Effects of intrathecal bupivacaine on the NR2B/CaMKIIα/CREB signaling pathway in the rat lumbar spinal cord
- Authors:
- Published online on: January 17, 2018 https://doi.org/10.3892/mmr.2018.8448
- Pages: 4508-4514
-
Copyright: © Zhao et al. This is an open access article distributed under the terms of Creative Commons Attribution License.
Abstract
Introduction
Combined neuraxial and general anesthesia markedly reduces the sedative/anesthetic requirements of general anesthetics (1–3). Similar sensorial levels with neuraxial anesthesia reduce the need for propofol in similar amounts (4), and the high spinal blocking is associated with increased sedation (5). Previous studies have indicated that the sensorial levels of epidural and spinal anesthesia serve a key role in de-afferentation-dependent sedation. These findings have been verified by further studies, demonstrating that neuraxial anesthesia affects the degree of subcortical arousal via peripheral de-afferentation (6,7). This may occur principally through the ablation of ascending nociceptive information transmission, which reduces arousal from surgical stimuli. A second subtle mechanism of neuraxial anesthesia may involve the blockade of motor and sensory activities, leading to reduced arousal even in the absence of any noxious stimuli (8–10).
Local anesthetics (LAs) including bupivacaine may not only block sodium channels, however may also affect synaptic transmission, modulate neurotransmitter release and interact with numerous other membrane proteins (11–13). These actions of LAs suggest that they may exert effects on glutamatergic synaptic transmission (11). Although sensory de-afferentation in the spinal cord may explain the sedative effect of neuraxial anesthesia, the neurochemical basis of de-afferentation-dependent sedation remains unknown. There is a lack of data on the neurochemical alterations that occur in the spinal cord following subarachnoid blockade with bupivacaine, particularly regarding synaptic transmission, which may be important for de-afferentation-dependent sedation during spinal anesthesia (14). In the transmission of somatosensory activity from the periphery to the brain, the N-methyl-D-aspartate (NMDA) subtype of glutamate receptors is considered to be essential (15,16).
In the present study, subarachnoid bupivacaine blockade was hypothesized to produce a sensory de-afferentation by downregulating the N-methyl-D-aspartate receptor 2B subunit/calcium-calmodulin-dependent protein kinase II α/cAMP response element-binding protein (NR2B/CaMKIIα/CREB) excitatory signaling pathway in the spinal cord. To test this hypothesis, the effect of intrathecal bupivacaine at clinically relevant concentrations was investigated on the expression levels of NR2B, CaMKIIα/p-CaMKIIα, and CREB/p-CREB in rat lumbar spinal cord samples. The NR2B subunit of the NMDA receptor was investigated as it is considered to be specifically required for nociceptive afferents (16).
Materials and methods
Animals
The present study was approved by the Ethics Committee of Ningxia Medical University (Yinchuan, China). A total of 47 adult male Sprague-Dawley (SD) rats (weight, 280–320 g; age, 7–8 weeks) were obtained from the Animal Center of Ningxia Medical University. All of the procedures were performed in accordance with the National Institute of Health guidelines on animal care. Rats were housed in separate plastic cages with unlimited access to water and food, and kept in temperature-controlled rooms (20–24°C, relative humidity 50–60%) under a 12/12 h light/dark cycle (with the dark cycle beginning at 7:00 p.m.). All experiments were conducted during light hours.
Catheter modification and intrathecal catheter placement
Rats were allowed to acclimate to the laboratory environment for 3 days. A rat model of intrathecal catheterization was established as previously described (17). To construct the intrathecal catheter, sections of polyethylene tubing of sizes PE-20 (8 cm) and PE-10 (2 cm) were connected to each other to produce a catheter with a decreasing diameter profile. Animals were anesthetized with an intraperitoneal (i.p.) injection of pentobarbital sodium at a dose of 60 mg kg−1. A longitudinal 1.5-cm incision was made in the lumbar L4-L5 vertebra of each rat placed in a prone position. One end of the small profiled catheter was inserted into the subarachnoid space through the incision, while the other was tunneled subcutaneously toward the occiput, exposing the distal tip out of the neck skin. Cefazolin sodium (100 mg) was injected intramuscularly to prevent infection. Following 24 h of catheterization, all rats with any sign of neurological deficit, infection, catheter displacement, or clogging, and those without bilateral lower extremity paralysis following intrathecal injection of 15 µl 2% lidocaine (Yinhu Shiyao Pharmaceutical Co, Ltd., Yuncheng, Shanxi, China) within 5 min, were excluded from the study.
Treatment groups
Following catheterization, 36 male SD rats were included and assigned to a normal saline (NS) group (n=18) or a bupivacaine (Bup) group (n=18). Each rat was intrathecally administered once with 20 µl saline solution or 0.5% bupivacaine (Shanghai Zhaohui Pharmaceutical Co, Ltd., Shanghai, China) and injected for 10 sec. From each group, samples from six rats were used for western blotting, samples from another six rats were used for reverse transcription-quantitative polymerase chain reaction (RT-qPCR), and samples from the remaining six rats were used for immunohistochemical (IHC) analysis.
Western blot analysis
Following 10 min of intrathecal administration of bupivacaine or saline, the rats were sacrificed by spinal dislocation. The lumbar enlargement of the spinalrd, which was estimated to be in the L1-L5 region (18), was collected and homogenized in lysis buffer (Nanjing KeyGEN Biotech, Co., Ltd., Nanjing, China). The homogenate was centrifuged at 4°C and 14,000 × g for 15 min. The supernatant (50 µg per sample, as quantified by a bicinchoninic acid protein assay kit (Nanjing KeyGEN Biotech, Co., Ltd.) was separated on 8% SDS-PAGE and transferred onto nitrocellulose membranes (GE Healthcare Bio-Sciences, Pittsburgh, PA, USA). Following incubation in 5% non-fat milk for 2 h at room temperature, membranes were incubated with primary antibodies against GAPDH (10494–1-AP; 1:2,000; ProteinTech Group, Inc., Chicago, IL, USA), NR2B (21920–1-AP; 1:1,000; ProteinTech Group, Inc.), CaMKIIα (13730–1-AP; 1:1,000; ProteinTech Group, Inc.), CREB (9197; 1:900; Cell Signaling Technology, Inc., Danvers, MA, USA), phosphorylated (p)-CaMKIIα (11278; 1:700; Signal-Aldrich; Merck KGaA, Darmstadt, Germany), and p-CREB (9198; 1:600; Cell Signaling Technology, Inc., USA) at 4°C overnight. Following washing, the membranes were incubated at room temperature for 2 h with secondary antibody (goat anti-rabbit IgG, SA00001-2; 1:2,000; ProteinTech Group, Inc.). Following washing with TBS with Tween-20, blots were visualized with an enhanced chemiluminescence kit (32109; Pierce; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and subsequently analyzed densitometrically with a western blotting detection system (Quantity One software version 4.6.2; Bio-Rad Laboratories, Inc., Hercules, CA, USA). Results were normalized to the protein level of GAPDH in each sample.
Quantification of the spinal cord mRNA expression levels
RT-qPCR analysis was conducted following a previously described method (19). Total RNA from the spinal cord (L1-L5) tissue was isolated with a Total RNA Miniprep Kit (Axygen A; Corning Incorporated, Corning, NY, USA) and reverse transcribed into cDNA (TransGen Biotech, China). Primer sequences used for PCR are presented in Table I. PCR was performed in a Light Cycler system (qTOWER 2.0; Analytic Jena AG, Jena, Germany), using SYBR Green I dye (TransGen Biotech, Beijing, China) to detect the mRNA expression levels of NR2B, CaMKIIα, CREB, and GAPDH. The cDNA products (3.5 µl), 10 µM forward primer (1 µl), 10 µM reverse primer (1 µl), and 2XTransStart Top Green qPCR SuperMix (12.5 µl) were mixed and added with double distilled water to obtain a total reaction volume of 25 µl. The reaction conditions were 50°C for 2 min, 95°C for 10 min, and 40 cycles of 30 sec at 94°C and 30 sec at 60°C. Data were analyzed by the 2−ΔΔCq method, using GAPDH as a reference gene (20).
IHC analysis
Under deep anesthesia with sodium pentobarbital at a dose of 60 mg kg−1, six male SD rats from each experimental group were transcardially perfused with 500 ml NS, followed by 500 ml 4% paraformaldehyde in PBS. The spinal cord was removed and post-fixed in 10% formalin solution for 12 h at 4°C. All samples were embedded in paraffin and transversely cut into 4-µm thick sections with a sliding microtome. Following deparaffinization and hydration, sections were blocked in 3% H2O2 for 10 min at room temperature, and were incubated with primary antibodies, including anti-NR2B (ab216621; 1:250; Abcam, Cambridge, UK), anti-p-CaMKIIα (ab5683; 1:300; Abcam), and anti-p-CREB (9198; 1:700; Cell Signaling Technology, Inc.), for 2 h at 37°C. Slides were washed with 0.1 M PBS 3 times for 2 min and subsequently incubated with a goat anti-rabbit secondary antibody (PV-9001; 1:500; ZSGB-BIO; OriGene Technologies, Inc., Rockville, MD, USA) for 30 min at 37°C, then stained with diaminobenzidine (DAB kit; ZSGB-BIO; OriGene Technologies, Inc.) and counterstained with hematoxylin for 30 sec at room temperature. Images were captured with a light microscope (Olympus Corporation, Tokyo, Japan). Quantitative image analysis of the relative optical density (OD) was performed using Image Pro Plus software version 6.0 (Media Cybernetics, Inc., Rockville, MD, USA).
Statistical analysis
All analyses were performed with SPSS software, version 17.0 (SPSS, Inc., Chicago, IL, USA). All parametric data were presented as the mean ± standard deviation. Comparisons between the two groups were performed with independent samples t-tests. P<0.05 was considered to indicate a statistically significant difference.
Results
Of the 47 SD rats intrathecally catheterized, 36 were included in the experiments. Of the rats excluded, five exhibited a neurological deficit and six exhibited failed catheterization.
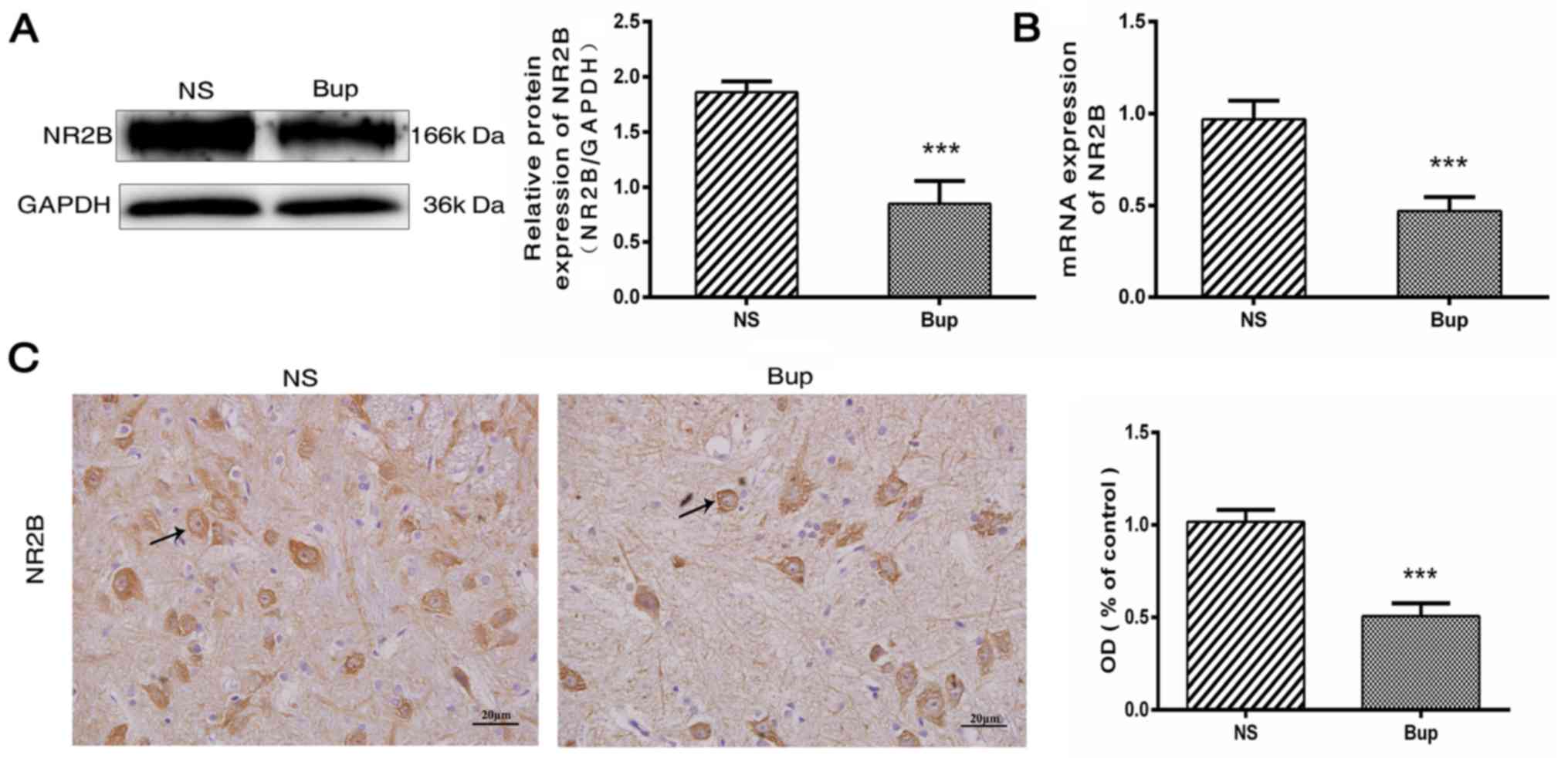
Intrathecal bupivacaine decreases NR2B expression in the spinal cord
Following the intrathecal administration of bupivacaine, protein and mRNA expression levels of NR2B were decreased by ~54 and 51%, respectively, compared with the NS rat group (Fig. 1A and B; P<0.001). This was further verified by IHC analysis, which demonstrated that the NR2B protein expression was decreased by ~50% in the spinal cord tissues following bupivacaine treatment (Fig. 1C; P<0.001). Together these observations indicated that the decreased expression of NR2B is associated with subarachnoid bupivacaine blockade.
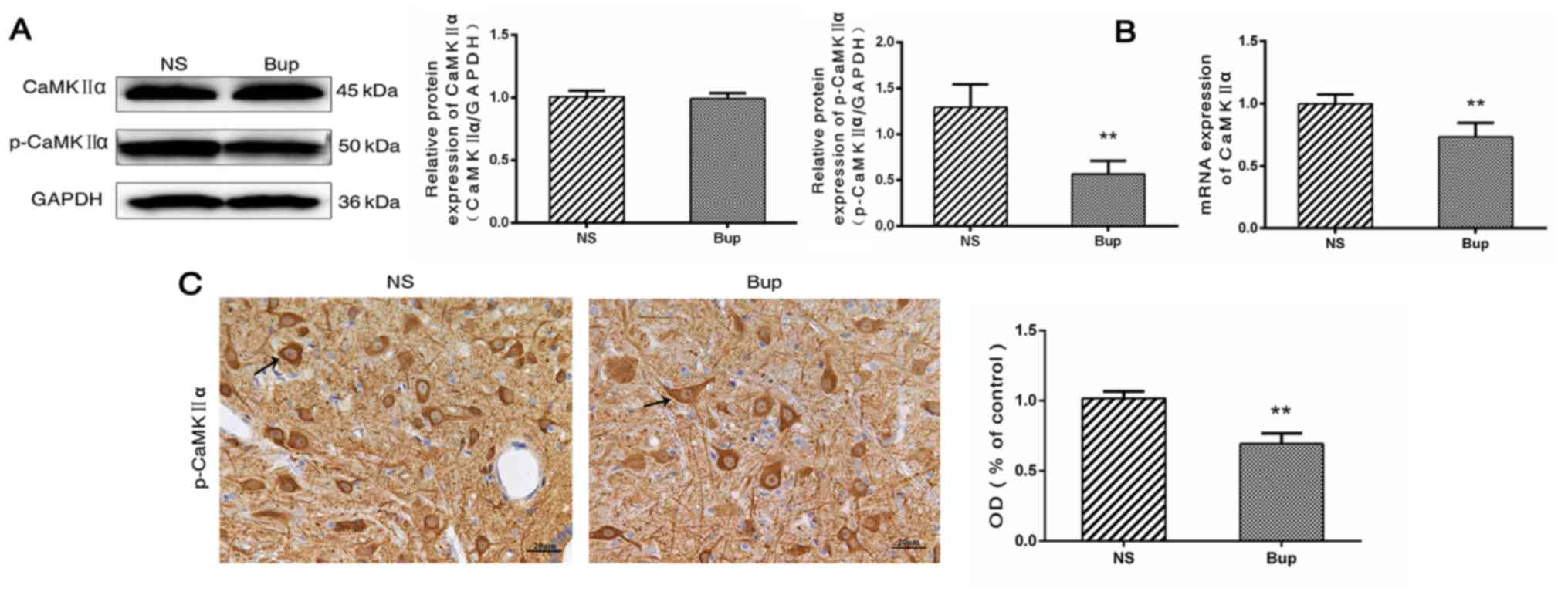
Effect of intrathecal bupivacaine on CaMKIIα and p-CaMKIIα levels in the spinal cord
Western blotting revealed that the protein expression level of total CaMKIIα in the spinal cord did not differ between the Bup and NS groups (Fig. 2A; P>0.05). Compared with the NS group, the mRNA level of CaMKIIα was reduced by ~36% in the Bup group (Fig. 2B; P<0.01). Western blotting and IHC indicated that the protein expression levels of p-CaMKIIα were decreased by ~56 and 32%, respectively in the Bup group compared with the NS group (Fig. 2A and C; P<0.01).
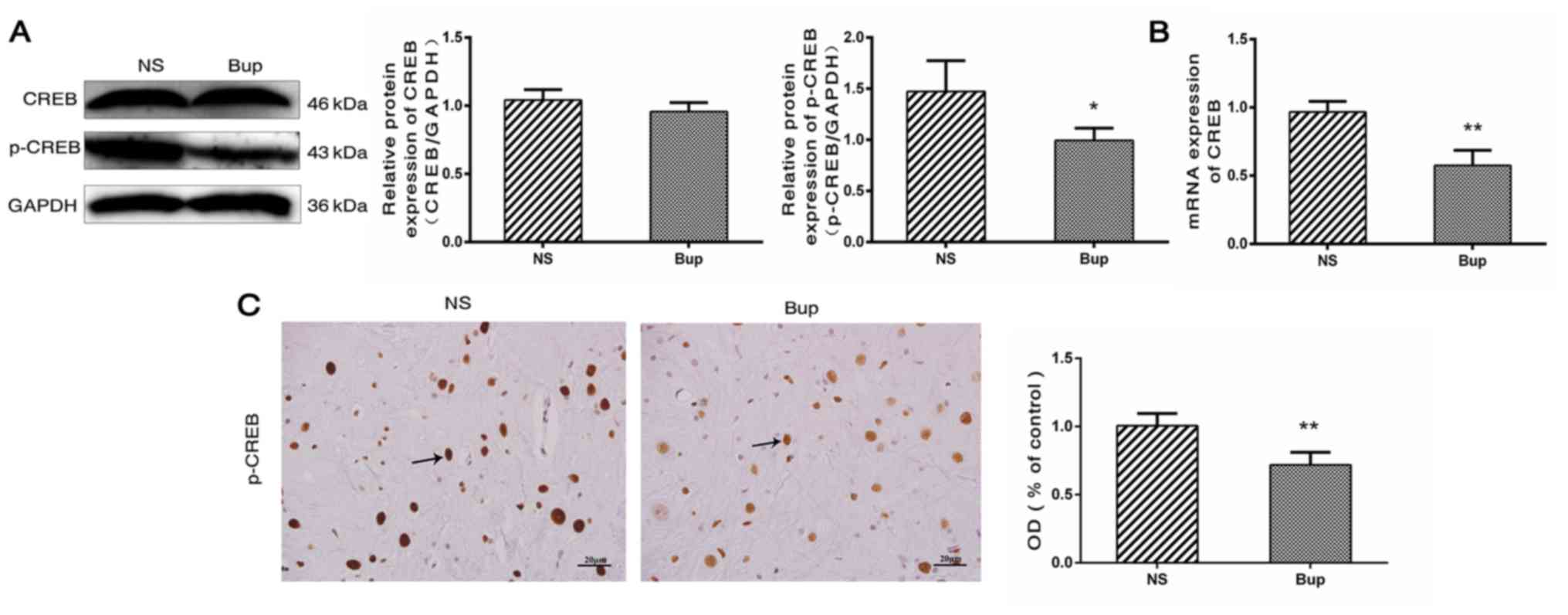
Effect of intrathecal bupivacaine on CREB and p-CREB levels in the spinal cord
Based on the results of western blot analysis, the alterations in the expression of CREB in the spinal cord were similar to those of CaMKIIα (Fig. 3A; P>0.05). Compared with the NS group, mRNA levels of CREB were reduced by ~41% in the Bup group (Fig. 3B; P<0.01). The protein expression level of p-CREB was indicated to be decreased by ~33 and 28% by western blotting and IHC, respectively in the Bup group, compared with the NS group (Fig. 3A and C; P<0.05 and P<0.01).
Discussion
Sedation during neuraxial blockade was highlighted in 1994 when Tverskoy et al (21) reported that subarachnoid blockade with bupivacaine reduces the hypnotic requirements of midazolam and thiopental. In animal (22) and clinical studies (23,24), these findings were verified, and neuraxial blocking was reported to elicit sedative effects. While the neurochemical mechanism underlying the reduced requirement for general anesthetic remains poorly understood at the spinal level, the systemic general anesthetic effects of absorbed local anesthetics (25) and de-afferentation (26) have been revealed to be mechanisms that may explain the interaction between local and general anesthetics. The afferentation theory suggests that tonic sensory and muscle-spindle activities maintain a state of wakefulness (26,27). For de-afferentation, neuraxial anesthesia reduces the spinal afferent input and thus affects the level of consciousness, enabling decreases in subsequent doses of the agent to achieve a defined level of sedation (27). The most speculated mechanism for sedation during neuraxial anesthesia is the de-afferentation phenomenon, which affects sensitivity to sedative/anesthetic drugs. Additionally, afferent somatosensory information, particularly nociceptive information, considerably affects brain activity (28,29). Complete spinal cord transection immediately slows down cortical electroencephalogram activity (30,31). Similarly, the degree of subcortical arousal is significantly affected when spinal and epidural blocking reduce or prevent afferent inputs from the blocked region of the body to the brain. However, the potential neurochemical mechanisms involved remain unknown.
The effect of subarachnoid bupivacaine blockade on the NMDA receptor subunit NR2B and its associated signal transduction pathway were investigated in the lumbar spinal cord of rats, using bupivacaine at clinically relevant concentrations. NR2B, p-CaMKIIα, and p-CREB were considerably downregulated following intrathecal administration of bupivacaine. Similar alterations in expression were observed by IHC analysis. Based on the current findings, it may be suggested that the sedative effect of subarachnoid bupivacaine blockade is associated with a decrease in spinal afferent input, possibly due to the downregulation of NR2B, p-CaMKIIα, and p-CREB expression levels in the lumbar spinal cord.
Glutamate acts as an excitatory neurotransmitter in primary afferent terminals and serves a critical role in spinal synaptic transmission through its activation of glutamate receptors (32). NMDA receptors are glutamate receptors in the central nervous system. The binding of glutamate to the NMDA receptor results in an influx of extracellular Ca2+, which controls membrane excitability and synaptic transmission (33). CaMKIIα is a multifunctional serine/threonine protein kinase, which is activated when cytosolic Ca2+ increases and in the presence of peripheral pain stimuli, CaMKIIα is phosphorylated at Thr286 (p-CaMKIIα) in the spinal cord (34). The p-CaMKIIα then translocates to the nucleus where it phosphorylates CREB at Ser133 in the dorsal horn of the spinal cord. Subsequently, p-CREB initiates gene transcription and translation and is thus possibly involved in the ascending transmission of nociceptive information. Peripheral nociceptive information, including visceral, inflammatory, and neuropathic pain, may activate NR2B, p-CaMKIIα, and p-CREB and may facilitate nociceptive transmission in the spinal cord. Therefore, reduced expression of NR2B, p-CaMKIIα, and p-CREB may suppress various types of hyperalgesia, including inflammatory pain (35,36), neuropathic pain (36–39), opioid-induced hyperalgesia (40), visceral pain (41–43), and central pain (44–46). A key step in the transmission of nociceptive information from the spinal cord to the brain is the activation of NMDA receptors in the spinal dorsal horn neurons (47). This suggests that ascending nociceptive transmission may be reduced by downregulating NMDA receptor expression and the associated signal transduction pathways. Furthermore, bupivacaine inhibits NMDA-induced glutamatergic transmission in rat dorsal horn neurons (48,49). This result is concordant with the present results. In the present study, spinally administered bupivacaine was indicated to inhibit NMDA-induced glutamatergic transmission in the spinal cord through downregulation of NR2B, p-CaMKIIα, and p-CREB.
Several important limitations were noted in the current study. Firstly, although protein and mRNA expression levels of NR2B, p-CaMKIIα, and p-CREB were all downregulated following intrathecal bupivacaine, the results indirectly indicate that the NR2B/CaMKIIα/CREB signaling pathway is involved in sedation during spinal anesthesia. Therefore, future studies are necessary in order to verify the association between sedation during spinal anesthesia and NR2B/CaMKIIα/CREB signaling by using agonists or antagonists. Secondly, the results demonstrated that expression levels of total CaMKIIα, and CREB mRNA were decreased, although the levels of their protein expression were not altered following intrathecal bupivacaine. The association between mRNA and protein expression is not strictly linear, however has a more intrinsic and complex dependence (50). Therefore, this variation may be due to post-transcriptional and post-translation regulation and experimental errors (50,51). Thirdly, the upstream and downstream molecules of NR2B/CaMKIIa/CREB were not detected. Future studies may elucidate the function of these molecular components.
In conclusion, sedation during spinal anesthesia from decreased spinal afferent input may be associated with the downregulation of NR2B, p-CaMKIIα, and p-CREB expression levels in the spinal cord. Therefore, stimulatory input to the brain may be indirectly reduced by inhibiting the NR2B/CaMKIIα/CREB signaling pathway in spinal neurons, with this inhibition rendering the brain susceptible to the effects of sedative drugs. The present findings may prompt anesthesiologists to plan appropriate dosage guidelines for the prevention of anesthetic overdose and investigate the mechanism of sedation during neuraxial anesthesia.
Acknowledgements
The present study was supported by the National Natural Science Foundation of China (grant no. 81660198) and Ningxia Natural Science Foundation (grant. no. NZ15137).
References
|
Kim SH, Chun DH, Chang CH, Kim TW, Kim YM and Shin YS: Effect of caudal block on sevoflurane requirement for lower limb surgery in children with cerebral palsy. Paediatr Anaesth. 21:394–398. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Xiang Y, Chen CQ, Chen HJ, Li M, Bao FP and Zhu SM: The effect of epidural lidocaine administration on sedation of propofol general anesthesia: A randomized trial. J Clin Anesth. 26:523–529. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Zhang J, Zhang W and Li B: The effect of epidural anesthesia with different concentrations of ropivacaine on sevoflurane requirements. Anesth Analg. 104:984–986. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Sentürk M, Gücyetmez B, Ozkan-Seyhan T, Karadeniz M, Dinçer S, Akpir D, Sengül T and Denkel T: Comparison of the effects of thoracic and lumbar epidural anaesthesia on induction and maintenance doses of propofol during total i.v. anaesthesia. Br J Anaesth. 101:255–260. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Gentili M, Huu PC, Enel D, Hollande J and Bonnet F: Sedation depends on the level of sensory block induced by spinal anaesthesia. Br J Anaesth. 81:970–971. 1998. View Article : Google Scholar : PubMed/NCBI | |
|
Pollock JE, Neal JM, Liu SS, Burkhead D and Polissar N: Sedation during spinal anesthesia. Anesthesiology. 93:728–734. 2000. View Article : Google Scholar : PubMed/NCBI | |
|
Yang W, Geng Y, Liu Y, Li A, Liu J, Xing J and Li W: Comparison of effects of thoracic epidural and intravenous administration of lidocaine on target-controlled infusion of propofol and tracheal intubation response during induction of anesthesia. J Cardiothorac Vasc Anesth. 27:1295–1300. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Hodgson PS and Liu SS: Epidural lidocaine decreases sevoflurane requirement for adequate depth of anesthesia as measured by the Bispectral Index monitor. Anesthesiology. 94:799–803. 2001. View Article : Google Scholar : PubMed/NCBI | |
|
Doufas AG, Wadhwa A, Shah YM, Lin CM, Haugh GS and Sessler DI: Block-dependent sedation during epidural anaesthesia is associated with delayed brainstem conduction. Br J Anaesth. 93:228–234. 2004. View Article : Google Scholar : PubMed/NCBI | |
|
Antognini JF, Atherley R and Carstens E: Isoflurane action in spinal cord indirectly depresses cortical activity associated with electrical stimulation of the reticular formation. Anesth Analg. 96:999–1003. 2003.PubMed/NCBI | |
|
Lin TY, Chung CY, Lu CW, Huang SK, Shieh JS and Wang SJ: Local anesthetics inhibit glutamate release from rat cerebral cortex synaptosomes. Synapse. 67:568–579. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Cherng CH, Wong CS, Wu CT and Yeh CC: Glutamate release and neurologic impairment after intrathecal administration of lidocaine and bupivacaine in the rat. Reg Anesth Pain Med. 36:452–456. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Nishizawa N, Shirasaki T, Nakao S, Matsuda H and Shingu K: The inhibition of the N-methyl-D-aspartate receptor channel by local anesthetics in mouse CA1 pyramidal neurons. Anesth Analg. 94:325–330. 2002. View Article : Google Scholar : PubMed/NCBI | |
|
Lanier WL, Iaizzo PA, Milde JH and Sharbrough FW: The cerebral and systemic effects of movement in response to a noxious stimulus in lightly anesthetized dogs. Possible modulation of cerebral function by muscle afferents. Anesthesiology. 80:392–401. 1994. View Article : Google Scholar : PubMed/NCBI | |
|
Zhai QZ and Traub RJ: The NMDA receptor antagonist MK-801 attenuates c-Fos expression in the lumbosacral spinal cord following repetitive noxious and non-noxious colorectal distention. Pain. 83:321–329. 1999. View Article : Google Scholar : PubMed/NCBI | |
|
Tong CK and MacDermott AB: Synaptic GluN2A and GluN2B containing NMDA receptors within the superficial dorsal horn activated following primary afferent stimulation. J Neurosci. 34:10808–10820. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Saito Y, Kaneko M, Kirihara Y, Sakura S and Kosaka Y: Interaction of intrathecally infused morphine and lidocaine in rats (part I): Synergistic antinociceptive effects. Anesthesiology. 89:1455–1463. 1998. View Article : Google Scholar : PubMed/NCBI | |
|
Oklinski MK, Lim JS, Choi HJ, Oklinska P, Skowronski MT and Kwon TH: Immunolocalization of water channel proteins AQP1 and AQP4 in rat spinal cord. J Histochem Cytochem. 62:598–611. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Wang HL, Li YX, Niu YT, Zheng J, Wu J, Shi GJ, Ma L, Niu Y, Sun T and Yu JQ: Observing anti-inflammatory and anti-nociceptive activities of glycyrrhizin through regulating COX-2 and pro-inflammatory cytokines expressions in mice. Inflammation. 38:2269–2278. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Livak KJ and Schmittgen TD: Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI | |
|
Tverskoy M, Shagal M, Finger J and Kissin I: Subarachnoid bupivacaine blockade decreases midazolam and thiopental hypnotic requirements. J Clin Anesth. 6:487–490. 1994. View Article : Google Scholar : PubMed/NCBI | |
|
Eappen S and Kissin I: Effect of subarachnoid bupivacaine block on anesthetic requirements for thiopental in rats. Anesthesiology. 88:1036–1042. 1998. View Article : Google Scholar : PubMed/NCBI | |
|
Agarwal A, Pandey R, Dhiraaj S, Singh PK, Raza M, Pandey CK, Gupta D, Choudhury A and Singh U: The effect of epidural bupivacaine on induction and maintenance doses of propofol (evaluated by bispectral index) and maintenance doses of fentanyl and vecuronium. Anesth Analg. 99:1684–1688. 2004. View Article : Google Scholar : PubMed/NCBI | |
|
Ingelmo PM, Ferri F and Fumagalli R: Interactions between general and regional anesthesia. Minerva Anestesiol. 72:437–445. 2006.PubMed/NCBI | |
|
Inagaki Y, Mashimo T, Kuzukawa A, Tsuda Y and Yoshiya I: Epidural lidocaine delays arousal from isoflurane anesthesia. Anesth Analg. 79:368–372. 1994. View Article : Google Scholar : PubMed/NCBI | |
|
Foffani G, Humanes-Valera D, Calderon-Muñoz F, Oliviero A and Aguilar J: Spinal cord injury immediately decreases anesthetic requirements in rats. Spinal Cord. 49:822–826. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Hodgson PS, Liu SS and Gras TW: Does epidural anesthesia have general anesthetic effects? A prospective, randomized, double-blind, placebo-controlled trial. Anesthesiology. 91:1687–1692. 1999. View Article : Google Scholar : PubMed/NCBI | |
|
Antognini JF and Carstens E: Isoflurane blunts electroencephalographic and thalamic-reticular formation responses to noxious stimulation in goats. Anesthesiology. 91:1770–1779. 1999. View Article : Google Scholar : PubMed/NCBI | |
|
Antognini JF, Carstens E, Sudo M and Sudo S: Isoflurane depresses electroencephalographic and medial thalamic responses to noxious stimulation via an indirect spinal action. Anesth Analg. 91:1282–1288. 2000. View Article : Google Scholar : PubMed/NCBI | |
|
Aguilar J, Humanes-Valera D, Alonso-Calviño E, Yague JG, Moxon KA, Oliviero A and Foffani G: Spinal cord injury immediately changes the state of the brain. J Neurosci. 30:7528–7537. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Alonso-Calviño E, Martinez-Camero I, Fernández-López E, Humanes-Valera D, Foffani G and Aguilar J: Increased responses in the somatosensory thalamus immediately after spinal cord injury. Neurobiol Dis. 87:39–49. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Aanonsen LM and Wilcox GL: Nociceptive action of excitatory amino acids in the mouse: Effects of spinally administered opioids, phencyclidine and sigma agonists. J Pharmacol Exp Ther. 243:9–19. 1987.PubMed/NCBI | |
|
Tang Q, Bangaru ML, Kostic S, Pan B, Wu HE, Koopmeiners AS, Yu H, Fischer GJ, McCallum JB, Kwok WM, et al: Ca2+-dependent regulation of Ca2+ currents in rat primary afferent neurons: Role of CaMKII and the effect of injury. J Neurosci. 32:11737–11749. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Zeitz KP, Giese KP, Silva AJ and Basbaum AI: The contribution of autophosphorylated alpha-calcium-calmodulin kinase II to injury-induced persistent pain. Neuroscience. 128:889–898. 2004. View Article : Google Scholar : PubMed/NCBI | |
|
Nakanishi M, Hata K, Nagayama T, Sakurai T, Nishisho T, Wakabayashi H, Hiraga T, Ebisu S and Yoneda T: Acid activation of Trpv1 leads to an up-regulation of calcitonin gene-related peptide expression in dorsal root ganglion neurons via the CaMK-CREB cascade: A potential mechanism of inflammatory pain. Mol Biol Cell. 21:2568–2577. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Descalzi G, Fukushima H, Suzuki A, Kida S and Zhuo M: Genetic enhancement of neuropathic and inflammatory pain by forebrain upregulation of CREB-mediated transcription. Mol Pain. 8:902012. View Article : Google Scholar : PubMed/NCBI | |
|
Miletic G, Pankratz MT and Miletic V: Increases in the phosphorylation of cyclic AMP response element binding protein (CREB) and decreases in the content of calcineurin accompany thermal hyperalgesia following chronic constriction injury in rats. Pain. 99:493–500. 2002. View Article : Google Scholar : PubMed/NCBI | |
|
Ma W, Hatzis C and Eisenach JC: Intrathecal injection of cAMP response element binding protein (CREB) antisense oligonucleotide attenuates tactile allodynia caused by partial sciatic nerve ligation. Brain Res. 988:97–104. 2003. View Article : Google Scholar : PubMed/NCBI | |
|
Katano T, Nakazawa T, Nakatsuka T, Watanabe M, Yamamoto T and Ito S: Involvement of spinal phosphorylation cascade of Tyr1472-NR2B, Thr286-CaMKII, and Ser831-GluR1 in neuropathic pain. Neuropharmacology. 60:609–616. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Wang Z, Ma W, Chabot JG and Quirion R: Calcitonin gene-related peptide as a regulator of neuronal CaMKII-CREB, microglial p38-NFκB and astroglial ERK-Stat1/3 cascades mediating the development of tolerance to morphine-induced analgesia. Pain. 151:194–205. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Wang Y, Wu J, Lin Q, Nauta HJ, Yue Y and Fang L: Effects of general anesthetics on visceral pain transmission in the spinal cord. Mol Pain. 4:502008. View Article : Google Scholar : PubMed/NCBI | |
|
Pan X, Chen J, Wang W, Chen L, Wang L, Ma Q, Zhang J, Chen L, Wang G, Zhang M, et al: Resveratrol-induced antinociception is involved in calcium channels and calcium/caffeine-sensitive pools. Oncotarget. 8:9399–9409. 2017.PubMed/NCBI | |
|
Miranda A, Mickle A, Bruckert M, Kannampalli P, Banerjee B and Sengupta JN: NMDA receptor mediates chronic visceral pain induced by neonatal noxious somatic stimulation. Eur J Pharmacol. 744:28–35. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Crown ED, Gwak YS, Ye Z, Yu Tan H, Johnson KM, Xu GY, McAdoo DJ and Hulsebosch CE: Calcium/calmodulin dependent kinase II contributes to persistent central neuropathic pain following spinal cord injury. Pain. 153:710–721. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Fang L, Wu J, Zhang X, Lin Q and Willis WD: Calcium/calmodulin dependent protein kinase II regulates the phosphorylation of cyclic AMP-responsive element-binding protein of spinal cord in rats following noxious stimulation. Neurosci Lett. 374:1–4. 2005. View Article : Google Scholar : PubMed/NCBI | |
|
Mitsikostas DD, Knight YE, Lasalandra M, Kavantzas N and Goadsby PJ: Triptans attenuate capsaicin-induced CREB phosphorylation within the trigeminal nucleus caudalis: A mechanism to prevent central sensitization? J Headache Pain. 12:411–417. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Liu H, Zhang Y, Qi D and Li W: Downregulation of the spinal NMDA receptor NR2B subunit during electro-acupuncture relief of chronic visceral hyperalgesia. J Physiol Sci. 67:197–206. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Paganelli MA and Popescu GK: Actions of bupivacaine, a widely used local anesthetic, on NMDA receptor responses. J Neurosci. 35:831–842. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Furutani K, Ikoma M, Ishii H, Baba H and Kohno T: Bupivacaine inhibits glutamatergic transmission in spinal dorsal horn neurons. Anesthesiology. 112:138–143. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
de Sousa Abreu R, Penalva LO, Marcotte EM and Vogel C: Global signatures of protein and mRNA expression levels. Mol Biosyst. 5:1512–1526. 2009.PubMed/NCBI | |
|
Fraser HB, Hirsh AE, Giaever G, Kumm J and Eisen MB: Noise minimization in eukaryotic gene expression. PLoS Biol. 2:e1372004. View Article : Google Scholar : PubMed/NCBI |