Introduction
Abdominal aortic aneurysm (AAA) is characterized as
a permanent dilation and degradation of the aortic wall. It is a
life-threatening condition affecting 8.3% of the population over 65
years of age (1). Currently, there
is no effective medication preventing or limiting the enlargement
of AAA other than surgical intervention. Thus, it is important to
investigate the mechanisms of AAA initiation and progression in
order to develop new therapies. Through bioinformatics analysis, we
found that focal adhesion was an important pathway in the
development of AAA. Moreover, numerous basic researches and
clinical studies have also reported that upregulation of adhesion
molecules could promote the initiation of AAA through an
inflammatory response induced by inflammatory cells infiltration
(2–5).
P-selectin is an adhesion molecule localized to the
surface of α granule in activated platelets and endothelial cells
in human. It could bind to P-selectin ligand on leukocyte mediating
the interaction between leukocytes with platelets or endothelial
cells. In the elastase aortic perfusion model, P-selectin knockout
could attenuate aneurysm formation (6). Previous study showed that Angiotensin
(Ang) II could increase the expression of P-selectin on endothelia
cells, which induced significant selectin-dependent leukocyte
rolling, recruitment, and infiltration. Nevertheless, fucoidan
could inhibit the leukocyte-endothelial cells interaction mediated
by P-selectin (7). Low molecular
weight fucoidan (LMWF) is a highly sulfated fraction degraded from
fucoidan in Saccharina Japonica (8). It has been reported to present high
affinity for P-selectin and abolish selectin-dependent recruitment
of leukocytes (9). Thus, this
study aimed to examine the effect of LMWF, as a competitive binding
agent of P-selectin, in ApoE(−/-) mice on inflammatory infiltration
and aneurysmal growth.
Materials and methods
Acquisition of microarray
datasets
Two gene expression profiles (GSE47472 and GSE57691)
were retrieved and downloaded from the National Center for
Biotechnology Information GEO database (http://www.ncbi.nlm.nih.gov/geo). The dataset
GSE47472, based on the platform of GPL10558 (Illumina HumanHT-12
V4.0 expression beadchip), consisted of 14 AAA neck samples from
patients and 8 normal samples from donors following brain
mortality. The array data of GSE57691, based on the same platform,
included 20 AAA samples from patients with small AAA (mean maximum
aortic diameter=54.3±2.3 mm), 29 AAA samples from patients with
large AAA (mean maximum aortic diameter=68.4±14.3 mm) and 10
control aortic specimen of organ donors.
Data processing and enrichment
analysis of DEGs
GEO2R (http://www.ncbi.nlm.nih.gov/geo/geo2r/) is an
interactive tool that allows comparing two groups of samples in a
GEO series. In this study, GEO2R was applied to screen
differentially-expressed genes (DEGs) between AAA and normal aortic
samples. The P-values were adjusted using Benjamini and Hochberg
false discovery rate method. The threshold for the DEGs was set as
adjusted P-value <0.05. Particularly, in GSE57691, the normal
aortic group was compared with small AAA group and large AAA group,
respectively. Venn plot was performed to determine the DEGs in all
three datasets, which was then submitted to the Database for
Annotation, Visualization and Integrated Discovery (DAVID;
http://david.abcc.ncifcrf.gov/) for
functional annotation analysis (10,11).
The significant enrichment analysis of DEGs was assessed based on
Kyoto Encyclopedia of Genes and Genomes (KEGG; http://www.genome.jp/kegg/kegg2.html)
with P-value <0.05 as the threshold.
Experimental design and AAA model
All experiments involving live animals were
conducted in compliance with the ‘Guide for the Care and Use of
Laboratory Animals’ and were approved by the institutional review
board of Zhongshan Hospital, Fudan University (Shanghai, China).
Six-month-old male apolipoprotein E-deficient (apoE-/-) mice
weighing 35–40 g were obtained from Shanghai Lab. Animal Research
Center. Ang II (1,000 ng/kg/min; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) or saline was administered subcutaneously for
28 days via Alzet osmotic minipumps (model 2004; DURECT, Cupertino,
CA, USA) as described previously (12). Thirty-six mice were randomly
allocated to 3 groups: Control, Ang II and LMWF group. LMWF group
mice were given 200 mg/kg/d LMWF (dissolved in 0.9% saline) by
gavage for 35 days, which was started 1 week before minipump
implantation. Identical volume of 0.9% saline were given to the
other 2 groups for the same period by gavage. All these mice were
fed with standard diet and water, and housed with a 12 h light/dark
cycle. The preparation method of LMWF was detailed before (8).
Measurement of aortic size using
ultrasonography
Aortic diameter measurements, intraluminal thrombus
and intimal flap recognition were obtained via a high-frequency
ultrasound system (Vevo 3100; VisualSonics, Toronto, Canada) in a
blind manner for each group. The short axis view was applied to
measure the maximum anterior-posterior diameter of abdominal aorta.
The aortic size was measured before minipump implantation and 14,
and 28 day after. The AAA is defined as a 50% increase in maximum
diameter of the abdominal aorta or the presence of abdominal aortic
dissection. One experienced operators who were blind to the study
design performed the quantitative analysis of the ultrasound
imaging. In addition, mice were daily monitored for mortality
analysis.
Peripheral blood monocytes count
After 4-weeks intervention, mice were anesthetized
by intraperitoneal injection of 0.12–0.15 ml sodium pentobarbital
(10 mg/ml). Blood was collected in citrated tubes for complete
blood count analysis. Peripheral blood monocytes were classified
and quantified based on the size and granularity of cells, and
content of nucleic acid using an automated hematology analyzer
(Mindray BC-2800vet; Mindray Bio-Medical Electronics Co., Ltd.,
Shenzhen, China). Counts were expressed as 109/l of
blood.
Histologic studies and
immuno-histochemical staining
Mice were sacrificed by cervical dislocation and
perfused briefly with phosphate-buffered saline (PBS). Aortic
segments were harvested and divided into two parts: one for
biochemical analysis and one was fixed with 4% paraformaldehyde in
PBS overnight. Then, it was embedded in paraffin and sectioned (5
µm in thickness). Hematoxylin and eosin (H&E) staining and
Elastin van Gieson (EVG) staining was used to show the gross
structure of aorta and medial elastin, respectively.
Immune-histochemical (IHC) assay was performed to identify
P-selectin expression in endothelium, F4/80, and matrix
metalloproteinases (MMP) expression in the aortic wall. In brief,
sections were de-paraffinized, rehydrated, and subjected to
heat-mediated antigen retrieval. After blocking with 3% bovine
serum albumin for 30 min at room temperature, the sections were
incubated with primary antibodies against P-selectin (1:100;
sc-8419; Santa Cruz Biotechnology, Inc., Dallas, TX, USA), F4/80
(1:100; ab6640), MMP-2 (1:100; ab37150), MMP-9 (1:100; ab38898;
Abcam, Cambridge, UK) overnight at 4°C. Then, incubated with second
antibody (1:500; Miaotong, Shanghai, China) for 50 min at room
temperature. Histological sections were examined under a light
microscope (BX53; Olympus Corp., Tokyo, Japan). Five random
microscopic fields in each mouse (n=6 in each group) were
evaluated. Macrophage was expressed as number of F4/80-positive
cells per square millimeter. P-selectin were quantified by the
intensity and range of IHC staining in endothelium.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Pro-inflammatory cytokines in the aortic tissues,
including monocyte chemotactic protein-1 (MCP-1), tumor necrosis
factor-α (TNF-α), and interleukin 1β (IL-1β), were determined by
qPCR. Total RNA was isolated using 1 ml TRIzol Reagent (Takara Bio
Inc., Kusatsu, Japan). Complementary DNA was prepared with 1 µg
total RNA using a Reverse Transcription kit (Takara Bio Inc.).
Specific mRNAs were amplified using SYBR-Green PCR Mix (Takara Bio
Inc.) in an ABI PRISM 7500 thermal cycler (Applied Biosystems;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). Cycling
conditions were 95°C for 30 sec, followed by 40 cycles at 95°C for
5 sec, 60°C for 34 sec. The primer sequences were as follows:
MCP-1: forward, 5′-CTTCTGGGCCTGCTGTTCA-3′ and reverse,
5′-CCAGCCTACTCATTGGGATCA-3′; TNF-α forward,
5′-CCCTCACACTCAGATCATCTTCT-3′ and reverse,
5′-GCTACGACGTGGGCTACAG-3′; IL-1β forward,
5′-GCAACTGTTCCTGAACTCAACT-3′ and reverse,
5′-ATCTTTTGGGGTCCGTCAACT-3′; β-actin forward,
5′-CGAGGCCCAGAGCAAGAGAGGTAT-3′ and reverse,
5′-CACGGTTGGCCTTAGGGTTCA-3′; The relative expression levels of mRNA
were calculated with β-actin mRNA as the invariant control using
the 2−ΔΔCq formula (13).
Western blotting
Aortic tissues were lysed by RIPA buffer. The
protein concentration was measured using a BCA protein Assay kit.
Then, 50 µg total proteins were separated by 10% SDS-PAGE and
transferred onto 0.45 µm PVDF transfer membranes (EMD Millipore,
Billerica, MA, USA). After blocked with 5% non-fat milk for 1 h at
room temperature, the membranes were incubation with primary
antibodies at 4°C overnight, including MMP2 (1:1,000; ab37150),
MMP9 (1:1,000; ab38898; both Abcam), β-actin (1:1,000; BS6007M;
Bioworld Technology, Inc., St. Louis Park, MN, USA). Then the
membranes were washed and incubated with secondary antibody
(1:3,000, Bioworld) for 30 min at room temperature and detected
using an enhanced chemiluminescence system. The bands relative
intensities were analyzed using AlphaEase FC software (USA).
Statistical analysis
Results are expressed as means ± standard deviations
or proportions. Comparisons between groups were performed by
one-way ANOVA analysis followed by a post hoc Bonferroni test for
intergroup pairwise comparisons (Prism 5; GraphPad Software Inc.,
San Diego, USA). For the incidence difference of AAA and
intraluminal thrombus between Ang II group and LMWF group, a
Fisher's exact test was applied (Stata 12.0; StataCorp, College
Station, TX, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Focal adhesion is involved in the
initiate and development of AAA
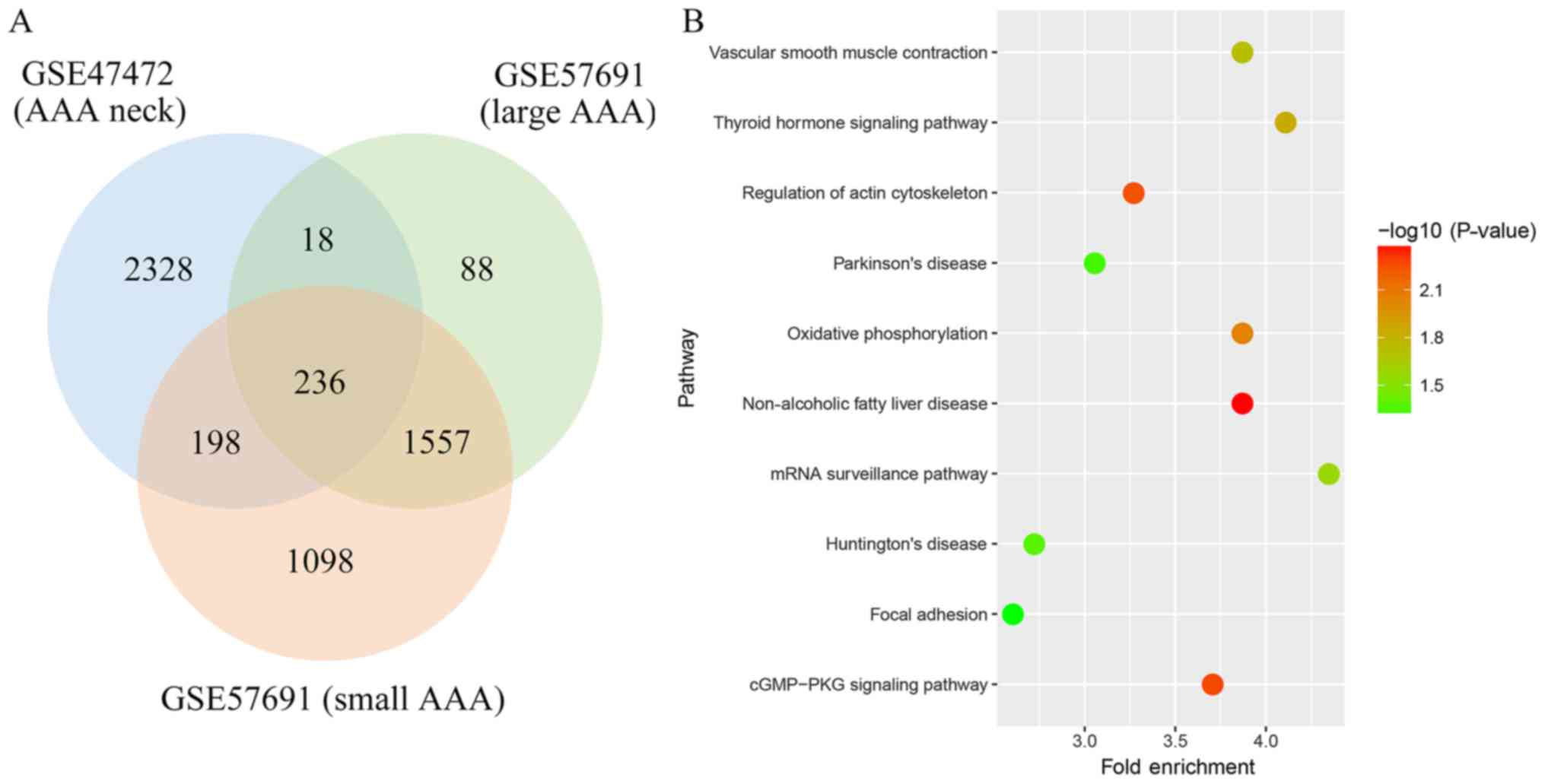
A total of 2,780, 1,899 and 3,089 genes were
identified as the DEGs from these datasets, respectively. Among
these DEGs, 236 genes were found in all three datasets (Fig. 1). Through pathway enrichment
analysis, these DEGs were identified to be significantly enriched
in non-alcoholic fatty liver disease, cGMP-PKG signaling pathway,
regulation of actin cytoskeleton, oxidative phosphorylation,
thyroid hormone signaling pathway, vascular smooth muscle
contraction, mRNA surveillance pathway, Huntington's disease,
Parkinson's disease and focal adhesion (Fig. 1).
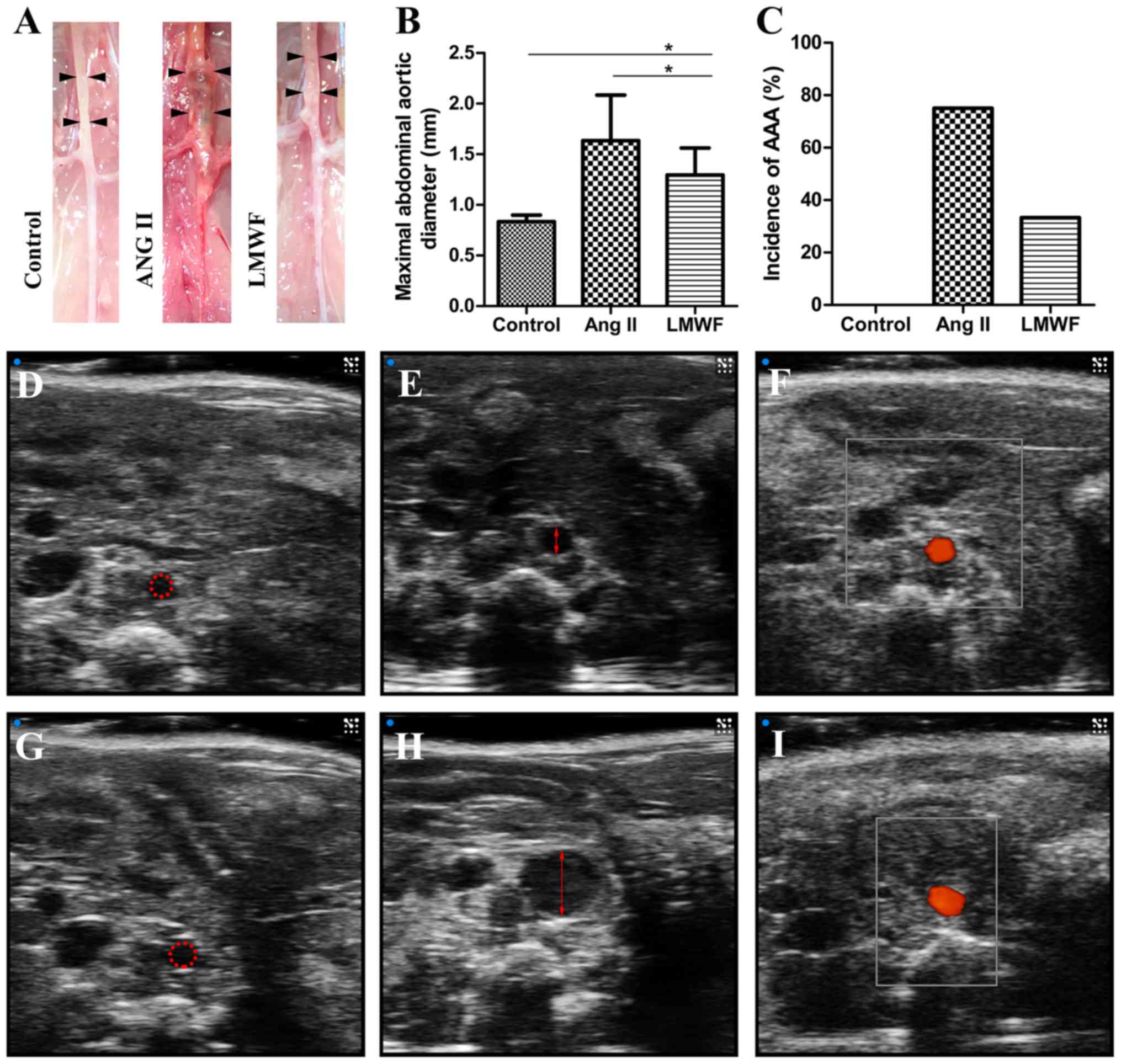
LMWF limits the enlargement of
experimental AAA but not the incidence
All experimental AAA induced by Ang II infusion
developed above and around renal arteries. There were no
significant differences in body weight among the 3 groups before
and after the intervention. One mouse died from abdominal aortic
rupture 18 day after minipump implantation in the Ang II group. All
mice survived till the end of intervention in the other 2 groups.
The incidence of AAA formation was not significantly different
between LMWF group and Ang II group (LMWF group, 33%; Ang II group,
75%; P=0.100). However, LMWF administration significantly decreased
the maximal aortic diameter (Control group, 0.84±0.06 mm; LMWF
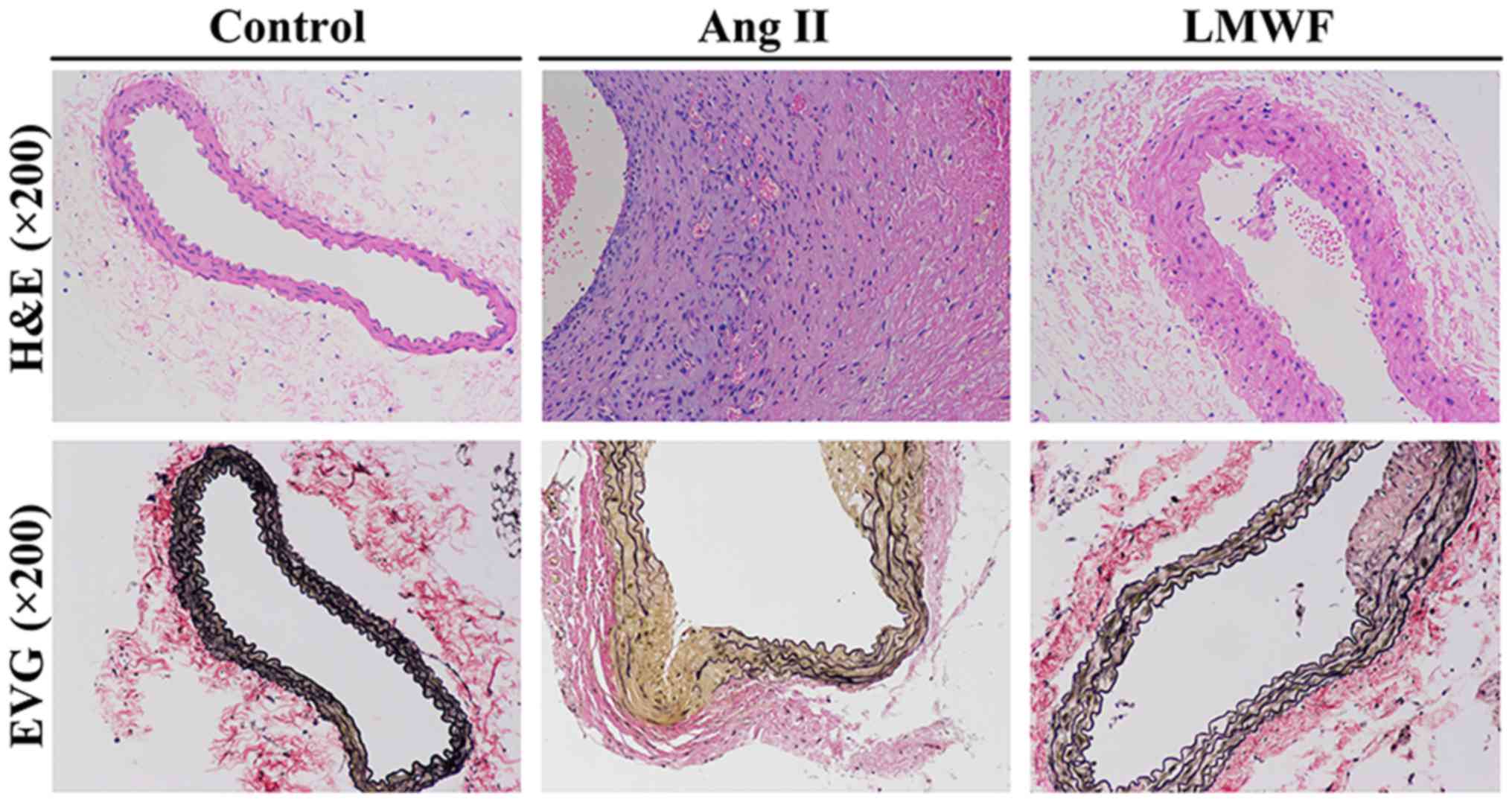
group, 1.30±0.27 mm; Ang II group, 1.63±0.45 mm) (Fig. 2). As to the histological changes of
AAA formation, H&E staining showed unusual medial thickening
and infiltration of inflammatory cells. EVG staining revealed the
degeneration and destruction of the medial elastic layers. However,
LMWF administration prevented the thickening of medial and
degeneration of the aortic wall (Fig.
3).
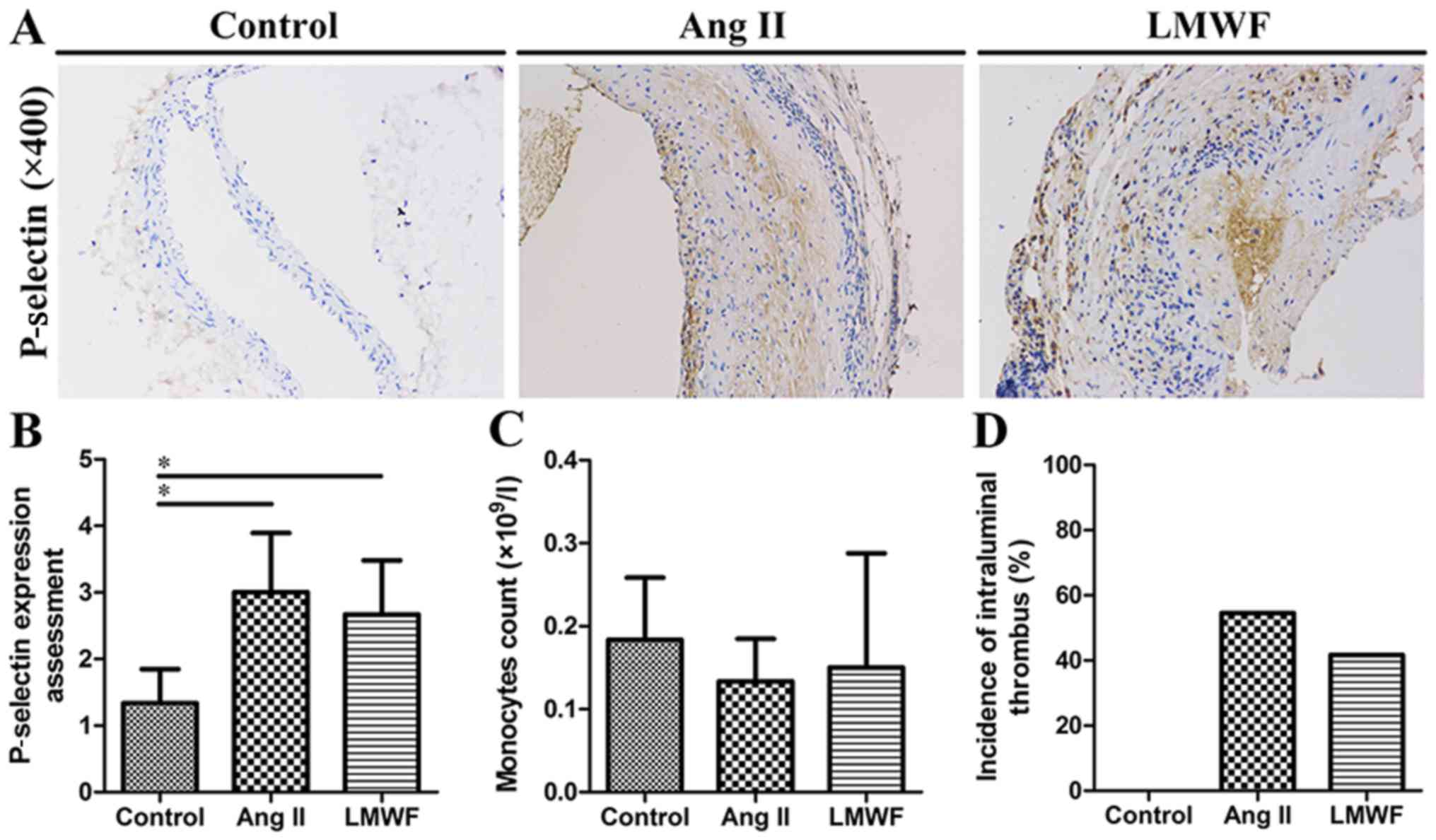
Circulating monocytes, intraluminal
thrombus and P-selectin are not suppressed by LMWF
administration
Complete blood count analysis showed that peripheral
blood monocytes were not significantly different in blood samples
among the three groups. Compared with Ang II group, the incidence
of intraluminal thrombus was also not decreased by LMWF
administration (LMWF group, 41.7%; Ang II group, 54.5%; P=0.414).
In addition, the upregulation of P-selectin in aortic intima was
found in Ang II group. However, this effect of Ang II on P-selectin
expression was independent of LMWF administration (Fig. 4).
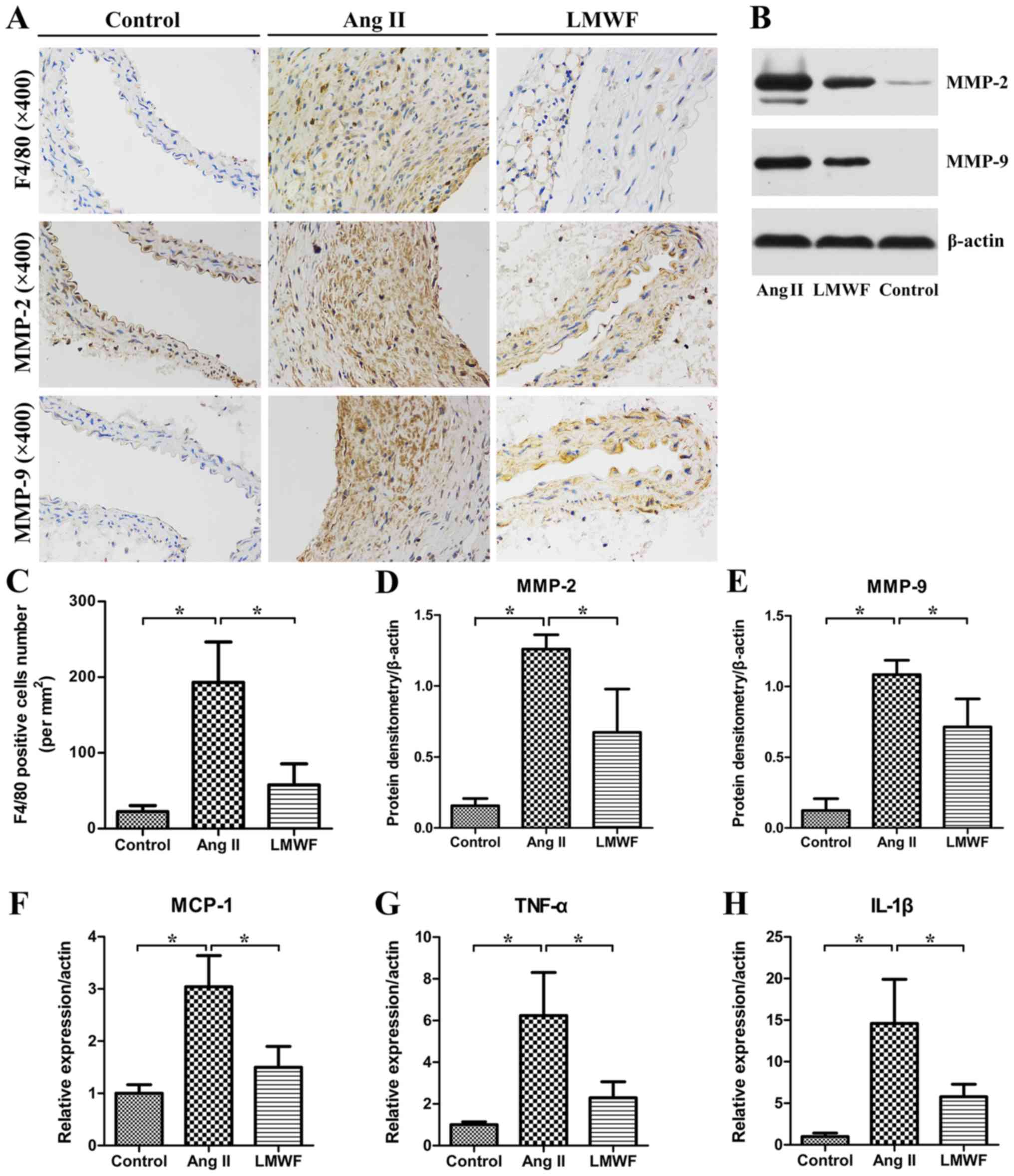
LMWF inhibits inflammatory
infiltration
In the Ang II group, an abundance of invasive
macrophages was observed in the aortic wall. The number of
F4/80-positive cells were significantly lower in the LMWF group
than the Ang II group. Moreover, the expression of TNF-α, MCP-1 and
IL-1β were significantly higher in the Ang II group than in the
Control group. LMWF significantly decreased TNF-α, MCP-1 and IL-1β
expression induced by inflammatory cells infiltration. Besides, IHC
and Western Blot assay of AAA tissues demonstrated that MMP
expression was enhanced in AAA tissues than that in Control group.
MMP-2 and MMP-9 expressions were significantly lower in LMWF group
than that in Ang II group (Fig.
5).
Discussion
It is well documented that the pathogenesis of AAA
is complicated and multifactorial. In this study, we attempted to
detect the potential mechanism by bioinformatic analysis. Based on
the microarray data contributed by Biros et al, we
identified 236 common DEGs and 10 pathways, which may affect the
development of AAA (14,15). Notably, the tissue used in the
microarray analysis included AAA neck, small-AAA aortic wall and
large-AAA aortic wall, which indicated the formation and
development of AAA. Although the fold enrichment of focal adhesion
was relatively low, it still suggested the importance of leukocytes
adhering to endothelium except for apoptosis of vascular smooth
muscle cells and degradation of the extracellular matrix in the
development of AAA. Thus, we aimed to detect the possibility of
limiting the enlargement of AAA by interfering
leukocyte-endothelial interactions.
P-selectin is an adhesion molecule expressed on the
endothelial cells. In vitro, P-selectin expression in
endothelial cells and macrophage adhesion were increased by Ang II,
which could be inhibited by fucoidan (7). Besides, accumulating evidence has
determined that medial and adventitial macrophage activity promotes
aneurysmal degeneration via production of MMP, pro-inflammatory
cytokines, and reactive oxygen species (16,17).
In this study, we found circulating monocytes was not changed by
LMWF administration, as well as P-selectin expressed in the
endothelial cells of AAA. However, the invasive macrophages were
significantly lower in the medial of abdominal aorta. PCR and IHC
analysis also confirmed that inflammatory mediators and MMP
expressions were also significantly decreased in AAA tissue. These
results concluded that dietary intake of LMWF could limit the
enlargement of AAA in Ang II-infused ApoE-/-mice, at least in part,
by blocking the adhesion site of macrophages and endothelium, and
thus decreasing the inflammatory mediators, MMP-2 and MMP-9
production from macrophage infiltration. Furthermore, LMWF was also
proved the potential to directly modulate the expression of MMP-2
(18).
In a previous study, Alsac et al referred
that neutrophil rolling and trapping in the intraluminal thrombus
were mainly mediated by P-selectin and P-selectin glycoprotein
ligand-1 on the activated platelets and neutrophil surface, which
led to the death and releasing proteolytic enzymes of neutrophil
(19). Thus, they detected the
effect of fucoidan on leukocyte-thrombus interaction in an AAA
model, which was characterized by neutrophil-rich intraluminal
thrombus induced by repeated episodes of weak bacteremia. Finally,
they found that fucoidan could limit the enlargement of AAA by
decreasing neutrophil activation. However, the AAA model used in
our study was different from theirs. It was induced by traditional
cardiovascular risk factors, including atherosclerotic lesions and
hypertension. The incidence of intraluminal thrombus and P-selectin
expression in endothelial cells were not significantly different
between Ang II group and LMWF group. Moreover, circulating
monocytes were not significantly different among these groups.
Nevertheless, the invasive macrophages in the media were
significantly lower by LMWF through interfering the
leukocyte-endothelial cells interaction. Thus, the mechanism we
clarified was a supplement to theirs based on a different animal
model, which also indicated that both endothelial cell and platelet
P-selectin contributed to the formation of AAA.
In fact, increased infiltration of inflammatory
cells was an established pathological hallmark of AAA. During the
initiate of inflammatory cells migrating into sub-endothelial
space, focal adhesion plays an important role. Previously, numerous
efforts have been made to interfere the leukocyte-endothelial cell
interaction (20,21). Considering that Ang II could induce
endothelial activation and dysfunction, calcium channel inhibitors
and Ang II receptor antagonists have been proved to prevent the Ang
II-induced over-expression of adhesion molecules and chemokines,
and presented a good endothelial protective activity (22,23).
Another important mechanism was the monocyte chemoattraction
induced by Ang II, leukocyte-endothelial cell interaction could be
inhibited by anti-chemokines without affecting the endothelial
expression of P-selectin and VCAM-1 (24). Iida et al also detected that
peptide inhibitor of CXCL4-CCL5 heterodimer formation could
attenuate AAA in mice due to that it could inhibit CCL5-mediated
monocyte chemotaxis and arrest on activated endothelial cells in
vitro (25). In this study,
the decreased inflammatory mediators also confirm the importance of
chemokines.
The adhesion character of fucoidan was also applied
in the imaging of AAA. In some studies, P-selectin was described as
a molecular imaging target for AAA early detection, dilatation, and
rupture risk assessment (26,27).
A team from France developed microcapsules functionalized with
fucoidan to target P-selectin, enabling detection of activated
endothelium and thrombosis of AAA in vivo (28). Notably, LMWF is highly sulfated
fractions degraded from fucoidan, with higher affinity for
P-selectin. The structure property of fucoidan was associated with
its specific biological activity. LMWF was more potent than
fucoidan in endothelial protection (29). Moreover, LMWF could induce
endothelial cell migration via the PI3K/AKT pathway and promote
angiogenesis in a rat model of critical hindlimb ischemia (30). As to the character of binding to
activated human platelets, LMWF could suppress the thrombin-induced
platelet aggregation and has a potential to become an oral
antithrombotic agent (31–33). In addition, Deux et al found
that LMWF prevented neointimal hyperplasia in rabbit iliac artery
in-stent restenosis model (34).
Recently, LMWF has been used in food supplements and pharmaceutical
products (35). Toxicological
evaluation of LMWF in vitro and in vivo have verified
that there were no toxicological effects at a high dose (36). Overall, LMWF showed a promising
future in the detection and treatment of vascular diseases.
Our study has several limitations. First, although
the incidence of ILT was not significantly different between the
two groups, the volume of ILT was not quantified. We still could
not exclude the potential effect of activated platelet and
thrombosis. Second, a single dose of LMWF was applied in this
study. Third, macrophages also produce pro-inflammatory cytokines,
which we could not identify whether LMWF could directly reduce
these chemokines. Fourth, MMPs activities were not tested in the
present study, which was suggested for future studies. Last, the
expression of P-selectin in the endothelial cells could not be
quantified by western blot analysis. The single section of IHC
analysis may be unstable.
In conclusion, focal adhesion involved in the
initiate and development of AAA. Dietary intake of LMWF limited AAA
enlargement through inhibiting P-selectin-dependent leukocytes roll
on the endothelium to initiate leukocyte recruitment, adhesion and
infiltration. These results suggest that LMWF may have beneficial
effects against AAA enlargement.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81370424).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZS and CL contributed to the conception and design;
LC and YW performed the data collection and interpretation; MZ and
YD analyzed the datasets and wrote this article. ZS obtained the
funding. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experiments involving live animals were
conducted in compliance with the ‘Guide for the Care and Use of
Laboratory Animals’ and approved by the Institutional Review Board
of Zhongshan Hospital, Fudan University.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sampson UK, Norman PE, Fowkes FG, Aboyans
V, Song Y, Harrell FE Jr, Forouzanfar MH, Naghavi M, Denenberg JO,
McDermott MM, et al: Estimation of global and regional incidence
and prevalence of abdominal aortic aneurysms 1990 to 2010. Global
heart. 9:159–170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thomas M, Gavrila D, McCormick ML, Miller
FJ Jr, Daugherty A, Cassis LA, Dellsperger KC and Weintraub NL:
Deletion of p47phox attenuates angiotensin II-induced abdominal
aortic aneurysm formation in apolipoprotein E-deficient mice.
Circulation. 114:404–413. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sharma AK, Lu G, Jester A, Johnston WF,
Zhao Y, Hajzus VA, Saadatzadeh MR, Su G, Bhamidipati CM, Mehta GS,
et al: Experimental abdominal aortic aneurysm formation is mediated
by IL-17 and attenuated by mesenchymal stem cell treatment.
Circulation. 126 11 Suppl 1:S38–S45. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Harrison SC, Smith AJ, Jones GT, Swerdlow
DI, Rampuri R, Bown MJ; Aneurysm Consortium, ; Folkersen L, Baas
AF, de Borst GJ, et al: Interleukin-6 receptor pathways in
abdominal aortic aneurysm. Eur Heart J. 34:3707–3716. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li MW, Mian MO, Barhoumi T, Rehman A, Mann
K, Paradis P and Schiffrin EL: Endothelin-1 overexpression
exacerbates atherosclerosis and induces aortic aneurysms in
apolipoprotein E knockout mice. Arterioscler Thromb Vasc Biol.
33:2306–2315. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hannawa KK, Cho BS, Sinha I, Roelofs KJ,
Myers DD, Wakefield TJ, Stanley JC, Henke PK and Upchurch GR Jr:
Attenuation of experimental aortic aneurysm formation in P-selectin
knockout mice. Ann N Y Acad Sci. 1085:353–359. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Piqueras L, Kubes P, Alvarez A, O'Connor
E, Issekutz AC, Esplugues JV and Sanz MJ: Angiotensin II induces
leukocyte-endothelial cell interactions in vivo via AT(1) and AT(2)
receptor-mediated P-selectin upregulation. Circulation.
102:2118–2123. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu Y, Zhang Q, Luo D, Wang J and Duan D:
Low molecular weight fucoidan modulates P-selectin and alleviates
diabetic nephropathy. Int J Biol Macromol. 91:233–240. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jin W, Wang J, Jiang H, Song N, Zhang W
and Zhang Q: The neuroprotective activities of
heteropolysaccharides extracted from Saccharina Japonica.
Carbohydr Polym. 97:116–120. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang da W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: Paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13S. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Daugherty A and Cassis LA: Mouse models of
abdominal aortic aneurysms. Arterioscler Thromb Vasc Biol.
24:429–434. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Biros E, Moran CS, Rush CM, Gäbel G,
Schreurs C, Lindeman JH, Walker PJ, Nataatmadja M, West M, Holdt
LM, et al: Differential gene expression in the proximal neck of
human abdominal aortic aneurysm. Atherosclerosis. 233:211–218.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Biros E, Gäbel G, Moran CS, Schreurs C,
Lindeman JH, Walker PJ, Nataatmadja M, West M, Holdt LM,
Hinterseher I, et al: Differential gene expression in human
abdominal aortic aneurysm and aortic occlusive disease. Oncotarget.
6:12984–12996. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yoshida S, Fuster JJ and Walsh K:
Adiponectin attenuates abdominal aortic aneurysm formation in
hyperlipidemic mice. Atherosclerosis. 235:339–346. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lu HY, Huang CY, Shih CM, Chang WH, Tsai
CS, Lin FY and Shih CC: Dipeptidyl peptidase-4 inhibitor decreases
abdominal aortic aneurysm formation through GLP-1-dependent
monocytic activity in mice. PLoS One. 10:e01210772015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hlawaty H, Suffee N, Sutton A, Oudar O,
Haddad O, Ollivier V, Laguillier-Morizot C, Gattegno L, Letourneur
D and Charnaux N: Low molecular weight fucoidan prevents intimal
hyperplasia in rat injured thoracic aorta through the modulation of
matrix metalloproteinase-2 expression. Biochem Pharmacol.
81:233–243. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Alsac JM, Delbosc S, Rouer M, Journé C,
Louedec L, Meilhac O and Michel JB: Fucoidan interferes with
Porphyromonas gingivalis-induced aneurysm enlargement by decreasing
neutrophil activation. J Vasc Surg. 57:796–805. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
da Cunha V, Tham DM, Martin-McNulty B,
Deng G, Ho JJ, Wilson DW, Rutledge JC, Vergona R, Sullivan ME and
Wang YX: Enalapril attenuates angiotensin II-induced
atherosclerosis and vascular inflammation. Atherosclerosis.
178:9–17. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cui W, Zheng Y, Zhang Q, Wang J, Wang L,
Yang W, Guo C, Gao W, Wang X and Luo D: Low-molecular-weight
fucoidan protects endothelial function and ameliorates basal
hypertension in diabetic Goto-Kakizaki rats. Lab Invest.
94:382–393. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Iida Y, Xu B, Schultz GM, Chow V, White
JJ, Sulaimon S, Hezi-Yamit A, Peterson SR and Dalman RL: Efficacy
and mechanism of angiotensin II receptor blocker treatment in
experimental abdominal aortic aneurysms. PLoS One. 7:e496422012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mieth A, Revermann M, Babelova A, Weigert
A, Schermuly RT and Brandes RP: L-type calcium channel inhibitor
diltiazem prevents aneurysm formation by blood pressure-independent
anti-inflammatory effects. Hypertension. 62:1098–1104. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mateo T, Abu Nabah YN, Abu Taha M, Mata M,
Cerdá-Nicolás M, Proudfoot AE, Stahl RA, Issekutz AC, Cortijo J,
Morcillo EJ, et al: Angiotensin II-induced mononuclear leukocyte
interactions with arteriolar and venular endothelium are mediated
by the release of different CC chemokines. J Immunol.
176:5577–5586. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Iida Y, Xu B, Xuan H, Glover KJ, Tanaka H,
Hu X, Fujimura N, Wang W, Schultz JR, Turner CR and Dalman RL:
Peptide inhibitor of CXCL4-CCL5 heterodimer formation, MKEY,
inhibits experimental aortic aneurysm initiation and progression.
Arterioscler Thromb Vasc Biol. 33:718–726. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li B, Juenet M, Aid-Launais R, Maire M,
Ollivier V, Letourneur D and Chauvierre C: Development of polymer
microcapsules functionalized with fucoidan to target P-selectin
overexpressed in cardiovascular diseases. Adv Healthc Mater.
6:2017. View Article : Google Scholar
|
|
27
|
Bonnard T, Serfaty JM, Journé C, Ho Tin
Noe B, Arnaud D, Louedec L, Derkaoui SM, Letourneur D, Chauvierre C
and Le Visage C: Leukocyte mimetic polysaccharide microparticles
tracked in vivo on activated endothelium and in abdominal aortic
aneurysm. Acta Biomater. 10:3535–3545. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rouzet F, Bachelet-Violette L, Alsac JM,
Suzuki M, Meulemans A, Louedec L, Petiet A, Jandrot-Perrus M,
Chaubet F and Michel JB: Radiolabeled fucoidan as a p-selectin
targeting agent for in vivo imaging of platelet-rich thrombus and
endothelial activation. J Nucl Med. 52:1433–1440. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen A, Lan Y, Liu J, Zhang F, Zhang L, Li
B and Zhao X: The structure property and endothelial protective
activity of fucoidan from Laminaria japonica. Int J Biol
Macromol. 105:1421–1429. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bouvard C, Galy-Fauroux I, Grelac F,
Carpentier W, Lokajczyk A, Gandrille S, Colliec-Jouault S, Fischer
AM and Helley D: Low-molecular-weight fucoidan induces endothelial
cell migration via the PI3K/AKT pathway and modulates the
transcription of genes involved in angiogenesis. Mar Drugs.
13:7446–7462. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhu Z, Zhang Q, Chen L, Ren S, Xu P, Tang
Y and Luo D: Higher specificity of the activity of low molecular
weight fucoidan for thrombin-induced platelet aggregation. Thromb
Res. 125:419–426. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bachelet L, Bertholon I, Lavigne D, Vassy
R, Jandrot-Perrus M, Chaubet F and Letourneur D: Affinity of low
molecular weight fucoidan for P-selectin triggers its binding to
activated human platelets. Biochim Biophys Acta. 1790:141–146.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao X, Guo F, Hu J, Zhang L, Xue C, Zhang
Z and Li B: Antithrombotic activity of oral administered low
molecular weight fucoidan from Laminaria Japonica. Thromb Res.
144:46–52. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Deux JF, Meddahi-Pellé A, Le Blanche AF,
Feldman LJ, Colliec-Jouault S, Brée F, Boudghène F, Michel JB and
Letourneur D: Low molecular weight fucoidan prevents neointimal
hyperplasia in rabbit iliac artery in-stent restenosis model.
Arterioscler Thromb Vasc Biol. 22:1604–1609. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tsai HL, Tai CJ, Huang CW, Chang FR and
Wang JY: Efficacy of low-molecular-weight fucoidan as a
supplemental therapy in metastatic colorectal cancer patients: A
double-blind randomized controlled trial. Mar Drugs. 15:pii: E122.
2017. View Article : Google Scholar
|
|
36
|
Hwang PA, Yan MD, Lin HT, Li KL and Lin
YC: Toxicological evaluation of low molecular weight fucoidan in
vitro and in vivo. Mar Drugs. 14:pii: E121. 2016. View Article : Google Scholar
|