Introduction
Cases of prostate cancer-associated morbidity and
mortality are rapidly increasing in China (1), and prostate cancer is the most
prevalent malignant tumor in the United States (2), thus affecting life expectancy and the
quality of life of patients with prostate cancer. Androgen
deprivation and radical resection surgery remain the primary
methods of treatment, with androgen blockade having an important
role in current therapeutic methods. However, during treatment,
many patients with prostate cancer rapidly develop hormone
resistance and enter the castration-insensitive phase of the
disease [castration-resistant prostate cancer (CRPC)], in which the
incidence of local and distant metastases markedly increases and
patients' prognoses are seriously affected.
Investigations to determine the underlying
mechanisms of prostate cancer metastasis have become an important
area of research. Epithelial-mesenchymal transition (EMT) is a
well-established mechanism of malignant tumor invasion and
metastasis (3–7). Clinical and pathological studies have
demonstrated that as the Gleason score of prostate cancer
increases, the structure and basement membranes of prostate cancer
epithelial cells gradually disappear, the appearance and growth of
cell clusters that rapidly become more invasive increases, and the
levels of epithelial cell marker E-cadherin (E-Cad) decrease,
whereas the mesenchymal cell markers N-cadherin (N-Cad) and
vimentin increase (8–10). Aberrant expression of these EMT
markers in numerous malignant tumors, including prostate cancer,
has an important role in cancer metastasis (8–13).
The mechanisms underlying EMT in prostate cancer
invasion and metastasis have not yet been fully determined.
Programmed cell death 1 ligand 1 (PD-L1) is a cell surface
glycoprotein belonging to the B7 family of co-stimulatory
molecules, which is widely expressed in numerous malignant tumor
types (14–16). PD-L1 is not only involved in tumor
immune evasion, but also in EMT, thus suggesting that PD-L1 is
involved in tumor metastasis and prognosis (14–16).
Studies have demonstrated that the Janus kinase (JAK)/signal
transducer and activator of transcription 3 (STAT3) signaling
pathway has an important role in tumor progression and metastasis
(17–19). The JAK/STAT3 pathway is not only
involved in prostate cancer progression, but is also closely
associated with the transition from hormone dependence to hormone
independence in prostate cancer (20–22).
A previous study revealed that increased ataxia
telangiectasia mutated kinase (ATM) expression in lymph node
metastases of breast cancer is closely associated with tumor
invasion and metastasis (23). It
has also been demonstrated that reduced expression of ATM can
inhibit ovarian cancer metastasis (24). Considering these findings, the
present study aimed to investigate the clinical findings in samples
obtained from patients with prostate cancer; the results revealed
that ATM expression levels in prostate tissue samples from patients
with CRPC were significantly enhanced compared with prostate tissue
samples from patients with hormone-dependent prostate cancer
(HDPC). In the present study, to investigate the mechanisms
underlying the elevated expression of ATM, the expression of the
ATM gene in tumor cells was inhibited via gene knockout and the
addition of inhibitors, after which cell migration and the
expression levels of EMT-associated markers were investigated. The
results of the present study revealed that, as determined by
inhibition of PD-L1 and the JAK/STAT3 signaling pathway, the
ATM-JAK-PD-L1 signaling pathway has an important role in the
metastatic progression of CRPC, and therefore may represent a novel
therapeutic approach for CRPC-targeted therapy.
Materials and methods
Clinical species
To investigate whether ATM is elevated in prostate
cancer tissues from patients with CRPC who are prone to develop
metastasis. We collected surgical specimens of prostate cancer from
April 2006 to April 2016 in our hospital. All patients were male,
aged 67–84 years, of whom 62 were HDPC and 50 were CRPC. 112
surgical specimens isolated from patients with prostate cancer were
obtained. All patients provided informed consent. The present study
was approved by the Ethics Committee of the Second Affiliated
Hospital of Soochow University (Suzhou, China). Among the total
patients included in the present study, 62 patients with HDPC but
without local and distant metastases were included, all of which
underwent radical prostatectomy. The remaining 50 patients with
CRPC had developed varying local or distant metastases and
underwent palliative surgery to alleviate urinary tract
symptoms.
Cell culture
CRPC cell lines C4-2 and CWR22Rv1 were used in the
present study. C4-2 cells were obtained from the China Center for
Type Culture Collection (Wuhan, China). CWR22Rv1 cells were
purchased from the American Type Culture Collection (Manassas, VA,
USA). Cells were cultured in RPMI-1640 (Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% charcoal stripped
fetal bovine serum (FBS; Thermo Fisher Scientific, Inc.). CP466722
(Selleck Chemicals, Houston, TX, USA), JAK inhibitor 1 (EMD
Millipore, Billerica, MA, USA) and Stattic (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) inhibitors were added to cultured tumor
cells (1:1,000 for all inhibitors) at 37°C, at 6, 12 and 24 h time
intervals to inhibit the expression of ATM, JAK and STAT3,
respectively. RT-qPCR, western blot and cell migration assay were
performed at 6, 12 and 24 h respectively.
Immunohistochemistry (IHC)
Tumor tissues were fixed in 10% (v/v) formaldehyde
in PBS, embedded in paraffin, and cut into 5-µm thick sections. The
sections were air-dried, then cleared and dehydrated. Antigen
retrieval was performed by boiling sections in sodium citrate
buffer. Subsequently, sections were incubated for 1 h at room
temperature in blocking solution 5% BSA (AR0004, Boster Biological
Technology, Pleasanton, CA, USA). Incubation with primary antibody
Phosphorylated (p)-ATM (MA1-46069, Thermo Fisher Scientific, Inc.
USA) was at 4°C overnight. Following washes with PBS three times,
incubation with secondary rabbit anti-mouse antibody (61–0200,
Thermo Fisher Scientific, Inc. USA) at room temperature for 20 min
followed by three additional washes with PBS. Following staining,
tissues were counterstained with hematoxylin. Error bars and
significance of values were determined by counting positively
stained cells in three randomly selected areas of each slide. Tumor
tissue sections were immunostained using a mouse and rabbit
specific HRP/DAB detection IHC kit (ab80436, Abcam, Cambridge, UK).
Following this, tissues were counterstained by hematoxylin. A light
microscope was used (Olympus Corporation, Tokyo, Japan), at
magnification, ×40. Error bars and significance values were
obtained by manually counting positively stained cells from one
randomly chosen area of slides of three different stains.
ATM knockout cell construction
ATM knockout cells and corresponding control cells
were established via lentivirus infection. The lentiviral pLenti-II
vector (Addgene, Inc., Cambridge, MA, USA) carried ATM small
interfering (si)RNA (ATMSi) or scrambled (Sc) gene sequences as
follows: ATM scramble forward, 5′-UUCUCCGAACGUGUCACGUdTdT-3′ and
reverse, 5′-ACGUGACACGUUCGGAGAAdTdT-3′ and ATM siRNA forward,
5′-CAUACUACUCAAAGACAUUdTdT-3′ and reverse,
5′-AAUGUCUUUGAGUAGUAUGdTdT-3′. pLent-II-ATMSi or Sc, psPAX2
(virus-packaging plasmid) and pMD2G (envelope plasmid; All from
Qiagen, Inc., Valencia, CA, USA) were transfected into 293T cells
at a ratio of 4:3:2 using PolyFect transfection reagent (Qiagen,
Inc.). Following C4-2 and CWR22Rv1 cells being virally infected
overnight, the culture media containing the virus were replaced
with normal culture media and maintained under normal cell culture
conditions. After subculturing cells, the ATM-knockdown cells were
selected by culturing cells in the presence of puromycin (2
µgxml−1; 540411; Millipore, Billerica, MA,
USA) and maintained in media containing 0.1
µgxml−1 puromycin.
Migration assay
Tumor cells (1×104) were seeded into the
upper chamber of a Transwell support (Corning Incorporated,
Corning, NY, USA) with serum-free media, whereas medium containing
10% FBS was added to the lower chamber. Following incubation for 24
h at 37°C, cells were fixed in methanol and then stained with
crystal violet. Positively stained cells in three randomly selected
visual fields were counted under a light microscope (Olympus
Corporation), magnification, ×20 (inlet, ×100). Each assay was
repeated three times. Antibodies used were: JAK inhibitor 1
(420099; EMD Millipore), CP466722 (S2245; Selleck Chemicals), PD-L1
Antibody (62-5982-80, Thermo Fisher Scientific, Inc.).
Cell proliferation assay
Tumor cells (1×103) were seeded in a
96-well plate, the ATM inhibitor CP466722 was added and a control
group was set up without CP466722. The growth of cells in MTT (5
mg/ml; Sigma-Aldrich; Merck KGaA) was assessed daily, DMSO was used
to dissolve formazan in cells and the optical density of each well
was determined at 570 nm relative to the control group.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from cells (1 µg) was subjected to reverse
transcription using Superscript III transcriptase (Invitrogen;
Thermo Fisher Scientific, Inc.) as follows: 25°C for 10 min, 42°C
for 50 min, 70°C for 15 min and 4°C hold. Quantitative PCR was
conducted using the appropriate primers and a Bio-Rad CFX96 system
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) with SYBR green to
determine the mRNA expression levels of genes of interest. Enzyme
activation and DNA initial denaturation occurred at 95°C for 3 min,
followed by for 40 cycles of: DNA denaturation at 95°C for 15 sec
and annealing/extension at 60°C for 45 sec. A melting curve
analysis was performed. GraphPad Prism 5 was used to analyze the
results. The 2−ΔΔCq method was used to calculate the
relative expression level of each gene. Expression levels were
normalized to GAPDH.
Western blotting
Tumor cells were collected following centrifugation
at 300 × g for 5 min at 4°C) and supernatant removal. Cells were
lysed in RIPA buffer (50 mM Tris-Cl at pH 7.5, 150 mM NaCl, 1%
NP-40, 0.5% sodium deoxycholate, 1 mM EDTA, 1 µg/ml leupeptin, 1
µg/ml aprotinin, 0.2 mM PMSF). Proteins (20–40 µg) were separated
on 8–10% SDS/PAGE gel and then transferred onto PVDF membranes (EMD
Millipore). The PVDF membrane was placed in blocking buffer and put
on shaker for 30 min at room temperature and wash three times with
PBS for 10 min. Subsequently, membranes were incubated with primary
antibodies (1:1,000), for 1 h at room temperature on a shaker.
Following 3 washes for 10 min each, with PBS with Tween 20,
HRP-conjugated secondary antibodies (1:5,000; ab97030; Abcam)
incubated for 1 h at room temperature on a shaker. Finally, band
intensities were visualized in Imager (Bio-Rad Laboratories) using
ECL system (34095; Thermo Fisher Scientific, Inc.). Primary
antibodies used in the assay included: Phosphorylated (p)-ATM
(MA1-46069; Thermo Fisher Scientific, Inc.), PD-L1, p-JAK1, p-JAK2
and p-STAT3 and GAPDH (14-5983-82, 44–422G, 710928, 44–384G,
MA5-15738; Thermo Fisher Scientific, Inc.).
Statistical analysis
All statistical analyzes were performed using SPSS
19.0 (IBM Corp., Armonk, NY, USA) statistical software. All
experiments were performed three times which are presented as the
mean ± standard deviation and differences between two groups were
analyzed using a two-tailed Student's t-test. One-way analysis of
variance followed by Fisher's least significant difference post hoc
test was used for comparisons among multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
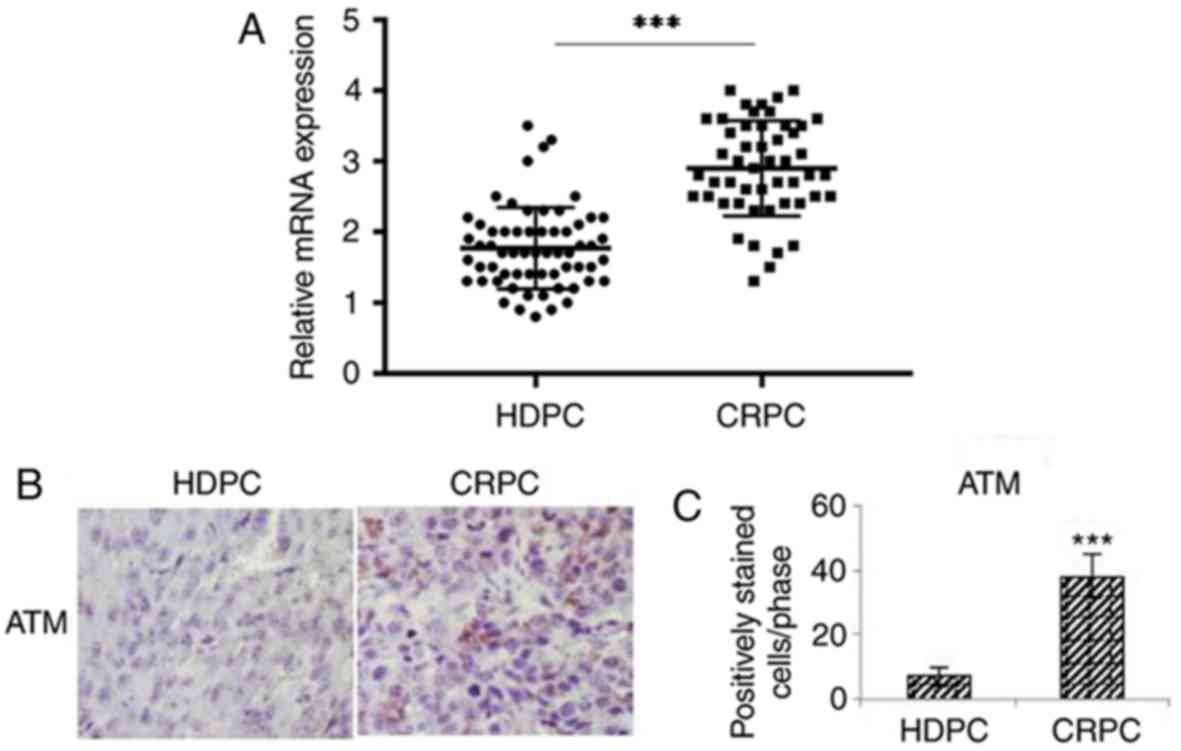
ATM expression in prostate cancer
tissues is higher in patients with CRPC compared with patients with
HDPC
ATM expression in prostate cancer specimens was
investigated via RT-qPCR and IHC. IHC was performed to determine
the levels of p-ATM, since ATM is predominantly activated by
phosphorylation. Based on clinical pathological findings of
prostate cancer specimens, the RT-qPCR results revealed that the
mRNA expression levels of p-ATM were significantly enhanced in
prostate cancer tissues obtained from patients with CRPC compared
with from patients with HDPC (P<0.01; Fig. 1A). The results of IHC analysis
demonstrated that the expression levels of p-ATM were significantly
enhanced in prostate cancer tissues obtained from patients with
CRPC compared with from patients with HDPC (P<0.01; Fig. 1B and C).
Migration and EMT of ATM knockout CRPC
cells, C4-2-ATMSi and CWR22Rv1-ATMSi, are suppressed
Using the CRPC cell lines C4-2 and CWR22Rv1 as model
systems, the successful establishment of ATM knockout cell lines
C4-2-ATMSi and CWR22Rv1-ATMSi, and control lines C4-2-Sc and
CWR22Rv1-Sc, was revealed (Fig.
2A). The migration of ATM knockout cells was significantly
decreased 24 h post-transfection in C4-2-ATMSi and CWR22Rv1-ATMSi
cells compared with in the control cells (Fig. 2B). The expression levels of
EMT-associated markers were investigated in ATM knockout cells by
RT-qPCR; the results demonstrated that the expression levels of
N-Cad, vimentin and vascular endothelial growth factor were
significantly decreased compared with in the control cells, whereas
E-Cad expression was significantly increased in ATM knockout cells
compared with in the control cells (Fig. 2C). These results suggested that ATM
knockout suppresses EMT in tumors.
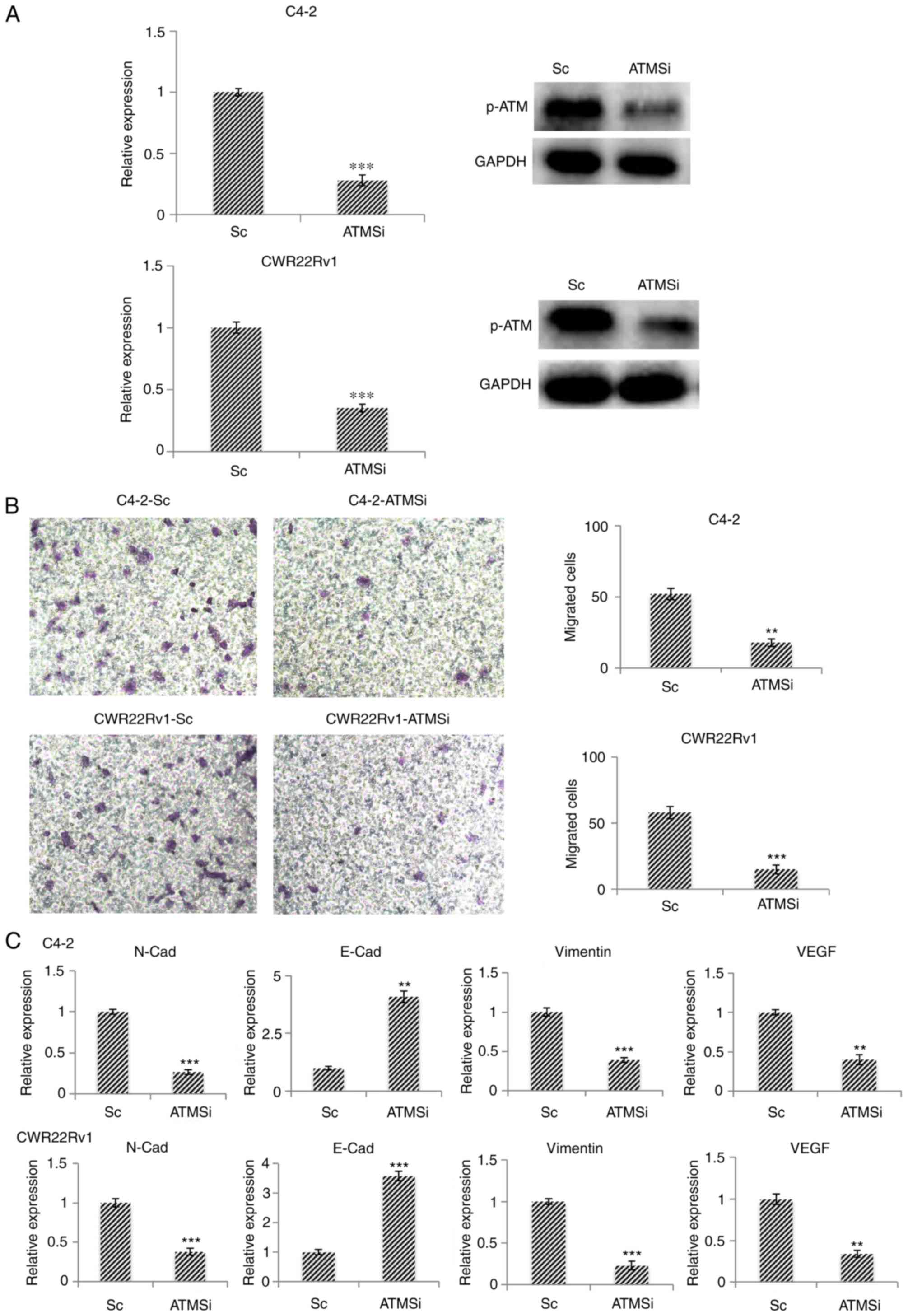 | Figure 2.In C4-2-ATMSi and CWR22Rv1-ATMSi
knockout cell lines, migration and EMT are significantly suppressed
compared with in the control group. (A) C4-2-ATMSi and
CWR22Rv1-ATMSi cells were established by lentivirus infection, and
decreased levels of p-ATM were demonstrated by reverse
transcription-quantitative polymerase chain reaction and western
blotting. (B) Migration ability of ATM knockout cells was
significantly decreased compared with in the Sc cells, as revealed
by a cell migration assay. Magnification, ×40. (C) EMT of the ATM
knockout cells was significantly decreased, as revealed by reverse
transcription-quantitative polymerase chain reaction. **P<0.01;
***P<0.001 vs. the Sc group. Each experiment was performed in
triplicate. ATM, ataxia telangiectasia mutated kinase;
E-Cad, E-cadherin; EMT, epithelial-mesenchymal transition; N-Cad,
cadherin-N; p-, phosphorylated; Sc, scramble; si, small interfering
RNA; VEGF, vascular endothelial growth factor. |
Expression of ATM is downregulated in
cells treated with an ATM inhibitor, and the proliferation and
migration of such cells are decreased
To validate the aforementioned experimental results,
an ATM inhibitor (CP466722) was incubated with C4-2, CWR22Rv1,
C4-2-Sc and CWR22Rv1-Sc cells. Following 6 h of incubation, a
significant decrease in p-ATM expression was revealed by RT-qPCR
analysis (Fig. 3A). Furthermore,
the proliferative ability of the cells was significantly decreased
3 and 4 days post incubation with CP466722 compared with in the
control group (Fig. 3B). In
addition, the migratory ability of ATM knockout cells and cells
incubated with CP466722 were significantly decreased, thus
suggesting that ATM inhibition may suppress the growth and
metastasis of CRPC cells (Fig.
3C).
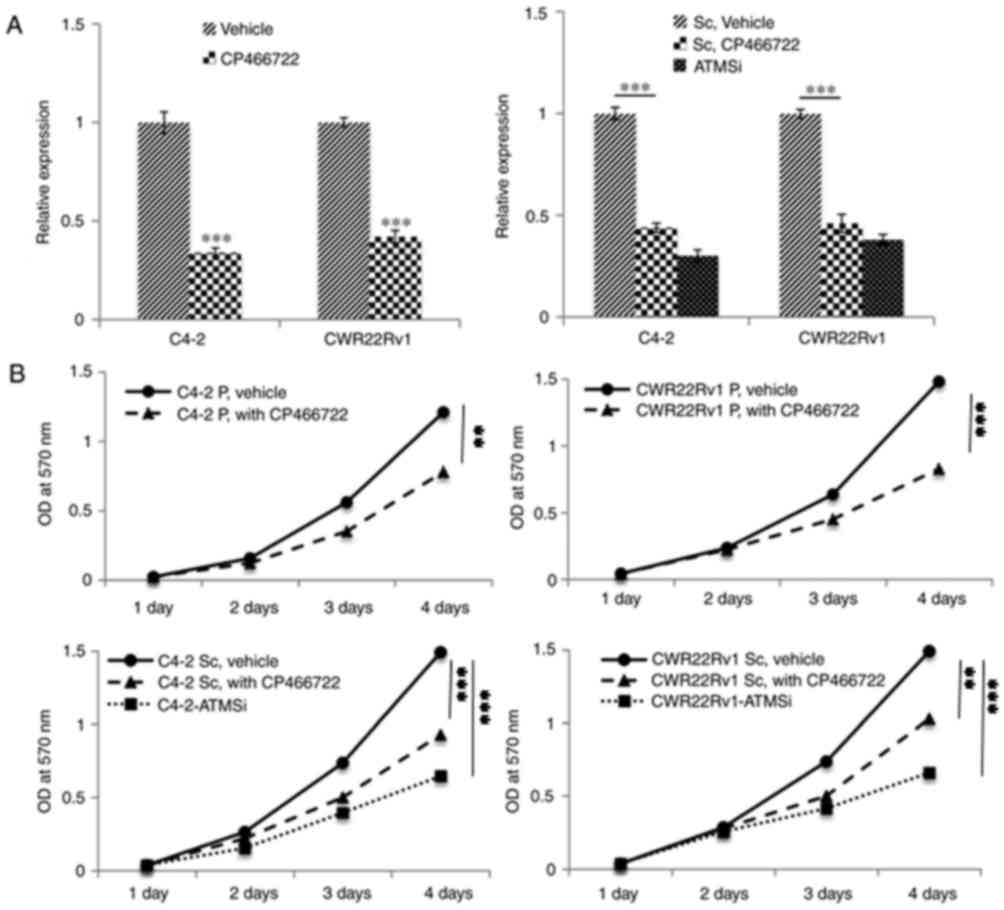 | Figure 3.Following inhibition of ATM
expression in C4-2, CWR22Rv1, C4-2-Sc and CWR22Rv1-Sc cells, the
proliferation and migration of cells is significantly decreased
compared with in the control cells. Similar to ATMSi, addition of
the ATM inhibitor CP466722 significantly inhibits the migration and
proliferation of prostate cancer cells. (A) Decreased p-ATM
expression was revealed by reverse transcription-quantitative
polymerase chain reaction analysis. Compared with ATMSi, the
ability of ATM inhibitor CP466722 to inhibit ATM was not
significantly different. (B) MTT assay demonstrated a significant
decrease in proliferation of the cells on days 3 and 4 compared
with in the control group. (C) Following addition of CP466722, the
migration of cells was significantly suppressed compared with in
the control group. There was no significant difference in the
migration of prostate cancer cells between the Sc, CP466722 and
ATMSi groups. Magnification, ×40. These results further
demonstrated that downregulation of ATM suppresses the migration
and epithelial-mesenchymal transition of CRPC cells. **P<0.01;
***P<0.001. ATM, ataxia telangiectasia mutated kinase; OD,
optical density; P, primary cells; Sc, scramble; si, small
interfering RNA. |
Expression of PD-L1 in ATM knockout
cells is suppressed, and downregulation of the JAK signaling
pathway can inhibit the expression of PD-L1
The aforementioned results demonstrated that ATM has
an important role in CRPC invasion and metastasis via mechanisms
that are not yet fully understood. Therefore, the present study
aimed to investigate the expression of PD-L1, which is closely
associated with EMT (14–16,25).
Firstly, the expression of PD-L1 in C4-2-ATMSi and CWR22Rv1-ATMSi
cells was investigated, and the results demonstrated that compared
with in the control group, the expression of PD-L1 was
significantly suppressed in ATM knockout cells (Fig. 4A). In order to determine the
mechanism underlying the alteration of PD-L1 levels, ATM downstream
regulatory factors were examined, particularly the extensively
studied JAK/STAT3 signaling pathway, which is involved in PD-L1
regulation (26,27). The expression levels of p-JAK1,
p-JAK2 and p-STAT3 in ATM knockout cells were decreased compared
with in the control cells (Fig.
4B). Following 6 h of co-culture with JAK inhibitor 1 or
Stattic, changes in PD-L1 mRNA expression were investigated by
RT-qPCR, the results of which revealed that incubation with JAK
inhibitor 1 resulted in a significant decrease in the expression
levels of PD-L1, whereas incubation with Stattic did not exert a
significant effect (Fig. 4C).
These results suggested that downregulation of the JAK signaling
pathway may inhibit the expression of PD-L1.
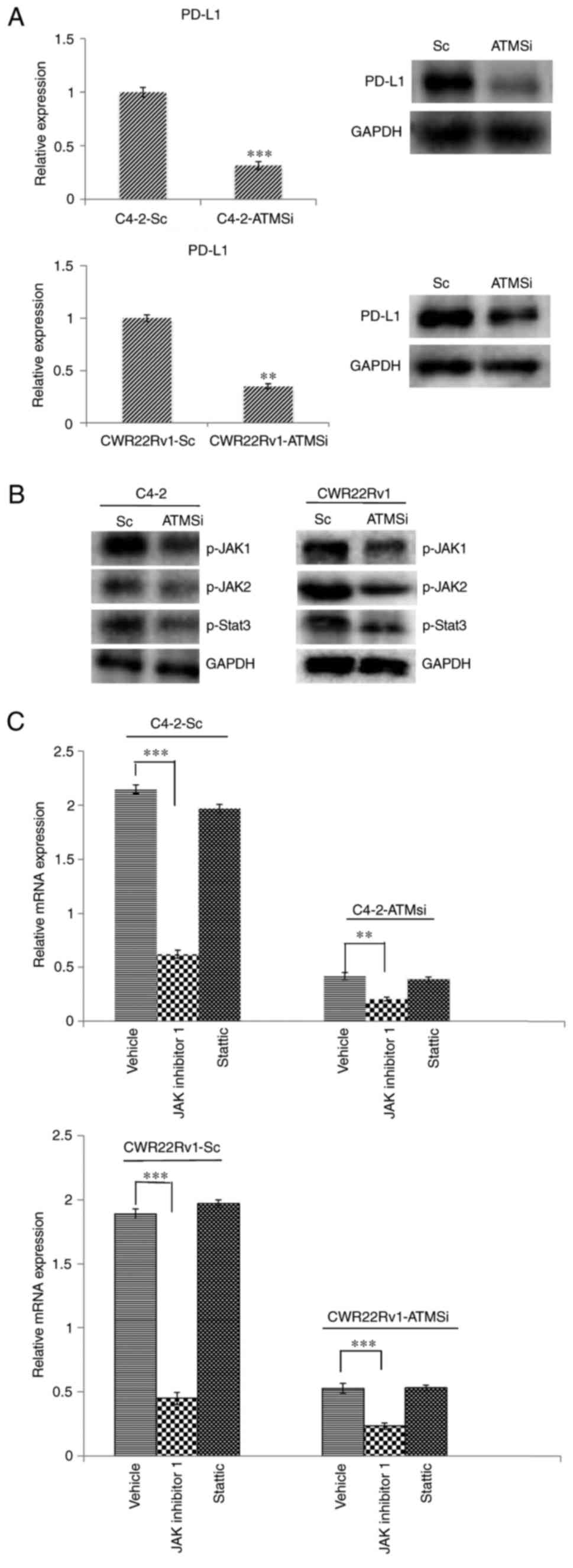 | Figure 4.Levels of PD-L1, p-JAK1, p-JAK2 and
p-STAT3 are suppressed in C4-2-ATMSi and CWR22Rv1-ATMSi cells
compared with in the control cells, and JAK inhibitor 1
significantly suppresses the expression of PD-L1 in ATM knockout
groups and control groups. (A) A significant decrease in PD-L1
expression was revealed in experimental groups by reverse
transcription-quantitative polymerase chain reaction and western
blotting. (B) Decreased levels of p-JAK1, p-JAK2 and p-STAT3 were
revealed in the experimental groups by western blotting. (C) JAK
inhibitor 1 and Stattic were used to treat all cell groups.
Downregulation of JAK significantly reduced PD-L1 expression,
whereas Stattic had no significant effect on PD-L1 expression.
**P<0.01; ***P<0.001. ATM, ataxia telangiectasia mutated
kinase; JAK, Janus kinase; p-, phosphorylated; PD-L1, programmed
cell death 1 ligand 1; Sc, scramble; si, small interfering RNA;
STAT3, signal transducer and activator of transcription 3. |
Suppressed expression of PD-L1 and the
JAK signaling pathway inhibits migration and EMT of CRPC cells, and
thereby inhibits metastasis
To investigate the roles of PD-L1 expression and the
JAK signaling pathway in ATM-associated regulation of CRPC
metastasis, PD-L1 antibodies and JAK inhibitor 1 were incubated
with C4-2-Sc and CWR22Rv1-Sc cells. The results demonstrated that
PD-L1 antibodies and JAK inhibitor 1 significantly decreased the
migration of cells (Fig. 5A).
Furthermore, the results demonstrated that following incubation
with either PD-L1 antibodies or JAK inhibitor 1, the overexpression
of EMT-associated marker genes was effectively normalized (Fig. 5B). These findings suggested that
downregulation of the JAK-PD-L1 signaling pathway may inhibit
metastasis and EMT in CRPC.
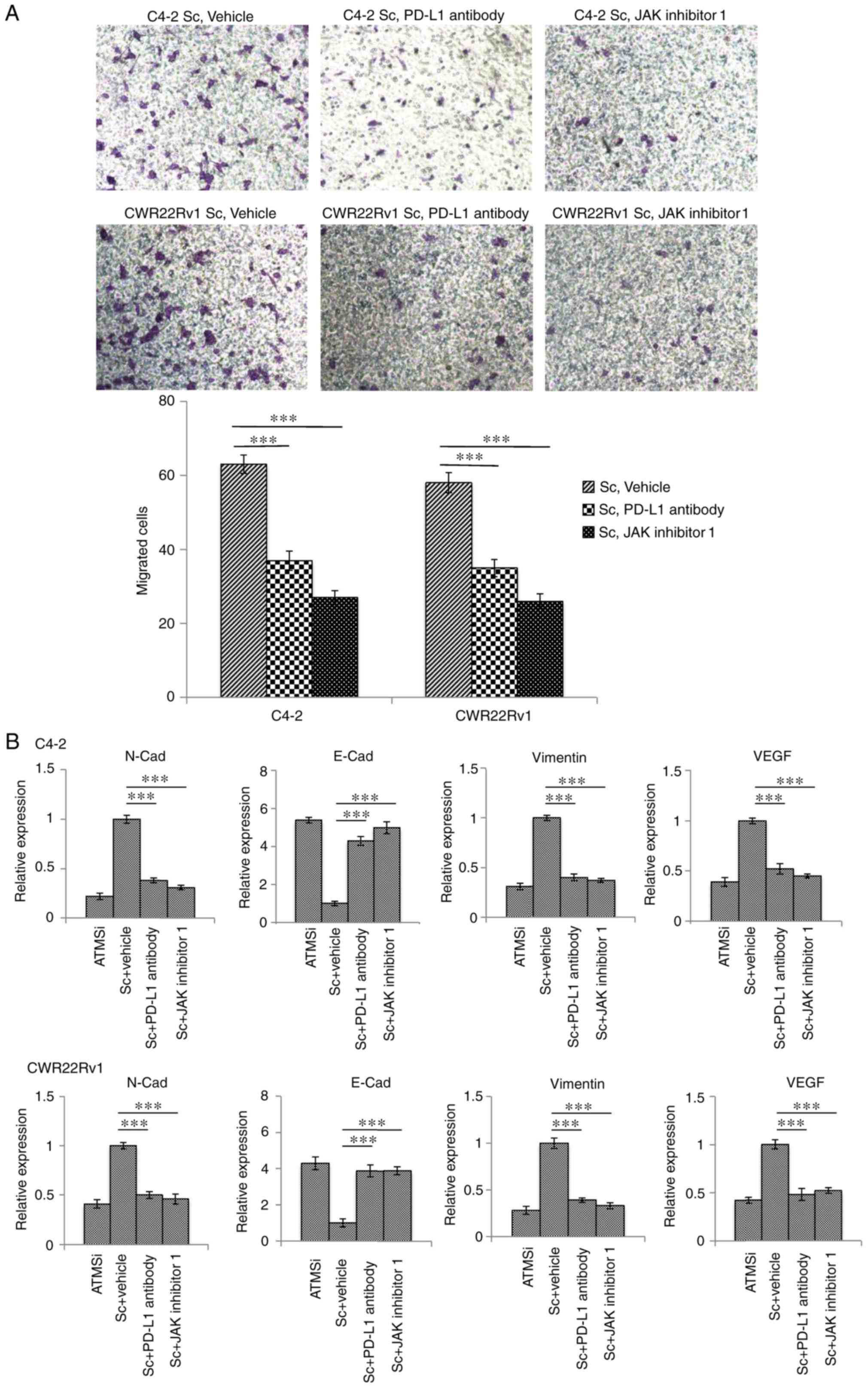 | Figure 5.Downregulation of PD-L1 and JAK
expression inhibits migration and EMT of C4-2-Sc and CWR22Rv1-Sc
cells. C4-2-Sc and CWR22Rv1-Sc cells were treated with PD-L1
antibodies and JAK inhibitor 1. (A) Migration of tumor cells was
significantly decreased following incubation with PD-L1 antibodies
and JAK inhibitor 1. Magnification, ×40. (B) Expression of
EMT-associated markers was reversed, and EMT of the tumor cells was
inhibited. One-way analysis of variance followed by Fisher's least
significant difference post hoc test was used for comparison among
groups. ***P<0.001. ATM, ataxia telangiectasia mutated kinase;
E-Cad, E-cadherin; EMT, epithelial-mesenchymal transition; JAK,
Janus kinase; N-Cad, N-cadherin; PD-L1, programmed cell death 1
ligand 1; Sc, scramble; si, small interfering RNA; VEGF, vascular
endothelial growth factor. |
Discussion
Numerous studies have revealed that ATM has an
important role in cell growth and DNA damage (28–31);
however, the role of ATM in tumor EMT has not been extensively
studied. Suppression of the ATM signaling pathway has been
demonstrated to inhibit ovarian cancer metastasis (24), and in a study of macrophage
paracrine signaling in breast cancer metastasis, downregulation of
ATM reduced the expression of EMT-associated markers in tumor
cells, and thus inhibited metastasis (32). In the present study, it was
revealed that the expression levels of EMT-associated genes were
decreased in ATM gene knockout CRPC cells and the migratory ability
of these cells was suppressed. To further investigate this, ATM
inhibitors were added to primary cells and to Sc cells, and similar
effects were observed. Furthermore, it was revealed that the
proliferative ability of cells was reduced in ATM knockout CRPC
cells on days 3 and 4. Therefore, the results of the present study
suggested that ATM may accelerate the progression of CRPC EMT and
metastasis.
The mechanisms underlying EMT and tumor metastasis
have been extensively studied in recent years. PD-L1 is closely
associated with EMT, invasion and metastasis of breast cancer, lung
cancer, kidney cancer, head and neck tumors, and other malignant
tumors. Furthermore, suppression of PD-L1 expression can inhibit
tumor metastasis (14–16,25).
PD-L1 is highly expressed in invasive prostate cancer and
enzalutamide-resistant prostate cancer (33), and can induce prostate cancer
immune evasion (34). Whether
PD-L1 is a downstream regulatory factor of ATM and whether its
regulation affects the EMT and metastasis of CRPC has not yet been
determined. Therefore, the present study aimed to investigate PD-L1
expression in ATM knockout cells. ATM knockout cells were revealed
to exhibit low PD-L1 expression, thus suggesting that ATM signaling
pathway inhibition suppresses PD-L1 expression.
Considering the aforementioned results, the
mechanisms by which ATM regulates PD-L1 expression and affects the
EMT and metastasis of CRPC cells will be investigated in future
studies. The roles of the JAK/STAT3 signaling pathway in tumor EMT
have been widely studied with regards to prostate cancer, lung
cancer, pancreatic cancer, neck tumors and other solid tumors
(35–38). JAK1/2 inhibitors have also been
revealed to modulate prostate cancer metastasis by regulating
interleukin (IL)-6 in animal experiments and in vitro;
therefore, IL-6 has been suggested to represent a therapeutic
target for the treatment of metastatic prostate cancer (39). Furthermore, previous studies have
demonstrated that the JAK/STAT3 signaling pathway is involved in
PD-L1 regulation (40,41). To investigate whether the JAK/STAT3
signaling pathway is associated with ATM-associated regulation of
PD-L1 expression, and thus whether the JAK/STAT3 signaling pathway
affects the EMT and metastasis of CRPC, the expression levels of
p-JAK1, p-JAK2 and PD-L1 were investigated, and the results
revealed that expression levels of p-JAK1, p-JAK2 and PD-L1were
suppressed in ATM knockout CRPC cells. In addition, inhibition of
the JAK signaling pathway by JAK inhibitor 1 was revealed to
significantly suppress the expression of PD-L1, whereas STAT3
inhibition did not affect the expression of PD-L1. Therefore, it
may be suggested that the JAK signaling pathway is involved in the
regulation of PD-L1 via ATM.
To investigate the effects of PD-L1 and JAK
expression on CRPC cell EMT and migration, PD-L1 antibodies and JAK
inhibitor 1 were used to treatC4-2-Sc and CWR22Rv1-Sc cells; the
results demonstrated that post incubation, the migratory ability of
CRPC cells was significantly decreased and the expression of
EMT-associated markers was effectively normalized. Therefore, these
results revealed that the ATM-JAK-PD-L1 signaling pathway may serve
an important role in the EMT and metastatic progression in
CRPC.
In conclusion, with continuous advances in prostate
cancer androgen deprivation therapy, CRPC has become difficult to
treat clinically and has been extensively studied (42,43).
Targeted therapy is considered a promising therapeutic approach for
numerous tumor types, including prostate cancer (44–46).
The present study aimed to investigate the mechanisms underlying
CRPC cell EMT and metastasis, and it was revealed that inhibition
of the ATM-JAK-PD-L1 signaling pathway may suppress EMT and
metastatic progression of CRPC cells. This may represent a novel
therapeutic approach for targeted therapy for the improvement of
prognosis and quality of life for patients with CRPC.
Acknowledgements
Authors thank Soo Ok Lee for assistance with
manuscript preparation.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81472776) for the
work of Professor Dr. Yang, Dr. Xu and Professor Shan; as well as
the Second Affiliated Hospital of Soochow University preponderant
clinic discipline group project funding (grant no. XKQ2015008).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
LZ, LJX and JZ performed the in vitro and
in vivo experiments and statistical analyses, and made the
figures. JL and B-XX contributed to the generation of knockdown
cell lines. JG and CYS provided and performed staining of human
tissues. YCZ and YBZ helped with the interpretation of data and
reviewed the manuscript. DRY and YXS conceived the idea and wrote
the manuscript. All authors reviewed and agreed to the information
in this manuscript.
Ethics approval and consent to
participate
All patients provided informed consent. The present
study was approved by the Ethics Committee of the Second Affiliated
Hospital of Soochow University (Suzhou, China).
Consent for publication
All patients provided written informed consent for
the publication of any associated data and accompanying images.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Miller KD, Siegel RL, Lin CC, Mariotto AB,
Kramer JL, Rowland JH, Stein KD, Alteri R and Jemal A: Cancer
treatment and survivorship statistics, 2016. CA Cancer J Clin.
66:271–289. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Meng F and Wu G: The rejuvenated scenario
of epithelial-mesenchymal transition (EMT) and cancer metastasis.
Cancer Metastasis Rev. 31:455–467. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang J, Liu D, Feng Z, Mao J, Zhang C, Lu
Y, Li J, Zhang Q, Li Q and Li L: MicroRNA-138 modulates metastasis
and EMT in breast cancer cells by targeting vimentin. Biomed
Pharmacother. 77:135–141. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li C, Wang J, Kong J, Tang J, Wu Y, Xu E,
Zhang H and Lai M: GDF15 promotes EMT and metastasis in colorectal
cancer. Oncotarget. 7:860–872. 2016.PubMed/NCBI
|
|
6
|
Suh SS, Yoo JY, Cui R, Kaur B, Huebner K,
Lee TK, Aqeilan RI and Croce CM: FHIT suppresses
epithelial-mesenchymal transition (EMT) and metastasis in lung
cancer through modulation of microRNAs. PLoS Genet.
10:e10046522014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ombrato L and Malanchi I: The EMT
universe: Space between cancer cell dissemination and metastasis
initiation. Crit Rev Oncog. 19:349–361. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu CH, Tang WC, Sia P, Huang CC, Yang PM,
Wu MH, Lai IL and Lee KH: Berberine inhibits the metastatic ability
of prostate cancer cells by suppressing epithelial-to-mesenchymal
transition (EMT)-associated genes with predictive and prognostic
relevance. Int J Med Sci. 12:63–71. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liang J, Li Y, Daniels G, Sfanos K, De
Marzo A, Wei J, Li X, Chen W, Wang J, Zhong X, et al: LEF1
targeting EMT in prostate cancer invasion is regulated by miR-34a.
Mol Cancer Res. 13:681–688. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kong D, Sethi S, Li Y, Chen W, Sakr WA,
Heath E and Sarkar FH: Androgen receptor splice variants contribute
to prostate cancer aggressiveness through induction of EMT and
expression of stem cell marker genes. Prostate. 75:161–174. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang L, Zhang J, Ma Y, Chen J, Dong B,
Zhao W, Wang X, Zheng Q, Fang F and Yang Y: Testicular orphan
receptor 4 (TR4) is a marker for metastasis and poor prognosis in
non-small cell lung cancer that drives the EMT phenotype. Lung
Cancer. 89:320–328. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ma X, Yan W, Dai Z, Gao X, Ma Y, Xu Q,
Jiang J and Zhang S: Baicalein suppresses metastasis of breast
cancer cells by inhibiting EMT via downregulation of SATB1 and
Wnt/β-catenin pathway. Drug Des Devel Ther. 10:1419–1441. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huan H, Wen X, Chen X, Wu L, Liu W, Habib
NA, Bie P and Xia F: C/EBPα short-activating RNA suppresses
metastasis of hepatocellular carcinoma through inhibiting
EGFR/β-catenin signaling mediated EMT. PLoS One. 11:e01531172016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Alsuliman A, Colak D, Al-Harazi O, Fitwi
H, Tulbah A, Al-Tweigeri T, Al-Alwan M and Ghebeh H: Bidirectional
crosstalk between PD-L1 expression and epithelial to mesenchymal
transition: Significance in claudin-low breast cancer cells. Mol
Cancer. 14:1492015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang Y, Wang H, Zhao Q, Xia Y, Hu X and
Guo J: PD-L1 induces epithelial-to-mesenchymal transition via
activating SREBP-1c in renal cell carcinoma. Med Oncol. 32:2122015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ock CY, Kim S, Keam B, Kim M, Kim TM, Kim
JH, Jeon YK, Lee JS, Kwon SK, Hah JH, et al: PD-L1 expression is
associated with epithelial-mesenchymal transition in head and neck
squamous cell carcinoma. Oncotarget. 7:15901–15914. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wen W, Liang W, Wu J, Kowolik CM, Buettner
R, Scuto A, Hsieh MY, Hong H, Brown CE, Forman SJ, et al: Targeting
JAK1/STAT3 signaling suppresses tumor progression and metastasis in
a peritoneal model of human ovarian cancer. Mol Cancer Ther.
13:3037–3048. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chang Q, Bournazou E, Sansone P, Berishaj
M, Gao SP, Daly L, Wels J, Theilen T, Granitto S, Zhang X, et al:
The IL-6/JAK/Stat3 feed-forward loop drives tumorigenesis and
metastasis. Neoplasia. 15:848–862. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sansone P and Bromberg J: Targeting the
interleukin-6/Jak/stat pathway in human malignancies. J Clin Oncol.
30:1005–1014. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao B, Shen X, Kunos G, Meng Q, Goldberg
ID, Rosen EM and Fan S: Constitutive activation of JAK-STAT3
signaling by BRCA1 in human prostate cancer cells. FEBS Lett.
488:179–184. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Teng Y, Ghoshal P, Ngoka L, Mei Y and
Cowell JK: Critical role of the WASF3 gene in JAK2/STAT3 regulation
of cancer cell motility. Carcinogenesis. 34:1994–1999. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tam L, McGlynn LM, Traynor P, Mukherjee R,
Bartlett JM and Edwards J: Expression levels of the JAK/STAT
pathway in the transition from hormone-sensitive to
hormone-refractory prostate cancer. Br J Cancer. 97:378–383. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sun M, Guo X, Qian X, Wang H, Yang C,
Brinkman KL, Serrano-Gonzalez M, Jope RS, Zhou B, Engler DA, et al:
Activation of the ATM-Snail pathway promotes breast cancer
metastasis. J Mol Cell Biol. 4:304–315. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yin S, Wang P, Yang L, Liu Y, Wang Y, Liu
M, Qi Z, Meng J, Shi TY, Yang G and Zang R: Wip1 suppresses ovarian
cancer metastasis through the ATM/AKT/Snail mediated signaling.
Oncotarget. 7:29359–29370. 2016.PubMed/NCBI
|
|
25
|
Kim S, Koh J, Kim MY, Kwon D, Go H, Kim
YA, Jeon YK and Chung DH: PD-L1 expression is associated with
epithelial-to-mesenchymal transition in adenocarcinoma of the lung.
Hum Pathol. 58:7–14. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Doi T, Ishikawa T, Okayama T, Oka K,
Mizushima K, Yasuda T, Sakamoto N, Katada K, Kamada K, Uchiyama K,
et al: The JAK/STAT pathway is involved in the upregulation of
PD-L1 expression in pancreatic cancer cell lines. Oncol Rep.
37:1545–1554. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bellucci R, Martin A, Bommarito D, Wang K,
Hansen SH, Freeman GJ and Ritz J: Interferon-γ-induced activation
of JAK1 and JAK2 suppresses tumor cell susceptibility to NK cells
through upregulation of PD-L1 expression. Oncoimmunology.
4:e10088242015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Subhash VV, Tan SH, Yeo MS, Yan FL,
Peethala PC, Liem N, Krishnan V and Yong WP: ATM expression
predicts veliparib and irinotecan sensitivity in gastric cancer by
mediating P53-independent regulation of cell cycle and apoptosis.
Mol Cancer Ther. 15:3087–3096. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Khoronenkova SV and Dianov GL: ATM
prevents DSB formation by coordinating SSB repair and cell cycle
progression. Proc Natl Acad Sci USA. 112:pp. 3997–4002. 2015;
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wojewoda M, Walczak J, Duszyński J and
Szczepanowska J: Selenite activates the ATM kinase-dependent DNA
repair pathway in human osteosarcoma cells with mitochondrial
dysfunction. Biochem Pharmacol. 95:170–176. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
McCabe N, Hanna C, Walker SM, Gonda D, Li
J, Wikstrom K, Savage KI, Butterworth KT, Chen C, Harkin DP, et al:
Mechanistic rationale to target PTEN-deficient tumor cells with
inhibitors of the DNA damage response kinase ATM. Cancer Res.
75:2159–2165. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Singh R, Shankar BS and Sainis KB:
TGF-β1-ROS-ATM-CREB signaling axis in macrophage mediated migration
of human breast cancer MCF7 cells. Cell Signal. 26:1604–1615. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bishop JL, Sio A, Angeles A, Roberts ME,
Azad AA, Chi KN and Zoubeidi A: PD-L1 is highly expressed in
Enzalutamide resistant prostate cancer. Oncotarget. 6:234–242.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Martin AM, Nirschl TR, Nirschl CJ,
Francica BJ, Kochel CM, van Bokhoven A, Meeker AK, Lucia MS, Anders
RA, DeMarzo AM and Drake CG: Paucity of PD-L1 expression in
prostate cancer: Innate and adaptive immune resistance. Prostate
Cancer Prostatic Dis. 18:325–332. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Aalinkeel R, Hu Z, Nair BB, Sykes DE,
Reynolds JL, Mahajan SD and Schwartz SA: Genomic analysis
highlights the role of the JAK-STAT signaling in the
anti-proliferative effects of dietary flavonoid-‘Ashwagandha’ in
prostate cancer cells. Evid Based Complement Alternat Med.
7:177–187. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu RY, Zeng Y, Lei Z, Wang L, Yang H, Liu
Z, Zhao J and Zhang HT: JAK/STAT3 signaling is required for
TGF-β-induced epithelial-mesenchymal transition in lung cancer
cells. Int J Oncol. 44:1643–1651. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Macha MA, Rachagani S, Gupta S, Pai P,
Ponnusamy MP, Batra SK and Jain M: Guggulsterone decreases
proliferation and metastatic behavior of pancreatic cancer cells by
modulating JAK/STAT3 and Src/FAK signaling. Cancer Lett.
341:166–177. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yadav A, Kumar B, Datta J, Teknos TN and
Kumar P: IL-6 promotes head and neck tumor metastasis by inducing
epithelial-mesenchymal transition via the JAK-STAT3-SNAIL signaling
pathway. Mol Cancer Res. 9:1658–1667. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gu L, Talati P, Vogiatzi P, Romero-Weaver
AL, Abdulghani J, Liao Z, Leiby B, Hoang DT, Mirtti T, Alanen K, et
al: Pharmacologic suppression of JAK1/2 by JAK1/2 inhibitor AZD1480
potently inhibits IL-6-induced experimental prostate cancer
metastases formation. Mol Cancer Ther. 13:1246–1258. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xu L, Shen M, Chen X, Zhu R, Yang DR, Tsai
Y, Keng PC, Chen Y and Lee SO: Adipocytes affect
castration-resistant prostate cancer cells to develop the
resistance to cytotoxic action of NK cells with alterations of
PD-L1/NKG2D ligand levels in tumor cells. Prostate. 78:353–364.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang N, Zeng Y, Du W, Zhu J, Shen D, Liu
Z and Huang JA: The EGFR pathway is involved in the regulation of
PD-L1 expression via the IL-6/JAK/STAT3 signaling pathway in
EGFR-mutated non-small cell lung cancer. Int J Oncol. 49:1360–1368.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee DJ, Cha EK, Dubin JM, Beltran H,
Chromecki TF, Fajkovic H, Scherr DS, Tagawa ST and Shariat SF:
Novel therapeutics for the management of castration-resistant
prostate cancer (CRPC). BJU Int. 109:968–985. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chandrasekar T, Yang JC, Gao AC and Evans
CP: Mechanisms of resistance in castration-resistant prostate
cancer (CRPC). Transl Androl Urol. 4:365–380. 2015.PubMed/NCBI
|
|
44
|
Kübler H and Miller K: New therapy
concepts for castration-resistant prostate cancer: Between hormone
manipulation, targeted therapy and chemotherapy. Urologe A.
52(1517–1518): 1520–1521, 1524–1526. 2013.(In German).
|
|
45
|
Qi WX, Fu S, Zhang Q and Guo XM: Efficacy
and toxicity of molecular targeted therapies in combination with
docetaxel for metastatic castration-resistant prostate cancer: A
meta-analysis of phase III randomized controlled trials. J
Chemother. 27:181–187. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Loriot Y, Eymard JC, Patrikidou A, Ileana
E, Massard C, Albiges L, Di Palma M, Escudier B and Fizazi K: Prior
long response to androgen deprivation predicts response to
next-generation androgen receptor axis targeted drugs in castration
resistant prostate cancer. Eur J Cancer. 51:1946–1952. 2015.
View Article : Google Scholar : PubMed/NCBI
|