Introduction
Glioma is the most common and aggressive of
malignant the brain tumours, and accounts for ~30% of all central
nervous system tumours and 80% of all malignant brain tumours
(1). Based on their pathology,
gliomas can be divided into astrocytomas, glioblastomas,
medulloblastomas, ependymomas and oligodendrogliomas (2). To date, the molecular mechanisms
underlying the occurrence and development of gliomas remain
unclear. Gliomas can be associated with several risk factors,
including tumour origin, genetic factors, biochemical environment,
ionizing radiation, nitroso compounds, air pollution, unhealthy
lifestyle habits and infection (3,4). At
present, surgery combined with radiotherapy, chemotherapy and
targeted therapy is the standard treatment for patients with glioma
(5). Despite an evident progress
in the therapeutic strategies for gliomas, the prognosis for
patients afflicted with this disease remains poor (6). The median survival time of patients
with glioma is ~9–12 months (7).
The poor prognosis of patients with glioma is partially due to the
highly invasive nature of glioma cells, making complete surgical
resection unattainable (8,9). Therefore, elucidation of the
mechanisms underlying the formation and progression of glioma and
development of effective diagnostic and therapeutic treatment
methods for patients with glioma are required.
MicroRNAs (miRNAs/miRs) are a class of endogenous,
short noncoding ~19–25 nucleotides-long RNA molecules (10). miRNAs negatively modulate gene
expression at the transcriptional level by base pairing with the
3′-untranslated region (3′-UTRs) of their target mRNAs, and
inducing mRNA degradation or inhibition of translation (11). Through negative regulation of the
target genes, miRNAs participate in many physiological processes,
including cell proliferation, cell cycle progression,
differentiation, apoptosis and metastasis (12–15).
Recent studies have reported that miRNAs are abnormally expressed
in human cancers, including glioma, breast, gastric, bladder and
lung cancers (16–20). In particular, abnormal expression
of certain miRNAs is associated with tumorigenesis and cancer
progression (21,22). Dysregulated miRNAs may serve
similar roles to oncogenes or tumour suppressor genes, depending on
their target mRNAs (23).
Therefore, miRNAs may demonstrate a potential as novel prognostic
indicators and therapeutic targets for patients with malignancies
(24).
miR-103 is abnormally expressed in various human
cancers (25–27). The present study aimed to elucidate
the expression pattern, biological functions and underlying
molecular mechanism of miR-103 in glioma.
Materials and methods
Tissue samples and cell lines
The present study was approved by the Ethical
Committee of Linyi People's Hospital (Linyi, China). Written
informed consent was obtained from all participants. A total of 26
surgically resected glioma tissues were collected from patients
with glioma (male, 25; female, 11; age range, 37–72 years) in Linyi
People's Hospital between July 2013 and February 2016. A total of
12 normal brain tissue samples were obtained from patients with
cerebral trauma (male, 7; female, 5; age range, 25–68 years)
subjected to a surgical procedure. None of the patients were
treated with radiotherapy or chemotherapy prior to the surgical
resection. All collected tissues were immediately snap-frozen in
liquid nitrogen at the time of surgery and stored at −80°C.
Normal human astrocytes (NHAs) were purchased form
ScienCell Research Laboratories, Inc. (San Diego, CA, USA) and
cultured in the astrocyte medium (ScienCell Research Laboratories,
Inc.). Glioma cell lines U251, LN18 and LN229 were obtained from
the Cell Bank Type Culture Collection of the Chinese Academy of
Sciences (Shanghai, China). All glioma cell lines were maintained
in Dulbecco's modified Eagle medium (DMEM) containing 10% fetal
bovine serum (FBS), 100 U/ml penicillin and 100 mg/ml streptomycin
(all Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
at 37°C in a humidified atmosphere containing 5%
CO2.
Cell transfection
miR-103 mimics and negative control miRNA (miR-NC)
were purchased from Guangzhou RiboBio Co., Ltd. (Guangzhou, China).
Brain-derived neurotrophic factor (BDNF) overexpression plasmid
(pcDNA3.1-BDNF) and blank plasmid (pcDNA3.1) were obtained from
Shanghai GenePharma Co., Ltd. (Shanghai, China). For transfection,
cells were seeded in 6-well plates at a density of 1×105
cells/per well and cultured until 60–70% confluence was attained.
Cells were separately treated with pcDNA3.1-BDNF or pcDNA3.1 using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.,),
according to the manufacturer's protocol. Following incubation at
37°C with 5% CO2 for 8 h, the transfection mixture was
replaced with fresh DMEM supplemented with 10% FBS, without
penicillin and streptomycin. Following transfection for 24 h, MTT
assay was performed. Reverse transcription-quantitative polymerase
chain reaction (RT-qPCR) and Transwell invasion assays were
performed at 48 h post-transfection. Western blotting analysis was
performed to detect protein expression at 72 h
post-transfection.
RT-qPCR
Total RNA was extracted from tissues or cells using
the TRIzol reagent (Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. To quantify the expression of miR-103,
total RNA was reverse transcribed into cDNA by using the TaqMan
MicroRNA Reverse Transcription kit (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The temperature protocol for reverse
transcription was as follows: 16°C for 30 min, 42°C for 30 min and
85°C for 5 min. Subsequently, the TaqMan MicroRNA PCR kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.) was used to detect
miR-103 expression in an RT-qPCR amplification reaction, with U6 as
an internal reference gene. To quantify the BDNF mRNA expression,
cDNA was synthesized using the M-MLV Reverse Transcriptase (Promega
Corporation, Madison, WI, USA), followed by a qPCR reaction with
the SYBR Green Real-Time PCR Master Mix (Toyobo Life Science,
Osaka, Japan) using a 7500 Sequence Detection system (Applied
Biosystems; Thermo Fisher Scientific, Inc.), with GAPDH as an
internal control gene. The thermocycling conditions for qPCR were
as follows: 95°C for 10 min, followed by 40 cycles of 95°C for 15
sec and 60°C for 1 min. The following PCR primer pairs were
designed: miR-103 forward: 5′-GAGCAGCATTGTACAG-3′, reverse:
5′-GTGCAGGGTCCGAGGT-3′; U6 forward: 5′-CTCGCTTCGGCAGCACA-3′,
reverse: 5′-AACGCTTCACGAATTTGCGT-3′; BDNF forward:
5′-TCCCTGGCTGACACTTTT-3′, reverse: 5′-ATTGGGTAGTTCGGCATT-3′; GAPDH
forward: 5′-CGGAGTCAACGGATTTGGTCGTAT-3′, reverse:
5′-AGCCTTCTCCATGGTGGTGAAGAC-3′. Relative gene expression was
calculated using the 2−ΔΔCq method (28).
MTT assay
MTT assay (Sigma-Aldrich, Merck KGaA, Darmstadt,
Germany) was used to determine cell proliferation. Cells were
seeded in 96-well plates at a density of 3×103
cells/well and cultured overnight at 37°C with 5% CO2.
Subsequently, transfection was performed, as described above, and
the samples were incubated at 37°C with 5% CO2 for 0,
24, 48 and 72 h. At each time point, 10 µl MTT solution (5 mg/ml)
was added into each well and incubated for additional 4 h, and
cells were then lysed in 150 µl dimethylsulfoxide at room
temperature for 10 min. The absorbance was measured at a wavelength
of 490 nm with a SpectraMax M3 microplate reader (Molecular
Devices, LLC, Sunnyvale, CA, USA). Each assay was performed in
triplicate and repeated independently at least three times.
Transwell invasion assay
Transwell filters (pore size, 8 µm; EMD Millipore,
Billerica, MA, USA) coated with Matrigel (BD Biosciences, Franklin
Lakes, NJ, CA, USA) were used to examine cell invasion ability.
Cells were harvested 48 h post-transfection and 5×104
cells were resuspended in 200-µl FBS-free DMEM and placed in the
upper chamber. A total of 500 µl DMEM containing 20% FBS was added
into the lower chamber as a chemo-attractant. Cells were then
incubated at 37°C with 5% CO2 for 24 h. Cells on the
upper surface of the membrane were removed with cotton swabs. Cells
that invaded to the bottom of the insert were fixed in 4%
paraformaldehyde at room temperature for 10 min, stained with 0.1%
crystal violet at room temperature for 10 min and washed in PBS.
Finally, the invasive cells were counted under a light microscope
(Olympus IX53; Olympus Corporation, Tokyo, Japan) in five
independent fields/well (magnification, ×200).
Bioinformatics analysis
TargetScan Human online software (version 7.0;
www.targetscan.org) and miRBase database
(http://www.mirbase.org/) were used to predict the
targets of miR-103.
Luciferase reporter assay
For the luciferase reporter assay, a reporter
plasmid containing the wild-type (pmirGLO-BDNF-3′-UTR Wt) or mutant
(pmirGLO-BDNF-3′-UTR Mut) 3′-UTR of BDNF were synthesized by
Shanghai GenePharma Co., Ltd. Cells were seeded in 24 well plates
at a density for 1.5×105 cells per well and incubated
overnight at 37°C with 5% CO2. Cells were then
transfected with miRNA mimics (50 pmol; miR-103 mimics or miR-NC)
and reporter plasmids (0.2 µg; pmirGLO-BDNF-3′-UTR Wt or
pmirGLO-BDNF-3′-UTR Mut) using Lipofectamine 2000, according to the
manufacturer's protocol. Following a 24 h transfection, the cells
were harvested and luciferase activities were determined using the
Dual-Luciferase Reporter Assay system (Promega Corporation)
according to the manufacturer's protocol. The Renilla
luciferase activity was normalized to the Firefly luciferase
activity. This assay was performed in triplicate and repeated three
times independently.
Protein isolation and western blot
analysis
Proteins were extracted from tissue samples or cells
using a protein lysis buffer (Xinyu Biotechnology, Shanghai, China)
supplemented with a protease and phosphatase inhibitor cocktail
(Roche Diagnostics, Basel, Switzerland). A Bicinchoninic Acid
Protein Assay kit (Pierce; Thermo Fisher Scientific, Inc.) was
applied to measure protein concentration. Equal quantities of
proteins (30 µg) were analyzed by 10% SDS-PAGE and transferred onto
polyvinylidene fluoride membranes (EMD Millipore). The membranes
were further blocked with 5% skimmed milk at room temperature for 2
h and incubated overnight at 4°C with monoclonal primary
antibodies: Anti-mouse anti-human BDNF (cat. no. ab10505; 1:1,000
dilution; Abcam, Cambridge, UK) or anti-mouse anti-human GAPDH
(cat. no. sc-47724; 1:1,000 dilution; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA). Subsequently, the membranes were incubated
with a corresponding horseradish peroxidase-conjugated secondary
antibody (cat. no. c-2005; 1:5,000 dilution; Santa Cruz
Biotechnology, Inc.) at room temperature for 2 h. Protein blots
were detected using the Enhanced Chemifluorescence western blotting
kit (Pierce; Thermo Fisher Scientific, Inc.). Signal intensity was
quantified using ImageJ software (version 1.48; National Institutes
of Health, Bethesda, MA, USA). GAPDH was used as control.
Statistical analysis
Data are presented as the mean ± standard deviation.
Comparisons between groups were made with Student's t-tests or one
way analysis of variance using SPSS software (version 18.0, SPSS,
Inc., Chicago, IL, USA). Student-Newman-Keuls analysis was used to
compare differences between groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
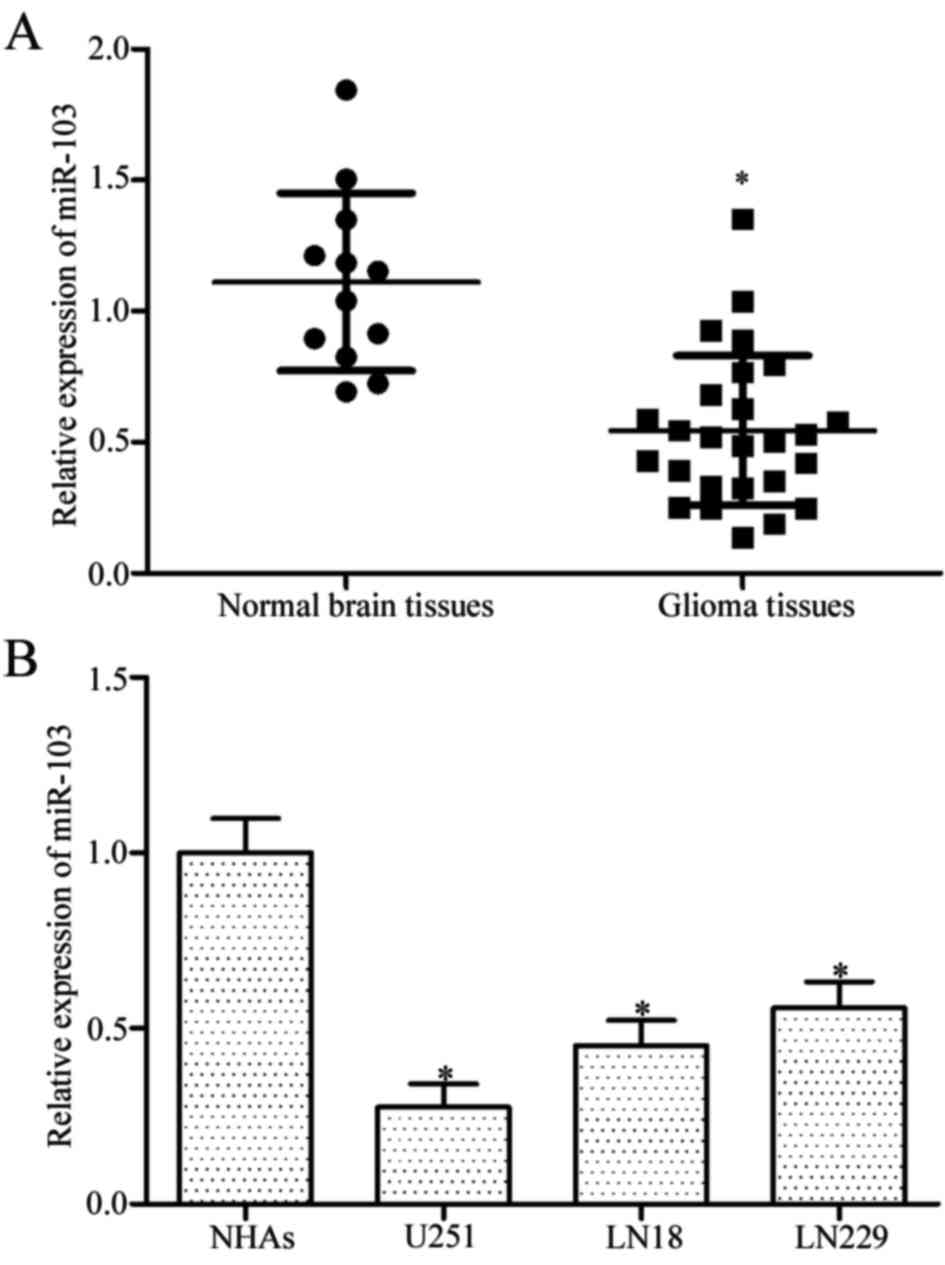
miR-103 is significantly
down-regulated in glioma tissues and cell lines
The expression level of miR-103 in 26 glioma tissues
and 12 normal brain tissues was measured using RT-qPCR to assess
the potential role of miR-103 in glioma. Results demonstrated that
miR-103 was down-regulated in glioma tissues compared with the
normal brain tissues (Fig. 1A,
P<0.05). Furthermore, miR-103 expression was detected using
RT-qPCR in three glioma cell lines (U251, LN18 and LN229) and NHAs.
The expression level of miR-103 decreased in glioma cell lines
compared with NHAs (Fig. 1B,
P<0.05). The above data support the hypothesis that miR-103 is
down-regulated in glioma tissues and cell lines, suggesting that
miR-103 downregulation may be involved in glioma initiation and
progression.
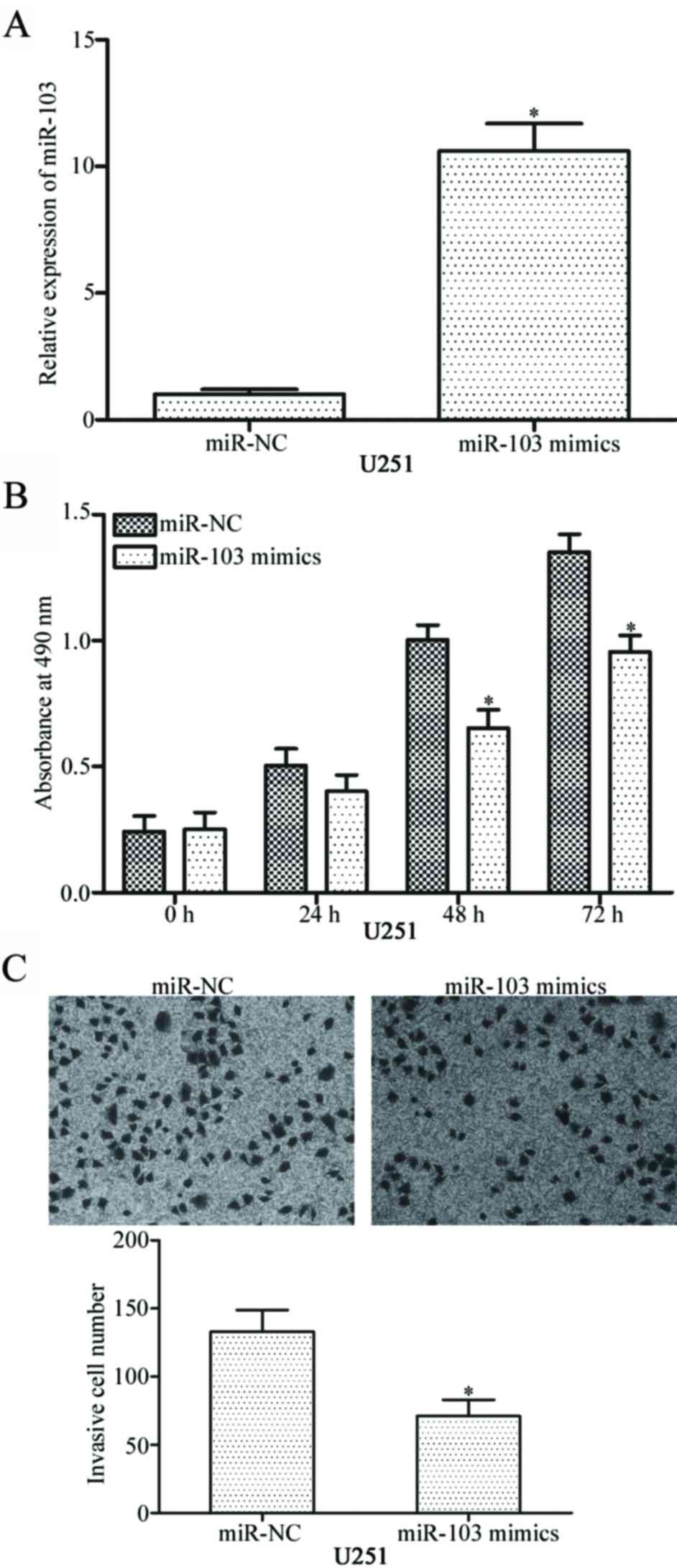
miR-103 inhibits the proliferation and
invasion of glioma cells in vitro
To investigate the roles of miR-103 in glioma, U251
cells were transfected with miR-103 mimics to increase the
intracellular levels of miR-103. RT-qPCR confirmed that miR-103
expression was markedly up-regulated in U251 cells transfected with
miR-103 mimics compared with cells transfected with miR-NC
(Fig. 2A, P<0.05).
MTT assay was performed to investigate the effect of
miR-103 overexpression on glioma cell proliferation. As presented
in Fig. 2B, miR-103 up-regulation
inhibited the proliferation of U251 cells compared with
miR-NC-transfected cells (P<0.05). Transwell invasion assay was
further used to examine the invasive capability of glioblastoma
cells. Ectopic expression of miR-103 significantly decreased the
invasive capacities of U251 cells (Fig. 2C, P<0.05). These results of the
present study indicate that miR-103 suppresses the proliferation
and invasion of glioma cells.
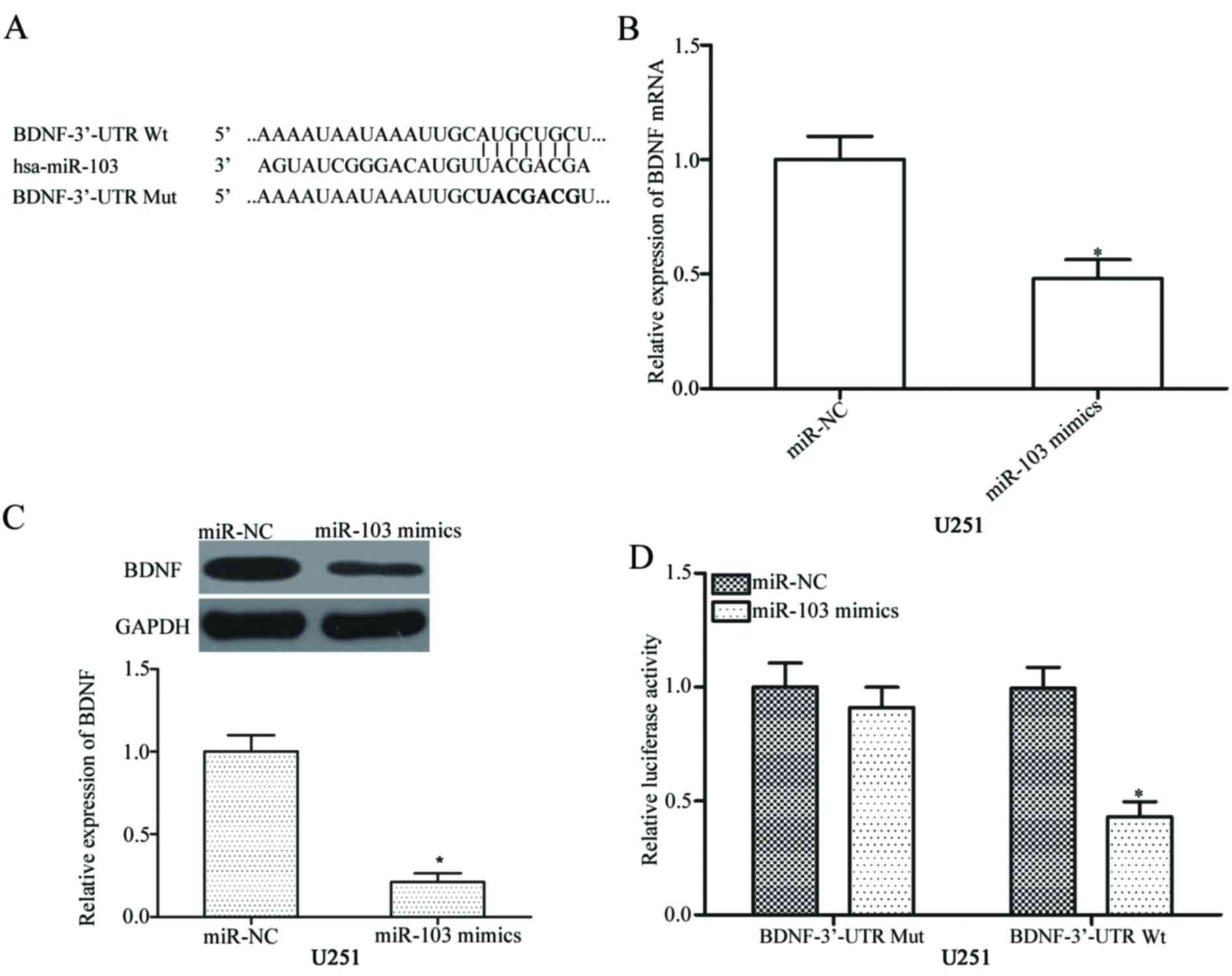
BDNF is a direct target of miR-103 in
glioma
Potential targets of miR-103 in glioma cells were
investigated to elucidate the molecular mechanisms underlying
miR-103-mediated inhibition of proliferation and invasion of glioma
cells. Bioinformatics analysis was performed to predict candidate
targets of miR-103. The analysis indicated a potentially
complementary region between the seed region sequence of miR-103
and nucleotide 299–305 region in the 3′-UTR of BDNF (Fig. 3A). To confirm this specificity,
regulatory effects of miR-103 on the endogenous BDNF expression
were investigated. RT-qPCR and western blot analysis revealed that
miR-103 overexpression suppressed BDNF expression in U251 cells at
both mRNA and protein levels (Fig. 3B
and C respectively, both P<0.05). Luciferase reporter assay
was conducted to further identify the direct interaction between
miR-103 and the 3′-UTR of BDNF. Luciferase activity was decreased
in the U251 cells co-transfected with the pmirGLO-BDNF-3′-UTR Wt
vector and miR-103 mimics (Fig.
3D, P<0.05). The pmirGLO-BDNF-3′-UTR Mut vector along with
the miR-103 mimic exerted no effect on luciferase activities. Above
data suggest that BDNF is a direct target of miR-103 in glioma.
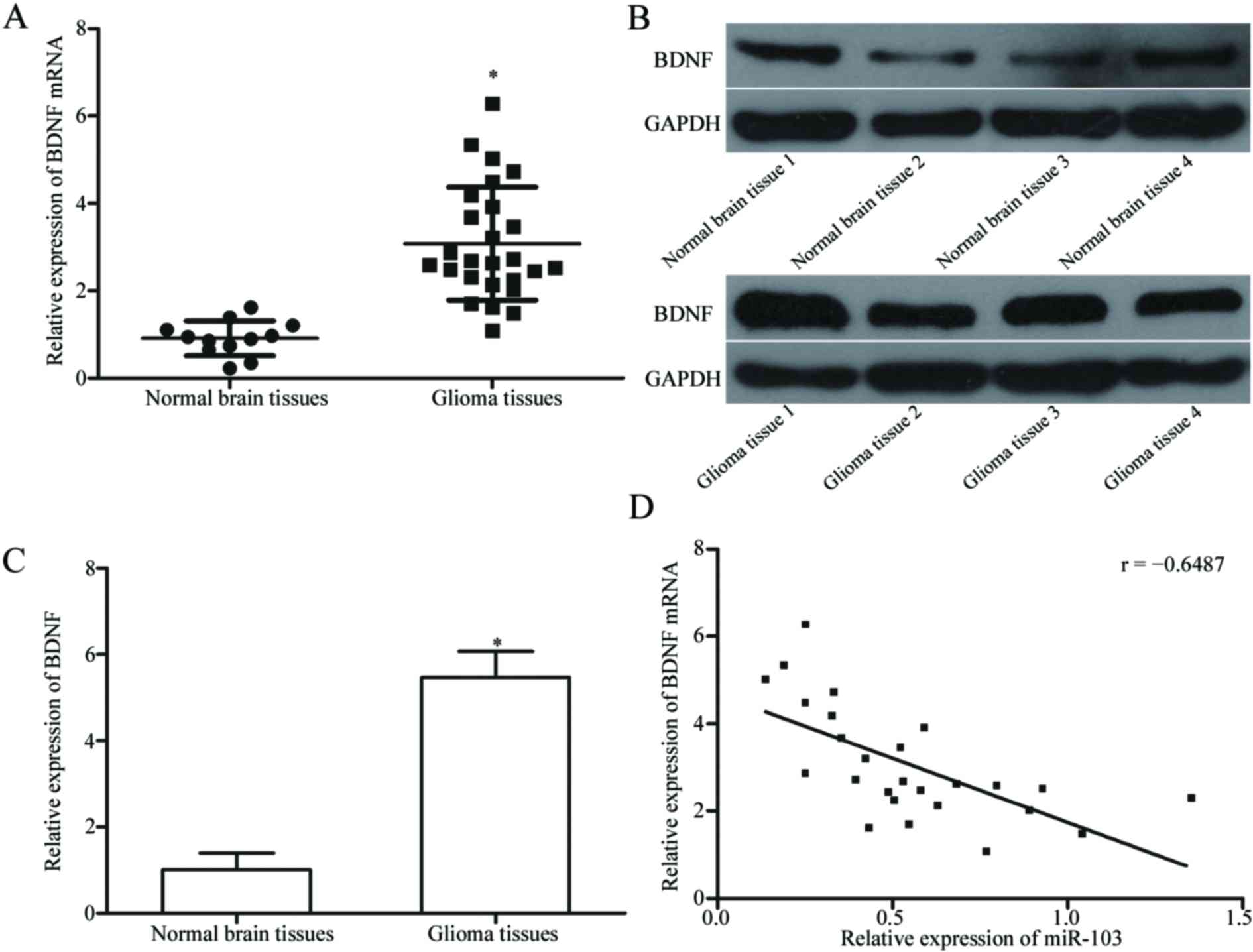
miR-103 expression is negatively
associated with BDNF levels in glioma tissues
mRNA and protein levels of BDNF in glioma tissues
and normal brain tissues were measured using RT-qPCR and western
blotting, respectively to verify the association between the
expression of miR-103 and BDNF. The results demonstrated that mRNA
and protein levels of BDNF were significantly increased in glioma
tissues compared with normal brain tissues (Fig. 4A-C, both P<0.05).
Spearman's correlation analysis demonstrated a
negative association between miR-103 and BDNF mRNA expression
levels in glioma tissues (Fig. 4D;
r=−0.6487; P=0.0003). The above results suggest that miR-103
down-regulation may cause BDNF up-regulation in glioma.
BDNF overexpression counteracts the
suppressive effects of miR-103 on glioma cells
Considering that BDNF is a direct target of miR-103,
it was investigated whether BDNF is involved in the regulation of
miR-103-mediated inhibition of glioma cell proliferation and
invasion. U251 cells were transfected with miR-103 mimics and
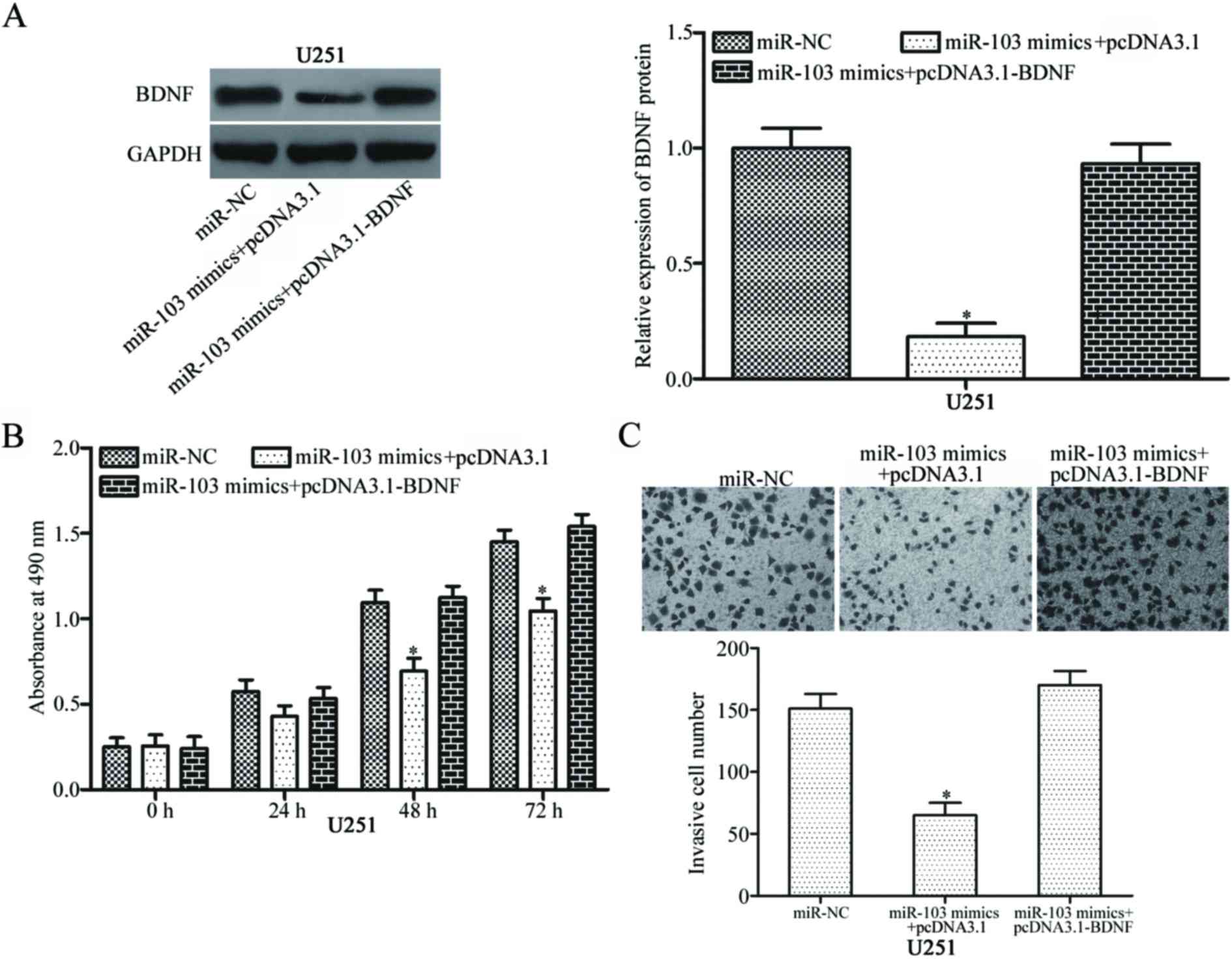
pcDNA3.1 or pcDNA3.1-BDNF. Western blot analysis indicated that
BDNF down-regulation induced by miR-103 was counteracted by BDNF
overexpression (Fig. 5A,
P<0.05). Functional rescue experiments revealed that BDNF
up-regulation significantly decreased the suppressive effects of
miR-103 on glioma cell proliferation and invasion (Fig. 5B and C; both P<0.05). The above
results suggest that miR-103 may inhibit the proliferation and
invasion of glioma cells by suppressing BDNF expression.
Discussion
miRNAs are involved in the tumour development of
various human cancers, including glioma (29–31).
Dysregulation of miRNAs is a common feature of human malignancies,
as they regulate the expression of oncogenes and tumour suppressors
(32). Therefore, miRNAs may
possess potential tumour diagnostic, prognostic and therapeutic
values in human glioma. In the present study, miR-103 was
down-regulated in glioma tissues and cell lines. In addition,
restoration of normal-cell levels of expression of miR-103
attenuated glioma cell proliferation and invasion in vitro.
Furthermore, BDNF was validated as a direct target of miR-103 in
glioma. The above results indicate that miR-103 acts as a tumour
suppressor in glioma, suggesting its potential use in the treatment
of patients with glioma.
miR-103 is down-regulated in several human cancers,
including gastric cancer tissues and cell lines. Low miR-103
expression is associated with pathological (p)M stage in the tumor,
node and metastasis classification of patients with gastric cancer
(33). In the prostate cancer,
miR-103 expression is reduced in tumour tissues (27). However, miR-103 is overexpressed in
colorectal cancer. High miR-103 expression is associated with an
aggressive phenotype of colorectal cancer and poor overall survival
of patients (34). miR-103 is also
up-regulated in breast cancer and hepatocellular carcinoma
(35,36). Serum levels of miR-103 in breast
cancer are associated with the advanced clinical stage and lymph
node metastasis (35). The above
results indicate that the expression patterns of miR-103 are tissue
specific and can be used as a predictive value for patient
prognosis in human cancer.
miR-103 has been previously identified to serve
tumour-suppressing roles in multiple human malignancies. For
instance, miR-103 up-regulation suppresses cell growth and
metastasis in gastric cancer (33). Restoration of the expression of
miR-103 inhibits prostate cancer cell proliferation and invasion
and increases the duration of the G1 phase of the cell cycle
(27). miR-103 was also identified
as an oncogene in colorectal cancer, promoting cell growth and
metastasis both in vitro and in vivo (25,34).
Yu et al (26) revealed
that miR-103 downregulation attenuates cell proliferation and
invasion of endometrial cancer. Xia et al (36) reported that miR-103 overexpression
promotes cell proliferation and decreases apoptosis in
hepatocellular carcinoma. The above, conflicting results revealed
that the biological functions of miR-103 are tissue specific, an
observation which can be explained by imperfect complementarity
between miRNAs and their target genes. Therefore, miR-103 should be
investigated as a potential target for the therapeutic treatment of
specific types of cancer.
Previous studies have identified several miR-103
targets, including: i) c-Myb proto-oncogene in gastric cancer; ii)
large tumor suppressor kinase 2, endoribonuclease DICER and
phosphatase and tensin homolog in colorectal cancer; iii) tissue
inhibitor of metallopeptidases-3 in endometrial cancer; iv)
programmed cell death 10 in prostate cancer; and v) A-kinase
anchoring protein 12 in hepatocellular carcinoma (25–27,33,34,36).
In the present study, BDNF was validated as a novel target of
miR-103. A bioinformatics prediction indicated that BDNF is a
potential target of miR-103. RT-qPCR and western blotting
demonstrated that miR-103 negatively modulated the mRNA and protein
expression levels of BDNF in glioma cells. Luciferase reporter
assay revealed that the 3′-UTR of BDNF could be directly targeted
by miR-103 in glioma cells. Furthermore, BDNF was significantly
upregulated in glioma tissues and negatively associated with
miR-103 expression. Rescue experiments demonstrated that BDNF
overexpression substantially reversed the suppressive effects of
miR-103 on glioma cell proliferation and invasion. The above
results indicate that BDNF is a direct and functional target gene
of miR-103 in glioma.
BDNF, located on the short arm of chromosome 11
(11p13), is up-regulated in various human cancer types, including
gastric, lung, cervical, breast and colorectal cancers, and
hepatocellular carcinoma (37–42).
BDNF has been recently identified to act as a cancer regulator and
serve a role in cancer occurrence and development (43). In glioma, BDNF is highly expressed
in tumour tissues and is associated with high pathological grading
scores (44). Functional assays
indicated that BDNF is an oncogene in glioma as it promotes cell
growth and metastasis, and inhibits apoptosis (45). Therefore, BDNF may be a promising
therapeutic target for the treatment of patients with glioma.
In conclusion, the present study provided evidence
that miR-103 is down-regulated in glioma tissues and cell lines.
miR-103 serves a tumour-suppressive role in glioma by
down-regulating BDNF. The results of the present study indicate
that the miR-103/BDNF interaction is a potential therapeutic target
for the treatment of glioma. However, the use of a single cell line
is a limitation of the present study, and the results should be
validated using multiple cell lines.
References
|
1
|
Goodenberger ML and Jenkins RB: Genetics
of adult glioma. Cancer Genet. 205:613–621. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Buckner JC, Brown PD, O'Neill BP, Meyer
FB, Wetmore CJ and Uhm JH: Central nervous system tumors. Mayo Clin
Proc. 82:pp. 1271–1286. 2007; View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu X, Wang S, Yuan A, Yuan X and Liu B:
MicroRNA-140 represses glioma growth and metastasis by directly
targeting ADAM9. Oncol Rep. 36:2329–2338. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhu GY, Shi BZ and Li Y: FoxM1 regulates
Sirt1 expression in glioma cells. Eur Rev Med Pharmacol Sci.
18:205–211. 2014.PubMed/NCBI
|
|
5
|
Chen J, McKay RM and Parada LF: Malignant
glioma: Lessons from genomics, mouse models, and stem cells. Cell.
149:36–47. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stupp R, Mason WP, Van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Van Meir EG, Hadjipanayis CG, Norden AD,
Shu HK, Wen PY and Olson JJ: Exciting new advances in
neuro-oncology: The avenue to a cure for malignant glioma. CA
Cancer J Clin. 60:166–193. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lefranc F, Brotchi J and Kiss R: Possible
future issues in the treatment of glioblastomas: Special emphasis
on cell migration and the resistance of migrating glioblastoma
cells to apoptosis. J Clin Oncol. 23:2411–2422. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Onishi M, Ichikawa T, Kurozumi K and Date
I: Angiogenesis and invasion in glioma. Brain Tumor Pathol.
28:13–24. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Carrington JC and Ambros V: Role of
microRNAs in plant and animal development. Science. 301:336–338.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qin B, Yang H and Xiao B: Role of
microRNAs in endothelial inflammation and senescence. Mol Biol Rep.
39:4509–4518. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kotaja N: MicroRNAs and spermatogenesis.
Fertil Steril. 101:1552–1562. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Imbar T and Eisenberg I: Regulatory role
of microRNAs in ovarian function. Fertil Steril. 101:1524–1530.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Garg D and Cohen SM: miRNAs and aging: A
genetic perspective. Ageing Res Rev. 17:3–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dogini DB, Pascoal VD, Avansini SH, Vieira
AS, Pereira TC and Lopes-Cendes I: The new world of RNAs. Genet Mol
Biol. 37 1 Suppl:S285–S293. 2014. View Article : Google Scholar
|
|
16
|
Hong L, Ya-Wei L, Hai W, Qiang Z, Jun-Jie
L, Huang A, Song-Tao Q and Yun-Tao L: MiR-519a functions as a tumor
suppressor in glioma by targeting the oncogenic STAT3 pathway. J
Neurooncol. 128:35–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Teoh SL and Das S: The role of MicroRNAs
in diagnosis, prognosis, metastasis and resistant cases in breast
cancer. Curr Pharm Des. 23:1845–1859. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu HT, Wang YW, Xing AY, Shi DB, Zhang H,
Guo XY, Xu J and Gao P: Prognostic value of microRNA signature in
patients with gastric cancers. Sci Rep. 7:428062017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dong F, Xu T, Shen Y, Zhong S, Chen S,
Ding Q and Shen Z: Dysregulation of miRNAs in bladder cancer:
Altered expression with aberrant biogenesis procedure. Oncotarget.
8:27547–27568. 2017.PubMed/NCBI
|
|
20
|
Liu L, Bi N, Wu L, Ding X, Men Y, Zhou W,
Li L, Zhang W, Shi S, Song Y and Wang L: MicroRNA-29c functions as
a tumor suppressor by targeting VEGFA in lung adenocarcinoma. Mol
Cancer. 16:502017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen Y, Gao DY and Huang L: In vivo
delivery of miRNAs for cancer therapy: Challenges and strategies.
Adv Drug Deliv Rev. 81:128–141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Turner JD, Williamson R, Almefty KK,
Nakaji P, Porter R, Tse V and Kalani MY: The many roles of
microRNAs in brain tumor biology. Neurosurg Focus. 28:E32010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kong YW, Ferland-McCollough D, Jackson TJ
and Bushell M: microRNAs in cancer management. Lancet Oncol.
13:e249–e258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Geng L, Sun B, Gao B, Wang Z, Quan C, Wei
F and Fang XD: MicroRNA-103 promotes colorectal cancer by targeting
tumor suppressor DICER and PTEN. Int J Mol Sci. 15:8458–8472. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu D, Zhou H, Xun Q, Xu X, Ling J and Hu
Y: microRNA-103 regulates the growth and invasion of endometrial
cancer cells through the downregulation of tissue inhibitor of
metalloproteinase 3. Oncol Lett. 3:1221–1226. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fu X, Zhang W, Su Y, Lu L, Wang D and Wang
H: MicroRNA-103 suppresses tumor cell proliferation by targeting
PDCD10 in prostate cancer. Prostate. 76:543–551. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Neviani P and Fabbri M: Exosomic microRNAs
in the tumor microenvironment. Front Med (Lausanne).
2:472015.PubMed/NCBI
|
|
30
|
Zhao Y, Pang D, Wang C, Zhong S and Wang
S: MicroRNA-134 modulates glioma cell U251 proliferation and
invasion by targeting KRAS and suppressing the ERK pathway. Tumour
Biol. 37:11485–11493. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tian LQ, Liu EQ, Zhu XD, Wang XG, Li J and
Xu GM: MicroRNA-197 inhibits cell proliferation by targeting GAB2
in glioblastoma. Mol Med Rep. 13:4279–4288. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in Cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liang J, Liu X, Xue H, Qiu B, Wei B and
Sun K: MicroRNA-103a inhibits gastric cancer cell proliferation,
migration and invasion by targeting c-Myb. Cell Prolif. 48:78–85.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zheng YB, Xiao K, Xiao GC, Tong SL, Ding
Y, Wang QS, Li SB and Hao ZN: MicroRNA-103 promotes tumor growth
and metastasis in colorectal cancer by directly targeting LATS2.
Oncol Lett. 12:2194–2200. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang X, Wu X, Yan L and Shao J: Serum
miR-103 as a potential diagnostic biomarker for breast cancer. Nan
Fang Yi Ke Da Xue Xue Bao. 32:631–634. 2012.(In Chinese).
PubMed/NCBI
|
|
36
|
Xia W, Ni J, Zhuang J, Qian L, Wang P and
Wang J: MiR-103 regulates hepatocellular carcinoma growth by
targeting AKAP12. Int J Biochem Cell Biol. 71:1–11. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Choi B, Lee EJ, Shin MK, Park YS, Ryu MH,
Kim SM, Kim EY, Lee HK and Chang EJ: Upregulation of brain-derived
neurotrophic factor in advanced gastric cancer contributes to bone
metastatic osteolysis by inducing long pentraxin 3. Oncotarget.
7:55506–55517. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang SY, Hui LP, Li CY, Gao J, Cui ZS and
Qiu XS: More expression of BDNF associates with lung squamous cell
carcinoma and is critical to the proliferation and invasion of lung
cancer cells. BMC Cancer. 16:1712016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Moon A, Won KY, Lee JY, Kang I, Lee SK and
Lee J: Expression of BDNF, TrkB, and p53 in early-stage squamous
cell carcinoma of the uterine cervix. Pathology. 43:453–458. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Patani N, Jiang WG and Mokbel K:
Brain-derived neurotrophic factor expression predicts adverse
pathological & clinical outcomes in human breast cancer. Cancer
Cell Int. 11:232011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang X, Martin TA and Jiang WG: Biological
influence of brain-derived neurotrophic factor (BDNF) on colon
cancer cells. Exp Ther Med. 6:1475–1481. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yang ZF, Ho DW, Lam CT, Luk JM, Lum CT, Yu
WC, Poon RT and Fan ST: Identification of brain-derived
neurotrophic factor as a novel functional protein in hepatocellular
carcinoma. Cancer Res. 65:219–225. 2005.PubMed/NCBI
|
|
43
|
Tajbakhsh A, Mokhtari-Zaer A, Rezaee M,
Afzaljavan F, Rivandi M, Hassanian SM, Ferns GA, Pasdar A and Avan
A: Therapeutic potentials of BDNF/TrkB in breast cancer; current
status and perspectives. J Cell Biochem. 118:2502–2515. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yan Q, Yu HL and Li JT: Study on the
expression of BDNF in human gliomas. Sichuan Da Xue Xue Bao Yi Xue
Ban. 40:415–417. 2009.(In Chinese). PubMed/NCBI
|
|
45
|
Xiong J, Zhou L, Lim Y, Yang M, Zhu YH, Li
ZW, Zhou FH, Xiao ZC and Zhou XF: Mature BDNF promotes the growth
of glioma cells in vitro. Oncol Rep. 30:2719–2724. 2013. View Article : Google Scholar : PubMed/NCBI
|