inhibit the
fatty degeneration of liver cells (24). In addition, it has previously been
revealed that Notch-1 can inhibit adipogenic differentiation via
suppression of PPAR-γ and PPAR-α expression levels (26). The small interfering RNA-mediated
suppression of Hes-1 expression can also inhibit adipogenesis
(27). The results of these
studies suggest that Notch signaling boosts lipogenesis and
inhibits lipolysis via regulation of the expression of SREBP-1 and
PPAR-α/γ transcription factors.
In the present study, an increased level of Notch-1
in fatty liver of rats was identified, coupled with the
upregulation of Hes-1 expression, and downregulation of PPAR-α/γ
expression. By contrast, expression of the SREBP-1, FAS and ACC
lipolysis genes were upregulated in the fatty livers of rats. The
results of the present study suggest that administration of
curcumin inhibits Notch-1 signaling, downregulates SREBP-1
expression, and suppresses the expression of FAS and ACC
lipogenesis genes in the liver. Furthermore, the present study also
demonstrated that curcumin promotes expression of the PPAR-α/γ and
CPT-1 lipolysis genes in rats, which reduces the development of
hyperglycemia and fatty liver.
Fatty liver is occasionally accompanied by
inflammation (28).
Pro-inflammatory cytokines, such as TNF-α, can directly increase
inflammation in liver tissues and impair insulin sensitivity. NF-κB
is heavily involved in the regulation of inflammatory-associated
gene expression in mammals. Activation of NF-κB can enhance the
expression of pro-inflammatory cytokines, which are major factors
in HFD-induced liver damage (29).
High expression of Notch-1 upregulates NF-κB transcriptional
activity; whereas downregulation of Notch-1 suppresses NF-κB
expression (30). The present
study demonstrated that curcumin can attenuate liver injury,
inhibit NF-κB activation and suppress downstream activation of
COX-2 and TNF-α.
With regards to the aforementioned results, the
current study has revealed that curcumin can downregulate Notch
signaling and differentially regulate SREBP-1, PPAR-α/γ and NF-κB
transcription factors; which may contribute to the therapeutic
amelioration of fatty liver and insulin resistance produced by
curcumin. The results of the present study therefore suggest that
the Notch-1 pathway may represent an attractive therapeutic target
for the treatment of insulin resistance and fatty liver induced by
a HFD.
Although the underlying mechanisms of the
curcumin-induced effects in fatty liver and insulin resistance are
complex, the present study, to the best of our knowledge, is the
first to demonstrate that the Notch signaling pathway may be
involved in the pathogenesis of fatty liver and insulin resistance,
and thus represent a potential novel therapeutic target. Further
research is required to validate the therapeutic potential of the
Notch signaling pathway as a target for the treatment of NAFLD and
insulin resistance.
Acknowledgements
The present study was supported by the Science
Foundation of Shanghai (grant no. 15401932900), the Natural Science
Foundation of Xiamen City (grant no. 3502Z20154011), and the
National Natural Science Foundation of China (grant no.
81503579).
References
|
1
|
Chartoumpekis DV, Palliyaguru DL,
Wakabayashi N, Khoo NK, Schoiswohl G, O'Doherty RM and Kensler TW:
Notch intracellular domain overexpression in adipocytes confers
lipodystrophy in mice. Mol Metab. 4:543–550. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carr RM, Oranu A and Khungar V:
Nonalcoholic fatty liver disease: Pathophysiology and Management.
Gastroenterol Clin North Am. 45:639–652. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schwanbeck R: The role of epigenetic
mechanisms in Notch signaling during development. J Cell Physiol.
230:969–981. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu J, Xia Y, Chen K, Zheng Y, Wang J, Lu
W, Yin Q, Wang F, Zhou Y and Guo C: Oncogenic role of the Notch
pathway in primary liver cancer. Oncol Lett. 12:42782016.PubMed/NCBI
|
|
5
|
Gridley T and Kajimura S: Lightening up a
notch: Notch regulation of energy metabolism. Nat Med. 20:811–812.
2014. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim GW, Lin JE, Blomain ES and Waldman SA:
Antiobesity pharmacotherapy: New drugs and emerging targets. Clin
Pharmacol Ther. 95:53–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kuo JJ, Chang HH, Tsai TH and Lee TY:
Positive effect of curcumin on inflammation and mitochondrial
dysfunction in obese mice with liver steatosis. Int J Mol Med.
30:673–679. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sha J, Li J, Wang W, Pan L, Cheng J, Li L,
Zhao H and Lin W: Curcumin induces G0/G1 arrest and apoptosis in
hormone independent prostate cancer DU-145 cells by down regulating
Notch signaling. Biomed Pharmacother. 84:177–184. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hackler L Jr, Ózsvári B, Gyuris M, Sipos
P, Fábián G, Molnár E, Marton A, Faragó N, Mihály J, Nagy LI, et
al: The Curcumin Analog C-150, Influencing NF-κB, UPR and Akt/notch
pathways has potent anticancer activity in vitro and in vivo. PLoS
One. 11:e01498322016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Z, Zhang Y, Banerjee S, Li Y and
Sarkar FH: Notch-1 down-regulation by curcumin is associated with
the inhibition of cell growth and the induction of apoptosis in
pancreatic cancer cells. Cancer. 106:2503–2513. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang Y, Duan W, Liang Z, Yi W, Yan J, Wang
N, Li Y, Chen W, Yu S, Jin Z and Yi D: Curcumin attenuates
endothelial cell oxidative stress injury through Notch signaling
inhibition. Cell Signal. 25:615–629. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Leclercq IA, Farrell GC, Sempoux C, dela
Peña A and Horsmans Y: Curcumin inhibits NF-kappaB activation and
reduces the severity of experimental steatohepatitis in mice. J
Hepatol. 41:926–934. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu ZC, Yang ZX, Zhou JS, Zhang HT, Huang
QK, Dang LL, Liu GX and Tao KS: Curcumin regulates hepatoma cell
proliferation and apoptosis through the Notch signaling pathway.
Int J Clin Exp Med. 7:714–718. 2014.PubMed/NCBI
|
|
14
|
Feng XT, Tang SY, Jiang YX and Zhao W:
Anti-diabetic effects of zhuoduqing formula, a chines herbal
decoction, on a rat model of type 2 diabetes. Afr J Tradit
Complement Altern Med. 14:42–50. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ha SK and Chae C: Inducible nitric oxide
distribution in the fatty liver of a mouse with high fat
diet-induced obesity. Exp Anim. 59:595–604. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ1 and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lallukka S and Yki-Järvinen H:
Non-alcoholic fatty liver disease and risk of type 2 diabetes. Best
Pract Res Clin Endocrinol Metab. 30:385–395. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kohjima M, Higuchi N, Kato M, Kotoh K,
Yoshimoto T, Fujino T, Yada M, Yada R, Harada N, Enjoji M, et al:
SREBP-1c, regulated by the insulin and AMPK signaling pathways,
plays a role in nonalcoholic fatty liver disease. Int J Mol Med.
21:507–511. 2008.PubMed/NCBI
|
|
19
|
Gross B, Pawlak M, Lefebvre P and Staels
B: PPARs in obesity-induced T2DM, dyslipidaemia and NAFLD. Nat Rev
Endocrinol. 13:36–49. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Brzozowa-Zasada M, Piecuch A, Dittfeld A,
Mielańczyk Ł, Michalski M, Wyrobiec G, Harabin-Słowińska M, Kurek J
and Wojnicz R: Notch signalling pathway as an oncogenic factor
involved in cancer development. Contemp Oncol (Pozn). 20:267–272.
2016.PubMed/NCBI
|
|
21
|
Cai Z, Zhao B, Deng Y, Shangguan S, Zhou
F, Zhou W, Li X, Li Y and Chen G: Notch signaling in
cerebrovascular diseases (Review). Mol Med Rep. 14:2883–2898. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Valenti L, Mendoza RM, Rametta R, Maggioni
M, Kitajewski C, Shawber CJ and Pajvani UB: Hepatic notch signaling
correlates with insulin resistance and nonalcoholic fatty liver
disease. Diabetes. 62:4052–4062. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lai PY, Tsai CB and Tseng MJ: Active form
Notch4 promotes the proliferation and differentiation of 3T3-L1
preadipocytes. Biochem Biophys Res Commun. 430:1132–1139. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pajvani UB, Shawber CJ, Samuel VT,
Birkenfeld AL, Shulman GI, Kitajewski J and Accili D: Inhibition of
Notch signaling ameliorates insulin resistance in a FoxO1-dependent
manner. Nat Med. 17:961–967. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pajvani UB, Qiang L, Kangsamaksin T,
Kitajewski J, Ginsberg HN and Accili D: Inhibition of Notch
uncouples Akt activation from hepatic lipid accumulation by
decreasing mTorc1 stability. Nat Med. 19:1054–1060. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ba K, Yang X, Wu L, Wei X, Fu N, Fu Y, Cai
X, Yao Y, Ge Y and Lin Y: Jagged-1-mediated activation of notch
signalling induces adipogenesis of adipose-derived stem cells. Cell
Prolif. 45:538–544. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lei T, Bi Y, Gao MJ, Gao SM, Zhou LL,
Zheng HL and Chen XD: HES1 inhibits adipogenesis of porcine
mesenchymal stem cells via transcriptional repression of FAD24.
Domest Anim Endocrinol. 45:28–32. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Musso G, Cassader M, Cohney S, De Michieli
F, Pinach S, Saba F and Gambino R: Fatty liver and chronic kidney
disease: Novel mechanistic insights and therapeutic opportunities.
Diabetes Care. 39:1830–1845. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yao L, Kan EM, Kaur C, Dheen ST, Hao A, Lu
J and Ling EA: Notch-1 signaling regulates microglia activation via
NF-κB pathway after hypoxic exposure in vivo and in vitro. PLoS
One. 8:e784392013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu Y, Su C, Shan Y, Yang S and Ma G:
Targeting Notch1 inhibits invasion and angiogenesis of human breast
cancer cells via inhibition Nuclear Factor-κB signaling. Am J
Transl Res. 8:2681–2692. 2016.PubMed/NCBI
|