Introduction
Restoring normal physiological function after
traumatic spinal cord injury (SCI) is difficult under conditions of
oxygen and energy deprivation, due to the severely compromised
energy metabolism in the injured spinal cord. Furthermore, new
nerves cannot grow because of the limited regenerative ability of
the central nervous system at the sites of injury, leading to a
loss of sensorimotor function below the point of injury (1,2).
Various measures to repair SCI have been developed; these can, to a
certain extent, promote axonal regeneration and functional
recovery, but there is still a long way to go to achieve complete
repair (3–8).
Polyunsaturated fatty acids (PUFAs) are essential
for mammals, but n-3 PUFAs cannot be synthesized by the human body
and must be obtained from foods. Research has indicated that n-3
PUFAs are closely involved in the physiological activities of the
nervous and immune systems, among others (9–13).
Investigations have revealed that n-3 PUFAs play an important role
in regulating the mammalian target of rapamycin complex 1 (mTORC1)
signaling pathway in the body (14–16).
Autophagy is a highly conserved cell degradation
process, which involves isolating part of the cytoplasm and
organelles in a bilayer vesicle and delivering them to lysosomes
for degradation, eventually recycling the large decomposed
molecules; LC3-II is one of the markers of autophagy (17,18).
Autophagy activity is widespread in injured spinal cords, and
studies have shown that autophagy is significantly activated in the
days just after SCI (19–22). Activation of autophagy can clear
intracellular damaged proteins and protect against neuronal loss to
promote recovery of motor function after SCI (23–25).
The role of autophagy in the experimental study of SCI is
attracting more and more attention.
mTOR is a highly-conserved serine/threonine protein
kinase consisting of two different compounds in the body, mTORC1
and mTORC2. Through nutrient, energy and growth factor signaling
pathways, mTORC1 regulates cell metabolism, growth, proliferation,
survival and autophagy (26,27).
Studies have shown that inhibiting mTORC1 can enhance autophagy and
help repair injuries (13,28–37).
It is known that the mTORC1 signaling pathway and autophagy
activity are involved in the repair of articular cartilage and
nervous tissues, playing a vital role in the recovery of damaged
tissue (13,15), but its role in SCI remains
unclear.
In this study, we created a rat SCI model and fed
the rats with a diet high in n-3 PUFAs to enhance the content of
n-3 PUFAs and the n-3/n-6 PUFA composition, and then explored
whether n-3 PUFAs can regulate autophagy through the mTORC1
signaling pathway to promote repair of SCI.
Materials and methods
In vitro experiments
Primary Schwann cells were obtained from National
Infrastructure of Cell Line Resource (RSC-96, 3111C0001CCC000664;
Beijing, China) and cultured according to previously-published
protocols (38). mTORC1 is a
signaling pathway sensitive to amino acids (aa). Removal and
readdition of aa eliminates and stimulates mTORC1 activity,
respectively. Thus, aa starvation was used in these experiments.
Exogenous docosahexaenoic acid (DHA) and arachidonic acid (AA) were
used as a representative n-3 PUFA and n-6 PUFA, respectively.
Neuronal cells were starved of amino acids by culturing in
Dulbecco's phosphate-buffered saline for 30 min. After this, either
DHA (50 µM) or AA (50 µM) (both from Cayman Chemical Company, Ann
Arbor, MI, USA) was added, cells were returned to culture for 30
min, then proteins were extracted and the activity of mTORC1
downstream protein p-S6 was analyzed to investigate the influence
of PUFAs on mTORC1. To investigate expression of the
autophagy-related protein LC3-II (Cell Signaling Technology, Inc.,
Danvers, MA, USA), cultures were treated in the same way but
returned to culture for 3 h before proteins were extracted to study
the influence of n-3 PUFAs on autophagy.
Animal groupings
Animal experiments were approved by the animal
experimental ethics committee of Southern Medical University
(GuangZhou, China) and all animals were purchased from the
Laboratory Animal Centre at Southern Medical University. Adult
Sprague-Dawley (SD) rats (90 female, weight: 200–250 g) were
randomly divided into three groups: A normal diet group (CON)
(n=30), a low n-3/n-6 diet group (Low), and a high n-3/n-6 diet
group (High) (Table I).
 | Table I.Polyunsaturated fatty acid
composition and content in the diets of experimental rats. |
Table I.
Polyunsaturated fatty acid
composition and content in the diets of experimental rats.
| Type of fatty
acids | CON (g/kg
diet) | Low n-3/n-6 (g/kg
diet) | High n-3/n-6 (g/kg
diet) |
|---|
| C18:3, n-3,
α-linoleic acid | 0.41 | 0.55 | 0.41 |
| C20:5, n-3,
eicosapentaenoic acid (EPA) | 2.80 | 4.47 | 29.75 |
| C22:5, n-3,
docosapentaenoic acid (DPA) | 0.79 | 0.85 | 0.83 |
| C22:6, n-3,
docosahexaenoic acid (DHA) | 2.41 | 3.63 | 17.19 |
| n-3, total | 6.41 | 9.50 | 48.18 |
| C18:2, n-6,
linoleic acid | 50.31 | 9.75 | 6.43 |
| C20:4, n-6,
arachidonic acid (AA) | 3.70 | 0.54 | 0.39 |
| n-6, total | 54.01 | 10.29 | 6.82 |
| n-3/n-6 | 0.118 | 0.923 | 7.04 |
SCI model
The three groups of rats were fed in separate cages
for one week before operation, and continued on the same diet after
model establishment. Rats were anesthetized with chloral hydrate
(10% chloral hydrate, 40 mg/kg) and placed in the prone position.
Laminectomy was performed at level T9-10 to expose the spinal cord.
In the CON group, laminectomy was performed without SCI; in the
other two groups, SCI was created by the modified Allen method at
level T10, and paralysis of both hind legs was successfully
achieved. The wound was then sterilized and closed. Rats received
intraperitoneal injection of 5 ml saline for rehydration and daily
administration of penicillin (100 mg/kg) for 3 days to prevent
infection. Manual bladder expression was performed twice a day
until re-establishment of the voiding reflex. All operations were
performed by the same surgeon to reduce inter-operator
variation.
Gas chromatography
Blood samples were collected from the caudal vein,
and the rats were killed. A 2-cm long section of the spinal cord at
the injured location was quickly removed before and 12 h after
surgery (n=6). Fatty acids were extracted from serum and the spinal
cord and the content was analyzed according to existing published
methods (39,40). A Perkin-Elmer Clarus 500
chromatographic analyzer (PerkinElmer, Inc., Waltham, MA, USA) was
used for gas chromatographic analysis. The fatty acid composition
was identified by comparing the peak time to that of standard
specimens (purchased from Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) by gas chromatography.
Behavioral assessment
Hindlimb motor function was assessed using the
open-field Basso, Beattie and Bresnahan (BBB) locomotor scoring
behavioral assessment system (41,42)
with scores ranging from 0 to 21. Scoring was performed by two
observers on the first postoperative day and then weekly up to 8
weeks.
Five rats were selected randomly from each group at
week 8 after the operation, anesthetized and placed on a quiet
desktop. The head of the electrodes was inserted into the scalp at
the motor cortex and the end into the quadriceps muscle. The
latency and amplitude of the motor-evoked potential were detected
according to the manufacturer's instructions. Measurements were
performed three times for each rat.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Spinal cords were harvested at 8 weeks after
surgery, and RT-qPCR was performed to determine the relative mRNA
levels of MBP, Galc, GFAP and TUBB3 in the spinal cords containing
the lesion site (n=3). RNA was extracted using TRIzol and cDNA was
synthesized using an RT Reagent kit (Takara Biotechnology Co.,
Ltd., Dalian, China) according to the manufacturer's instructions.
The cDNA concentration was tested using a NanoDrop 2000
spectrophotometer (Thermo Fisher Scientific, Inc., Wilmington, DE,
USA) after reverse transcription. According to the instructions of
the TaKaRa Perfect Real Time Reagent kit (Takara Biotechnology Co.,
Ltd.), qPCRs were performed in 0.2-ml thin-walled reaction tubes
using an ABI Prism 7500 Sequence Detection System (Applied
Biosystems; Thermo Fisher Scientific, Inc., Waltham, MA, USA). The
sequences of the primers (Sangon, Shanghai, China) were listed in
Table II.
| Table II.Sequences of polymerase chain
reaction target genes. |
Table II.
Sequences of polymerase chain
reaction target genes.
| MBP |
5′-GGCAATGGTGGGACTCAAAA-3′ |
5′-GGGACCCGCTCCTTCAAC-3′ |
| Galc |
5′-GAGTCCACAACCATCCTTCTGAG-3′ |
5′-ACACCAGGCTGCTTGAACAC-3′ |
| GFAP |
5′-GCATCGCTTCACACTGCGCC-3′ |
5′-ACACACCGCCAGTCCGAGGA-3′ |
| TUBB3 |
5′-GCCCAAGTGAAGTTGCTTGC-3′ |
5′-TGCCCTGAAGAGCTGGTAG-3′ |
| GAPDH |
5′-CCACTCCTCCACCTTTGAC-3′ |
5′-ACCCTGTTGCTGTAGCCA-3′ |
GAPDH was used as an endogenous control to normalize
expression levels of the target genes between the different groups.
Relative expression of the PCR products was determined using the
ΔΔCq method.
Immunofluorescence analysis
Rats were deeply anesthetized with chloral hydrate
12 h after operation (n=3), and transcardiac perfusion was
performed as previously described (43). A 2-cm-long spinal cord segment
centered on the injured area was removed and placed into 4%
paraformaldehyde overnight, and then samples were cryoprotected in
30% sucrose buffer for cryosectioning. The spinal cords were
sectioned into 5-µm-thick coronal sections with a cryostat and used
for immunofluorescent staining. Each section was washed with
phosphate-buffered saline (PBS) and treated with 0.1% Triton X-100
for 5 min. Sections were then blocked with 10% bovine serum albumin
(BSA) for 1 h, then incubated with primary antibody at 4°C
overnight as follows: MBP (1:150), Galc (1:100), GFAP (1:100) (Cell
Signaling Technology, Inc.). After washing with PBS for 1 h,
sections were incubated with the appropriate fluorescent-labeled
goat-anti rabbit secondary antibody (Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA), and then examined under a fluorescence
microscope (Nikon, Tokyo, Japan). Specimens were collected from
rats killed at 12 h, 1 week and 2 weeks postoperatively and stained
with LC3-II (1:50; Cell Signaling Technology, Inc.), and then
observed under a fluorescence microscope (Nikon).
Western blotting
The fresh spinal cords of rats killed 2 weeks
postoperatively (n=3) were pounded in a mortar to a powder and
centrifuged at 14,000 × g for 15 min at 4°C. The supernatant
was then collected and boiled for 10 min at 100°C. The appropriate
amounts of proteins were loaded onto a 10% sodium dodecyl sulfate
polyacrylamide gel and subjected to electrophoresis (SDS-PAGE),
then electroblotted onto a polyvinylidene fluoride membrane. After
blocking for 1 h at room temperature in a solution of 5% (w/v) skim
milk in Tris-buffered saline containing 0.05% Tween-20, the
membrane was incubated with antibodies against the mTOR-related
proteins p-S6 (1:100), p-Akt (1:50), p-S6K (1:100), or 4EBP1
(1:100) (all from Cell Signaling Technology, Inc.) at 4°C
overnight, then incubated with the appropriate secondary goat
anti-rabbit antibody (Bioworld, Dublin, OH, USA) after washing the
membrane for 1 h at room temperature. Finally, after exposure and
development, the blots were photographed and scanned for
analysis.
Statistical analysis
All quantitative data are presented as mean ±
standard error of the mean, and differences among groups were
considered significant at P<0.05. Statistical analysis was
performed by analysis of variance followed by Bonferroni's post-hoc
test (multiple-comparison tests) among three groups, using the
statistical analysis software SPSS 13.0 (SPSS, Inc., Chicago, IL,
USA).
Results
PUFAs altered the activity of mTORC1
and the expression of autophagy-related genes in vitro
When the same dose of DHA was added to neurons with
decreased p-S6 induced by aa or AA, the p-S6 activities in both the
groups decreased (Fig. 1A). After
adding the same dose of AA or DHA to the two groups starved of aa,
the expression of autophagy protein LC3-II decreased or increased,
respectively. The effect on LC3I was opposite to that of LC3-II and
the effect on P62 was the same as that of LC3-II (Fig. 1B). Addition of AA or DHA to the
neurons provided with aa led to an increase or decrease in p-S6
activity, respectively; neurons given the same dose of DHA and AA
simultaneously showed no significant change in p-S6 activity
compared with those without treatment (Fig. 1C).
Contents and composition of PUFAs in
serum and spinal cord
Gas chromatography was used to analyze the serum and
spinal cord preoperatively and postoperatively to study the effects
of a high-n-3 PUFA diet on the percentage of PUFAs. The results
showed that, preoperatively, the content of n-3 PUFAs increased,
while that of n-6 PUFAs decreased, significantly in both the spinal
cord and serum of the group fed a high-n-3 PUFA diet compared with
those fed a low-n-3 PUFA diet or the control group (P<0.05;
Fig. 2A and B); the same result
was observed postoperatively, (P<0.05; Fig. 2C and D). The percentages of PUFAs
in the serum and spinal cord of each group before operation were
lower than those after operation, but the difference was not
significant. In summary, a high-n-3/n-6 PUFA diet significantly
increased both the n-3 PUFA content and n-3/n-6 PUFA ratio in the
serum and spinal cord of rats in vivo.
Assessment of locomotor behavior
Complete paralysis of the hind legs after surgery
and a BBB score of 0 indicated successful establishment of the SCI
model. BBB scores revealed gradual restoration of movement of rats
in each group by 8 weeks after surgery. Compared with the group fed
a low-n-3 PUFA diet and the control group at each time-point, BBB
scores of the rats in the high-n-3 PUFA diet group were higher, but
the group that received n-3 PUFA lavage had much higher scores
(P<0.05; Fig. 3A). Rats in the
high-n-3 PUFA diet group showed shorter latency times of motor
evoked potential (5.3±0.32 msec) and higher amplitude (60.30±4.72
msec) than those in the low-n-3 PUFA diet group (6.19±0.42 and
33.19±4.49 msec, respectively) or the control group (9.51±0.39 and
31.61±4.22 msec, respectively), and all differences were
statistically significant (P<0.05; Fig. 3B-D). These results indicate that
the high-n-3/n-6 PUFA diet was more effective in promoting
functional recovery.
Changes in tissue repair related
proteins and mRNA expression after operation
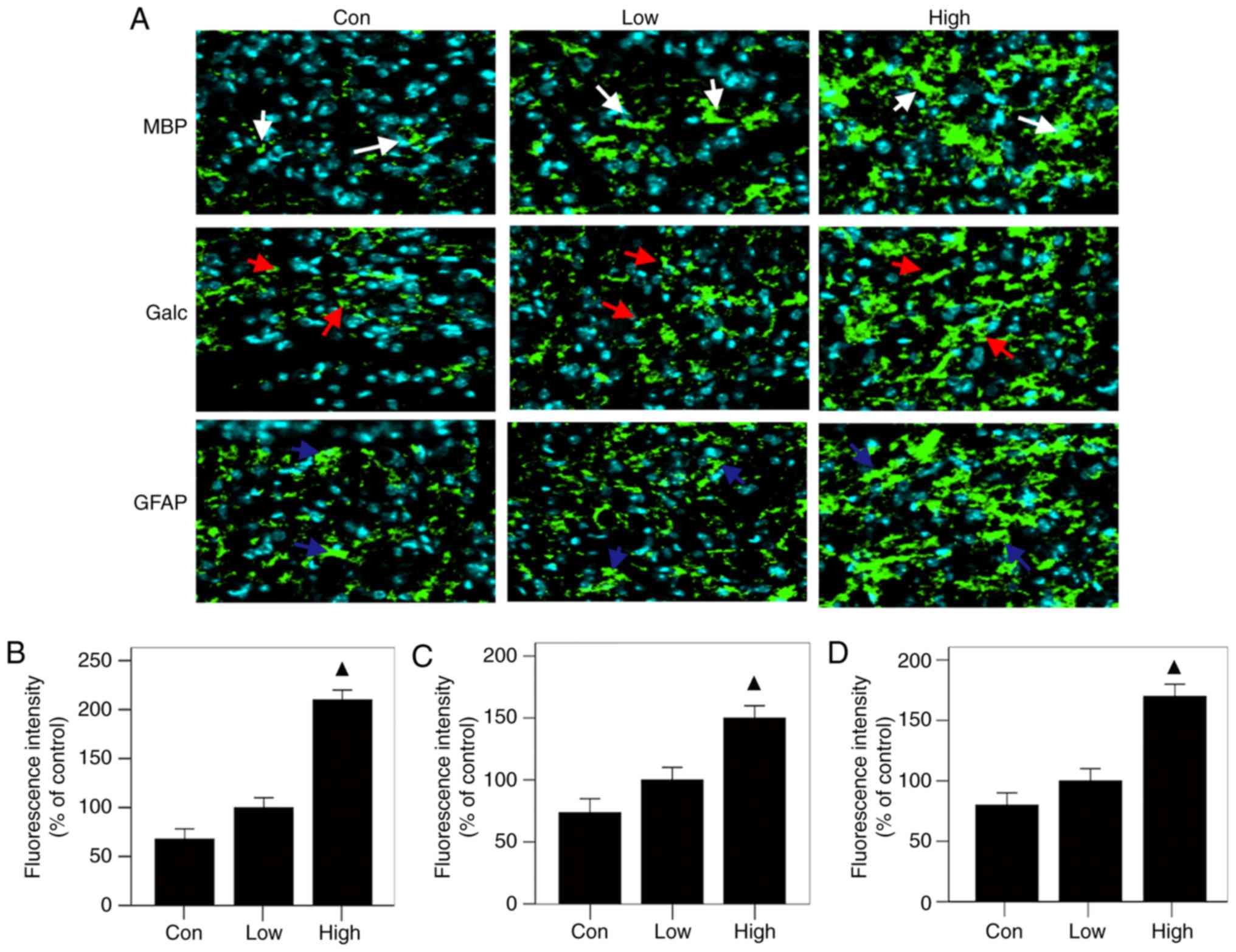
The results of immunofluorescence staining showed
that all three groups had the same patterns of expression of MBP,
Galc and GFAP at the injured sites: the high-n-3 PUFA diet group
expressed the highest levels, followed by the low-n-3 PUFA diet
group and finally the control group (Fig. 4A); the differences were
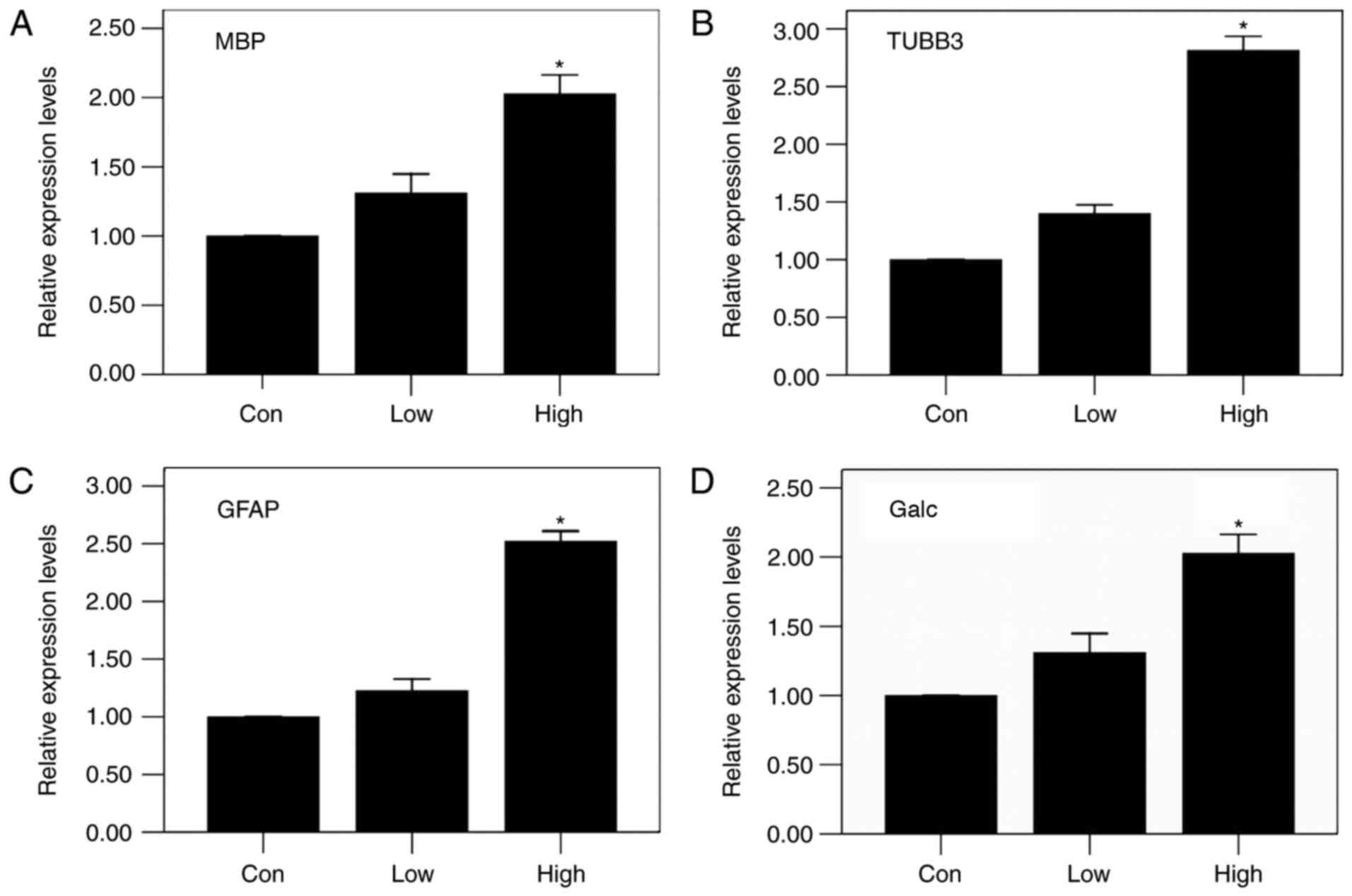
statistically significant (P<0.05) (Fig. 4 B-D). The results of RT-PCR were in
agreement with immunofluorescent staining, showing the same trends,
with statistically significant differences (P<0.05) (Fig. 5). These results indicate that a
diet high in n-3/n-6 PUFAs can significantly increase MBP, Galc,
GFAP and TUBB3 mRNA expression in injured spinal cords.
A diet high in n-3PUFAs promoted
autophagy
No significant difference in LC3-II expression was
found among the three groups, as shown by the autophagy protein
test performed 12 h postoperatively (Fig. 6A and B). However, expression of the
LC3-II autophagy protein was significantly higher in the high-n-3
PUFA diet group at 1 and 2 weeks after operation compared to the
low-n-3 PUFA diet group or the control group (P<0.05; Fig. 6A, C and D). Thus, a high n-3/n-6
PUFA diet promoted LC3-II expression in the injured spinal
cords.
A diet high in n-3 PUFAs inhibited
activity of the mTORC1 signaling pathway
The high-n-3 PUFA diet group showed lower activities
of p-Akt (S473), p-S6 (S235/S236), p-S6 (S240/S244), and p-S6K in
the spinal cords at 2 weeks after surgery compared to the low-n-3
PUFA diet group or the control group (Fig. 7A), and comparison of the density of
the bands produced by western blotting showed that the difference
was statistically significant (P<0.05; Fig. 7B). There was no significant
difference in the level of p-Akt (T308) in the spinal cord between
the high-n-3 PUFA diet group and the other two groups (Fig. 7B); however, the high-n-3 PUFA diet
group had significantly higher levels of 4EBP1 activity in the
spinal cord compared with the other two groups (P<0.05; Fig. 7A and B). Thus, a high n-3/n-6 PUFA
diet could inhibit activity of the mTORC1 signaling pathway.
Discussion
In this study, we created a spinal cord contusion
model in SD rats and fed them with a diet high in n-3 PUFAs to
increase n-3 PUFA content as well as the n-3/n-6 PUFA composition;
we investigated the effects of this change on recovery of SCI. As
our results show, a diet high in n-3 PUFAs inhibited the mTORC1
signaling pathway, increased expression of autophagy proteins,
provided energy for regeneration of the injured spinal cord and
promoted the recovery of hind limb motor function.
Recently, a series of studies has demonstrated that
n-3 PUFAs play crucial roles in tissue repair mechanisms (44–46),
so the study of the influence of n-3 PUFAs on physiological and
pathological processes in the human body is very important
(47–49). Traditional oral administration can
change the content of n-3 PUFAs and the proportions of n-3/n-6
PUFAs in the body; such administration has been used widely in the
clinic and is much safer than transgenic technology (13,15).
Consequently, we supplied rats with n-3 PUFAs in their diet to try
to reproduce drug delivery in the clinic through the enteral route
and thus provide reference data for future clinical studies.
Better understanding of the effect of n-3 PUFAs on
the mTORC1 signaling pathway in tissue repair will help greatly in
increasing understanding of the repair of SCI and provide
information which could lead to better treatments. A recent study
reported that a diet high in n-3 PUFAs reduced expression of p-S6
and p-S6K, suggesting an inhibitory effect of n-3 PUFAs on the
mTORC1 pathway (50). It has also
been reported that both exogenous and endogenous n-3 PUFAs can
target mTOR to inhibit the mTORC1/2 signaling pathway and the
downstream proteins (51). Our
western blot results showed that expression of p-S6K, p-Akt, and
4-EBP1 in the injured spinal cord of rats fed a high-n-3 PUFA diet
were significantly reduced by 8 weeks after injury, consistent with
the above findings. As a result, we concluded that a diet high in
n-3 PUFAs can inhibit activity of the mTOR signaling pathway in
SCI.
After SCI, the local blood supply, and hence the
supply of energy needed for physiological and pathological
activities, is damaged due to local vascular system damage and
regeneration of the injured spinal cord is difficult. Many
investigators have carried out preliminary research and found that
autophagy is significantly activated in the days just after SCI in
a contusion model, peaking within a week postoperatively and then
decreasing to levels close to normal after 2 weeks (38,52–55).
In this experiment, expression of autophagy markers showed a trend
consistent with reports in the existing literature, and mTORC1
activity in the injured spinal cord of rats fed a high-n-3 PUFA
diet was obviously reduced (Fig.
4), while LC3-II protein expression was enhanced. Increased
cell autophagy activity removes damaged tissue and provides a large
amount of macromolecular material and energy for the repair of
local injured tissue, thus promoting regeneration of the injured
spinal cord and restoration of motor function after SCI in
rats.
In conclusion, this study demonstrates that a
high-n-3 PUFA diet downregulates the activity of the mTORC1
signaling pathway, improves autophagy capability, and provides
energy to promote repair of the injured spinal cord and restoration
of motor function in a rat model of SCI. Thus, it can be seen that
changes of n-3 PUFA content and n-3/n-6 PUFA ratio play an
important role in motor function recovery of SCI rats. These
results provide important reference data for the potential
treatment of SCI by n-3 PUFAs.
Acknowledgements
The present study was supported by National Natural
Sciences Foundation of China (grant no. 81560213), Chongqing Three
Gorges Central Hospital PhD Research Startup Fund (grant no.
2017BSKYQDJJ02), Chongqing Postdoctoral Research Special Fund
(grant no. Xm2017181), and North Sichuan Medical College and
Chongqing Three Gorges Central Hospital Cooperation Fund (grant no.
2016XY01).
References
|
1
|
Schwab ME: Regenerative nerve fiber in the
adult central nervous system. News Physiol Sci. 13:294–298.
1998.PubMed/NCBI
|
|
2
|
Fry EJ: Central nervous system
regeneration: Mission impossible? Clin Exp Pharmacol Physiol.
28:253–258. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ritfeld GJ, Nandoe Tewarie RD, Vajn K,
Rahiem ST, Hurtado A, Wendell DF, Roos RA and Oudega M: Bone marrow
stromal cell-mediated tissue sparing enhances functional repair
after spinal cord contusion in adult rats. Cell Transplant.
21:1561–1575. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ribatti D, Conconi MT, Nico B, Baiguera S,
Corsi P, Parnigotto PP and Nussdorfer GG: Angiogenic response
induced by acellular brain scaffolds grafted onto the chick embryo
chorioallantoic membrane. Brain Res. 989:9–15. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Johnson PJ, Tatara A, Shiu A and
Sakiyama-Elbert SE: Controlled release of neurotrophin-3 and
platelet-derived growth factor from fibrin scaffolds containing
neural progenitor cells enhances survival and differentiation into
neurons in a subacute model of SCI. Cell Transplant. 19:89–101.
2011. View Article : Google Scholar
|
|
6
|
Zurita M, Vaquero J, Oya S and Miguel M:
Schwann cells induce neuronal differentiation of bone marrow
stromal cells. Neuroreport. 16:505–508. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu GM, Luo YG, Li J and Xu K: Knockdown
of Nogo gene by short hairpin RNA interference promotes functional
recovery of spinal cord injury in a rat model. Mol Med Rep.
13:4431–4436. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guo J and Li Y, Chen Z, He Z, Zhang B and
Li Y, Hu J, Han M, Xu Y and Li Y: N-acetylcysteine treatment
following spinal cord trauma reduces neural tissue damage and
improves locomotor function in mice. Mol Med Rep. 12:37–44. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen KB, Uchida K, Nakajima H, Yayama T,
Hirai T, Watanabe S, Guerrero AR, Kobayashi S, Ma WY, Liu SY and
Baba H: Tumor necrosis factor-α antagonist reduces apoptosis of
neurons and oligodendroglia in rat spinal cord injury. Spine (Phila
Pa 1976). 36:1350–1358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Figueroa JD and De Leon M:
Neurorestorative targets of dietary long-chain omega-3 fatty acids
in neurological injury. Mol Neurobiol. 50:197–213. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zeman M, Jirak R, Vecka M, Raboch J and
Zak A: N-3 polyunsaturated fatty acids in psychiatric diseases:
Mechanisms and clinical data. Neuro Endocrinol Lett. 33:736–748.
2012.PubMed/NCBI
|
|
12
|
Hasadsri L, Wang BH, Lee JV, Erdman JW,
Llano DA, Barbey AK, Wszalek T, Sharrock MF and Wang HJ: Omega-3
fatty acids as a putative treatment for traumatic brain injury. J
Neurotrauma. 30:897–906. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang MJ, Wang L, Jin DD, Zhang ZM, Chen
TY, Jia CH, Wang Y, Zhen XC, Huang B, Yan B, et al: Enhancement of
the synthesis of n-3 PUFAs in fat-1 transgenic mice inhibits mTORC1
signalling and delays surgically induced osteoarthritis in
comparison with wild-type mice. Ann Rheum Dis. 73:1719–1727. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lim SN, Gladman SJ, Dyall SC, Patel U,
Virani N, Kang JX, Priestley JV and Michael-Titus AT: Transgenic
mice with high endogenous omega-3 fatty acids are protected from
spinal cord injury. Neurobiol Dis. 51:104–112. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wen ZH, Su YC, Lai PL, Zhang Y, Xu YF,
Zhao A, Yao GY, Jia CH, Lin J, Xu S, et al: Critical role of
arachidonic acid activated mTOR signaling in breast carcinogenesis
and angiogenesis. Oncogene. 32:160–170. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gerber M: Omega-3 fatty acids and cancers:
A systematic update review of epidemiological studies. Br J Nutr.
107 Suppl 2:S228–S239. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fleming JC, Norenberg MD, Ramsay DA,
Dekaban GA, Marcillo AE, Saenz AD, Pasquale-Styles M, Dietrich WD
and Weaver LC: The cellular inflammatory response in human spinal
cords after injury. Brain. 129:3249–3269. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Donnelly DJ and Popovich PG: Inflammation
and its role in neuroprotection, axonal regeneration and functional
recovery after spinal cord injury. Exp Neurol. 209:378–388. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Daniel J and Klionsky: The molecular
machinery of autophagy: unanswered questions. J Cell Sci. 118:7–18.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kanki T and Klionsky DJ: The molecular
mechanism of mitochondria autophagy in yeast. Mol Microbiol.
75:795–800. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kanno H, Ozawa H, Sekiguchi A, Yamaya S,
Tateda S, Yahata K and Itoi E: The role of mTOR signaling pathway
in spinal cord injury. Cell Cycle. 11:3175–3179. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu K, Lu Y, Lee JK, Samara R, Willenberg
R, Sears-Kraxberger I, Tedeschi A, Park KK, Jin D, Cai B, et al:
PTEN deletion enhances the regenerative ability of adult
corticospinal neurons. Nat Neurosci. 13:1075–10781. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lamming DW and Sabatini DM: A central role
for mTOR in lipid homeostasis. Cell Metab. 18:465–469. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Laplante M and Sabatini DM: An emerging
role of mTOR in lipid biosynthesis. Curr Biol. 19:R1046–R1052.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Laplante M and Sabatini DM: mTOR signaling
at a glance. J Cell Sci. 122:3589–3594. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Laplante M and Sabatini DM: mTOR
signaling. Cold Spring Harb Perspect Biol. 4(pii):
a0115932012.PubMed/NCBI
|
|
27
|
Su M, Guan H, Zhang F, Gao Y, Teng X and
Yang W: HDAC6 regulates the chaperone-mediated autophagy to prevent
oxidative damage in injured neurons after experimental spinal cord
injury. Oxid Med Cell Longev. 2016:72637362016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou KL, Zhou YF, Wu K, Tian NF, Wu YS,
Wang YL, Chen DH, Zhou B, Wang XY, Xu HZ and Zhang XL: Stimulation
of autophagy promotes functional recovery in diabetic rats with
spinal cord injury. Sci Rep. 5:171302015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou Y, Zheng B, Ye L, Zhang H, Zhu S,
Zheng X, Xia Q, He Z, Wang Q, Xiao J and Xu H: Retinoic acid
prevents disruption of blood-spinal cord barrier by inducing
autophagic flux after spinal cord injury. Neurochem Res.
41:813–825. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gao K, Wang G, Wang Y, Han D, Bi J, Yuan
Y, Yao T, Wan Z, Li H and Mei X: Neuroprotective effect of
simvastatin via inducing the autophagy on spinal cord injury in the
rat model. Biomed Res Int. 2015:2601612015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guo Y, Liu S, Zhang X, Wang L, Gao J, Han
A and Hao A: G-CSF promotes autophagy and reduces neural tissue
damage after spinal cord injury in mice. Lab Invest. 95:1439–1449.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lotz MK and Caramés B: Autophagy and
cartilage homeostasis mechanisms in joint health, aging and OA. Nat
Rev Rheumatol. 7:579–587. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kanno H, Ozawa H, Sekiguchi A, Yamaya S
and Itoi E: Induction of autophagy and autophagic cell death in
damaged neural tissue after acute spinal cord injury in mice. Spine
(Phila Pa 1976). 36:E1427–E1434. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Basso DM, Beattie MS and Bresnahan JC: A
sensitive and reliable locomotor rating scale for open field
testing in rats. J Neurotrauma. 12:1–21. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang XY, Xue H, Liu JM and Chen D:
Chemically extracted acellular muscle: A new potential scaffold for
spinal cord injury repair. J Biomed Mater Res A. 100:578–587. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wei D, Li J, Shen M, Jia W, Chen N, Chen
T, Su D, Tian H, Zheng S, Dai Y and Zhao A: Cellular production of
n-3 PUFAs and reduction of n-6-to-n-3 ratios in the pancreatic
beta-cells and islets enhance insulin secretion and confer
protection against cytokine-induced cell death. Diabetes.
59:471–478. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kang JX: A transgenic mouse model for
gene-nutrient interactions. J Nutrigenet Nutrigenomics. 1:172–177.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nakamura M, Houghtling RA, MacArthur L,
Bayer BM and Bregman BS: Differences in cytokine gene expression
profile between acute and secondary injury in adult rat spinal
cord. Exp Neurol. 184:313–325. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Manku MS, Horrobin DF, Huang YS and Morse
N: Fatty acids in plasma and red cell membranes in normal humans.
Lipids. 18:906–908. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dyall SC, Michael GJ, Whelpton R, Scott AG
and Michael-Titus AT: Dietary enrichment with omega-3
polyunsaturated fatty acids reverses age-related decreases in the
GluR2 and NR2B glutamate receptor subunits in rat forebrain.
Neurobiol Aging. 28:424–439. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jiang W, Zhu Z, McGinley JN, El Bayoumy K,
Manni A and Thompson HJ: Identification of a molecular signature
underlying inhibition of mammary carcinoma growth by dietary n-3
fatty acids. Cancer Res. 72:3795–3806. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen Z, Zhang Y, Jia C, Wang Y, Lai P,
Zhou X, Wang Y, Song Q, Lin J, Ren Z, et al: mTORC1/2 targeted by
n-3 polyunsaturated fatty acids in the prevention of mammary
tumorigenesis and tumor progression. Oncogene. 33:4548–4557. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Berquin IM, Edwards IJ and Chen YQ:
Multi-targeted therapy of cancer by omega-3 fatty acids. Cancer
Lett. 269:363–377. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Larsson SC, Kumlin M, Ingelman-Sundberg M
and Wolk A: Dietary long-chain n-3 fatty acids for the prevention
of cancer: A review of potential mechanisms. Am J Clin Nutr.
79:935–945. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Streit WJ, Semple-Rowland SL, Hurley SD,
Miller RC, Popovich PG and Stokes BT: Cytokine mRNA profiles in
contused spinal cord and axotomized facial nucleus suggest a
beneficial role for inflammation and gliosis. Exp Neurol.
152:74–87. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bracken MB, Shepard MJ, Collins WF,
Holford TR, Young W, Baskin DS, Eisenberg HM, Flamm E, Leo-Summers
L, Maroon J, et al: A randomized, controlled trial of
methylprednisolone or naloxone in the treatment of acute
spinal-cord injury. Results of the second national acute spinal
cord injury study. N Engl J Med. 322:1405–1411. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang CX, Olschowka JA and Wrathall JR:
Increase of interleukin-1beta mRNA and protein in the spinal cord
following experimental traumatic injury in the rat. Brain Res.
13:190–196. 1997. View Article : Google Scholar
|
|
48
|
Lu Y, Jiang BC, Cao DL, Zhang ZJ, Zhang X,
Ji RR and Gao YJ: TRAF6 upregulation in spinal astrocytes maintains
neuropathic pain by integrating TNF-α and IL-1β signaling. Pain.
155:2618–2629. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Grau JW, Huie JR, Lee KH, Hoy KC, Huang
YJ, Turtle JD, Strain MM, Baumbauer KM, Miranda RM, Hook MA, et al:
Metaplasticity and behavior: How training and inflammation affect
plastic potential within the spinal cord and recovery after injury.
Front Neural Circuits. 8:1002014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Erlich S, Alexandrovich A, Shohami E and
Pinkas-Kramarski R: Rapamycin is a neuroprotective treatment for
traumatic brain injury. Neurobiol Dis. 26:86–93. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hou H, Zhang L, Zhang L and Tang P: Acute
spinal cord injury in rats should target activated autophagy. J
Neurosurg Spine. 20:568–577. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hao HH, Wang L, Guo ZJ, Bai L, Zhang RP,
Shuang WB, Jia YJ, Wang J, Li XY and Liu Q: Valproic acid reduces
autophagy and promotes functional recovery after spinal cord injury
in rats. Neurosci Bull. 29:484–492. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wang ZY, Lin JH, Muharram A and Liu WG:
Beclin-1-mediated autophagy protects spinal cord neurons against
mechanical injury-induced apoptosis. Apoptosis. 19:933–945. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Serhan CN, Dalli J, Colas RA, Winkler JW
and Chiang N: Protectins and maresins: New pro-resolving families
of mediators in acute inflammation and resolution bioactive
metabolome. Biochim Biophys Acta. 1851:397–413. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen Z, Fu Q, Shen B, Huang X, Wang K, He
P, Li F, Zhang F and Shen H: Enhanced p62 expression triggers
concomitant autophagy and apoptosis in a rat chronic spinal cord
compression model. Mol Med Rep. 9:2091–2096. 2014. View Article : Google Scholar : PubMed/NCBI
|