Introduction
Steroid-induced avascular necrosis of the femoral
head (SANFH), as the most common type of femoral head necrosis,
also leads to femoral head collapse and osteoarthritis if not being
treated properly (1,2). Finally, most patients require total
hip arthroplasty. Nowadays, Dexamethasone (Dex) and other
glucocorticoids (GCs) are of great importance in curing many
diseases such as serious infection, inflammation, anti-shock,
allergic disease, hematological system diseases and replacement
therapy (3–6). Since the wide application of Dex and
other GCs, the incidence of SANFH has already exceeded
trauma-induced femoral head necrosis (7). However, the pathogenesis of SANFH is
still expecting for further research. It is urgent to clarify the
pathogenesis of SANFH, and find out a proper treatment for
SANFH.
The imbalance between osteoblast and osteoclast
activity, by inhibiting osteoblastogenesis and promoting osteoblast
apoptosis, will finally cause osteonecrosis (8,9).
Osteoblast, which is the main target of Dex and other GCs, plays a
vital role in promoting bone formation (10,11).
Dex-induced osteoblasts apoptosis was used to create a cellular
model of SANFH, which has been proved to be feasible in our recent
study (12).
Salidroside (Sal), which is one important medicinal
ingredient of Rhodiola rosea L, is useful in anti-aging,
memory improvement, anti-fatigue, anti-oxidation, anti-depressant
and so on (13–17). Previous studies found Sal could
reduce apoptosis induced by glutamate in rat primary cultured
hippocampal neurons (18). Sal
attenuates colistin-induced neurotoxicity in RSC96 Schwann cells
via activating the phosphoinositide 3-kinase (PI3K)/ protein kinase
B (Akt) signaling pathway and downregulating caspase-3 expression
(19). The PI3K/Akt pathway is a
critical pathway related to several fundamental cellular processes,
such as survival, proliferation, growth and differentiation
(20–24). Recently, a study showed that the
suppression of the PI3K/Akt signaling pathway caused osteoblast
damage (25). Caspase-3, a subtype
of the caspase proteins as dormant proenzymes in cells, cleaves
cellular substrates. Caspase-3 also causes cell apoptosis whether
it is activated by the intrinsic pathway or the extrinsic pathway
(26,27).
In our study, we employed primary osteoblasts in
vitro and rats in vivo to find out the anti-apoptosis
effect of Sal on SANFH. Furthermore, we also tried to explore the
mechanism of the protective effect of Sal. We assumed Sal had an
anti-apoptosis effect on SANFH via activating the PI3K/Akt
signaling pathway and subsequently downregulating the caspase-3
expression.
Materials and methods
Preparation of primary murine
osteoblasts
According to our previous study (12), osteoblasts were isolated and
extracted by digesting neonatal rats' calvariae using collagenase I
(Sigma-Aldrich, St. Louis, MA, USA). Digests 3–6 were neutralized,
pooled, and filtered. After we resuspended the single-cell
suspensions, the single-cell suspensions was cultured in
alpha-Minimal Essential Medium (α-MEM) which supplemented with 12%
fetal bovine serum (FBS) and antibiotics. The operation procedures
were approved by the Animal Care and Use Committee at our
institution and complied with the guidelines of the international
regulations.
Cell survival assay, cell apoptosis
assay and LDH release assay
The Cell Counting kit-8 (CCK-8; Dojindo
Laboratories, Kumamoto, Japan) and Cell Apoptosis-DNA ELISA Plus
kit (Roche, Palo Alto, CA, USA) was respectively used as
quantitative measurement to assess the cell survival and the cell
apoptosis. We recorded both the optic density (OD) of each well at
450 nm.
The lactate dehydrogenase (LDH) in the conditional
medium was also measured by a two-step enzymatic reaction LDH assay
kit (Takara, Tokyo, Japan) to assess the cell apoptosis. The
percentage of LDH release=LDH (conditional medium)/LDH (conditional
medium + cell lysates).
TUNEL assay
We used TUNNEL staining to detect the apoptotic
osteoblasts by the In Situ Cell Death Detection kit (Roche). The
cultured osteoblasts were fixed with freshly 4% paraformaldehyde
for 1 h in a six-well plate. Then they were incubated with 3%
H2O2 and 0.1% Triton X-100 for 10 min. During
each step, they were washed three times with phosphate buffered
saline (PBS). Finally, cells were stained with DAPI. A fluorescence
microscope (Olympus, Tokyo, Japan) was used to obtain the images
and assess the apoptotic cells. The TUNEL positive cells were
calculated by three different authors.
Western blot analysis
As described (12),
we used six-well plates to culture osteoblasts (3×105
cells/well) and added Dex or/and Sal at different time. In the Sal
group and Dex + Sal group, osteoblasts were pretreated with the
optimum concentration of Sal (100 nM) for 2 h before the addition
of Dex. We used radioimmunoprecipitation assay (RIPA) buffer
containing phenylmethane sulfonyl fluoride (PMSF) to extract
proteins from the osteoblasts. Then the protein concentration was
calculated by the BCA Protein Assay kit (Beyotime Biotechnology,
China). 12% sodium dodecyl sulfate polyacrylamide gel
electrophoresis (SDS-PAGE) was adopted to resolve each protein
lysate samples (30 µg in average). Then the solution was
transferred onto polyvinylidene fluoride (PVDF) membranes. After
the membranes were blocked and incubated with the primary antibody
(1:1,000) overnight, they were incubated with each secondary
antibody (1:3,000) for 2 h. Electrochemiluminescence Plus Reagent
(Invitrogen Life Technologies, Carlsbad, CA, USA) was used to make
the bands visible. We also used Image Lab 3.0 software (Bio-Rad,
Hercules, CA, USA) to assess the band intensity. All the primary
antibodies against cleaved-caspase-3, cleaved caspase-9, BAX,
Bcl-2, cytochrome C, Akt and phosphor-Akt (Ser473) were purchased
from Cell Signaling Technology (Danvers, MA, USA).
Rat model in vivo
We chose 12-weeks-old adult male Sprague-Dawley rats
(Animal Center of the Chinese Academy of Sciences, Shanghai, China)
to build our animal model. We divided 45 rats equally into three
groups (the control group, the model group and the Sal group).
According to our previous study (12), The experimental rats in the model
group and the Sal group were administered with double doses of
lipopolysaccharide (LPS; Escherichia coli oup:B5;
Sigma-Aldrich) on day 0 and 1 and received intramuscular injection
of 40 mg/kg methlprednisolone (MPS) sodium succinate (Pfizer
Pharmaceutical, Beijing, China) on day 3, 4, and 5. The rats in the
control group were injected with normal saline (NS) instead. Then,
the rats in the Sal group received an intraperitoneal injection of
Sal (25 mg/kg/d) for seven consecutive days. The rest of the rats
were injected with NS. All the injections were carried out at the
same time each day and the time interval was 24 h.
We used 50×35×20 cm plexiglass cages to raise the
rats. We also controlled the laboratory conditions (relative
humidity of 50±10%, constant temperature of 20±2°C and 12 h
light/dark cycle). All the rats were allowed free access to food
and water. 8 weeks after the final injection of MPS, all the rats
were sacrificed and bilateral femoral heads were collected for
histological analysis. Our protocol was carried out in accordance
with the National Institutes of Health guide for the care and use
of Laboratory animals and was also approved by the Animal Care and
Use Committee of Wenzhou Medical University.
Histological analysis
Femoral head samples, collected from rat models
in vivo, were fixed in paraformaldehyde for 24 h at 4°C.
Then we transferred the samples to 10% EDTA solution at 4°C for 2
months. After the samples were dehydrated in ethanol, the samples
were embedded in paraffin and cut into 4-µm thick sections in the
coronal plane. We used hematoxylin-eosin for tissue section
staining. With a microscope (Leica Microsystems, Wetzlar, Germany),
we observed the pyknotic nuclei of osteocytes, empty lacunae, bone
marrow cell necrosis and bone trabeculae broken. The percentage of
empty lacunae was assessed blindly by three authors.
Statistical analysis
We repeated all the experiments at least three times
with consistent outcomes. We used SPSS 21.0 software (IBM SPSS,
Armonk, NY, USA) to convert all outcome measurements. The one-way
ANOVA followed by Tukey's post hoc test was used to calculate
statistical significance, and P<0.05 was considered to indicate
a statistically significant difference.
Results
In our previous study (12), we found Dex induced osteoblast
apoptosis related to dose and time. Cells exposed to 1 µM Dex for
24 h obviously damaged cell viability, and the dose and time of Dex
were used in our subsequent experiments.
Sal attenuates Dex-induced osteoblasts
apoptosis
Sal showed a significant protective effect on cell
survival assay and against Dex-induced cell death (cell apoptosis
assay and LDH release assay) when the concentration reached 10 nM
(P<0.05). And the protective effect of Sal was most obvious at
100 nM (P<0.01) (Fig. 1A-C).
Furthermore, we used CCK-8 to value the cytotoxic effect of Sal. As
shown in Fig. 1D, Sal had no
potential cytotoxic effect on the primary murine osteoblasts in Sal
alone group, which reflected in the decline of CCK-8 OD, when the
concentration was not more than 100 nM (P<0.05).
 | Figure 1.Sal protects osteoblasts from
Dex-induced damage. Primary murine osteoblasts were pretreated with
the indicated concentrations of Sal (0–1,000 nM) for 2 h, followed
by treatment with Dex (1 µM). Cells were then cultured in
conditional medium for 24 h, after which cell survival and
apoptosis were measured by the CCK-8 assay (A and D) and histoneDNA
ELISA (B). Furthermore, the LDH release was assayed (C). Sal showed
a significant protective effect on cell survival assay and against
Dex-induced cell death (cell apoptosis assay and LDH release assay)
when the concentration reached 10 nM (P<0.05) and 100 nM
(P<0.01) (A-C). Respectively, Sal had no potential cytotoxic
effect when the concentration was not more than 100 nM (P<0.05)
(D). Data are presented as the mean ± standard deviation.
Experiments in this figure were repeated five times. Significant
differences between two groups are indicated as **P<0.01 and
*P<0.05. Sal, salidroside; Dex, dexamethasone; CCK-8, cell
counting kit-8; OD, optical density; LDH, lactate
dehydrogenase. |
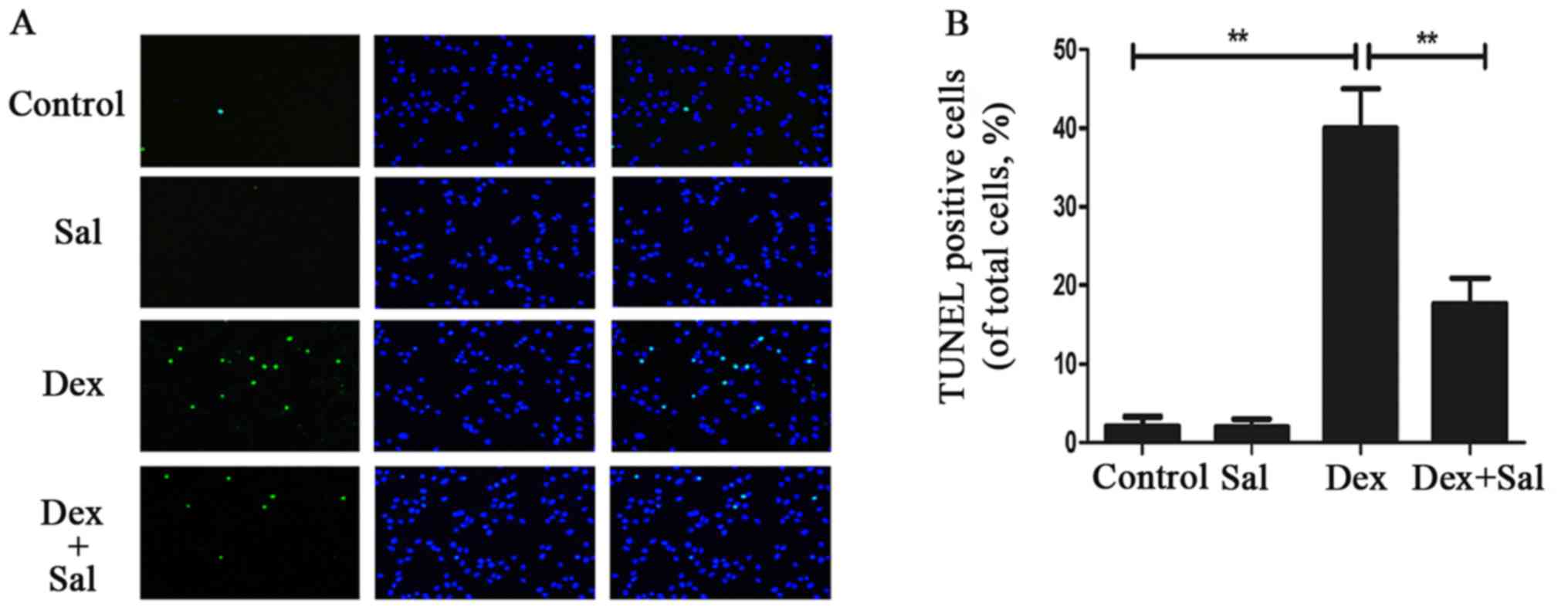
The TUNEL assay revealed that TUNEL positive cells
significantly increased when treated with Dex (P<0.01) (Fig. 2). And the pretreatment of Sal
obviously decreased TUNEL positive cells compared with the Dex
group (P<0.01) (Fig. 2). What's
more, there is no significant difference between the Sal alone
group and the control group (P>0.05).
Sal alleviated Dex-induced osteoblasts apoptosis via
activating the PI3K/Akt signaling pathway and downregulating the
caspase protein expression. To explore the mechanism by which Sal
decreased Dex-induced apoptosis, western blot analysis was
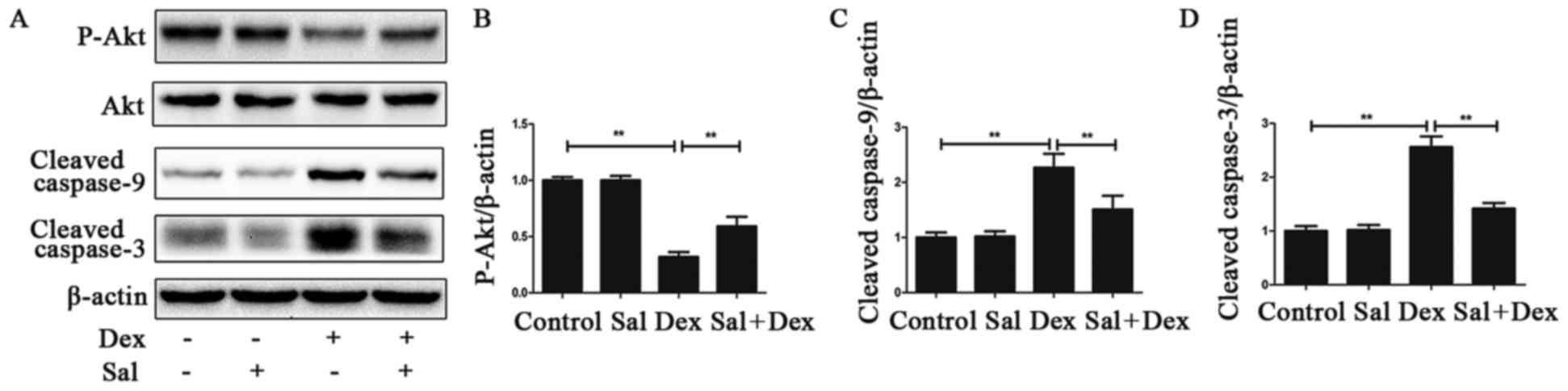
performed to study changes in the PI3K/Akt signaling pathway. As
shown in Fig. 3A, B, Dex
significantly decreased p-Akt level in the Dex alone group compared
with the control group (P<0.01). Pretreatment with Sal for 2 h
significantly increased p-Akt expression in Dex-induced osteoblasts
apoptosis (P<0.01). And the expression of p-Akt in the Sal alone
group did not show any change compared with the control group
(P>0.05). Total Akt levels in each group did not change.
To further investigate the effect of PI3K/Akt
signaling in the caspase protein expression, cells were pretreated
with Sal for 2 h and then stimulated with Dex for 24 h. Sal
significantly downregulated the expression of Dex-induced cleaved
caspase-9 and cleaved caspase-3 (P<0.01) (Fig. 3C, D).
Sal protected against Dex-induced
osteoblasts apoptosis by inhibiting the mitochondrial apoptosis
pathway
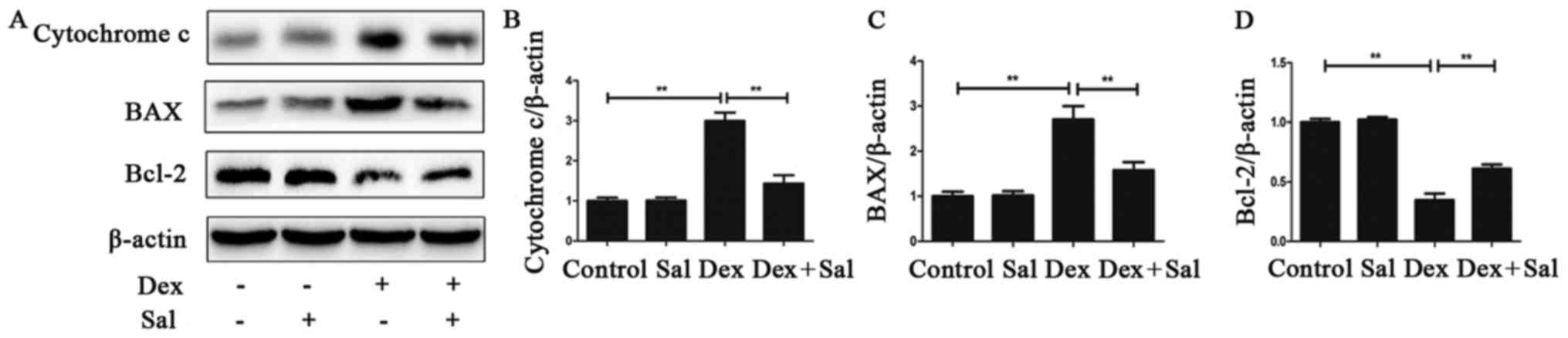
We used western blot analysis to explore whether the
mitochondrial pathway is involved in the Sal-induced protective
effect against Dex-induced osteoblasts apoptosis. The expression
levels of BAX, Bcl-2 and cytochrome C were detected. As shown in
Fig. 4, compared with the control
group, the expression level of pro-apoptotic protein Bax
significantly increased (P<0.01) while the expression level of
anti-apoptotic protein Bcl-2 was on the contrary in the Dex group
after exposure to Dex for 24 h (P<0.01). Pretreatment with Sal
for 2 h significantly decreased BAX expression and increased Bcl-2
expression in Dex-induced osteoblasts apoptosis (P<0.01).
Moreover, we found cytochrome C increased significantly by Dex
(P<0.01), but the expression could be inhibited by Sal
(P<0.01). And the expression of BAX, Bcl-2 and cytochrome C in
Sal alone group did not show any change compared with the control
group.
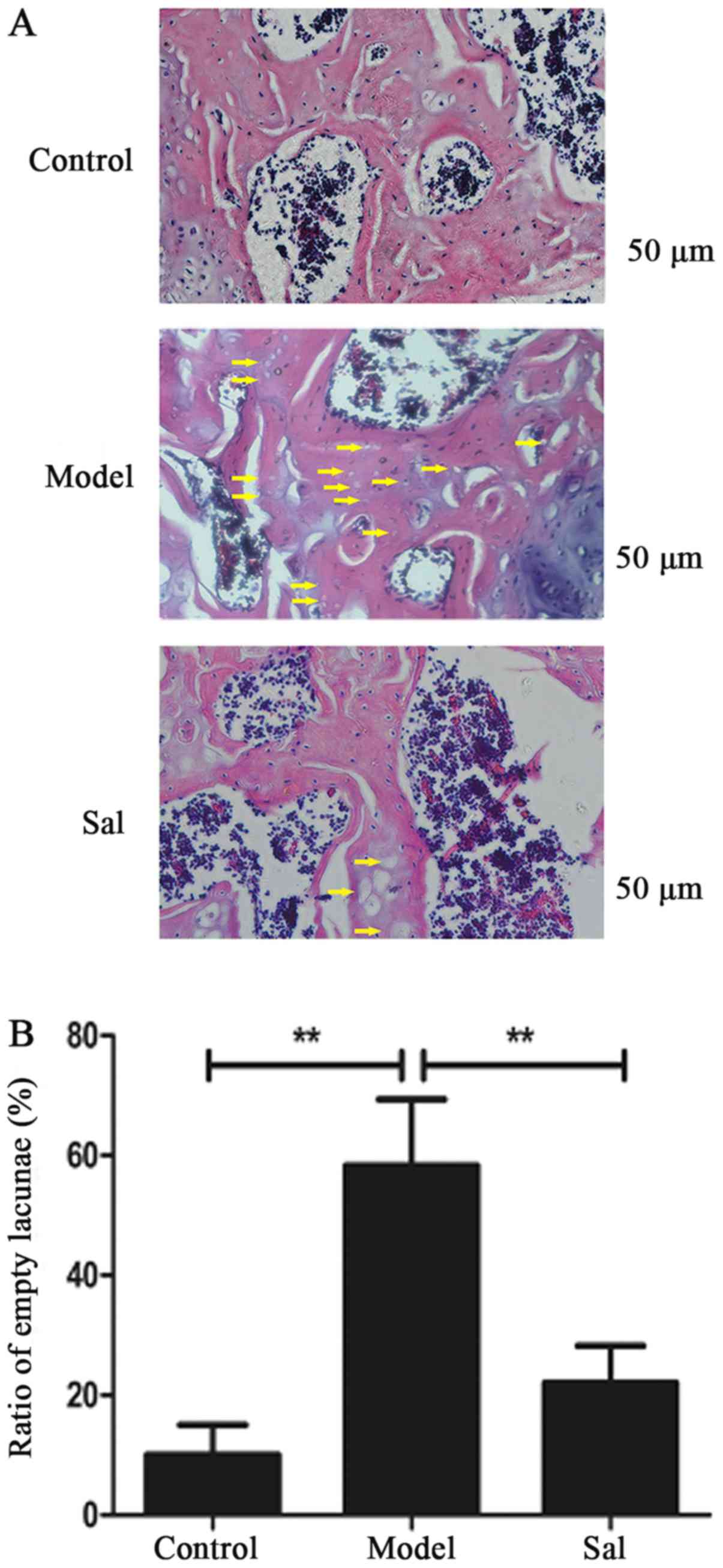
Sal protected against SANFH in rats by
decreasing the percentage of empty lacunae
The Sal group (2/15) significantly decreased the
incidence rate of SANFH compared with the model group (13/15)
(P<0.01). As shown in Fig. 5,
the percentage of empty lacunae increased significantly in the
model group (P<0.01). Additionally, Sal reduced the percentage
of empty lacunae compared with the model group (P<0.01) while
Sal would not increase the percentage of empty lacunae compared
with the control group.
Discussion
The present study showed that pretreatment with Sal
attenuated Dex-induced osteoblasts apoptosis by increasing cell
survival rate and decreasing cell apoptosis rate and LDH release
rate. Furthermore, the change of TUNEL positive cells in TUNEL
staining also proved our point. Sal decreased the expression of
cleaved caspase-3, cleaved caspase-9, BAX, cytochrome C and
increased Bcl-2 expression in Dex-induced osteoblasts apoptosis.
Sal protected against Dex-induced osteoblasts apoptosis in part
through the activation of the PI3K/Akt signaling pathway, which
might result in the inhibition of caspase-3 and caspase-9
expression. In vivo, the reduction of empty lacunae in
hematoxylin-eosin staining indicated Sal protected against
SANFH.
Nowadays, Dex and other GCs have been long used for
serious infection and immunological diseases and so on (3–6).
However, lots of studies indicate Dex and other GCs can induce
potential osteoblastic apoptosis which also leads to SANFH
(7,12). The mechanism of SANFH and the
protective method against SANFH are still unclear. Our previous
studies indicated that Dex could induce osteoblasts apoptosis and
SANFH in rats or human specimens via activating STAT1-mediated
upregulation of caspase-3 (12,28).
A recent study showed the activation of the PI3K/Akt signaling
Pathway was important in anti-apoptosis of osteoblasts (25). The mechanism of anti-osteoblasts
apoptosis seems to be not unique, and the PI3K/Akt signaling
pathway plays an important role. Studies which focused on nerve
cells showed the PI3K/Akt signaling pathway was a protective factor
of Sal on anti-apoptosis (19,29).
However, the protective effect of Sal on anti-osteoblasts apoptosis
has not been reported before. In our present study, we investigated
the effects of Sal in osteoblasts in vitro and in a rat
SANFH model in vivo.
Previous studies showed Dex and other GCs increased
the expression of caspase protein family especially caspase-3 which
was related to osteoblasts apoptosis (30) and other cell apoptosis (31,32).
Caspase-3 is of great importance in osteoblasts apoptosis related
to osteonecrosis (33), and the
obvious elevation is one of the typical pathological
characteristics of SANFH (28).
The PI3K/Akt pathway is a very important component of cell
survival, proliferation, and growth pathways in many different cell
types including osteoblast (19,29,30).
Akt which serves as the substrate of PI3K plays a vital role in the
antiapoptotic pathway (34,35).
Activated by PI3K, the Akt pathway caused the production of
phosphatidylinositol trisphosphate and the phosphorylation of
threonine-308 and serine-473 induces the activation of Akt
(19). Activation of the PI3K/Akt
pathway can prevent osteoblast against apoptosis, and the activated
Akt can reduce the risk of cell injury during oxidative stress,
free radical exposure and DNA damage (19,36).
Previous studies show Sal has an anti-apoptosis on cardiac H9c2
cells, PC12 cells and RSC96cells via activating the PI3K/Akt
pathway (19,37,38).
In our present study, we found that the activation of the PI3K/Akt
pathway played a crucial role in osteoblasts anti-apoptosis. We
guessed that Sal downregulating cleaved caspase-3 might be related
to the activation of the PI3K/Akt signaling pathway. However, the
exact mechanism is still unclear and further studies shall be done
to find out the truth.
Bcl-2 and BAX serve respectively as the
antiapoptotic and proapoptotic regulators. And the imbalance
between Bcl-2 and BAX, which stands for mitochondrial dysfunction,
is responsible for Dex-induced osteoblasts apoptosis (39,40).
Mitochondria induce cellular apoptosis by the release of cytochrome
C, and Sal can reverse it in Dex-induced osteoblasts apoptosis
(41). Cytochrome C promote the
generation of caspase-9, which subsequently activates caspase-3
expression (12). Our present
study demonstrated that Sal showed up anti-apoptotic effect partly
by inhibiting the mitochondrial pathway.
As described in our previous study (12), we adopted double dosage of LPS and
MPS to generate a modified rat SANFH model. Previous study
mentioned that the diffuse presence of empty lacunae in the bone
trabeculae could stand for the severity of SANFH (27). Steffen et al found a
significantly greater percentage of empty lacunae within the
trabecular bone in the avascular necrosis of femoral necrosis
(42). The empty lacunae represent
the bone necrosis degree and reflect the effect of bone repair to
some extent. As a result, we used hematoxylin-eosin staining to
calculate the percentage of empty lacunae in rats to evaluate the
effect of Sal on preventing SANFH. The reduction of empty lacunae
in Sal treated group indicated Sal protected against SANFH.
In conclusion, our present study demonstrated that
Sal alleviated Dex-induced osteoblasts apoptosis via activating the
PI3K/Akt signaling pathway and downregulating caspase-3 expression
in osteoblasts. Sal also protected against SANFH in SANFH rat
models by decreasing the percentage of empty lacunae. The
inhibition of the mitochondrial apoptosis pathway was involved.
Further research is necessary to decipher the detailed mechanism of
Sal in protecting against Dex-induced SANFH.
Acknowledgements
The authors thank the staff of the Laboratory of the
Orthopaedic Research Institute and the Scientific Research Center
of the Second Affiliated Hospital of Wenzhou Medical University.
The present study was generously supported by grants from the
Zhejiang Province Public Welfare Technology Application Research
Project, China (2015C33209) and Wenzhou Public Welfare Science and
Technology Project, Zhejiang Province, China (Y20150243).
References
|
1
|
Simon JP, Berger P and Bellemans J: Total
hip arthroplasty in patients less than 40 years old with avascular
necrosis of the femoral head. A 5 to 19-year follow-up study. Acta
Orthop Belg. 77:53–60. 2011.PubMed/NCBI
|
|
2
|
Motomura G, Yamamoto T, Yamaguchi R,
Ikemura S, Nakashima Y, Mawatari T and Iwamoto Y: Morphological
analysis of collapsed regions in osteonecrosis of the femoral head.
J Bone Joint Surg Br. 93:184–187. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tanaka Y, Yoshikawa N, Hattori S, Sasaki
S, Ando T, Ikeda M and Honda M; Japanese Study Group for Renal
Disease in Children, : Combination therapy with steroids and
mizoribine in juvenile SLE: A randomized controlled trial. Pediatr
Nephrol. 25:877–882. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hodson EM and Alexander SI: Evaluation and
management of steroid-sensitive nephrotic syndrome. Curr Opin
Pediatr. 20:145–150. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tönshoff B, Höcker B and Weber LT: Steroid
withdrawal in pediatric and adult renal transplant recipients.
Pediatr Nephrol. 20:409–417. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schäcke H, Döcke WD and Asadullah K:
Mechanisms involved in the side effects of glucocorticoids.
Pharmacol Ther. 96:23–43. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nagasawa K, Tada Y, Koarada S, Tsukamoto
H, Horiuchi T, Yoshizawa S, Murai K, Ueda A, Haruta Y and Ohta A:
Prevention of steroid-induced osteonecrosis of femoral head in
systemic lupus erythematosus by anti-coagulant. Lupus. 15:354–357.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Weinstein RS, Jilka RL, Parfitt AM and
Manolagas SC: Inhibition of osteoblastogenesis and promotion of
apoptosis of osteoblasts and osteocytes by glucocorticoids.
Potential mechanisms of their deleterious effects on bone. J Clin
Invest. 102:274–282. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim HJ: New understanding of
glucocorticoid action in bone cells. BMB Rep. 43:524–529. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Souttou B, Raulais D and Vigny M:
Pleiotrophin induces angiogenesis: Involvement of the
phosphoinositide-3 kinase but not the nitric oxide synthase
pathways. J Cell Physiol. 187:59–64. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Himburg HA, Muramoto GG, Daher P, Meadows
SK, Russell JL, Doan P, Chi JT, Salter AB, Lento WE, Reya T, et al:
Pleiotrophin regulates the expansion and regeneration of
hematopoietic stem cells. Nat Med. 16:475–482. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Feng Z, Zheng W, Tang Q, Cheng L, Li H, Ni
W and Pan X: Fludarabine inhibits STAT1-mediated up-regulation of
caspase-3 expression in dexamethasone-induced osteoblasts apoptosis
and slows the progression of steroid-induced avascular necrosis of
the femoral head in rats. Apoptosis. 22:1001–1012. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen X, Zhang Q, Cheng Q and Ding F:
Protective effect of salidroside against
H2O2-induced cell apoptosis in primary
culture of rat hippocampal neurons. Mol Cell Biochem. 332:85–93.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang SC, Lee FT, Kuo TY, Yang JH and
Chien CT: Attenuation of long-term Rhodiola rosea supplementation
on exhaustive swimming-evoked oxidative stress in the rat. Chin J
Physiol. 52:316–324. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ma L, Cai D, Li H, Tong B, Song L and Wang
Y: Anti-fatigue effects of salidroside in mice. J Med Coll PLA.
23:88–93. 2008. View Article : Google Scholar
|
|
16
|
Van Diermen D, Marston A, Bravo J, Reist
M, Carrupt PA and Hostettmann K: Monoamine oxidase inhibition by
Rhodiola rosea L. roots. J Ethnopharmacol. 122:397–401. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang L, Yu H, Sun Y, Lin X, Chen B, Tan
C, Cao G and Wang Z: Protective effects of salidroside on hydrogen
peroxide-induced apoptosis in SH-SY5Y human neuroblastoma cells.
Eur J Pharmacol. 564:18–25. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen X, Liu J, Gu X and Ding F:
Salidroside attenuates glutamate-induced apoptotic cell death in
primary cultured hippocampal neurons of rats. Brain Res.
1238:189–198. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lu Z, Jiang G, Chen Y, Wang J, Muhammad I,
Zhang L, Wang R, Liu F, Li R, Qian F and Li J: Salidroside
attenuates colistin-induced neurotoxicity in RSC96 Schwann cells
through PI3K/Akt pathway. Chem Biol Interact. 271:67–78. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Aziz SA, Davies M, Pick E, Zito C,
Jilaveanu L, Camp RL, Rimm DL, Kluger Y and Kluger HM:
Phosphatidylinositol-3-kinase as a therapeutic target in melanoma.
Clin Cancer Res. 15:3029–3036. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cantrell DA: Phosphoinositide 3-kinase
signaling pathways. J Cell Sci. 114:1439–1445. 2001.PubMed/NCBI
|
|
22
|
Gu YX, Du J, Si MS, Mo JJ, Qiao SC and Lai
HC: The roles of PI3K/Akt signaling pathway in regulating MC3T3-E1
preosteoblast proliferation and differentiation on SLA and SLActive
titanium surfaces. J Biomed Mater Res A. 101:748–754. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guntur AR and Rosen CJ: The skeleton: A
multi-functional complex organ: New insights into osteoblasts and
their role in bone formation: The central role of PI3Kinase. J
Endocrinol. 211:123–130. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li L, Xia Y, Wang Z, Cao X, Da Z, Guo G,
Qian J, Liu X, Fan Y, Sun L, et al: Suppression of the PI3K-Akt
pathway is involved in the decreased adhesion and migration of bone
marrow-derived mesenchymal stem cells from nonobese diabetic mice.
Cell Biol Int. 35:961–966. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang J, Ma XY, Feng YF, Ma ZS, Ma TC,
Zhang Y, Li X, Wang L and Lei W: Magnesium ions promote the
biological behaviour of rat calvarial osteoblasts by activating the
PI3K/Akt signaling pathway. Biol Trace Elem Res. 179:284–293. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Abdi A, Sadraie H, Dargahi L, Khalaj L and
Ahmadiani A: Apoptosis inhibition can be threatening in Aβ-induced
neuroinflammation, through promoting cell proliferation. Neurochem
Res. 36:39–48. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yamamoto T, Irisa T, Sugioka Y and Sueishi
K: Effects of pulse methylprednisolone on bone and marrow tissues:
Corticosteroid-induced osteonecrosis in rabbits. Arthritis Rheum.
40:2055–2064. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu X, Wen H, Hu Y, Yu H, Zhang Y, Chen C
and Pan X: STAT1-caspase 3 pathway in the apoptotic process
associated with steroid-induced necrosis of the femoral head. J Mol
Histol. 45:473–485. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang W, He H, Song H, Zhao J, Li T, Wu L,
Zhang X and Chen J: Neuroprotective effects of salidroside in the
MPTP mouse model of Parkinson's disease: Involvement of the
PI3K/Akt/GSK3β pathway. Parkinsons Dis. 2016:94501372016.PubMed/NCBI
|
|
30
|
Dai WW, Wang LB, Jin GQ, Wu HJ, Zhang J,
Wang CL, Wei YJ, Lee JH, Lay YE and Yao W: Beta-ecdysone protects
mouse osteoblasts from glucocorticoid-induced apoptosis in vitro.
Planta Med. 83:888–894. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chua CC, Chua BH, Chen Z, Landy C and
Hamdy RC: Dexamethasone induces caspase activation in murine
osteoblastic MC3T3-E1 cells. Biochim Biophys Acta. 1642:79–85.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ryu JS, Ko JH, Kim MK, Wee WR and Oh JY:
Prednisolone induces apoptosis in corneal epithelial cells through
the intrinsic pathway. Sci Rep. 7:41352017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Calder JD, Buttery L, Revell PA, Pearse M
and Polak JM: Apoptosis-a significant cause of bone cell death in
osteonecrosis of the femoral head. J Bone Joint Surg Br.
86:1209–1213. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Manning BD and Cantley LC: AKT/PKB
signaling: Navigating downstream. Cell. 129:1261–1274. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pap M and Cooper GM: Role of glycogen
synthase kinase-3 in the phosphatidylinositol 3-kinase/Akt cell
survival pathway. J Biol Chem. 273:19929–19932. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kang JQ, Chong ZZ and Maiese K: Critical
role for Akt1 in the modulation of apoptotic phosphatidylserine
exposure and microglial activation. Mol Pharmacol. 64:557–569.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhu Y, Shi YP, Wu D, Ji YJ, Wang X, Chen
HL, Wu SS, Huang DJ and Jiang W: Salidroside protects against
hydrogen peroxide-induced injury in cardiac H9c2 cells via PI3K-Akt
dependent pathway. DNA Cell Biol. 30:809–819. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang L, Ding W, Sun H, Zhou Q, Huang J,
Li X, Xie Y and Chen J: Salidroside protects PC12 cell from
MPP+-induced apoptosis via activation of the PI3K/Akt
pathway. Food Chem Toxicol. 50:2591–2597. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li DW, Liu ZQ, Chen W, Yao M and Li GR:
Association of glycogen synthase kinase-3β with Parkinson's disease
(review). Mol Med Rep. 9:2043–2050. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chao DT and Korsmeyer SJ: BCL-2 family:
Regulators of cell death. Annu Rev Immunol. 16:395–419. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hu WP, Yu HS, Sung PJ, Tsai FY, Shen YK,
Chang LS and Wang JJ: DC-81-Indole conjugate agent induces
mitochondria mediated apoptosis in human melanoma A375 cells. Chem
Res Toxicol. 20:905–912. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Steffen RT, Athanasou NA, Gill HS and
Murray DW: Avascular necrosis associated with fracture of the
femoral neck after hip resurfacing: Histological assessment of
femoral bone from retrieval specimens. J Bone Joint Surg Br.
92:787–793. 2010. View Article : Google Scholar : PubMed/NCBI
|