Introduction
Spinal cord injury (SCI) leading to either temporary
or permanent disability always undergoes two pathological phases:
primary injury and secondary injury (1). Primary injury is always instantaneous
and irreversible. Secondary injury involves multiple pathological
processes such as edema, initiation of an ischemic cascade,
inflammation and apoptotic cell death, and can occur over minutes
or weeks following the injury. Currently, the study and treatment
of SCI are mainly focused on reducing secondary injury and
promoting neuronal regeneration (1–4).
Autophagy is a highly conserved process that can be
induced by a variety of conditions such as hypoxia, hunger and
mechanical damage (5). Previous
studies have shown that autophagy plays an important role in SCI
and is closely associated with apoptosis. Autophagy can reduce
neuronal damage and promote locomotor recovery by inhibiting
apoptosis following SCI in rats. Further, disruption of autophagy
may contribute to neuronal cell death (6–8).
However, the role of autophagy in SCI is still controversial and
may vary depending on the type or degree of SCI (7,9,10).
Neuroserpin (NSP) is a member of the serine
proteinase inhibitor (serpin) family and is widely expressed in the
nervous system, in areas such as the cerebral cortex, hippocampus
and the spinal cord (11,12). NSP is a tissue-type plasminogen
activator (t-PA) inhibitor and has been shown to reduce cerebral
infarct volume and protect neurons from ischemia-induced apoptosis
by preventing the harmful effects of t-PA activation. However, NSP
can also protect neurons from death independently of t-PA
inhibition, though the molecular mechanisms underlying this effect
are still poorly understood (13,14).
In the present study, we aimed to build a clip compress model of
SCI and identify the characteristic markers of autophagy following
the injury as well as the effect of NSP treatment on autophagy. We
hope to provide a novel therapeutic strategy for the treatment of
SCI based on intervention of autophagy.
Materials and methods
Animals
Adult male Sprague-Dawley (SD) rats weighing 250–300
g were provided by the Department of Laboratory Animal Science of
Fudan University. All animal interventions strictly complied with
the regulations of the Animals Ethics Committee of Fudan
University. Animals were kept in a specific pathogen-free (SPF)
barrier system for 1 week to adapt to the environment and were fed
ad libitum. The room temperature was held constant at
27°C.
SCI model
Before the operation, animals were anesthetized with
10% chloral hydrate via intraperitoneal injection (0.3 ml/100 g).
After sterilization with iodophor, they were placed in a prone
position. T10 was located according to the bony process on the
dorsum, and the skin and muscle were cut and separated to expose
the lamina. A laminectomy was performed with a mini-toothed clamp
to fully expose the spinal cord at T9-T10, then a smooth vascular
clamp (30 g force; Oscar, Shanghai, China) was used to compress the
spinal cord vertically for 1 min (15–17).
Postoperative care included manual assisted urination twice daily.
If hematuresis was noted, penicillin was administered to prevent
infection. The rat hairs were kept dry, and food and water were
placed so that the animals had easy and free access.
NSP treatment
Immediately following the injury, rats randomly
received NSP (20 µg/ml, intrathecal injection; PeproTech) 25 µl or
an equal volume of saline vehicle. A group of sham-operated rats
was included as an additional control group.
Western blot analysis
At different time-points (2, 4, 24, 72 h and 7 days)
after SCI and 3 d after the sham operation (n=4), rats were
anesthetized with 10% chloral hydrate, and an intracardiac
perfusion was performed with normal saline. A spinal cord segment
(0.5 cm) across the epicenter of injury was collected and rapidly
stored at −80°C for further study. Protein was extracted with RIPA
[150 mM NaCl, 50 mM Tris (8.0), 1% NP40, 1% sodium deoxycholate,
0.1% SDS pH 7.4] that contained a protease inhibitor cocktail
(Roche, Germany). The protein concentration was determined with a
BCA protein assay kit (Tiangen, China). A total of 40 µg of protein
was separated via 12.5% polyacrylamide gel (Promoton, Shanghai,
China) and transferred onto a PVDF membrane. The membrane was
blocked with 5% skim milk in TBST (TBS contain 0.1% Tween-20) for 1
h, then incubated with primary antibody rabbit anti-LC3 (1:3,000;
Sigma-Aldrich, St. Louis, MO, USA), mouse anti-p62 (1:2,000; Abcam,
Cambridge, UK), rabbit anti-beclin1 (1:1,000; Novus Biologicals,
Littleton, CO, USA) at 4°C overnight. Thereafter, the membrane was
incubated with secondary antibody linked to horseradish peroxidase
(HRP) (goat anti-mouse or goat anti-rabbit, 1:5,000; Sigma-Aldrich)
for 2 h at room temperature. β-actin (mouse anti-β-actin, 1:5,000;
Sigma-Aldrich) was set as an internal reference. Immunoreactive
bands were detected with an enhanced chemiluminescence system (ECL
kit; Sangon Biotech Co., Ltd., Shanghai, China). The image was
analyzed with Quantity One software (version 4.62; Bio-Rad,
Hercules, CA, USA). A commercial molecular weight marker ranging
from 10 to 250 kDa was used to locate the target protein.
Transmission electron microscopy
To evaluate the ultrastructural changes following
SCI, transmission electron microscopy (TEM) was further used for
autophagy detection at 4 h and 3 days and compared with the sham
group (n=3 for each group). After anesthesia, the rats were
perfused with normal saline (37°C) to clean the blood. Then, 50 ml
of fixed liquid (0.1 M phosphate buffer (pH 7.4) containing 4%
paraformaldehyde and 1% glutaraldehyde, 37°C) was perfused quickly,
after which 250 to 300 ml of fixed liquid (4°C) was perfused slowly
for 10 to 15 mins. The targeted spinal cord (2.5 mm around the
epicenter) was then quickly removed, and the anterior horn of the
spinal cord was cut into 1 mm3 pieces and postfixed in
2.5% glutaraldehyde for more than 3 h at 4°C. Subsequently,
according to the standard procedures, the pieces were osmicated
with 1% osmium tetroxide for 1 h, dehydrated in an ethanol series
and embedded in epon. Ultrathin sections were cut and stained with
uranyl acetate and lead citrate in that order. An electron
microscope (Hitachi, Tokyo, Japan) was used to evaluate the
ultrastructural changes.
Immunofluorescence double
staining
At 3 days following SCI, tissue sections from the
sham-group, the vehicle-group and the NSP-group were collected.
After anesthesia, the rats (SCI and Sham group, n=3) were placed in
the supine position. An intracardiac perfusion was performed with
0.01 M phosphate-buffered saline (PBS, pH 7.4) and followed with 4%
paraformaldehyde (PFA in 0.1 M PBS, pH 7.4). A spinal cord segment
(2 cm length) across the epicenter of the injury was harvested and
fixed with 4% PFA overnight. The tissue was subsequently soaked in
30% sucrose solution at 4°C until it sank to the bottom. Then, the
spinal cord was embedded in OCT (Polysciences, Inc., Warrington,
PA, USA) at −20°C and cut into a series of 10 µm transverse
sections. Ten sequential sections at 50 µm intervals were
collected, which spanned a 500 µm length across the epicenter site.
Frozen sections were placed on poly-L-lysine coated slides and
permeabilized in 0.1 mol/l PBS (pH 7.40) containing 0.25% Triton
X-100 for 30 min, blocked with 3% BSA in 0.1 mol/l PBS containing
0.05% Tween for 30 min at room temperature, and then incubated with
rabbit anti-LC3 antibody (1:200; Sigma-Aldrich) and mouse anti-MAP2
(1:200; ProteinTech, Chicago, IL, USA) overnight at 4°C. After
washing in PBS, the sections were incubated in a mixture of goat
anti-rabbit TRITC and goat anti-mouse FITC fluorescence-conjugated
secondary antibodies (1:500; Invitrogen, Carlsbad, CA, USA) for 2 h
at room temperature. The anterior horn in these slides was observed
under a laser confocal microscope (Nikon, Tokyo, Japan).
Nissl staining
At 14 days, tissue from the sham-, the vehicle- and
the NSP-groups were cut into a series of 50 µm transverse sections.
Ten sequential sections at 50 µm intervals were collected, which
spanned a 1,000 µm length across the epicenter site. Sections were
stained with Nissl staining solution (Beyotime, Nanjing, China)
according to the manufacturer's instructions. In brief, the spinal
cord was stained with Nissl solution for 10 min, followed by two 5
min washes with 95% alcohol. The sections were subsequently made
diaphanous twice with dimethylbenzene for 5 min. The overall
structure from each group was compared. The number of anterior horn
motor neurons were also counted and compared.
Behavioral analysis
Basso-beattle-bresnahan (BBB) locomotor rating scale
was applied to evaluate motor function recovery. The scale ranged
from 0 to 21. A score of 0 indicates complete paralysis of the hind
limb, while a score of 21 indicates normal locomotion. The rats
were set in an open-field, and two experienced researchers who were
blind to the operation participated in the evaluation (24, 72, 168
h and 14 days, for 5 min during each period of observation). The
NSP-group and vehicle-group scores were compared.
Statistical analysis
Data is presented as the mean ± SD and compared
using one-way analysis of variance (ANOVA). All experimental data
were analyzed using SPSS 20.0 (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered statistically significant.
Results
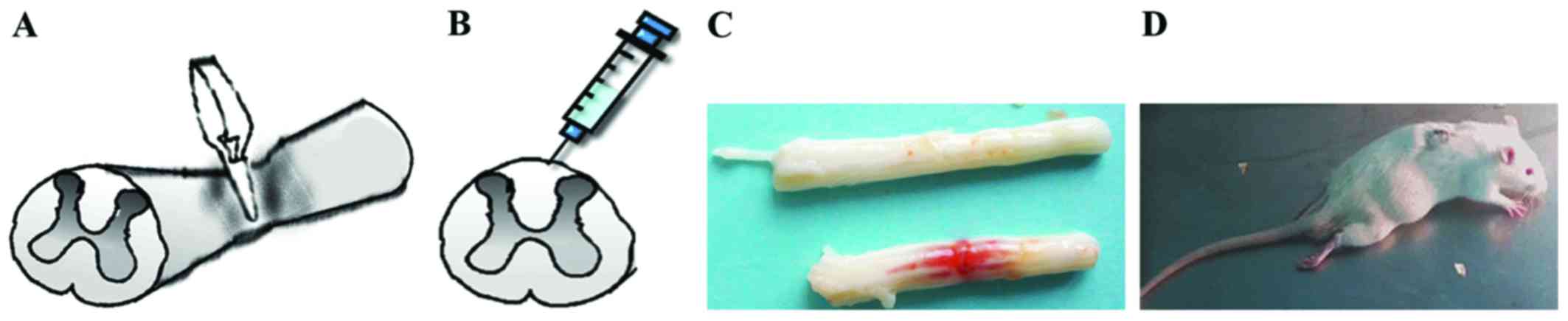
SCI model
The animals were anesthetized with 10% chloral
hydrate via intraperitoneal injection (0.3 ml/100 g). A smooth
vascular clamp (30 g force; Oscar) was used to compress the spinal
cord vertically for 1 min (Fig.
1A). The sham group underwent the same surgical procedure, but
no clip compression was applied to the spinal cord. After adequate
hemostasis, the incision was closed in layers. A strip hemorrhagic
focus was visible after the clamp was removed (Fig. 1C). When the animals awakened, they
were evaluated for double lower limb paralysis and the presence of
any obstacles to urination (Fig.
1D).
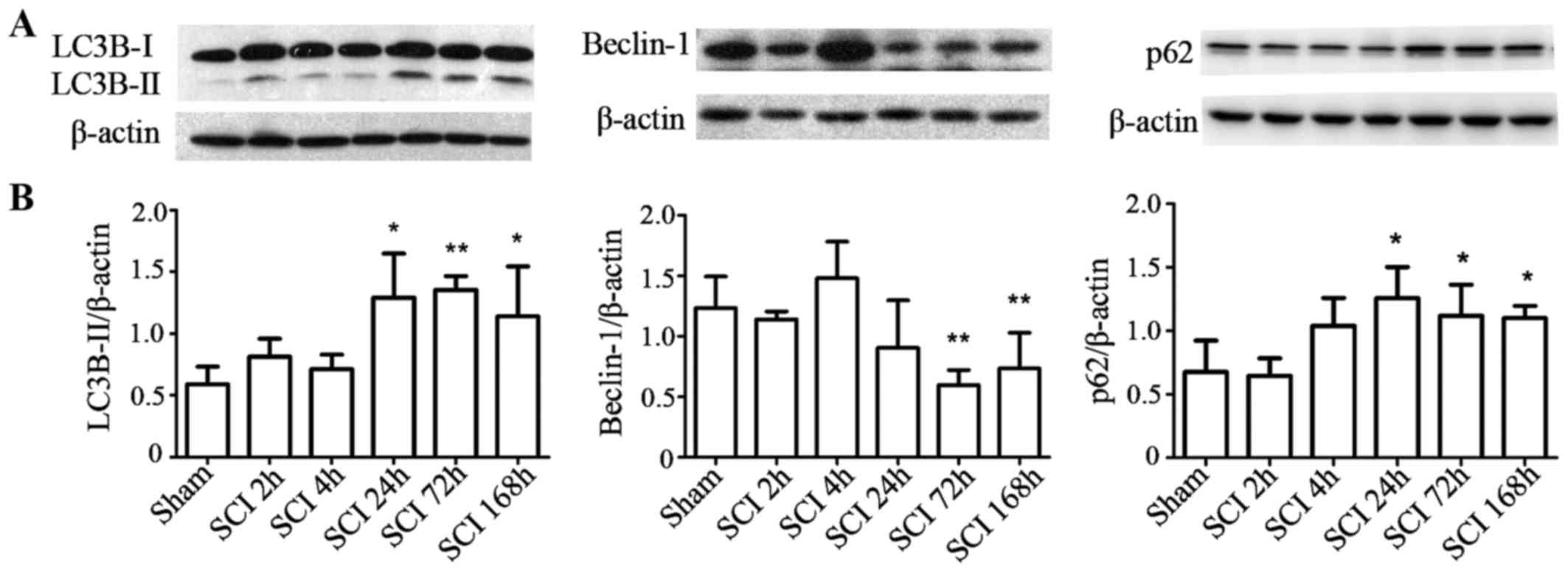
Expression of LC3-II and p62 in SCI
rats
To investigate the changes in autophagy at different
time-points following SCI, LC3-II and p62 protein levels were
measured by western blot analysis. Compared with the sham-operated
group, LC3-II and p62 protein levels increased at 1, 3, and 7 days
after injury in the experimental group (P<0.05), while Beclin-1
significantly dropped at 3 and 7 days post-surgery in the SCI
compared with the sham group (P<0.01; Fig. 2A and B).
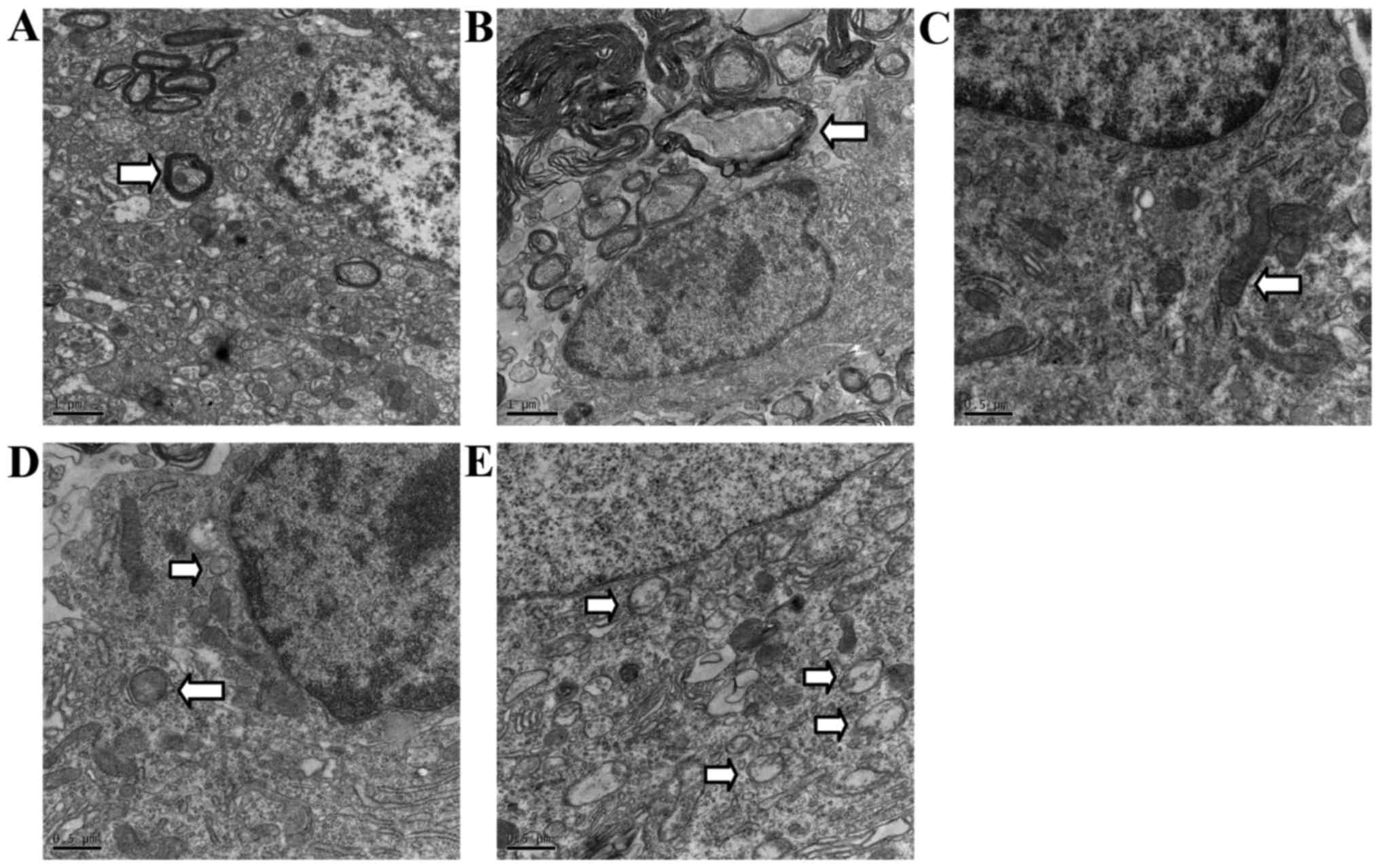
Ultrastructural examination for
autophagy in SCI rats
Normal mitochondria, endoplasmic reticulum and
regular myelinated axons were observed in TEM from the sham-group
rats in the areas corresponding to those in the SCI group, but
rarely in autophagic vacuoles (Fig. 3A
and C). In contrast, disordered structure of mitochondria,
myelinated axons, and numerous autophagic vacuoles with a
double-membrane structure and parts of cytoplasmic organelles were
detected near the epicenter of the rats at 2 h, and especially 72 h
after SCI. Autophagosomes were largely observed in the cytoplasm
near the nuclei (Fig. 3B, D and
E).
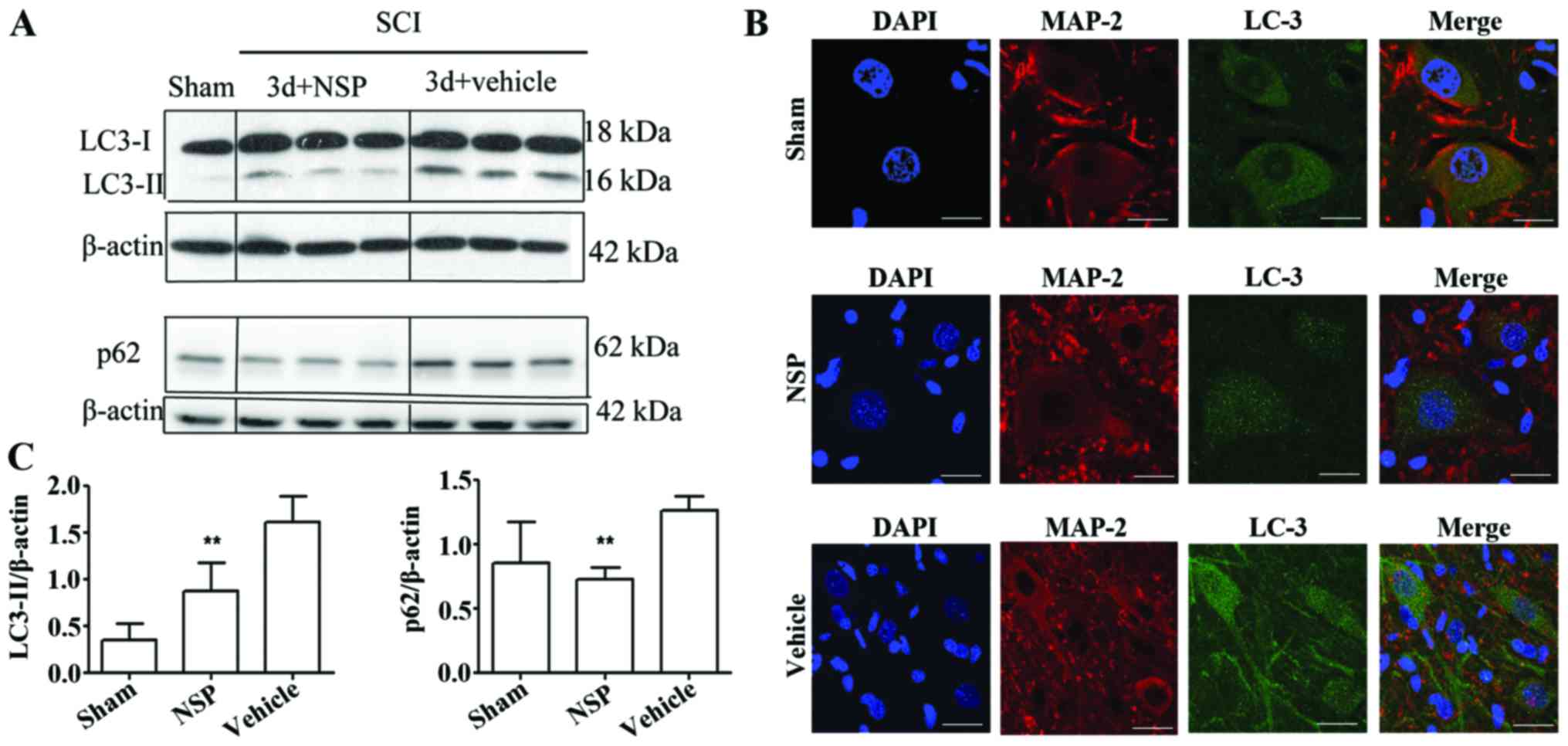
NSP reduced the levels of LC3-II and
p62
Western blot analysis indicated that NSP treatment
decreased LC3-II and p62 protein levels at 72 h after SCI when
compared with the vehicle-treated rats (P<0.01; Fig. 4A and B). Double immunofluorescence
staining of the epicenter sections also showed that at 72 h after
the injury, LC3 was increased in the SCI compared with the
sham-group, and displayed a punctate distribution in the neurons.
NSP decreased the LC3 expression as evidenced by the decreased
numbers of LC3 dots under confocal microscopy (Fig. 4C).
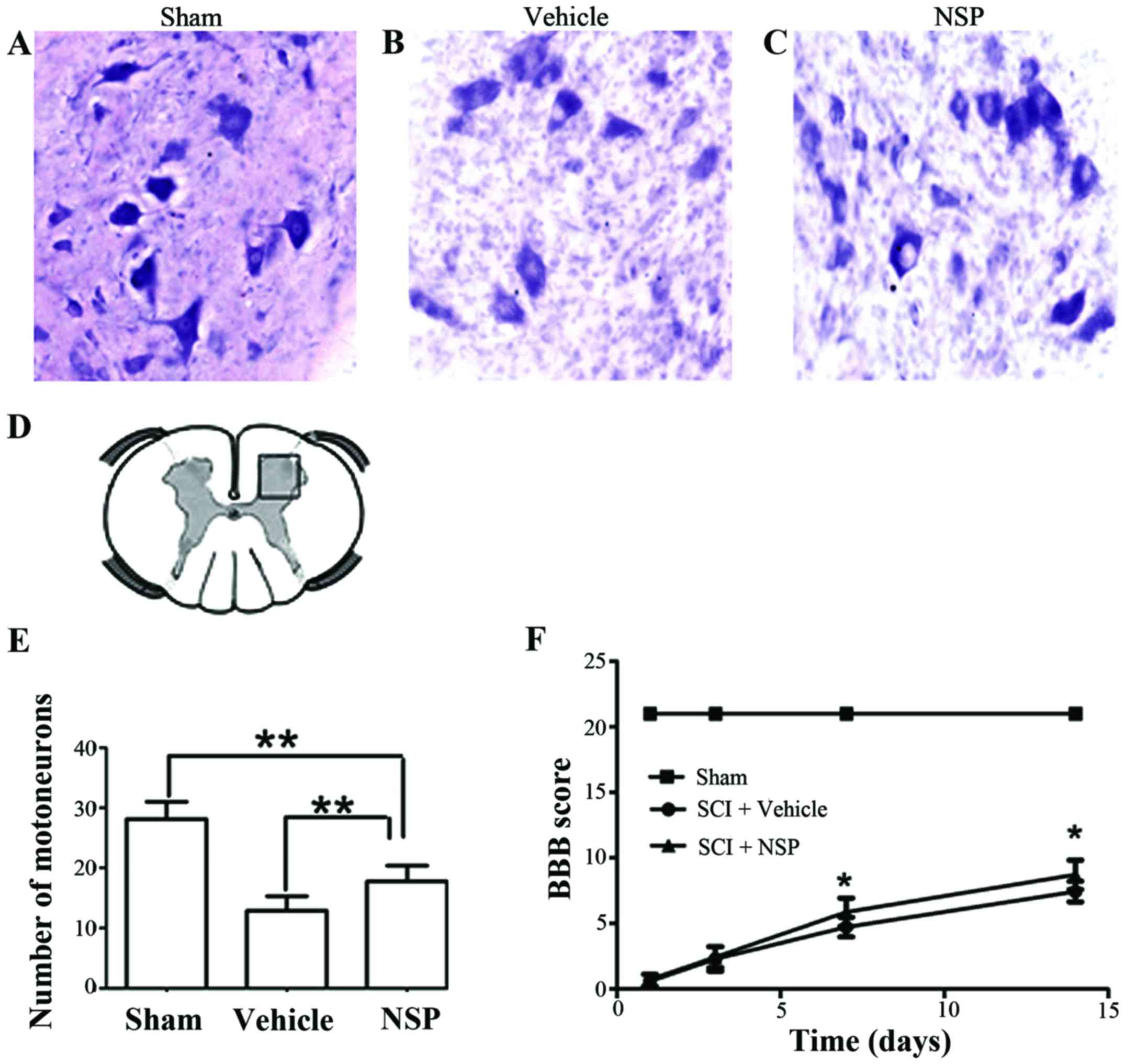
NSP promotes neuroprotection and
functional recovery following SCI
At 14 days after injury, there was an increase in
the number of motor neurons near the injury epicenter in NSP
treated rats compared with the vehicle group. Sections located a 2
mm distance from the epicenter revealed that NSP increased the
number of motor neurons However, there was also a significant
decrease in the number motor neurons in the NSP-treated rats
compared with sham-group (Fig.
5A-E). Representative photomicrographs of motor neurons from
NSP-treated and vehicle-treated rats are shown in Fig. 5A-C. The BBB score was measured at
1, 3, 7 and 14 days (Fig. 5F).
From 7 to 14 days after injury, the scores in the NSP-treated rats
were significantly higher than those in the vehicle-treated group.
However, the scores of the NSP-treated and vehicle-treated groups
both declined compared with those of the sham-group.
Discussion
Previous studies have identified that autophagy
could reduce neuronal damage and promote locomotor recovery via
inhibition of apoptosis after SCI in rats. Here, we showed that NSP
can restore autophagic flux impairment and reduce autophagosomes
accumulation by decreasing LC3-II and p62 protein levels at 72 h
after SCI. As expected, NSP significantly reduced neuronal damage
and promoted locomotor function after SCI. To the best of our
knowledge, this is the first study to demonstrate a link between
NSP-mediated neuroprotection and autophagy following SCI.
Autophagy has been reported to be involved in a
series of neurological disease, such as Alzheimer's disease,
Parkinson's disease, traumatic brain injury and spinal cord injury
(18–20). The process of autophagy entails the
degradation of expired proteins or damaged organelles to maintain
the stability of the intracellular environment, which is important
for cells to survive a temporary crisis. However, autophagy can
also induce cell death, which is often referred to as type II
programmed cell death (21).
Autophagy is also closely associated with apoptosis. The role that
autophagy plays in SCI is still controversial and varies depending
on the type and extent of SCI as well as the SCI animal model being
studied. A study by Chen et al showed that autophagy was
activated after SCI (in a contusion rat model, 10 g rod dropped
from 25 mm height) and inhibited by methylprednisolone suggesting
that methylprednisolone may protect neurons by inhibiting
autophagic cell death (22).
However, Sekiguchi et al reported that rapamycin also
promotes autophagy and reduces neural tissue damage after SCI (in a
contusion mice model, 10 g × 3 mm) (10). Another study by Tang et al
demonstrated that autophagy reduces neural damage and promotes
locomotor recovery via inhibition of apoptosis after SCI (a
hemisection model) (7). In our
study, using another SCI rat model with a simple and economic clip
compression, we demonstrated changes in the characteristic markers
of autophagy after SCI and, further, illustrated these changes with
TME.
LC3, a mammalian homolog of yeast Atg8, is the most
important and reliable marker of autophagy used in research
studies. During the formation of the autophagosome membrane, the
conversion of LC3-I to LC3-II is considered to be one of the
symbols of autophagic induction. Therefore, LC3-II is often used as
a marker for autophagosomes. Beclin-1 did not strictly follow the
same pattern as LC3-II and peaked at 4 h. Previous studies have
demonstrated that beclin-1 participates in the regulation of
autophagy as well as playing a role in apoptosis that is not
limited to autophagy (23). Our
study demonstrated that LC3-II protein levels trended upwards from
24 h after SCI and peaked at 72 h. p62 protein interacts with
LC3-II and transports altered proteins to autophagosomes for
degradation. The accumulation of p62 protein reflects the
disruption of autophagic flux (24). Our study showed that p62 rises from
24 h after SCI. The downregulation of autophagy indicated that
cells may be trapped in an energy crisis or have sustained injuries
to organelles and can no longer control autophagic degradation.
These cells may then proceed to undergo autophagic death or
apoptosis. TEM, which can display the double-membrane
autophagosome, is the gold standard for autophagy detection
(25). In this study, TEM showed
that autophagosomes largely accumulated at 72 h following
compression-induced SCI. In the sham group, few autophagosomes were
observed, while normal cell structure with layered myelin was
ubiquitous. This was in contrast to the numerous vacuole-like
structures and disordered myelin observed in the SCI group. The
loss of neural structure after SCI may directly contribute to the
functional impairment, which is difficult to reverse with simple
drug intervention.
To minimize the secondary cell death after SCI, it
is imperative that drug therapy be promptly instituted. NSP is a
tissue-type plasminogen activator (t-PA) inhibitor. A previous
study showed that in a focal cerebral ischemia/reperfusion model,
intracortical injection of 30 µmol/l NSP reduced infarct volume by
64% from 161 mm3 in control animals to 58 mm3
in NSP-treated animals 72 h after reperfusion. NSP also decreased
apoptotic cell counts in the ischemic penumbra by more than 50%
(14). Our previous study also
demonstrated that, in the early period of sustained spinal cord
compression. NSP is upregulated and plays a neuroprotective role
against neuronal apoptosis (12).
NSP does not appear to freely cross the blood-brain barrier. In the
present study, NSP was administered by intrathecal injection based
on the results from previous studies in which. intracisternal or
intracortical injections of NSP were used (14,26).
In addition, the doses of NSP used in this study were based on our
previous protocols (27). Our
results demonstrate that autophagic flux was restored by NSP,
possibly by lowering p62 and LC3-II protein levels.
Immunofluorescence double staining also indicated that the LC3
fluorescence dots, located in the cytoplasm were markedly decreased
in neurons at the epicenter in the NSP group. These data indicate
that NSP partially inhibited the accumulation of autophagosomes.
Additionally, NSP improved functional recovery after injury and
significantly increased the number of anterior horn motor neurons.
Therefore, we suggest that NSP-induced neuroprotection and
functional recovery following SCI may at least in part be
associated with a restoration of SCI-induced autophagic flux
impairment.
In conclusion, our results suggest that treatment
with NSP following SCI restores the impaired autophagic flux and
offers a new strategy for reducing secondary damage.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (81301047).
Glossary
Abbreviations
Abbreviations:
|
NSP
|
neuroserpin
|
|
SCI
|
spinal cord injury
|
|
TEM
|
transmission electron microscopy
|
|
SD
|
Sprague-Dawley
|
References
|
1
|
Kwon BK, Tetzlaff W, Grauer JN, Beiner J
and Vaccaro AR: Pathophysiology and pharmacologic treatment of
acute spinal cord injury. Spine J. 4:451–464. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yu WY and He DW: Current trends in spinal
cord injury repair. Eur Rev Med Pharmacol Sci. 19:3340–3344.
2015.PubMed/NCBI
|
|
3
|
Thuret S, Moon LD and Gage FH: Therapeutic
interventions after spinal cord injury. Nat Rev Neurosci.
7:628–643. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Varma AK, Das A, Wallace G IV, Barry J,
Vertegel AA, Ray SK and Banik NL: Spinal cord injury: A review of
current therapy, future treatments, and basic science frontiers.
Neurochem Res. 38:895–905. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mizushima N: Autophagy: Process and
function. Genes Dev. 21:2861–2873. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang ZY, Lin JH, Muharram A and Liu WG:
Beclin-1-mediated autophagy protects spinal cord neurons against
mechanical injury-induced apoptosis. Apoptosis. 19:933–945. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tang P, Hou H and Zhang L, Lan X, Mao Z,
Liu D, He C, Du H and Zhang L: Autophagy reduces neuronal damage
and promotes locomotor recovery via inhibition of apoptosis after
spinal cord injury in rats. Mol Neurobiol. 49:276–287. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu S, Sarkar C, Dinizo M, Faden AI, Koh
EY, Lipinski MM and Wu J: Disrupted autophagy after spinal cord
injury is associated with ER stress and neuronal cell death. Cell
Death Dis. 6:e15822015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kanno H, Ozawa H, Sekiguchi A and Itoi E:
Spinal cord injury induces upregulation of Beclin 1 and promotes
autophagic cell death. Neurobiol Dis. 33:143–148. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sekiguchi A, Kanno H, Ozawa H, Yamaya S
and Itoi E: Rapamycin promotes autophagy and reduces neural tissue
damage and locomotor impairment after spinal cord injury in mice. J
Neurotrauma. 29:946–956. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Miranda E and Lomas DA: Neuroserpin: A
serpin to think about. Cell Mol Life Sci. 63:709–722. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wan S, Feng Z, Chen Z, Wang X, Cao Y, Shao
Y and Jiang X: Neuroserpin upregulates in the early period of
sustained spinal cord compression. Clin Lab. 58:891–896.
2012.PubMed/NCBI
|
|
13
|
Wu J, Echeverry R, Guzman J and Yepes M:
Neuroserpin protects neurons from ischemia-induced plasmin-mediated
cell death independently of tissue-type plasminogen activator
inhibition. Am J Pathol. 177:2576–2584. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yepes M, Sandkvist M, Wong MK, Coleman TA,
Smith E, Cohan SL and Lawrence DA: Neuroserpin reduces cerebral
infarct volume and protects neurons from ischemia-induced
apoptosis. Blood. 96:569–576. 2000.PubMed/NCBI
|
|
15
|
Rivlin AS and Tator CH: Effect of duration
of acute spinal cord compression in a new acute cord injury model
in the rat. Surg Neurol. 10:38–43. 1978.PubMed/NCBI
|
|
16
|
von Euler M, Seiger A and Sundström E:
Clip compression injury in the spinal cord: A correlative study of
neurological and morphological alterations. Exp Neurol.
145:502–510. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang HY, Wang ZG, Wu FZ, Kong XX, Yang J,
Lin BB, Zhu SP, Lin L, Gan CS, Fu XB, et al: Regulation of
autophagy and ubiquitinated protein accumulation by bFGF promotes
functional recovery and neural protection in a rat model of spinal
cord injury. Mol Neurobiol. 48:452–464. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nixon RA: The role of autophagy in
neurodegenerative disease. Nat Med. 19:983–997. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Luo CL, Li BX, Li QQ, Chen XP, Sun YX, Bao
HJ, Dai DK, Shen YW, Xu HF, Ni H, et al: Autophagy is involved in
traumatic brain injury-induced cell death and contributes to
functional outcome deficits in mice. Neuroscience. 184:54–63. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ghavami S, Shojaei S, Yeganeh B, Ande SR,
Jangamreddy JR, Mehrpour M, Christoffersson J, Chaabane W, Moghadam
AR, Kashani HH, et al: Autophagy and apoptosis dysfunction in
neurodegenerative disorders. Prog Neurobiol. 112:24–49. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Smith CM, Chen Y, Sullivan ML, Kochanek PM
and Clark RS: Autophagy in acute brain injury: Feast, famine, or
folly? Neurobiol Dis. 43:52–59. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen HC, Fong TH, Lee AW and Chiu WT:
Autophagy is activated in injured neurons and inhibited by
methylprednisolone after experimental spinal cord injury. Spine
(Phila Pa 1976). 37:470–475. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kang R, Zeh HJ, Lotze MT and Tang D: The
Beclin 1 network regulates autophagy and apoptosis. Cell Death
Differ. 18:571–580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tanabe F, Yone K, Kawabata N, Sakakima H,
Matsuda F, Ishidou Y, Maeda S, Abematsu M, Komiya S and Setoguchi
T: Accumulation of p62 in degenerated spinal cord under chronic
mechanical compression: Functional analysis of p62 and autophagy in
hypoxic neuronal cells. Autophagy. 7:1462–1471. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gurusamy N and Das DK: Detection of cell
death by autophagy. Methods Mol Biol. 559:95–103. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang Z, Zhang L, Yepes M, Jiang Q, Li Q,
Arniego P, Coleman TA, Lawrence DA and Chopp M: Adjuvant treatment
with neuroserpin increases the therapeutic window for tissue-type
plasminogen activator administration in a rat model of embolic
stroke. Circulation. 106:740–745. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lv YC, Shi DY, Jiang XX, Feng ZZ, Jiang C
and Chen ZX: Repair effects of neuroserpin (NSP) on neural function
after acute spinal cord injury (SCI) in rats. Fudan Univ J Med Sci.
41:216–221. 2014.(In Chinese).
|