Introduction
In acute liver failure, hepatocellular death
surpasses hepatocyte regeneration. Studies have revealed that
hepatocellular death may be caused by a variety of biochemical
signaling pathways, and the morphological alterations are
characterized by large hepatocytes, necrosis or severe degeneration
accompanied by a large amount of hepatocellular apoptosis, which
occurs in response to viruses, drugs, ethanol, toxins and
hemorrhagic lesions, followed by necrosis (1). A large amount of hepatocellular
apoptosis and necrosis defines acute liver failure (2,3).
However, the underlying molecular mechanism of apoptosis in acute
liver failure remains to be understood.
It is believed that the phosphoinositide-3 kinase
(PI3K)/protein kinase B (AKT) signaling pathway mediates cell
survival and a variety of biological effects, resulting in an
anti-apoptotic effect (4,5). Activated AKT may promote the
phosphorylation of downstream substrates such as glycogen synthase
kinase 3β (GSK3β) (12). Tournier
et al (6) argued that the
P38 and c-Jun N-terminal kinase (JNK) pathway is involved in
apoptosis via mitogen-activated protein kinase (MAPK) signal
transduction, and P38 also causes inflammatory responses. Activated
JNK promotes apoptosis, and is involved in the regulation of
apoptosis-associated gene expression and the activation of the
caspase family, making it more complex than P38.
Patrinia is commonly used for reducing heat,
detoxification, eliminating carbuncle abscess, reducing pain and
dissipating blood stasis. It is a spicy and bitter heat-clearing
herb, and may exhibit anti-viral, neuroprotective,
heart-protective, antioxidant and anti-tumor effects (7). As it has a variety of pharmacological
effects, Patrinia has attracted widespread attention. However, the
inhibition of hepatocellular apoptosis and its underlying mechanism
of liver failure have yet to be fully elucidated. The present study
aimed to investigate the inhibitory effect of Patrinia on
hepatocyte apoptosis in a rat model of acute liver failure, in
order to provide experimental evidence for the prevention and
treatment of acute liver failure with Patrinia. It was also
determined whether this inhibition was mediated through the
TLR4/PI3K/AKT/GSK3β and TLR4/P38/JNK signaling pathways.
Materials and methods
Materials
The rat liver BRL-3A cell line was provided by the
Sun Yat-sen University cell bank (Guangzhou, China). Patrinia was
purchased from Xi'an Four Seasons Biotechnology Co., Ltd. (Xi'an,
China).
Cell Counting Kit-8 (CCK-8) assay to
determine BRL-3A cell viability
To measure the effect of Patrinia on BRL-3A cell
viability, BRL-3A cells were seeded at a density of
1×104 cells/ml in 96 well plates for 48 h at 37°C in
RMPI-1640 (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), 10% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc.) and 1% streptomycin and penicillin. Cells were subsequently
divided into five groups: The control group (treated with
RPMI-1640), and Patrinia treated groups at concentrations of 0.25,
0.5, 1.0, 2.0 and 4.0 g/l. The wells of all groups were replaced
with fresh medium and cells were cultured for a further 24 h at
37°C. Cells were incubated with tetrazolium salt for 4 h and the
absorbance in each well was subsequently measured at 450 nm with a
microplate reader (Beijing Biocoen Biotechnology Co., Ltd.,
Beijing, China). The control group viability was set at 100% and
BRL-3A cell viability of the Patrinia-treated groups were
calculated accordingly.
To determine the effect of Patrinia on LPS-induced
BRL-3A cell viability, BRL-3A cells were seeded at a density of
1×104 cells/ml in 96-well plates for 48 h, and then
divided into five groups: A, the normal group; B, the LPS-treated
group; C, the low-density Patrinia (0.5 g/l)-treated group; D, the
middle-density Patrinia (1 g/l)-treated group; E, the high-density
Patrinia (2 g/l)-treated group. Cells were pretreated with Patrinia
for 2 h and then treated with LPS at 10 µg/ml for 24 h, prior to
determining the BRL-3A cell viability as described above.
Measuring the effect of Patrinia on
lactate dehydrogenase (LDH) release by LPS stimulation in BRL-3A
cells via a biochemical method
BRL-3A cells (1×104/ml) were seeded in
96-well culture plates for 24 h, and grouped and treated as
described above. LDH release in each BRL-3A cell was determined
using a lactate dehydrogenase (LDH) kit (Genmed Scientifics, Inc.,
Wilmington, DE, USA). Cells were incubated with LDH reagent for 30
min at room temperature and absorbance was measured at 490 nm with
a microplate reader.
Hoechst 33342 staining to observe the
effect of Patrinia on apoptosis of BRL-3A cells after LPS
stimulation
BRL-3A cell suspension was prepared and the cell
density was adjusted to 1×104 cells/ml. The cells were
cultured in 12-well culture plates with round coverslips. After
incubation for 24 h, the cells were fixed with 4% paraformaldehyde
for 20 min and subsequently incubated with Hoechst stain solution
(1:1,000; Dojindo Molecular Technologies, Inc., Kumamoto, Japan)
for 15 min at room temperature. Cells were observed with a
fluorescent microscope (IX51; Olympus Corporation, Tokyo, Japan) at
a magnification of ×100.
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) double labeling
Cells (1×104/ml) were cultured in 6-well
plates for 24 h. Cells were placed into a centrifuge tube, then
digested with 0.25% EDTA-free trypsin (Gibco, Thermo Fisher
Scientific, Inc.). Cells were collected and centrifuged at 447.2 ×
g for 5 min at 37°C and the supernatant was discarded before
washing the cells with PBS twice. The cell density was adjusted to
5×105 cells/ml.
A total of 1 ml cell suspension was centrifuged at
111.8 × g for 10 min at 4°C and the supernatant was discarded. A
total of 1 ml cold PBS was added and the cells were agitated for
efficient suspension. The cells were centrifuged again at 111.8 × g
for 10 min at 4°C and the supernatant was discarded. The above
procedure was repeated three or four times. The cells were
re-suspended in 200 µl binding buffer (cat. no. RVBB-02; Biomiga,
Shanghai, China). Following this, the apoptotic rate was detected
using an Annexin V-FITC apoptosis jut (Biouniquer Technology Co.,
Ltd., Hangzhou, China). A total of 10 µl Annexin V-FITC was added
and cells were agitated in the dark for 1 min at room temperature.
Following the addition of a further 300 µl binding buffer and 5 µl
PI, the apoptotic rate was measured by flow cytometry (BD FACS
Calibur™; BD Biosciences, Franklin Lakes, NJ, USA). The
experiment was repeated three times.
Western blot analysis
Cells were collected from all treatment groups and
washed with PBS twice. A total of 400 µl cell lysate was mixed with
40 µl 10 mmol/l phenylmethylsulfonyl fluoride, gently agitated and
put on ice for 10 min for sufficient lysis. The cells were gauged
with sterile syringes repeatedly. The lysates were placed into
Eppendorf tubes, placed on an ice-bath for 30 min and centrifuged
at 12,000 × g at 4°C for 15 min. The supernatant was subsequently
transferred to fresh Eppendorf tubes. Protein concentration was
determined using a BCA protein assay kit. Protein sample 6X buffer
(20 µl; Beijing Biocoen Biotechnology Co., Ltd.) was added to each
sample (100 µl), boiled for 5 min and then stored at 80°C. Protein
(30 µg/lane) was separated by 12% SDS-PAGE and transferred onto
polyvinylidene difluoride membranes. The primary antibodies against
toll-like receptor 4 (TLR4 (cat. no. 76B357; Novus Biologicals,
LLC, Littleton, CO, USA), AKT (cat. no. 9272S; CST Biological
Reagents Co., Ltd., Shanghai, China), phosphorylated
(P)-AKTSer473 (cat. no. 9271S; CST Biological Reagents
Co., Ltd.), GSK3β (cat. no. 9315S; CST Biological Reagents Co.,
Ltd.), P-GSK3βSer9 (cat. no. 9323S; CST Biological
Reagents Co., Ltd.), P38 (cat. no. ab32142; Abcam, Cambridge, UK),
P-P38 (ab47363, Abcam, USA), JNK (cat. no. ab110724; Abcam), P-JNK
(cat. no. ab76572; Abcam), B-cell lymphoma-2 (Bcl-2; cat. no.
S1460; Selleck Chemicals, Houston, TX, USA), Bcl-2 associated X
protein (Bax; cat. no. S1076; Selleck Chemicals) and
active-caspase-3 (cat. no. S7524; Selleck Chemicals) were added at
a 1:1,000 dilution and incubated overnight at 4°C. The cells were
washed with PBS with 0.05% Tween (PBST) three times. Horseradish
peroxidase-conjugated goat anti-rabbit IgG (cat. no. 01-15-06;
1:1,000; Sigma-Aldrich; Merck KGaA) was added and incubated for 1 h
at 4°C. The cells were washed with PBST three times. Proteins were
visualized using a chemiluminescence substrate (Merck KGaA). The
expression levels of the above proteins were measured using the
ChemiDox XRS+ system (version 4.6; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Effect of Patrinia on GSK3β
translocation in BRL-3A cells after LPS stimulation
A BRL-3 cell suspension was prepared and the cell
density was adjusted to 1×104 cells/ml. After incubation
for 24 h in a 12-well culture plate with coverslips, cells were
grouped and treated as described above. Following this, the
coverslips containing cells were removed and immunofluorescence
staining of GSK3β was performed. Cells were fixed in 4%
paraformaldehyde for 30 min at room temperature and blocked with 1%
bovine serum albumin (Santa Cruz Biotechnology, Inc., Dallas, TX,
USA) for 20 min at room temperature. Cells were then incubated with
GSK3β primary antibody (cat. no. 9315S; 1:500; CST Biological
Reagents Co., Ltd.) for 12 h at 4°C followed by incubation with
Alexa Fluor® 488 Donkey Anti-Rabbit IgG (H+L) secondary
antibody (cat. no. A121206; Invitrogen; Thermo Fisher Scientific,
Inc.) at 37°C for 1 h. GSK3β nuclear translocation was observed and
images were captured using a IX51 fluorescence microscope (Olympus
Corporation).
Statistical analysis
Data are expressed as mean ± standard deviation of
three independent experiments. Comparison between two groups was
performed via a Student's t-test, whereas comparisons between >2
groups was determined by a one-way analysis of variance followed by
Tukey's post-hoc test for multiple comparisons. P<0.05 was
considered to indicate a statistically significant difference. Data
were analyzed using GraphPad Prism version 5.0 software (GraphPad
Software, Inc., La Jolla, CA, USA).
Results
Effect of Patrinia on BRL-3A cell
viability
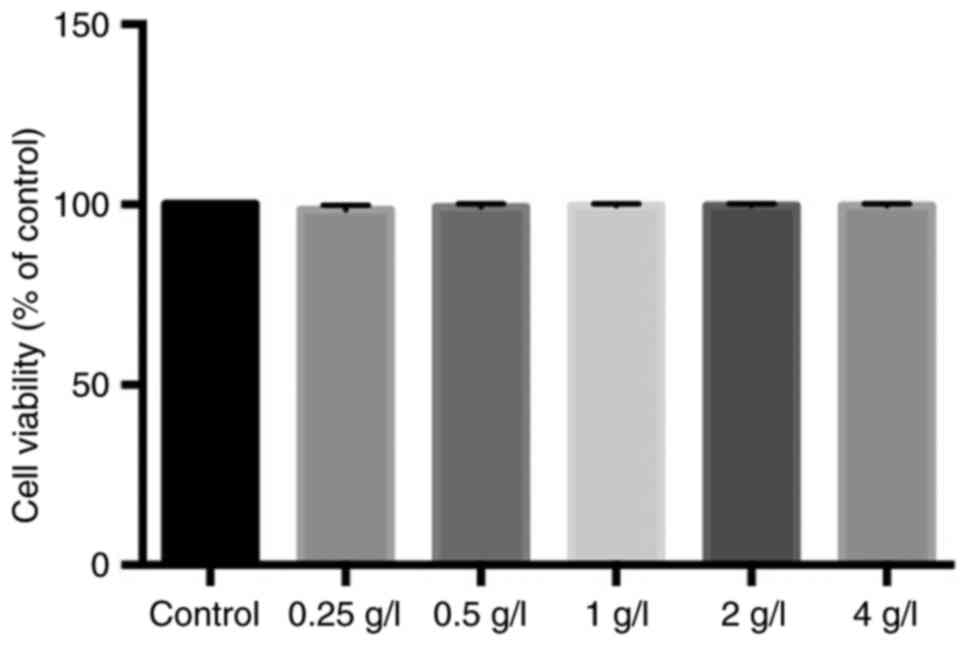
The CCK-8 assay was used to determine the effect of
Patrinia on the viability of BRL-3A cells. BRL-3A cells were
cultured in medium containing different concentrations of Patrinia
for 24 h. The results demonstrated that the cell viability was 100%
in the control group. Compared with the control group, the
Patrinia-treated groups did not show cytotoxicity, suggesting that
the subsequent inhibitory effect of Patrinia was not due to its
cytotoxicity (Fig. 1).
Effects of Patrinia on the viability
of BRL-3A cells after LPS stimulation
The CCK-8 assay was used to determine the effect of
Patrinia on the viability of BRL-3A cells after LPS stimulation.
The results revealed that the cell viability of the LPS-stimulated
group decreased significantly compared with the control group.
However, compared with the LPS-stimulated group, the cell viability
of the Patrinia-pretreated groups increased in a dose-dependent
manner (Fig. 2).
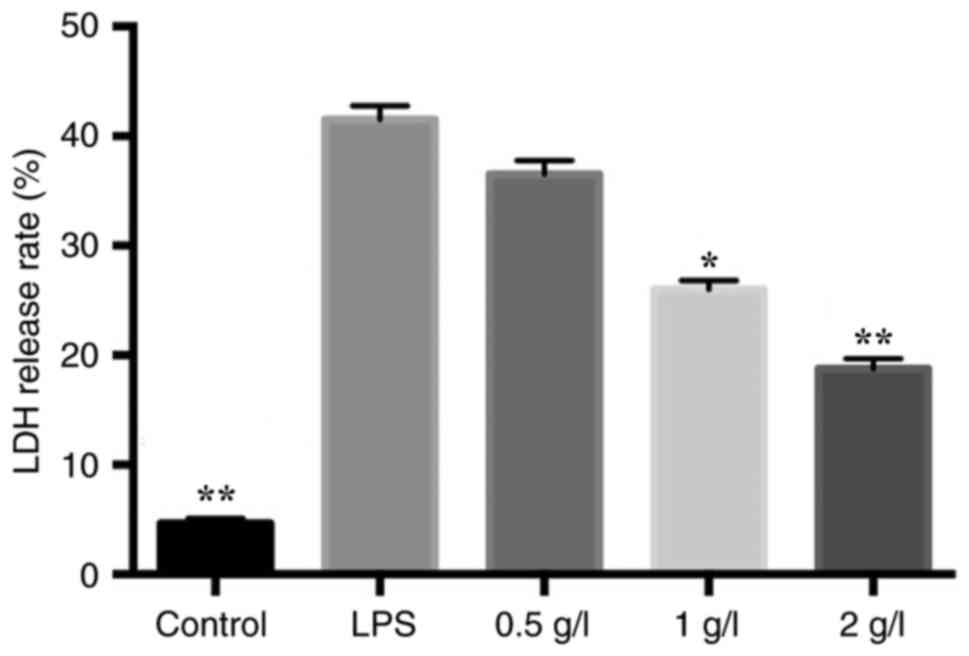
Effect of Patrinia on the release of
LDH in BRL-3A cells after LPS stimulation
The release of LDH was an indicator of toxicity of
BRL-3A cells after LPS stimulation. The results demonstrated that,
compared with the control group, LDH release was significantly
increased in the LPS-stimulated groups. However, pre-treatment with
Patrinia significantly reduced LDH release, in a dose-dependent
manner (Fig. 3).
Morphological effects of Patrinia on
BRL-3A cell apoptosis after LPS stimulation
Fluorescence microscopy was employed to observe the
morphological features of apoptosis. The control group exhibited
normal nucleus morphology and nuclear membrane integrity, with no
signs of apoptosis. The LPS group displayed enhanced staining of
apoptotic cells and nuclear debris and pyknosis were observed. The
fluorescence intensity of the Patrinia-treated groups appeared to
be lower, suggesting a reduced number of apoptotic cells (Fig. 4).
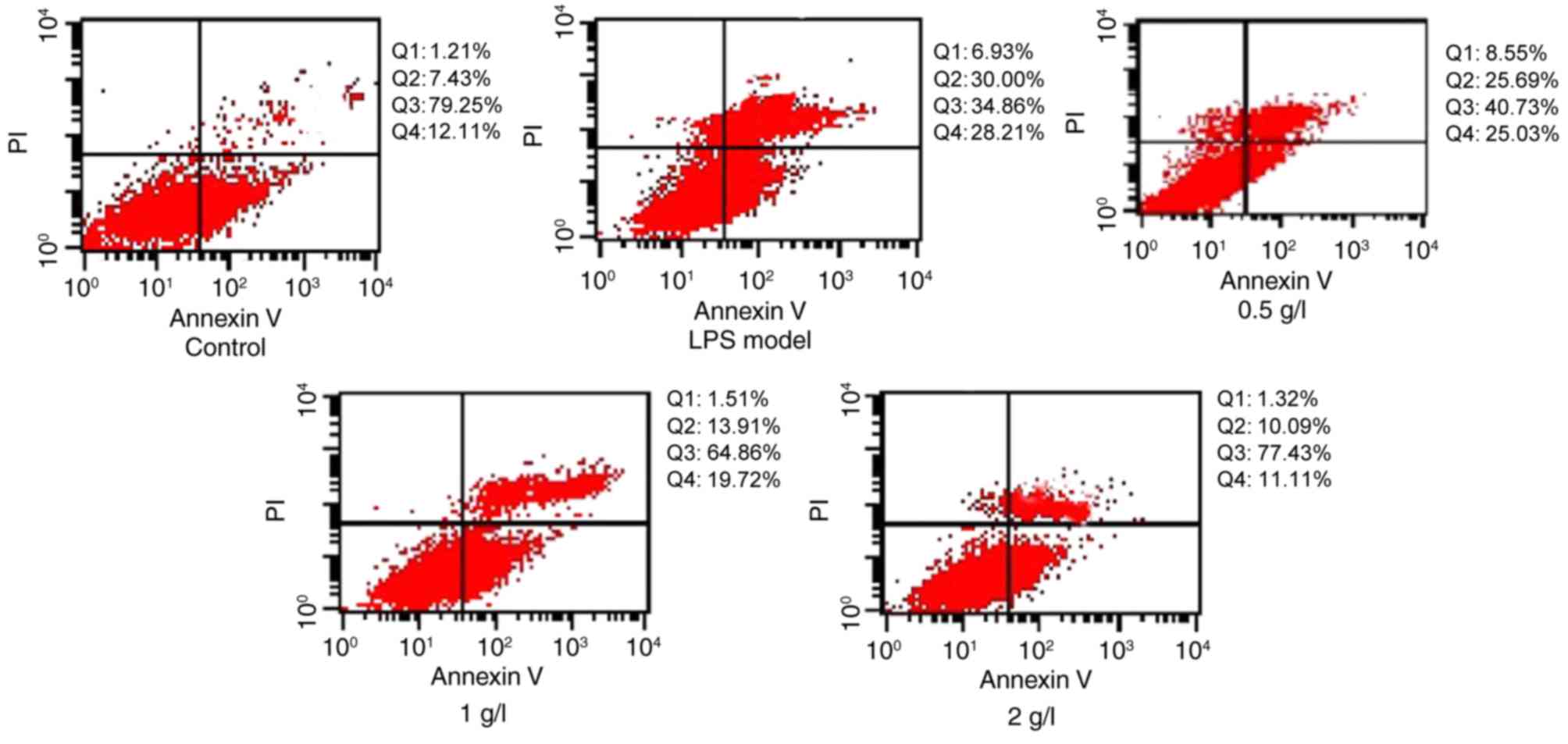
Effect of Patrinia on apoptosis of
BRL-3A cells after LPS stimulation
Flow cytometric analysis demonstrated that apoptosis
of BRL-3A cells in the LPS group was significantly higher compared
with the control group. However, the Patrinia-treated groups
exhibited reduced apoptosis of BRL-3A cells induced by LPS, with a
statistical significance (Fig.
5).
Western blotting to detect the
expression levels of TLR4, AKT, P-AKTSer473, GSK3β,
P-GSK3βSer9, P38, P-P38, JNK, P-JNK, Bax, Bcl-2 and
active-caspase-3 proteins
The results revealed that the expression levels of
TLR4, P-P38, P-JNK, Bax and active-caspase-3 were reduced by
treatment with Patrinia, whereas the expression of
P-GSK3βSer9, P-AKTSer473 and Bcl-2 were
increased, compared with the LPS group (Fig. 6A-C).
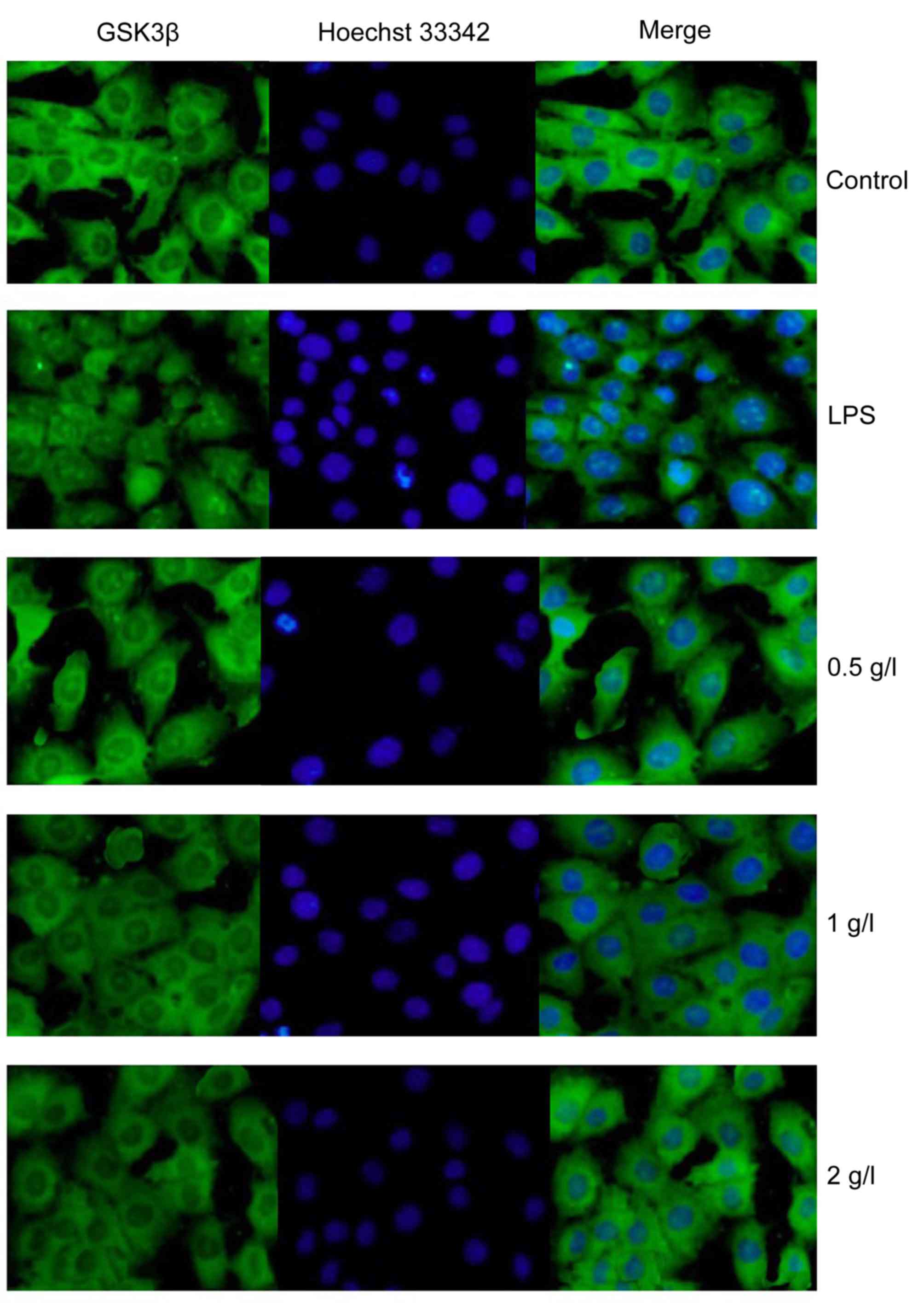
Effects of Patrinia on GSK3β
translocation in BRL-3A cells after LPS stimulation
Immunofluorescence staining was performed to observe
the nuclear translocation of GSK3β, which was localized primarily
in the cytoplasm (green fluorescence) in the control group. After
24 h of LPS stimulation, the majority of the GSK3β may have
translocated to the nucleus, as the green fluorescence appeared to
be co-localized with the blue nucleus. Compared with the LPS group,
the Patrinia-treated groups exhibited a decrease in GSK3β nuclear
translocation (Fig. 7).
Discussion
TLR4 is a member of the TLR family and is a
pattern-recognition receptor (8).
Studies have demonstrated that TLR4 expression is increased in
liver injury and acute liver failure (9).
The PI3K/AKT signaling pathway is an important
regulator of cell survival and apoptosis (10). PI3K is a conservative signal
transduction enzyme, involved in regulating cell proliferation and
inhibiting apoptosis (11). PI3K
is an enzyme complex consisting of a regulatory subunit (p85) and a
catalytic subunit (p110) (12).
Activation of PI3 K may promote the formation of a secondary
messenger, phosphatidylinositol (3–5)-triphosphate, which activates AKT
phosphorylation. Activated AKT may then activate or inhibit its
downstream substrates, including the phosphorylation of GSK3,
thereby regulating cell proliferation, differentiation, apoptosis
and migration processes (13–15).
AKT may also inhibit apoptosis by inhibiting the release of
cytochrome c via the suppression of Bax activities, and also by
phosphorylating Bcl-2 at Ser136 site to prevent the inhibition of
B-cell lymphoma extra-large. In the present study, the underlying
mechanism of Patrinia inhibition of BRL-3A cell apoptosis was
investigated. Western blotting was performed to detect the
expression levels of certain proteins. The expression of TLR4 in
BRL-3A cells was significantly increased by LPS stimulation, the
expression of P-AKTSer473 and P-GSK3βSer9 was
decreased by LPS, and the PI3K/AKT/GSK3β signaling pathway was
activated. However, treatment with Patrinia reduced the expression
of TLR4 induced by LPS, and promoted the phosphorylation of AKT and
GSK3β. This suggested that Patrinia inhibited LPS-induced apoptosis
of BRL-3A cells.
MAPK, as a signal transporter, carries external
signals to the nucleus, and mediates a number of biological
effects, including cell proliferation, differentiation and
apoptosis, via a series of protein kinase cascade reactions.
Studies have demonstrated that the MAPK signaling pathway includes
P38 and JNK signaling pathways (16). p38 is a 38 kDa protein consisting
of 360 amino acids, which belongs to the stress protein kinase
family. It has been revealed that P38 has five isomers. The P38
pathway may be activated by inflammatory factors, such as tumor
necrosis factor and interleukin-1, stress stimuli such as UV light,
hypoxia and heat shock, as well as LPS and gram-positive bacterial
cell wall components. The phosphorylation of P38 indicates its
activation (17).
JNK is a serine/threonine protein kinase identified
in 1990 (18). In mammalian cells,
JNK in encoded by the JNK1, JNK2 and JNK3 genes. Cell
model studies have shown that activation of the JNK pathway is
involved in the initiation of apoptosis induced by various stimuli.
JNK activation is involved not only in drug-induced apoptosis, but
also in DNA damage-induced apoptosis (19). P-JNK, the phosphorylated product of
JNK, indicates the activation of JNK. JNK is activated in the
presence of inflammatory factors, such as heat shock, ionizing
radiation, oxidative stress and DNA damage (20,21).
JNK may phosphorylate a variety of transcription factors, such as
JNK, p53 and ETS domain containing protein Elk-1, which have been
involved in the expression of genes associated with cell
proliferation and apoptotic processes. In the present study, BRL-3A
cells were stimulated with LPS, and western blotting was performed
to detect the expression levels of P38, JNK and their
phosphorylated proteins. LPS increased the expression levels of
phosphorylated P38 and JNK proteins, suggesting that the activation
of P38 and JNK promote apoptosis of BRL-3A cells. However, the
phosphorylation levels of JNK and P38 in BRL-3A cells were
significantly reduced by Patrinia treatment; thus apoptosis of
BRL-3A cells was inhibited. The inhibition of LPS-induced BRL-3A
cell apoptosis by Patrinia was associated with a decrease in P-JNK
and P-P38 expression. Patrinia may serve an anti-apoptotic role
through inhibition of the TLR4/P38/JNK signaling pathway.
In conclusion, treatment with Patrinia may reduce
apoptosis of BRL-3A cells induced by LPS. Through inhibition of the
expression levels of TLR4 in BRL-3A cells, Patrinia increased the
expression of P-AKTSer473 and P-GSK3βSer9,
decreased GSK3β nuclear translocation, reduced P38 and JNK
phosphorylation levels and decreased Bax/Bcl-2 ratio and caspase-3
expression. Therefore, Patrinia may inhibit BRL-3A cell apoptosis
via the TLR4/PI3 K/AKT/GSK3β and TLR4/P38/JNK signaling pathways.
However, further research is required to quantitatively confirm
that Patrinia treatment in LPS-stimulated BRL-3A cells leads to
GSK3β nuclear translocation, as this was determined by eye in the
present study. In conclusion, evidence was provided for the effects
of TLR4-mediated signaling on hepatocyte apoptosis in acute liver
failure, and targeting of this signaling may aid in the development
of effective drugs for the prevention and treatment of hepatocyte
apoptosis in acute liver failure.
References
|
1
|
Lopez M, Kopec AK, Joshi N, Geddings JE,
Cline H, Towery KL, Rockwell CE and Mackman N: Fas-induced
apoptosis increases hepatocyte tissue factor procoagulant activity
in vitro and in vivo. Toxicol Sci. 141:453–464. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jiang X and Wang X: Cytochrome c promotes
caspase-9 activation by inducing nucleotide binding to Apaf-1. J
Biol Chem. 275:31199–31203. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tait SW and Green DR: Mitochondria and
cell death: Outer membrane permeabilization and beyond. Nat Rev Mol
Cell Biol. 11:621–632. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fulda S: Modulation of mitochondrial
apoptosis by PI3K inhibitors. Mitochondrion. 13:195–198. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Johnson-Farley NN, Patel K, Kim D and
Cowen DS: Interaction of FGF-2 with IGF-1 and BDNF in stimulating
Akt, ERK, and neuronal survival in hippocampal cultures. Brain Res.
1154:40–49. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tournier C, Hess P, Yang DD, Xu J, Turner
TK, Nimnual A, Bar-Sagi D, Jones SN, Flavell RA and Davis RJ:
Requirement of JNK for stress-induced activation of the cytochrome
c-mediated death pathway. Science. 288:870–874. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang YF: The research progress of
Patrinia. J Chin Med Mat. 1:148–152. 2009.(In Chinese).
|
|
8
|
Du C, Fang M, Li Y, Li L and Wang X: Smac,
a mitochondrial protein that promotes cytochrome c-dependent
caspase activation by eliminating IAP inhibition. Cell. 102:33–42.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mohammadi A, Tajik N, Shah-Hosseini A,
Alavian SM, Sharifi Z and Jarahi L: FAS and FAS-ligand promoter
polymorphisms in hepatitis B virus infection. Hepat Mon.
15:e264902015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Muzio M, Chinnaiyan AM, Kischkel FC,
O'Rourke K, Shevchenko A, Ni J, Scaffidi C, Bretz JD, Zhang M,
Gentz R, et al: FLICE, a novel FADD-homologous ICE/CED-3-like
protease, is recruited to the CD95 (Fas/APO-1) death-inducing
signaling complex. Cell. 85:817–827. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Flanagan L, Sebastià J, Tuffy LP, Spring
A, Lichawska A, Devocelle M, Prehn JH and Rehm M: XIAP impairs Smac
release from the mitochondria during apoptosis. Cell Death Dis.
1:e492010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen HC, Kanai M, Inoue-Yamauchi A, Tu HC,
Huang Y, Ren D, Kim H, Takeda S, Reyna DE, Chan PM, et al: An
interconnected hierarchical model of cell death regulation by the
BCL-2 family. Nat Cell Biol. 17:1270–1281. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gross A: BCL-2 family proteins as
regulators of mitochondria metabolism. Biochim Biophys Acta.
1857:1243–1246. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Martinou JC and Youle RJ: Mitochondria in
apoptosis: Bcl-2 family members and mitochondrial dynamics. Dev
Cell. 21:92–101. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ha H, Han D and Choi Y: TRAF-mediated
TNFR-family signaling. Curr Protoc Immunol Chapter. 11:Unit11.9D.
2009. View Article : Google Scholar
|
|
16
|
Weichhaus M, Chung ST and Connelly L:
Osteoprotegerin in breast cancer: Beyond bone remodeling. Mol
Cancer. 14:1172015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guicciardi ME, Malhi H, Mott JL and Gores
GJ: Apoptosis and necrosis in the liver. Compr Physiol. 3:977–1010.
2013.PubMed/NCBI
|
|
18
|
Yarza R, Vela S, Solas M and Ramirez MJ:
c-Jun N-terminal kinase (JNK) signaling as a therapeutic target for
alzheimer's disease. Front Pharmacol. 6:3212016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim YR, Kang TW, To PK, Xuan Nguyen NT,
Cho YS, Jung C and Kim MS: HOXB13-mediated suppression of
p21WAF1/CIP1 regulates JNK/c-Jun signaling in prostate cancer
cells. Oncol Rep. 35:2011–2016. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ma M, Wang L, Ma Y, Yang Y, Chen B and Zhu
X: Effects of norepinephrine on proliferation and apoptosis of
neonatal cardiac fibroblasts in rats. Zhonghua xin xue guan bing za
zhi. 43:542–547. 2015.(In Chinese). PubMed/NCBI
|