Introduction
Gallbladder cancer (GBC) is a rare, aggressive
biliary tract malignancy, of which only ~20% of cases diagnosed in
the early stages (1). The majority
of patients with GBC present at an advanced stage, which is
associated with a poor prognosis and poor survival outcomes for
patients, with a 5-year survival rate ranging between 20 and 40%
(2). Although considerable
progress has been made in elucidating the genetic mechanisms
involved in GBC (3,4), the molecular pathogenesis of GBC
remains to be fully elucidated. Therefore, investigating novel
molecules involved in the progression of GBC is like to facilitate
understanding of the molecular mechanism underlying this disease
and facilitate the development of novel therapeutic strategies.
Long non-coding RNAs (lncRNAs) are >200
nucleotides in length and the extent or levels of expression of
lncRNAs generally appear to be specific to the tissues (5). Increasing evidence has demonstrated
that lncRNAs are important regulators of diverse cellular processes
and are emerging as being important in cancer (6). They have the potential to serve as
therapeutic targets and prognostic indicators. For example, Wang
et al (7) showed that the
expression level of lncRNA-regulator of reprogramming (ROR) was
upregulated in tissues from patients with GBC, and that the
overexpression of lncRNA-ROR promoted the proliferation, migration
and invasion of tumor cells, which was significantly associated
with a poor outcome. Another study analyzed the expression of
lncRNA actin filament associated protein 1 antisense RNA1
(AFAP1-AS1) using quantitative polymerase chain reaction analysis
in GBC tissues and produced survival plots, which demonstrated that
lncRNA AFAP1-AS1 was correlated with a poor prognosis in patients
with GBC (8). Additionally, a
competitive endogenous RNA (ceRNA) hypothesis has been suggested,
in which lncRNAs are involved in cancer progression via interacting
with microRNAs (miRNAs) (9,10).
For example, a previous study reported that lncRNA Gall bladder
cancer associated suppressor of pyruvate carboxylase, a target of
miRNA (miR)-17-3p, negatively regulates pyruvate
carboxylase-dependent cell proliferation in GBC (11). In addition, the findings of Wang
et al (12) suggested that
the lncRNA H19 may regulate the expression of forkhead box M1
(FOXM1) by competitively binding endogenous miR-342-3p in GBC.
Although progress has been made in understanding the roles of
lncRNAs in the progression of GBC, the molecular mechanisms
underlying of lncRNAs require detailed investigations.
In the present study, a set of bioinformatics
approaches were used to comprehensively analyze the publicly
available microarray data from the Gene Expression Omnibus (GEO)
database, which included five separate GBC tissue samples and fived
matched adjacent gallbladder normal tissue samples. The
differentially expressed lncRNAs and mRNAs were first identified in
separate GBC tissues, and compared with those of matched adjacent
normal gallbladder tissues. Subsequently, co-expressed lncRNA-mRNA
pairs were identified, followed by functional enrichment analysis
for these mRNAs. The shared upstream miRNAs regulating the
co-expressed lncRNA and mRNAs were also predicted. The present
study is aimed to examine key lncRNAs potentially involved in the
progression of GBC and to elucidate their molecular mechanisms in
the development of GBC.
Materials and methods
Affymetrix microarray data
The microarray data of GSE62335 was downloaded from
the GEO database (http://www.ncbi.nlm.nih.gov/geo/), which was deposited
by Ma et al (13) on 15th
October, 2014. In total, 10 samples were used to develop the array
data, which included five separate GBC tissues and five matched
adjacent normal gallbladder tissues. The raw data and annotation
files were downloaded based on the platform of the GPL16686
Affymetrix Human Gene 2.0 ST Array (Affymetrix Inc., Santa Clara,
CA, USA) for further analysis.
Data preprocessing and screening of
differentially expressed lncRNAs and mRNAs
All the raw data were preprocessed using the robust
multichip average method in the oligo package (available through
Bioconductor version 3.0; http://www.bioconductor.org) (14). The comparison of differentially
expressed lncRNAs and mRNAs in separate GBC tissues with normal
tissues were screened using the Limma package version 3.22.7 of
Bioconductor version 3.0 (http://www.bioconductor.org/packages/release/bioc/html/limma.html)
(15). P<0.05 and
|log2 fold-change (FC)|>0.58 were defined as the
cut-off values for screening.
RNA binding protein (RBP)
analysis
The starBase v2.0 database (http://starbase.sysu.edu.cn/) (16,17)
is an experimentally supported database, which provides the most
comprehensive protein-RNA, miRNA-mRNA and miRNA-lncRNA interactions
supported by large-scale cross-linking immunoprecipitation
(CLIP)-Seq (HITS-CLIP, PAR-CLIP, iCLIP and CLASH) data. The newly
developed starBase v2.0 database has been used in several studies
to identify between miRNAs and lncRNAs, which have also been
validated by experiments (18,19).
In the present study, RBPs directly interacting with differentially
expressed lncRNAs and mRNAs were identified based on the
information in starBase v2.0. The lncRNAs and mRNAs, which were
able to bind to the same RBP, were then selected for subsequent
analysis.
Co-expression network construction and
functional enrichment analysis
The Pearson correlation coefficient (20,21)
is widely used as a co-expression similarity measure. In the
present study, the lncRNAs and mRNAs found to interact with the
same RBP were selected for co-expression analysis, and the
co-expressed lncRNA-mRNA pairs were screened out with the absolute
value of a Pearson correlation coefficient >0.85. The
co-expression network was then constructed with the significant
lncRNA-mRNAs using Cytoscape software version 3.0.2 (http://www.cytoscape.org/) (22).
The Gene Ontology (GO) database (http://www. geneontology.org/) is widely used for
the unification of biological functions and provides gene
annotation terms for large-scale genomic or transcriptomic data
(23,24). The Database for Annotation
Visualization and Integrated Discovery (DAVID; http://david.abcc.ncifcrf.gov/) (25) is used for systematically
associating the functional terms with gene or protein lists. The
DAVID Bioinformatics Resources consist of the DAVID Knowledgebase
and a set of integrated, web-based functional annotation tool
suites, which can be used to screen large gene lists in detail by
comprehensively integrating several different biological angles to
extract associated biological meanings (26). In the present study, the DAVID
online tool was used for GO enrichment analysis of mRNAs in the
co-expressed lncRNA-mRNA pairs identified. P<0.01 was considered
the cut-off value.
Upstream miRNA analysis
Based on the information obtained on the
miRNA-lncRNA and miRNA-mRNA pairs in starBase v2.0, the upstream
miRNAs of lncRNAs and mRNAs were respectively predicted. The shared
upstream miRNAs of the co-expressed lncRNAs and mRNAs were then
identified according to the their expression levels.
Results
Screening of differentially expressed
lncRNAs and mRNAs
Using the Limma package with P<0.05 and
|log2 FC|>0.58 as cut-off thresholds, a total of 89
upregulated transcripts (13 lncRNAs and 76 mRNAs) and 261
downregulated transcripts (27 lncRNAs and 234 mRNAs) were
identified in the separate GBC tissues, compared with the normal
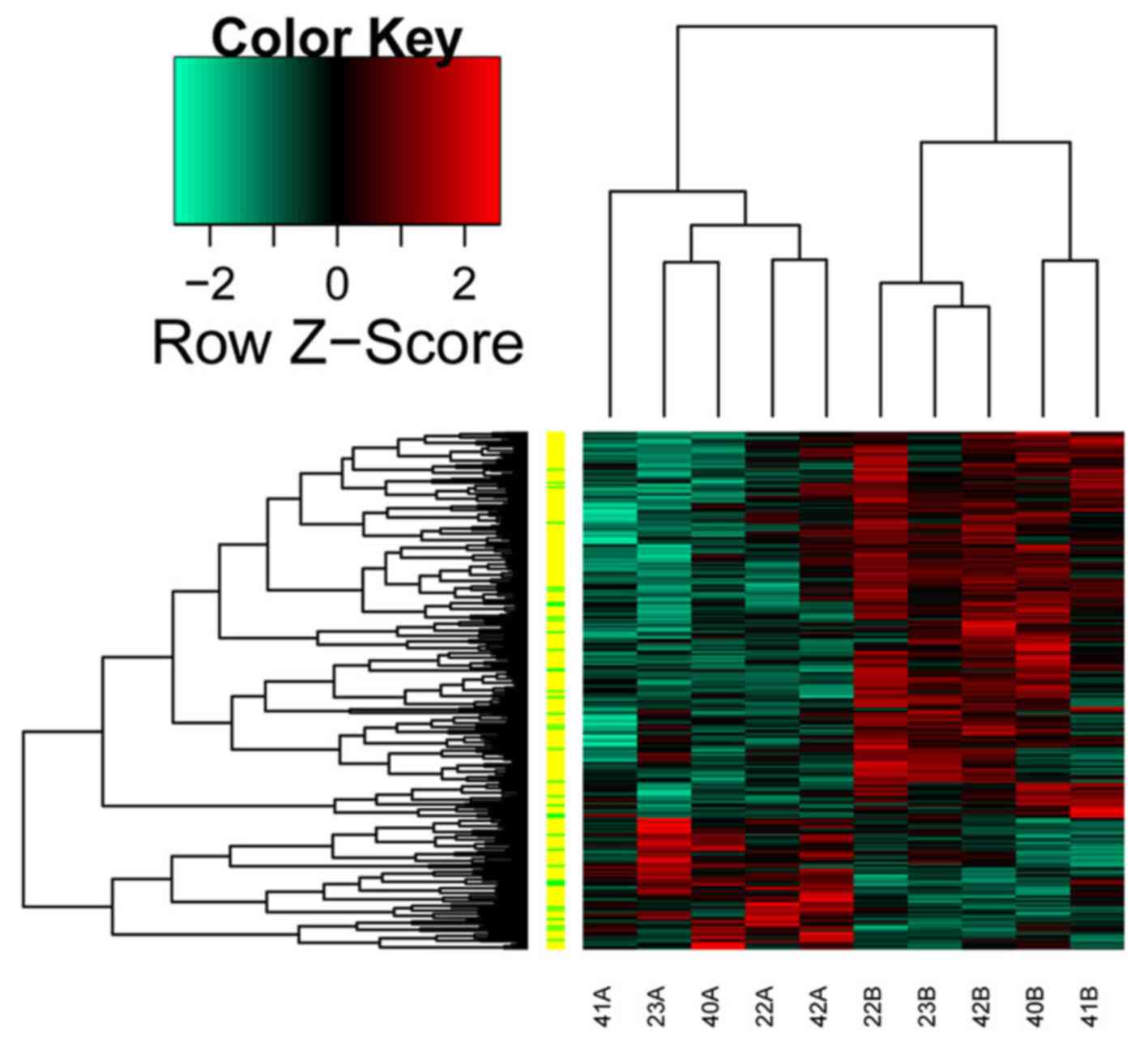
tissues. The heatmap of the differentially expressed transcripts is
shown in Fig. 1.
RBP analysis
A total of 2,759 RBP-RNA pairs were initially
identified according to the information of starBase v2.0, among
which 1,747 were screened out as they were able to interact with
not only lncRNAs, but also mRNAs.
Co-expression and functional
enrichment analysis
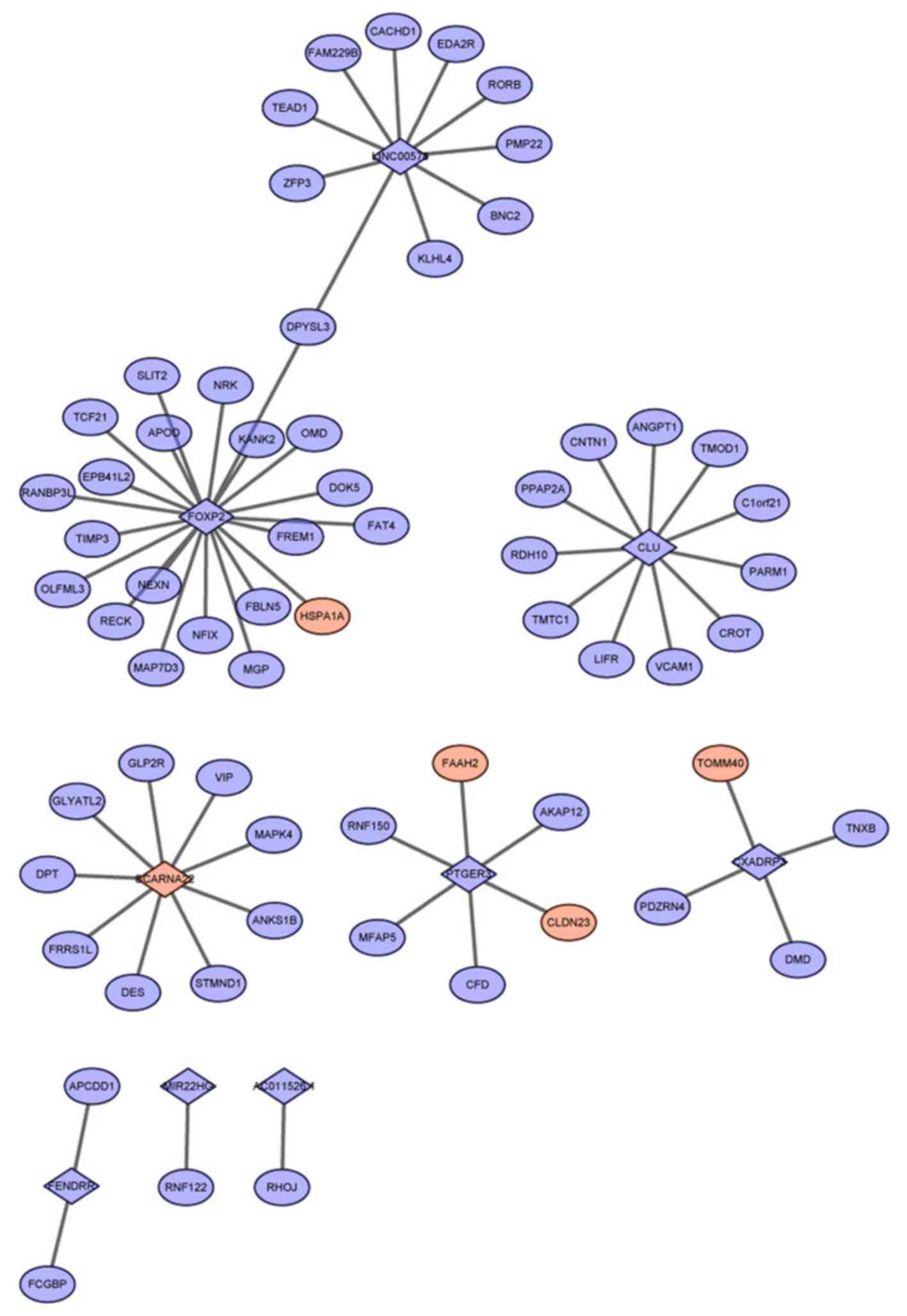
In the present study, a total of 287 co-expressed
lncRNA-mRNA pairs were screened with the absolute Pearson
correlation coefficient value of >0.85. Of these co-expressed
lncRNA-mRNA pairs, 65 were involved in the 1,747 identified RBP-RNA
pairs, from which a co-expression network was constructed (Fig. 2). Notably, all mRNAs within the 65
lncRNA-mRNA pairs were linked with only nine lncRNAs, including
FOXP2, clusterin (CLU) and coxsackie virus and adenovirus receptor
pseudogene 3 (CXADRP3), with co-expressed mRNAs (Table I). FOXP2 was also able to
co-express with the highest number of mRNAs, followed by CLU.
 | Table I.Number of co-expressed mRNAs
corresponding to lncRNAs. |
Table I.
Number of co-expressed mRNAs
corresponding to lncRNAs.
| LncRNA | Co-expressed mRNAs
(n) |
|---|
| AC011526.1 | 1 |
| CLU | 11 |
| CXADRP3 | 4 |
| FENDRR | 2 |
| FOXP2 | 21 |
| LINC00578 | 10 |
| MIR22HG | 1 |
| PTGER3 | 6 |
| SCARNA22 | 9 |
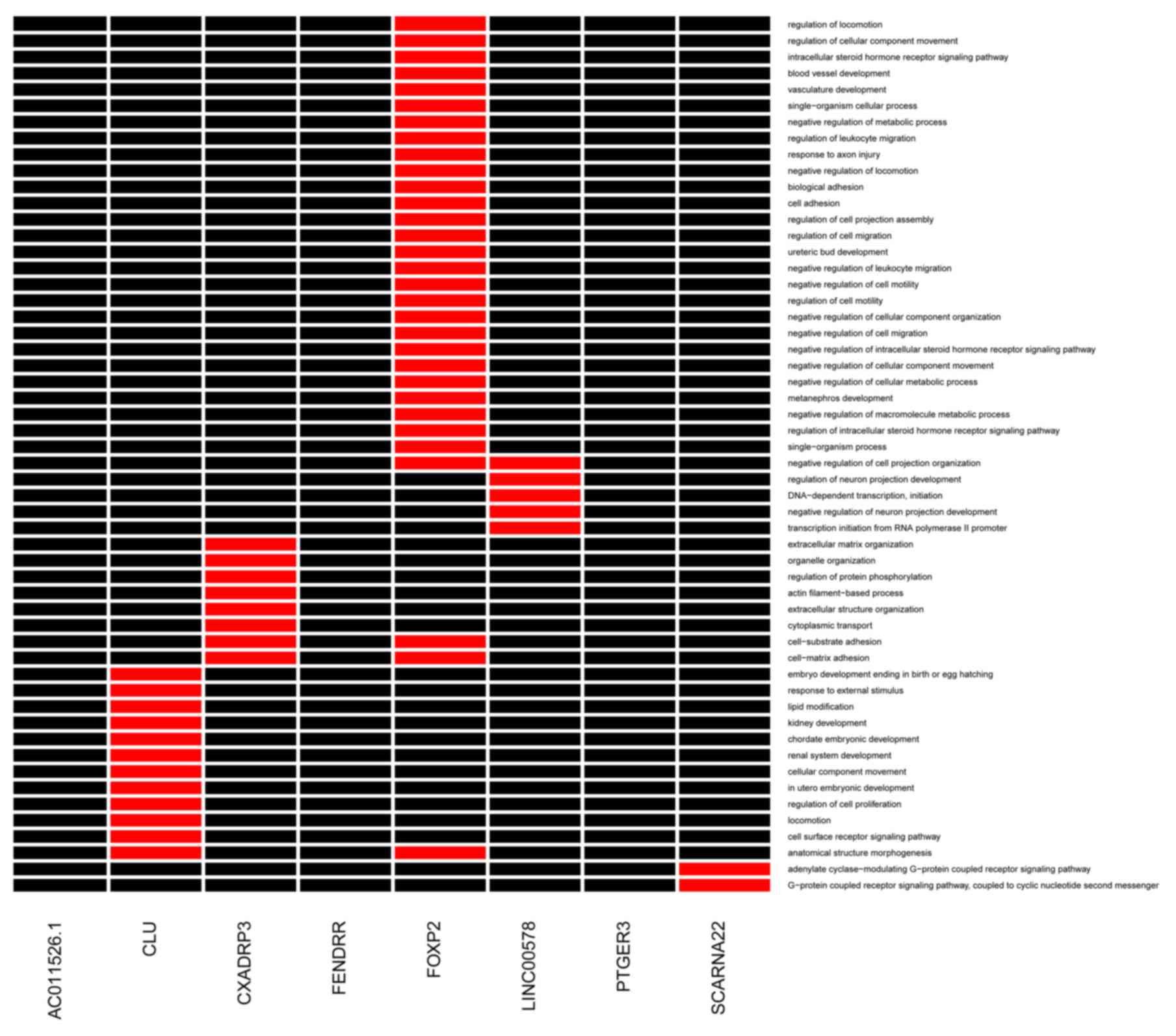
As shown in Fig. 3,
the co-expressed mRNAs connected with FOXP2 and CXADRP3 were
significantly enriched in several GO terms, including cell-matrix
adhesion, cell-substrate adhesion and cell adhesion. The
co-expressed mRNAs, which had interactions with CLU were
significantly associated with the regulation of cell proliferation
and cell surface receptor signaling pathway terms.
Upstream miRNA analysis
According to the information from starBase v2.0,
only lncRNA FOXF1 adjacent non-coding developmental regulatory RNA
(FENDRR) was able to interact with upstream miRNAs. FENDRR shared
common upstream miRNAs with 119 differentially expressed mRNAs.
However, based on the correlation information in the starBase
version 2.0 database, not all 119 differentially expressed mRNAs
exhibited a correlation with FENDRR; thus, it was not possible to
construct the ceRNA interaction network with these 119
differentially expressed mRNAs, FENDRR and the shared upstream
miRNAs.
However, FENDRR was revealed to be co-expressed with
adenomatosis polyposis coli downregulated 1 (APCDD1; correlation
coefficient, 0.905) and v-kit Hardy-Zuckerman 4 feline sarcoma
viral oncogene homolog (KIT; correlation coefficient, 0.846).
Notably, upstream miRNA analysis showed that miR-18b-5p, miR-28-5p
and miR-4735-3p were common upstream miRNAs of FENDRR and KIT,
whereas miR-18b-5p was the common upstream miRNA of FENDRR and
APCDD1.
Discussion
In the present study, a comprehensive biological
informatics approach was used to examine key lncRNAs involved in
GBC and to elucidate their molecular mechanisms in the development
of GBC. In total, 13 upregulated and 27 downregulated lncRNAs were
identified in the separate GBC tissues, compared with the normal
tissues. Among these lncRNAs, FOXP2 was found to be downregulated
in the GBC tissues and co-expressed with the highest number of
differentially expressed mRNAs, which were significantly enriched
in GO terms associated with cell adhesion. FENDRR was also a
downregulated lncRNA, which was found to share common upstream
miRNAs with 119 differentially expressed mRNAs, including
miR-18b-5p, and was co-expressed with KIT and APCDD1. These results
suggested that lncRNA FOXP2 and FENDRR were likely to be important
in the pathogenesis of GBC via cell adhesion and regulating
miR-18b-5p, or the interaction with KIT and APCDD1,
respectively.
Cell adhesion is an important event in biological
processes, which determines the polarity of cells and is the
morphological hallmark of malignant tumors (27). Cell-cell adhesiveness is usually
reduced in several types of cancer, which is considered to be
essential for cancer invasion and metastasis (27). Increasing evidence has highlighted
the roles of cell adhesion molecules in perineural invasion in GBC
(28). The overexpression of
epithelial cell adhesion molecule antigen is an independent
prognostic marker in GBC (29). In
addition, E-cadherin is a transmembrane cell-adhesion protein
mediating intercellular adhesion in epithelial tissues (30). It is reported that loss of the
expression of E-cadherin is important in the progression from tumor
formation to invasion and metastasis in several types of cancer
(31). Kohya et al
(32) confirmed that E-cadherin
has a critical role in the proliferation, motility and invasion of
GBC cells. Therefore, cell adhesion may be an important biological
process involved in GBC. By contrast, lncRNAs have been shown to
regulate gene expression through several mechanisms (33). LncRNAs have also been shown to
allosterically modify RBPs, which regulate transcription (33). In the present study, lncRNA FOXP2
was found to be downregulated in separate GBC tissues, compared
with that in matched adjacent normal gallbladder tissues, and the
majority of mRNAs co-expressed with lncRNA FOXP2 were significantly
associated with the function of cell adhesion. Therefore, it was
hypothesized that lncRNA FOXP2 may be crucial in the progression of
GBC via co-expression with important molecules in cell
adhesion.
KIT is a member of the class III receptor tyrosine
kinase family, which can be bound to and activated by stem cell
factor (34). KIT receptor
activation can cause the phosphorylation and activation of the Shc
adaptor protein (35) and Ras
(36). The Shc adaptor protein can
link c-KIT to the Ras/mitogen-protein kinase (MAPK) pathway
(37). K-ras gene mutation is
frequently found in GBC cells (38). Activated Ras induces
gefitinib-resistance by activating the epidermal growth factor
receptor signaling pathways in human GBC cells (39). In addition, KIT be crucial in the
majority of gastrointestinal stromal tumors via activation of the
phosphatidyl-inositol-3-kinase (PI3-K)/AKT survival pathway. The
activated MAPK and AKT pathways can lead to the uncontrolled growth
of GBC epithelium, and promote the invasion and metastasis of GBC
cells (40). By contrast, APCDD1
is a membrane-bound glycoprotein, which can interact in
vitro with WNT3A and low-density lipoprotein receptor-related
protein 5, which are two essential components of the Wnt signaling
pathway (40). A previous study
demonstrated that APCDD1 is the activated downstream target of the
WNT/β-catenin pathway, which may enhance the proliferation of
pediatric Wilms' tumor cells (41). Mutations of components of the
Wnt/β-catenin pathway, including β-catenin exon 3 mutations, have
been found to be associated with the progression of GBC (42). In addition, Wnt signaling can
cross-talk with the E-cadherin cell adhesion system, which is
involved in the morphogenesis of several types of cancer, including
GBC (27,32). In the present study, FENDRR, as a
downregulated lncRNA in GBC, was co-expressed with KIT and APCDD1.
In a previous study, the expression profiles of lncRNAs in
infantile hemangioma were assessed, and significant associations of
FENDRR and its associated mRNAs were identified (43). Based on the results of the present
study, and previous evidence confirming the importance of KIT and
APCDD1 in the progression of GBC, FENDRR may be crucial in the
progression of GBC through interaction with these two molecules by
targeting several signaling pathways, including the Wnt signaling
pathway.
In the present study, the analysis of upstream
miRNAs also showed that miR-18b-5p was a common upstream miRNA of
FENDRR, KIT and APCDD1. miR-18b-5p acts as an oncogene, and has
been reported to be involved in the development and pathogenesis of
human gastric cancer (44). The
expression of miR-18b-5p is associated with the tumor protein 53
status, and may predict clinical outcome in patients with head and
neck squamous cell carcinoma (45). Although the roles of miR-18b-5p in
the development of GBC remain to be fully elucidated, the present
study hypothesized that miR-18b-5p may be involved in the
progression of GBC via targeting KIT and APCDD1, and lncRNA FENDRR
may be involved in the development of GBC via regulating
miR-18b-5p.
However, there were several limitations to the
present study. Firstly, the size of samples was relatively small,
therefore, further investigations with a higher number of samples
are required. Secondly, statistical validation using other
datasets, including meta-analysis (46,47)
may be used to cross-check the results of the present study.
Thirdly, due to insufficient material, experimental validation was
not performed in the present study. The inclusion of these as a
central component of future investigations can assist in confirming
results using different approaches, including quantitative
real-time polymerase chain reaction analysis.
In conclusion, lncRNA FOXP2 and FENDRR may be
crucial in promoting the progression of GBC via cell adhesion and
regulating miR-18b-5p or interaction with KIT and APCDD1,
respectively. Therefore, these two factors may be recommended as
potential targets in the management of GBC. The findings of the
present study shed new light on the molecular mechanism of GBC and
may provide a foundation for the development of credible
therapeutic approaches. However, in the absence of experimental
validation in the present study, further analysis of the microarray
data and genetic experiments are required.
References
|
1
|
Takebe N and Yang SX: Sonic hedgehog
signaling pathway and gallbladder cancer: Targeting with precision
medicine approach. Chin Clin Oncol. 5:12016.PubMed/NCBI
|
|
2
|
Ma M, Weng M, Zhang M, Qin Y, Gong W and
Quan Z: Targeting gallbladder cancer: Hyaluronan sensitizes cancer
cells to chemo-therapeutics. Int J Clin Exp Pathol. 8:1822–1825.
2015.PubMed/NCBI
|
|
3
|
Nagahashi M, Ajioka Y, Lang I, Szentirmay
Z, Kasler M, Nakadaira H, Yokoyama N, Watanabe G, Nishikura K,
Wakai T, et al: Genetic changes of p53, K-ras, and microsatellite
instability in gallbladder carcinoma in high-incidence areas of
Japan and Hungary. World J Gastroenterol. 14:70–75. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ooi A, Suzuki S, Nakazawa K, Itakura J,
Imoto I, Nakamura H and Dobashi Y: Gene amplification of Myc and
its coamplification with ERBB2 and EGFR in gallbladder
adenocarcinoma. Anticancer Res. 29:19–26. 2009.PubMed/NCBI
|
|
5
|
Bhat SA, Ahmad SM, Mumtaz PT, Malik AA,
Dar MA, Urwat U, Shah RA and Ganai NA: Long non-coding RNAs:
Mechanism of action and functional utility. Non-coding RNA Res.
1:43–50. 2016. View Article : Google Scholar
|
|
6
|
Haemmerle M and Gutschner T: Long
non-coding RNAs in cancer and development: Where do we go from
here? Int J Mol Sci. 16:1395–1405. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang SH, Zhang MD, Wu XC, Weng MZ, Zhou D
and Quan ZW: Overexpression of LncRNA-ROR predicts a poor outcome
in gallbladder cancer patients and promotes the tumor cells
proliferation, migration, and invasion. Tumour Biol.
37:12867–12875. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ma F, Wang SH, Cai Q, Zhang MD, Yang Y and
Ding J: Overexpression of LncRNA AFAP1-AS1 predicts poor prognosis
and promotes cells proliferation and invasion in gallbladder
cancer. Biomed Pharmacother. 84:1249–1255. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jalali S, Bhartiya D, Lalwani MK,
Sivasubbu S and Scaria V: Systematic transcriptome wide analysis of
lncRNA-miRNA interactions. PloS One. 8:e538232013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Juan L, Wang G, Radovich M, Schneider BP,
Clare SE, Wang Y and Liu Y: Potential roles of microRNAs in
regulating long intergenic noncoding RNAs. BMC Med Genomics. 6
Suppl 1:S72013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma MZ, Zhang Y, Weng MZ, Wang SH, Hu Y,
Hou ZY, Qin YY, Gong W, Zhang YJ, Kong X, et al: Long noncoding RNA
GCASPC, a target of miR-17-3p, negatively regulates pyruvate
carboxylase-dependent cell proliferation in gallbladder cancer.
Cancer Res. 76:5361–5371. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang SH, Ma F, Tang ZH, Wu XC, Cai Q,
Zhang MD, Weng MZ, Zhou D, Wang JD and Quan ZW: Long non-coding RNA
H19 regulates FOXM1 expression by competitively binding endogenous
miR-342-3p in gallbladder cancer. J Exp Clin Cancer Res.
35:1602016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma MZ, Kong X, Weng MZ, Zhang MD, Qin YY,
Gong W, Zhang WJ and Quan ZW: Long non-coding RNA-LET is a positive
prognostic factor and exhibits tumor-suppressive activity in
gallbladder cancer. Mol Carcinog. 54:1397–1406. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Carvalho BS and Irizarry RA: A framework
for oligonucleotide microarray preprocessing. Bioinformatics.
26:2363–2367. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Smyth GK: Linear models and empirical
bayes methods for assessing differential expression in microarray
experiments. Stat Appl Genet Mol Biol. 3:Article32004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang JH, Li JH, Shao P, Zhou H, Chen YQ
and Qu LH: starBase: A database for exploring microRNA-mRNA
interaction maps from Argonaute CLIP-Seq and Degradome-Seq data.
Nucleic Acids Res. 39:(Database issue). D202–D209. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:(Database issue). D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cui J, Mo J, Luo M, Yu Q, Zhou S, Li T,
Zhang Y and Luo W: c-Myc-activated long non-coding RNA H19
downregulates miR-107 and promotes cell cycle progression of
non-small cell lung cancer. Int J Clin Exp Pathol. 8:12400–12409.
2015.PubMed/NCBI
|
|
19
|
Xiao H, Tang K, Liu P, Chen K, Hu J, Zeng
J, Xiao W, Yu G, Yao W, Zhou H, et al: LncRNA MALAT1 functions as a
competing endogenous RNA to regulate ZEB2 expression by sponging
miR-200s in clear cell kidney carcinoma. Oncotarget. 6:38005–38015.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
de la Fuente A, Bing N, Hoeschele I and
Mendes P: Discovery of meaningful associations in genomic data
using partial correlation coefficients. Bioinformatics.
20:3565–3574. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang J: Pearson Correlation
CoefficientEncyclopedia of Systems Biology. Springer; pp. 1671.
2013, View Article : Google Scholar
|
|
22
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Consortium GO: The gene ontology (GO)
database and informatics resource. Nucleic acids Res. 32:(Database
issue). D258–D261. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang DW, Sherman BT, Tan Q, Collins JR,
Alvord WG, Roayaei J, Stephens R, Baseler MW, Lane HC and Lempicki
RA: The DAVID gene functional classification tool: A novel
biological module-centric algorithm to functionally analyze large
gene lists. Genome Biol. 8:R1832007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang DW, Sherman BT, Tan Q, Kir J, Liu D,
Bryant D, Guo Y, Stephens R, Baseler MW, Lane HC and Lempicki RA:
DAVID bioinformatics resources: Expanded annotation database and
novel algorithms to better extract biology from large gene lists.
Nucleic Acids Res. 35:(Web Server issue). W169–W175. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hirohashi S and Kanai Y: Cell adhesion
system and human cancer morphogenesis. Cancer Sci. 94:575–581.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Seki H, Koyama K, Tanaka JI, Sato Y and
Umezawa A: Neural cell adhesion molecule and perineural invasion in
gallbladder cancer. J Surg Oncol. 58:97–100. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Varga M, Obrist P, Schneeberger S,
Mühlmann G, Felgel-Farnholz C, Fong D, Zitt M, Brunhuber T, Schäfer
G, Gastl G and Spizzo G: Overexpression of epithelial cell adhesion
molecule antigen in gallbladder carcinoma is an independent marker
for poor survival. Clin Cancer Res. 10:3131–3136. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gumbiner BM: Regulation of
cadherin-mediated adhesion in morphogenesis. Nat Rev Mol Cell Biol.
6:622–634. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Katagiri A, Watanabe R and Tomita Y:
E-cadherin expression in renal cell cancer and its significance in
metastasis and survival. Br J Cancer. 71:3761995. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kohya N, Kitajima Y, Jiao W and Miyazaki
K: Effects of E-cadherin transfection on gene expression of a
gallbladder carcinoma cell line: Repression of MTS1/S100A4 gene
expression. Int J Cancer. 104:44–53. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Su Y, Wu H, Pavlosky A, Zou LL, Deng X,
Zhang ZX and Jevnikar AM: Regulatory non-coding RNA: New
instruments in the orchestration of cell death. Cell Death Dis.
7:e23332016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu H, Chen X, Focia PJ and He X:
Structural basis for stem cell factor-KIT signaling and activation
of class III receptor tyrosine kinases. EMBO J. 26:891–901. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cutler RL, Liu L, Damen J and Krystal G:
Multiple cytokines induce the tyrosine phosphorylation of Shc and
its association with Grb2 in hemopoietic cells. J Biol Chem.
268:21463–21465. 1993.PubMed/NCBI
|
|
36
|
Duronio V, Welham MJ, Abraham S, Dryden P
and Schrader JW: p21ras activation via hemopoietin receptors and
c-kit requires tyrosine kinase activity but not tyrosine
phosphorylation of p21ras GTPase-activating protein. Proc Natl Acad
Sci USA. 89:pp. 1587–1591. 1992; View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hemesath TJ, Price ER, Takemoto C,
Badalian T and Fisher DE: MAP kinase links the transcription factor
Microphthalmia to c-Kit signalling in melanocytes. Nature.
391:298–301. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Masuhara S, Kasuya K, Aoki T, Yoshimatsu
A, Tsuchida A and Koyanagi Y: Relation between K-ras codon 12
mutation and p53 protein overexpression in gallbladder cancer and
biliary ductal epithelia in patients with pancreaticobiliary
maljunction. J Hepatobiliary Pancreat Surg. 7:198–205. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Qin B, Ariyama H, Baba E, Tanaka R, Kusaba
H, Harada M and Nakano S: Activated Src and Ras induce gefitinib
resistance by activation of signaling pathways downstream of
epidermal growth factor receptor in human gallbladder
adenocarcinoma cells. Cancer Chemother Pharmacol. 58:577–584. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Horiuchi H, Kawamata H, Furihata T,
Omotehara F, Hori H, Shinagawa Y, Ohkura Y, Tachibana M, Yamazaki
T, Ajiki T, et al: A MEK inhibitor (U0126) markedly inhibits direct
liver invasion of orthotopically inoculated human gallbladder
cancer cells in nude mice. J Exp Clin Cancer Res. 23:599–606.
2004.PubMed/NCBI
|
|
41
|
Zirn B, Samans B, Wittmann S, Pietsch T,
Leuschner I, Graf N and Gessler M: Target genes of the
WNT/beta-catenin pathway in Wilms tumors. Genes Chromosomes Cancer.
45:565–574. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yanagisawa N, Mikami T, Saegusa M and
Okayasu I: More frequent beta-catenin exon 3 mutations in
gallbladder adenomas than in carcinomas indicate different
lineages. Cancer Res. 61:19–22. 2001.PubMed/NCBI
|
|
43
|
Liu X, Lv R, Zhang L, Xu G, Bi J, Gao F,
Zhang J, Xue F, Wang F, Wu Y, et al: Long noncoding RNA expression
profile of infantile hemangioma identified by microarray analysis.
Tumour Biol. Oct 5–2016.(Epub ahead of print). View Article : Google Scholar
|
|
44
|
Li H, Xie S, Liu X, Wu H, Lin X, Gu J,
Wang H and Duan Y: Matrine alters microRNA expression profiles in
SGC-7901 human gastric cancer cells. Oncol Rep. 32:2118–2126. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ganci F, Sacconi A, Ben-Moshe NB,
Manciocco V, Sperduti I, Strigari L, Covello R, Benevolo M,
Pescarmona E, Domany E, et al: Expression of TP53
mutation-associated microRNAs predicts clinical outcome in head and
neck squamous cell carcinoma patients. Ann Oncol. 24:3082–3088.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang J, Zhang B, Wang T and Wang H:
LncRNA MALAT1 overexpression is an unfavorable prognostic factor in
human cancer: Evidence from a meta-analysis. Int J Clin Exp Med.
8:5499–5505. 2015.PubMed/NCBI
|
|
47
|
Wang X, Ning Y and Guo X: Integrative
meta-analysis of differentially expressed genes in osteoarthritis
using microarray technology. Mol Med Rep. 12:3439–3445. 2015.
View Article : Google Scholar : PubMed/NCBI
|