Introduction
Epidemiologic and case control studies have
consistently indicated that moderate and mild elevation of plasma
homocysteine (Hcy), an intermediate metabolite of methionine, is an
independent risk factor in the development of atherosclerosis (AS)
(1). Previous studies have
intensively focused on the involvement of Hcy in the dysfunction
and injury of vascular cells, including vascular smooth muscle
cells (VSMCs) (2,3). The authors previously demonstrated
that Hcy induces VSMC proliferation, however the underlying
mechanisms remain to be elucidated (4).
Hcy is a non-protein, sulfur containing amino acid,
which is a metabolic intermediate of the methionine cycle. One of
the precursors is S-adenosylmethionine (SAM), the unique methyl
group donor for DNA methylation (5). Following the transfer of the methyl
group, SAM is transformed into S-adenosylhomocysteine (SAH), which
is hydrolyzed to form Hcy (6). DNA
methylation refers to the addition of a methyl group to the 5
position of cytosine in the context of a CpG dinucleotide.
Increasing evidence indicates that human diseases, including AS,
are either caused or impacted by abnormal methylation (7). The authors previously demonstrated
that abnormal DNA methylation of genes including peroxisome
proliferator activated receptor-α, apolipoprotein E and genomic DNA
contribute to the development of AS induced by Hcy (8,9).
Phosphatase and tensin homologue on chromosome 10 (PTEN), is a
dual-specificity protein and lipid phosphatase that suppresses
multiple signaling networks involved in cell proliferation,
survival and inflammation (10).
PTEN, specifically expressed in VSMCs, is additionally involved in
regulation of a variety of physiological and pathological
processes, including cell proliferation, differentiation,
apoptosis, adhesion and migration (11). PTEN is expressed endogenously in
VSMCs, and overexpression of PTEN significantly inhibits both basal
and platelet derived growth factor-mediated VSMC proliferation and
migration (12). PTEN
overexpression in VSMCs using adenovirus transfection results in
inhibition of cell proliferation and migration induced by
angiotensin II (13). However, the
direct mechanisms responsible for the PTEN influence on Hcy-induced
VSMC proliferation have not been explored.
Resveratrol (RSV), naturally occurring in various
plant foods, and particularly in grapes and red wine, is the most
investigated and well-known member of this class of compounds
(14). It exhibits a protective
role against cardiovascular diseases, and is involved in the
‘French paradox’, characterized by the low incidence of
cardiovascular diseases in the French population, despite a high
intake of saturated fats, due to their moderate red wine
consumption (15). RSV is an
effective therapeutic candidate, due to its protective action in
vascular walls towards oxidation, inflammation, platelet oxidation
and thrombus formation (16).
However, the association between Hcy and RSV on PTEN regulation in
VSMC proliferation remains to be elucidated.
Therefore, the present study sought to address: i)
The role of PTEN DNA methylation in VSMC proliferation induced by
Hcy; ii) whether the mechanism of methylation is regulated by DNA
methyltransferase (DNMT)1.
Materials and methods
Cell culture
The T/G HA-VSM cell line was used, which was
obtained from American Type Culture Collection (Manassas, VA, USA).
Cells were cultured in Dulbecco's modified Eagle's medium-Han's F12
media (DMEM-F12; Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) supplemented with 10% FCS at 37°C in an incubator with an
environment containing 5% CO2. Hcy was applied at
concentrations of 50, 100, 200 and 500 µM in addition to 30 µM
folate and vitamin B12 (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) for 72 h.
Bromodeoxyuridine (BrdU) assay
VSMCs at 5×103 cells per well were
cultured in triplicate in 96-well plates in 0, 50, 100, 200 and 500
µM concentrations of Hcy and antagonist (30 µM folate and 30 µM
vitamin B12 were added to 100 µM Hcy) then incubated for 72 h. Cell
proliferation was measured by BrdU incorporation assay using a
commercial ELISA kit according to the manufacturer's protocol (cat.
no. 11647229001; Roche Diagnostics GmbH, Mannheim, Germany).
Cell viability assessment
MTT (Sigma-Aldrich; Merck KGaA) was used for the
evaluation of cell viability. The cells were grown in 96-well
microtiter plates at a density of 1×104 cells in 200 µl
per well. Subsequently, cells were treated with treated with a
range of 10–100 µM RSV to determine an optimal concentration.
Following incubation for 10 min, the plates were read on a
microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
at 490 nm.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) of DNMT1 and PTEN
RNA was then reverse transcribed by using the Revert
Aid first strand cDNA synthesis kit (Applied Biosystems; Thermo
Fisher Scientific, Inc.); the SYBR-Green kit (Fermentas; Thermo
Fisher Scientific, Inc., Pittsburgh, PA, USA) was then used for
RT-qPCR analysis. The primer sequences of the DNMT1 and PTEN genes
were as follows: Forward, 5′-AGCTCTTACCTTACCATC-3′ and reverse,
5′-CCATCTGATACTCTGAA-3′ for DNMT1; and forward,
5′-CAGCATCTTATCCGAGTG-3′ and reverse, 5′-GATGGTGGTACAGTCAGA-3′ for
PTEN. GAPDH was applied as an internal control: Forward,
5′-AGAAGGCTGGGGCTCATTT-3′ and reverse, 5′-AGGGGCCACAGTCTTCG-3′. The
thermal cycler (Funglyn Biotech, Inc., Toronto, ON, Canada)
conditions comprised an initial activation step at 95°C for 5 min,
followed by a 2-step PCR program of 95°C for 15 sec, annealing
temperatures for 15 sec and at 72°C for 30 sec for 30 cycles.
Subsequently, the relative alterations in the mRNA expression
levels of DNMT1 and PTEN were determined by fold-change analysis,
in which the degree of change was calculated as 2−∆∆Cq,
where Cq=(Cqgene-CqGAPDH)
treatment-(Cqgene-CqGAPDH) control (17).
Western blotting for DNMT1 and
PTEN
Total proteins were isolated from the cells using
cell lysis buffer (Keygen Biotech Co., Ltd., Nanjing, China), then
protein concentration was determined via a bicinchoninic acid
protein assay kit (KeyGen Biotech, Nanjing, China). Equal amounts
of protein (~80 µg) and known molecular weight marker were
separated by 12% SDS-PAGE and were transferred to polyvinylidene
fluoride membrane by electrophoresis at 300 mA for 50 min at 4°C,
the membrane was then blocked in 10 ml 5% skimmed milk for 2 h at
room temperature with gentle agitation on a platform shaker. The
DNMT1 and PTEN proteins were detected using DNMT1 (cat. no.
sc-271729) and PTEN (cat. no. sc-65604) antibodies were obtained
from Santa Cruz Biotechnology, Inc, (Dallas, TX, US) diluted 1:500,
and β-actin protein was detected using a rabbit anti-human β-actin
antibody (cat. no. sc-70319, Santa Cruz Biotechnology, Inc.)
diluted 1:2,000; all primary antibodies were incubated at 4°C. The
secondary antibody (goat anti-mouse immunoglobulin G-horseradish
peroxidase, cat. no. sc-2031, 1:2,000; Santa Cruz Biotechnology,
Inc.) was added for 2 h at room temperature. The protein bands were
visualized and analyzed by the Gel Documentation and Analysis
System ChemiDoc XRS system with Image Lab software, version 4.1
(Bio-Rad Laboratories, Inc.) and calculated by the gray value of
the bands.
Nested methylation-specific (nMS)-PCR
for PTEN methylation assay
DNA denaturation and bisulfite conversion processes
were integrated into one step by using the EZ DNA
Methylation-Gold™ kit (Zymo Research Corp., Irvine, CA,
USA). nMS-PCR consists of two-step PCR amplifications following a
standard sodium bisulfite DNA modification. The first step uses an
outer primer pair set that does not contain any CpGs. The
second-step PCR was conducted with the conventional PCR primers.
The primers of the nMS-PCR assays were as follows: i) PTEN-outer
primers: Forward, 5′-GTTTTGGTTTGAAGGATAGTAGT-3′ and reverse,
5′-AAAAACCCTAAAACTTAATAAAAAC-3′; ii) PTEN-methylated primers:
Forward, 5′-TTAGTTTTAGTTAGGATGGTTTCGA-3′ and reverse,
5′-GAAAAATAAACCGAAATCCCG-3′; iii) PTEN-unmethylated primers:
Forward, 5′-ATTAGTTTTAGTAGGATGGTTTTGA-3′ and reverse,
5′-CAAAAAATAAACCAAAATCCCAC-3′. DNA bands were visualized by
ultraviolet illumination, and the percentage of methylation was
calculated by using the following formula: Methylation
%=methylation/(methylation+unmethylation) ×100%.
Cell transfection
The recombinant plasmids EGFP-N1-PTEN/DNMT1 were
established obtain from Hanbio Biotechnology Co., Ltd., (Shanghai,
China). Cells were plated to 70–90% confluence at the time of
transfection; 100 ng plasmid and 0.5 µl for DNA-lipid complexes
were prepared and incubated for 30 min. The DNA-lipid complexed
were then applied to cells. Then, the VSMCs were transfected with
3–5 µg/µl recombinant plasmids: EGFP-N1-PTEN/DNMT1.
EGFP-N1-PTEN/DNMT1 molecule-Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) complexes were
established and then added to the plate wells, where the VSMCs had
grown to 80–90% confluence. The transfection efficiency was
determined by detecting the fluorescence intensity via an inverted
fluorescence phase contrast microscope (Olympus IX71, Olympus
Corporation, Tokyo, Japan). The transfected VSMCs were treated with
Hcy for 24 h and mRNA expression of PTEN/DNMT1 was detected with
RT-qPCR (17). Western blotting
was carried out to examine the protein expression of PTEN/DNMT1
(17).
Statistical analysis
Prism 5.0 (GraphPad Software, Inc., La Jolla, CA,
USA) was used for data processing. Data are expressed as the mean ±
standard deviation. Each experiment was repeated three times.
Statistical comparison of each parameter between two groups was
performed using the paired Student's t-test. One-way analysis of
variance was used to compare the means of multiple groups, followed
by Dunnett's test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Hcy promotes the proliferation of
VSMCs
In order to investigate the effect of Hcy on VSMCs,
the present study detected the cell proliferation ratio following
cell treatment with Hcy for 72 h, using an MTT assay. As presented
in Fig. 1A, the proliferation of
VSMCs was increased in the cells which were co-incubated with
different concentrations of Hcy, compared with control group, and
in 100 µM Hcy it was increased 1.72-fold (P<0.05). The
proliferation ratio in the folate and vitamin B12 group treated
with 100 µM Hcy was decreased by 35.46% (P<0.05). Furthermore,
the effect of Hcy on VSMC proliferation was evaluated with a BrdU
incorporation assay to determine the effects of Hcy on DNA
synthesis, and it was demonstrated that 100 µM Hcy treatment
significantly promoted DNA synthesis (Fig. 1B), whereas in the folate and
vitamin B12 group, DNA synthesis was decreased (P<0.05). These
data indicated that Hcy promoted the proliferation ratio, and this
effect was counteracted with folate and vitamin B12.
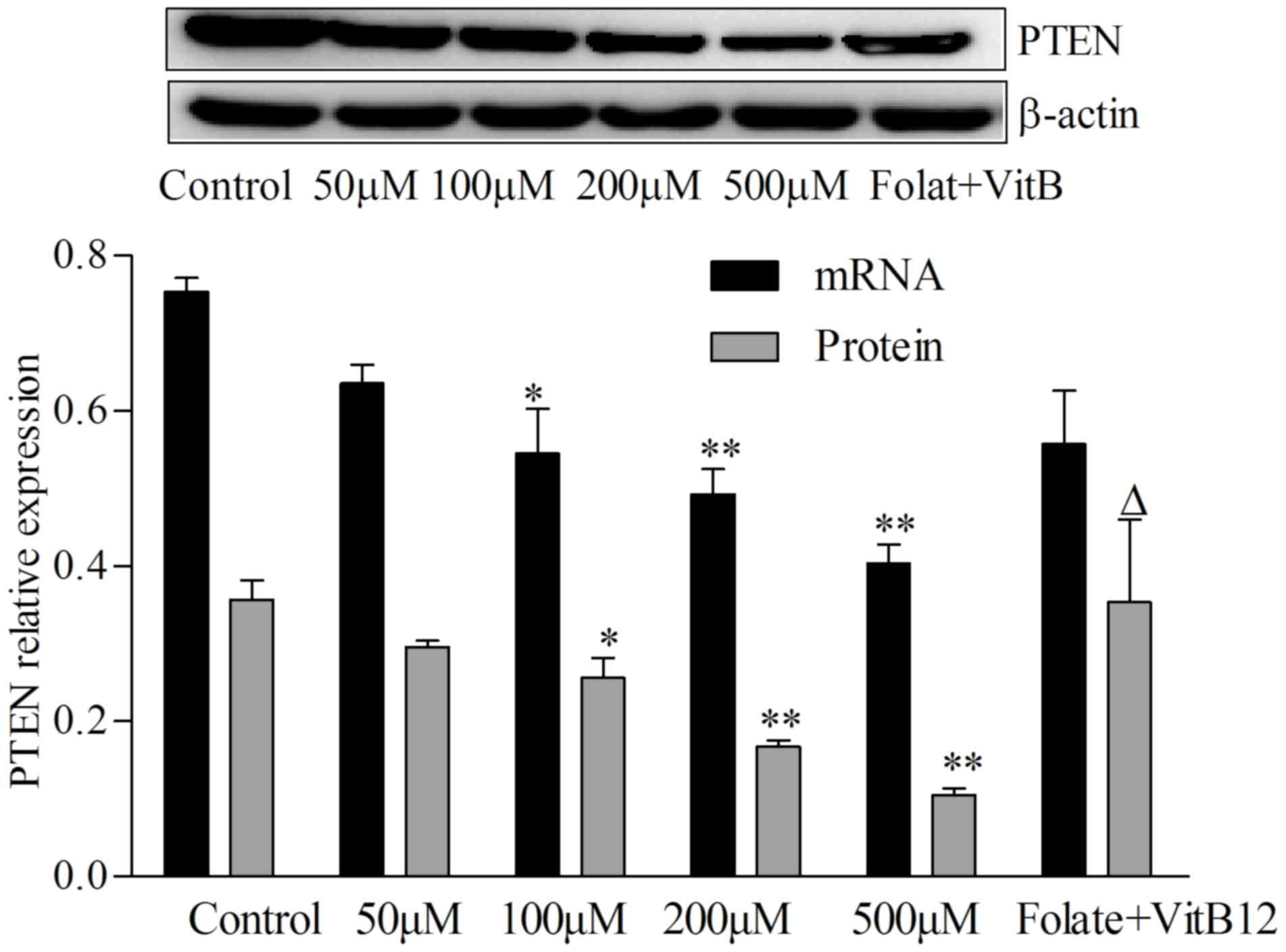
Hcy decreases PTEN expression in
VSMCs
PTEN is ranked the most mutated tumor suppressor
gene, which regulates numerous cellular processes, including cell
cycle, adhesion, migration, proliferation and cell survival
(18). To study whether Hcy
regulates PTEN expression, and if this was responsible for the VSMC
proliferation, the present study detected PTEN expression levels in
VSMCs following treatment with Hcy for 72 h. As presented in
Fig. 2, PTEN mRNA was decreased by
27.5, 34.7 and 46.7% in 100, 200 and 500 µM Hcy groups
respectively, compared with control group (P<0.05, P<0.01).
PTEN protein levels in VSMCs decreased by 28.6, 54.3 and 71.4% in
100, 200 and 500 µM Hcy groups, compared with control group
(P<0.05, P<0.01), whereas in folate and vitamin B12 group,
PTEN protein levels were increased 1.44-fold, compared with 100 µM
Hcy group (P<0.05). Therefore, PTEN may have an important role
in VSMCs proliferation induced by Hcy.
RSV antagonizes proliferation of VSMCs
induced by Hcy
RSV has biological activities, and similar to
estrogen, is known to have several effects on cardiovascular
disease and malignancies (19).
The present study co-incubated VSMCs with different concentrations
of RSV and 100 µM Hcy for 72 h at 37°C, then detected the VSMC
proliferation ratio by MTT, as presented in Fig. 3. Compared with 100 µM Hcy group,
the proliferation ratio of VSMCs was decreased by 45.16, 53.76 and
63.44% in 20, 50 and 100 µM RSV groups (P<0.05, P<0.01),
respectively. Compared with 100 µM RSV group, the proliferation
ratio in the folate and vitamin B12 group was increased by
1.62-fold (P<0.05). Notably, compared with 100 µM Hcy group, the
folate and vitamin B12 group proliferation ratio of VSMCs was
decreased by 40.86% (P<0.05). These data indicated that RSV
acted as an antagonist which decreased the proliferation ratio of
VSMCs, which was previously increased when induced by Hcy. The
inhibitory effect of RSV was more potent compared with folate and
vitamin B12.
 | Figure 3.Resveratrol antagonistic effects on
proliferation of VSMCs induced by Hcy. VSMCs were treated with Hcy
and different concentrations of Resveratrol for 72 h, then the
proliferation ratio of VSMCs was detected by MTT. A, 100 µM Hcy
group, as control group; B, 100 µM Hcy+10 µM Resveratrol; C, 100 µM
Hcy+20 µM Resveratrol; D, 100 µM Hcy+50 µM Resveratrol; E, 100 µM
Hcy+100 µM Resveratrol; F, cells were treated with 100 µM Hcy, 30
µM folate and 30 µM vitamin B12. *P<0.05, **P<0.01, compared
with 100 µM Hcy group; ΔP<0.05 vs. 100 µM Hcy+100 µM Resveratrol
group. Hcy, homocysteine; VSMC, vascular smooth muscle cells. |
Methylation status of PTEN in
VSMCs
DNA methylation is important in the development of
AS induced by Hcy (20). To
investigate whether PTEN DNA methylation alterations occur in VSMC
proliferation induced by Hcy, the present study detected PTEN DNA
methylation by nMS-PCR, following cell treatment with Hcy and
various antagonistic compounds for 72 h. As presented in Fig. 4A, compared with control group, PTEN
DNA methylation levels were increased 8.56-fold in 100 µM Hcy group
(P<0.01), and 2.56-fold in folate and vitamin B12 group
(P<0.05). Compared with 100 µM Hcy group, PTEN DNA methylation
levels decreased by 79.2 and 70.1% in RSV (P<0.01) and folate
and vitamin B12 groups (P<0.01), respectively. These data
suggested that PTEN DNA methylation was affected by Hcy in VSMCs
and this may act as the primary reason for proliferation of VSMCs
induced by Hcy.
 | Figure 4.PTEN DNA methylation and its
regulation mechanism in VSMCs induced by Hcy. (A) PTEN DNA
methylation levels in VSMCs detected by nested methylation-specific
PCR, following cell treatment with Hcy and other antagonists for 72
h. (B) DNMT1 expression levels in VSMCs detected by RT-qPCR and
western blotting, following cell treatment with Hcy and other
antagonists for 72 h. (C) pEGFP-N1-DNMT1 was transfected into VSMCs
using Lipofectamine® 2000, and detected with
fluorescence microscopy (magnification, ×1,000). (D) DNMT1 mRNA and
protein levels in VSMCs analyzed by RT-qPCR and western blotting,
following cell transfection with DNMT1 overexpression vector. (E)
Levels of PTEN DNA methylation in VSMCs, following cell
transfection with recombination vector of DNMT1, then co-incubation
with Hcy for 72 h. Control group, untreated cells; Hcy group: Cells
treated with 100 µM Hcy for 72 h; RSV group, cells treated with 100
µM Hcy and 100 µM Resveratrol for 72 h; Folate and vitamin B12
group, cells treated with 100 µM Hcy, 30 µM folate and 30 µM
vitamin B12; EGFP-N1, pEGFP-N1 control vector; DNMT1-N1,
recombination vector of DNMT1; DNMT1 group, cells transfected with
recombination vector of DNMT1; DNMT1+Hcy group, cells transfected
with recombination vector of DNMT1, then treated with 100 µM Hcy
for 72 h; DNMT1+Hcy+RSV group, cells transfected with recombination
vector of DNMT1, then treated with 100 µM Hcy and 100 µM
Resveratrol for 72 h; *P<0.05, **P<0.01, vs. control group;
ΔP<0.05, ΔΔP<0.01 vs. 100 µM Hcy group; #P<0.05 vs.
pEGFP-N1 vector group. M, methylation; U, unmethylated; Marker, DNA
marker (top to bottom, 2,000, 1,000, 750, 500, 250 and 100 bp).
Hcy, homocysteine; VSMC, vascular smooth muscle cells; PTEN,
phosphatase and tensin homologue on chromosome 10; DNMT1, DNA
methyltransferase; RSV, Resveratrol; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction. |
DNMT1 catalyzes the transfer of methyl groups to DNA
from SAM. In mammals, DNMT1 is the maintenance methyltransferase,
which preferentially methylates unmethylated or hemi-methylated
double-stranded DNA. To further study the regulation mechanism of
PTEN DNA methylation, the expression of DNMT1 in VSMCs was analyzed
by RT-qPCR and western blotting. As presented in Fig. 4B, DNMT1 mRNA levels were detected
in VSMCs. Compared with control group, DNMT1 mRNA increased
3.28-fold in 100 µM Hcy group (P<0.01), whereas compared with
100 µM Hcy group, DNMT1 mRNA levels decreased 47.4 and 57.9% in RSV
(P<0.01) and folate and vitamin B12 groups (P<0.01),
respectively. DNMT1 protein expression in VSMCs exhibited the same
trends as the DNMT1 mRNA levels. Compared with control group, DNMT1
protein expression increased 2.81-fold in the100 µM Hcy group
(P<0.01), whereas compared with 100 µM Hcy group, DNMT1 protein
levels decreased by 47.4 and 43.4% in RSV (P<0.01) and folate
and vitamin B12 groups (P<0.01), respectively.
To identify whether DNMT1 was a key molecule in
regulation of PTEN DNA methylation in VSMC proliferation induced by
Hcy, the levels of DNMT1 were overexpressed. The recombination
vector of DNMT1 was transfected into untreated VSMCs, and the
observed green fluorescence verified that the recombined vector had
been successfully transfected into VSMCs (Fig. 4C). The expression levels of DNMT1
were detected with RT-qPCR and western blotting. As presented in
Fig. 4D, DNMT1 mRNA and protein
expression levels were increased in the overexpression group by
3.57-fold and 2.03-fold (P<0.01), compared with control group,
and these data indicated that the transfection of the
overexpression recombination vector DNMT1 had been successful.
Following this, PTEN DNA methylation was analyzed with nMS-PCR. As
presented in Fig. 4E, compared
with control group, PTEN DNA methylation was increased 2-fold
(P<0.05) in DNMT1 overexpression group and 2.59-fold (P<0.01)
in DNMT1 overexpression and Hcy group. Overall, these data
suggested that DNMT1 has an important role in regulating PTEN DNA
methylation in VSMCs, induced by Hcy.
PTEN exhibits a primary role in the
proliferation of VSMCs induced by Hcy
To further verify the role of PTEN in VSMC
proliferation resulting from Hcy, a recombination PTEN
overexpression vector was transfected into untreated VSMCs and
proliferation of VSMCs induced by Hcy was analyzed, as presented in
Fig. 5. Following transfection
with recombination vector of PTEN, green fluorescence in VSMCs was
observed, and the levels of PTEN mRNA and protein were increased
1.91-fold (P<0.01) and 2.27-fold (P<0.01) in the PTEN
recombination group (Fig. 5A and
B).
 | Figure 5.PTEN is a pivotal molecule in
proliferation of VSMCs induced by Hcy. (A) pEGFP-N1 and
pEGFP-N1-PTEN were transfected into VSMC using Lipofectamine™ 2000,
and detected with fluorescence microscopy (magnification, ×100).
(B) Expression levels of PTEN in VSMCs, following cell transfection
with recombination vector. mRNA and protein levels were analyzed
with reverse transcription-quantitative polymerase chain reaction
and western blotting. (C) Proliferation ratio of VSMCs was detected
by MTT, following cell transfection with recombination vector PTEN,
and then co-incubation with Hcy and other antagonistic compounds.
EGFP-N1, pEGFP-N1 control vector; PTEN-N1, recombination vector of
PTEN; Hcy group, cells treated with 100 µM Hcy for 72 h;
N1-PTEN+Hcy group, cells transfected with recombination vector of
PTEN, then treated with 100 µM Hcy for 72 h; N-PTEN+Hcy+RSV group,
cells transfected with recombination vector of PTEN, then treated
with 100 µM Hcy and 100 µM Resveratrol for 72 h; *P<0.05,
**P<0.01 vs. control group; ΔP<0.05 vs. pEGFP-N1 vector
group; #P<0.05, vs. 100 µM Hcy group. Hcy, homocysteine; VSMC,
vascular smooth muscle cells; PTEN, phosphatase and tensin
homologue on chromosome 10. |
Following this, the proliferation ratio of VSMCs was
detected following cell transfection with the PTEN recombination
vector. As presented in Fig. 5C,
the proliferation ratio was increased 4.75-fold (P<0.01),
3.25-fold (P<0.01) and 2.1-fold (P<0.01) in Hcy, PTEN
overexpression cells treated with Hcy and PTEN overexpression cells
treated with Hcy and RSV groups, respectively. Compared with 100 µM
Hcy group, in PTEN overexpression cells treated with Hcy and PTEN
overexpression cells treated with Hcy and RSV, the proliferation
ratio was decreased by 33.3% (P<0.05) and 56.6% (P<0.05).
These results suggested that PTEN is important in proliferation of
VSMCs which is induced by Hcy, and RSV may relieve this nocuous
effect of Hcy.
Discussion
It has previously been demonstrated that elevated
Hcy levels are a diagnosed cause of VSMC proliferation and further
promote the formation of AS (21).
The present study demonstrated that Hcy induced the proliferation
of VSMCs, which may contribute to development of AS. Clinical
studies regarding treatment with RSV are not as promising as the
preclinical findings, with regards to the beneficial effects of RSV
on AS (22,23). The present study demonstrated that
Hcy promoted VSMC proliferation, and decreased PTEN expression via
an increase in PTEN DNA methylation levels. DNMT1 was important in
this process, and RSV relieved the pathological process via a
promotion of PTEN expression levels.
As an important independent risk factor of AS, Hcy
stimulates VSMC proliferation, which has been demonstrated in
previous studies (24,25). To investigate the effect of Hcy on
VSMCs, the present study detected the cell proliferation ratio
following cell treatment with Hcy for 72 h using an MTT assay, and
the proliferation of VSMCs was observed to be increased with
different concentrations of Hcy. PTEN is a tumor suppressor gene,
which regulates various cellular processes, including cell cycle,
adhesion, migration, proliferation and cell survival (26). PTEN has recently been demonstrated
to have an important role in cells of the cardiovascular system
(27). Schwartzbauer and Robbins
(28) demonstrated that PTEN
regulates cardiac myocyte hypertrophy and survival. The present
study demonstrated that PTEN expression levels were decreased in
Hcy groups, at the mRNA and protein level.
RSV is a naturally occurring polyphenolic
phytoalexin derived from plants such as Polygonum cuspidatum
and grapes, it has anti-aging, -apoptotic, -tumorigenic, -oxidant,
and -inflammatory properties (29). Guo (30) reported that RSV inhibits high
glucose-induced oxidative stress and VSMC proliferation by
suppressing reactive oxygen species generation, nicotinamide
adenine dinucleotide phosphate oxidase, AKT Serine/Threonine Kinase
phosphorylation, p38 mitogen activated protein kinase/c-Jun
N-terminal kinase/extracellular signal-regulated kinase
phosphorylation, and IκB-α and nuclear factor-11κB activities,
however the effects of RSV in anti-proliferation of VSMCs still
remain unclear. The present study co-incubated VSMCs with different
concentrations of RSV and 100 µM Hcy for 72 h, and demonstrated
that the VSMC proliferation ratio was decreased in different
concentrations of RSV groups. These data indicated that RSV acted
as an antagonist which decreased the proliferation ratio of VSMCs
induced by Hcy.
DNA methylation serves as an important mechanism
that controls gene expression in AS (31). As previously demonstrated, PTEN DNA
methylation levels are significantly decreased in Hcy groups. In
Hcy metabolism, SAM is a metabolic intermediate which is
synthesized from methionine catalysed by methionine
adenosyltransferase, providing methyl group moieties in several
dozens of transmethylation reactions (32). SAM is converted into SAH that is
the sole metabolic precursor of Hcy in a reversible reaction
catalyzed by SAH hydrolase, and inhibits DNMT1 (33). The present study demonstrated that
PTEN DNA methylation levels were increased in 100 µM Hcy group and
decreased in RSV group. The role of DNMT1 in the regulation of PTEN
was also observed. DNMT1, 3a, 3b have all previously been detected,
however only DNMT1 exhibits an important role in this process
(34). To test whether DNMT1 had
an effect on PTEN promoter hypermethylation, overexpression of
DNMT1 and PTEN occurred. Following transfection with pDNMT1, the
methylation levels of PTEN significantly increased.
In conclusion, Hcy impacted the methylation status
of PTEN involved in cell proliferation, and induced the
proliferation of VSMCs, whereas RSV alleviated VSMC proliferation
by antagonism of Hcy. The data from the present study provided
evidence for the mechanisms of VSMC proliferation in AS induced by
Hcy, and may act as a potential diagnostic marker for AS induced by
Hcy.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81570452, 81560084,
81670416, 81660088 and 81760076).
References
|
1
|
Devasia AJ, Joy B and Tarey SD: Serum
homocysteine as a risk factor for carotid intimal thickening in
acute stroke: A cross sectional observational study. Ann Indian
Acad Neurol. 19:48–51. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu T, Lin J, Ju T, Chu L and Zhang L:
Vascular smooth muscle cell differentiation to an osteogenic
phenotype involves matrix metalloproteinase-2 modulation by
homocysteine. Mol Cell Biochem. 406:139–149. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu X, Shen J, Zhan R, Wang X, Wang X,
Zhang Z, Leng X, Yang Z and Qian L: Proteomic analysis of
homocysteine induced proliferation of cultured neonatal rat
vascular smooth muscle cells. Biochim Biophys Acta. 1794:177–184.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Han XB, Zhang HP, Cao CJ, Wang YH, Tian J,
Yang XL, Yang AN, Wang J, Jiang YD and Xu H: Aberrant DNA
methylation of the PDGF gene in homocysteine-mediated VSMC
proliferation and its underlying mechanism. Mol Med Rep.
10:947–954. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Taysi S, Keles MS, Gumustekin K, Akyuz M,
Boyuk A, Cikman O and Bakan N: Plasma homocysteine and liver tissue
S-adenosylmethionine, S-adenosylhomocysteine status in vitamin
B6-deficient rats. Eur Rev Med Pharmacol Sci. 19:154–160.
2015.PubMed/NCBI
|
|
6
|
Elshorbagy AK, Jernerén F, Samocha-Bonet
D, Refsum H and Heilbronn LK: Serum S-adenosylmethionine, but not
methionine, increases in response to overfeeding in humans. Nutr
Diabetes. 6:e1922016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang L, Fu X, Peng X, Xiao Z, Li Z, Chen G
and Wang X: DNA methylation profiling reveals correlation of
differential methylation patterns with gene expression in human
epilepsy. J Mol Neurosci. 59:68–77. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang AN, Zhang HP, Sun Y, Yang XL, Wang N,
Zhu G, Zhang H, Xu H, Ma SC, Zhang Y, et al: High-methionine diets
accelerate atherosclerosis by HHcy-mediated FABP4 gene
demethylation pathway via DNMT1 in ApoE(−/−) mice. FEBS Lett.
589:3998–4009. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yideng J, Zhihong L, Jiantuan X, Jun C,
Guizhong L and Shuren W: Homocysteine-mediated PPARalpha, gamma DNA
methylation and its potential pathogenic mechanism in monocytes.
DNA Cell Biol. 27:143–150. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen WJ, Lin KH, Lai YJ, Yang SH and Pang
JH: Protective effect of propylthiouracil independent of its
hypothyroid effect on atherogenesis in cholesterol-fed rabbits:
PTEN induction and inhibition of vascular smooth muscle cell
proliferation and migration. Circulation. 110:1313–1319. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sedding DG, Widmer-Teske R, Mueller A,
Stieger P, Daniel JM, Gündüz D, Pullamsetti S, Nef H, Moellmann H,
Troidl C, et al: Role of the phosphatase PTEN in early vascular
remodeling. PLoS One. 8:e554452013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rice KM, Uddemarri S, Desai DH, Morrison
RG, Harris R, Wright GL and Blough ER: PGF2alpha-associated
vascular smooth muscle hypertrophy is ROS dependent and involves
the activation of mTOR, p70S6k, and PTEN. Prostaglandins Other
Lipid Mediat. 85:49–57. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hu C, Liu S, Sun Y, Shi G and Li Y: Effect
of recombinant hPTEN gene expression on PDGF induced VSMC
proliferation. Cell Biochem Biophys. 70:1185–1190. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moradi H and Vaziri ND: Effect of
Resveratrol on progression of polycystic kidney disease: A case of
cautious optimism. Nephrol Dial Transplant. 31:1755–1758. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pieszka M, Szczurek P, Ropka-Molik K,
Oczkowicz M and Pieszka M: The role of Resveratrol in the
regulation of cell metabolism-a review. Postepy Hig Med Dosw
(Online). 70:117–123. 2016.(In Polish). View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fišar Z, Hroudová J, Singh N, Kopřivová A
and Macečková D: Effect of simvastatin, coenzyme Q10, Resveratrol,
acetylcysteine and acetylcarnitine on mitochondrial respiration.
Folia Biol (Praha). 62:53–66. 2016.PubMed/NCBI
|
|
17
|
Ma SC, Cao JC, Zhang HP, Jiao Y, Zhang H,
He YY, Wang YH, Yang XL, Yang AN, Tian J, et al: Aberrant promoter
methylation of multiple genes in VSMC proliferation induced by Hcy.
Mol Med Rep. Sep 19–2017.(Epub ahead of print). View Article : Google Scholar
|
|
18
|
Hu C, Liu S, Sun Y, Shi G and Li Y: Effect
of recombinant hPTEN gene expression on PDGF induced VSMC
proliferation. Cell Biochem Biophys. 70:1185–1190. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dong X, Yu LG, Sun R, Cheng YN, Cao H,
Yang KM, Dong YN, Wu Y and Guo XL: Inhibition of PTEN expression
and activity by angiotensin II induces proliferation and migration
of vascular smooth muscle cells. J Cell Biochem. 114:174–182. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Greißel A, Culmes M, Napieralski R, Wagner
E, Gebhard H, Schmitt M, Zimmermann A, Eckstein HH, Zernecke A and
Pelisek J: Alternation of histone and DNA methylation in human
atherosclerotic carotid plaques. Thromb Haemost. 114:390–402. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Meng L, Liu L, Zhou C, Pan S, Zhai X,
Jiang C, Guo Y, Ji Z, Chi J, Peng F and Guo H: Polyphenols and
polypeptides in Chinese rice wine inhibit homocysteine-induced
proliferation and migration of vascular smooth muscle cells. J
Cardiovasc Pharmacol. 67:482–490. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Swiader A, Nahapetyan H, Faccini J,
D'Angelo R, Mucher E, Elbaz M, Boya P and Vindis C: Mitophagy acts
as a safeguard mechanism against human vascular smooth muscle cell
apoptosis induced by atherogenic lipids. Oncotarget. 7:28821–28835.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen WJ, Pang JH, Lin KH, Lee DY, Hsu LA
and Kuo CT: Propylthiouracil, independent of its antithyroid
effect, promotes vascular smooth muscle cells differentiation via
PTEN induction. Basic Res Cardiol. 105:19–28. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chawla V, Simionescu A, Langan EM III and
LaBerge M: Influence of clinically relevant mechanical forces on
vascular smooth muscle cells under chronic high glucose: An in
vitro dynamic disease model. Ann Vasc Surg. 34:212–226. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang C, Zhang H, Zhang W, Kong W, Zhu Y,
Zhang H, Xu Q, Li Y and Wang X: Homocysteine promotes vascular
smooth muscle cell migration by induction of the adipokine
resistin. Am J Physiol Cell Physiol. 297:C1466–C1476. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bonnevaux H, Lemaitre O, Vincent L, Levit
MN, Windenberger F, Halley F, Delorme C, Lengauer C,
Garcia-Echeverria C and Virone-Oddos A: Concomitant inhibition of
PI3Kβ and BRAF or MEK in PTEN-deficient/BRAF-mutant melanoma
treatment: Preclinical assessment of SAR260301 oral PI3Kβ-selective
inhibitor. Mol Cancer Ther. 15:1460–1471. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Morais CL, Guedes LB, Hicks J, Baras AS,
De Marzo AM and Lotan TL: ERG and PTEN status of isolated
high-grade PIN occurring in cystoprostatectomy specimens without
invasive prostatic adenocarcinoma. Hum Pathol. 55:117–125. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Schwartzbauer G and Robbins J: The tumor
suppressor gene PTEN can regulate cardiac hypertrophy and survival.
J Biol Chem. 276:35786–35793. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tang F, Guo S, Liao H, Yu P, Wang L, Song
X, Chen J and Yang Q: Resveratrol enhances neurite outgrowth and
synaptogenesis via sonic hedgehog signaling following
oxygen-glucose deprivation/reoxygenation injury. Cell Physiol
Biochem. 43:852–869. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guo R, Li W, Liu B, Li S, Zhang B and Xu
Y: Resveratrol protects vascular smooth muscle cells against high
glucose-induced oxidative stress and cell proliferation in vitro.
Med Sci Monit Basic Res. 20:82–92. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ma SC, Zhang HP, Kong FQ, Zhang H, Yang C,
He YY, Wang YH, Yang AN, Tian J, Yang XL, et al: Integration of
gene expression and DNA methylation profiles provides a molecular
subtype for risk assessment in atherosclerosis. Mol Med Rep.
13:4791–4799. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hai Z and Zuo W: Aberrant DNA methylation
in the pathogenesis of atherosclerosis. Clin Chim Acta. 456:69–74.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Grimaldi V, Vietri MT, Schiano C, Picascia
A, De Pascale MR, Fiorito C, Casamassimi A and Napoli C: Epigenetic
reprogramming in atherosclerosis. Curr Atheroscler Rep. 17:4762015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Benetatos L and Vartholomatos G: On the
potential role of DNMT1 in acute myeloid leukemia and
myelodysplastic syndromes: Not another mutated epigenetic driver.
Ann Hematol. 95:1571–1582. 2016. View Article : Google Scholar : PubMed/NCBI
|