Introduction
With a growing epidemic, chronic heart failure (CHF)
results in an economic burden worldwide and has become a major
factor causing disabilities and mortality (1,2). CHF
is characterized by progressive cardiac dysfunction and remodeling
(3,4). Volume and pressure overload are the
most common promoters in pathological process of CHF (5). Cellular and molecular mechanisms are
involved in the deterioration of CHF, including inflammation,
oxidative stress and fibrosis but the underlying mechanisms are not
fully understood (6).
Curcumin [1,7-bis
(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione] is the major
active component of turmeric isolated from the rhizome of
Zingiberaceae and has widely been studied as an anticancer agent
(7,8). Curcumin is a powerful antioxidant and
anti-inflammatory reagent (9), as
well as a modulator of protein kinase, DNA methyltransferase and
histone acetyltransferase (10).
Curcumin is closely associated with the expression and activity of
various regulatory proteins (11).
Several studies have reported the protective effects of curcumin on
cardiovascular diseases, including protection against cardiac
dysfunction and remodeling in heart failure in rats (8,12,13).
However, the mechanism by which curcumin exerts cardioprotective
effects has not been fully understood.
Dickkopf-related protein 3 (DKK-3), a member of the
Dickkopf glycoprotein family, is recognized as a negative regulator
of many tumors and is crucially involved in cell growth,
proliferation and immunity (14).
DKK-3 is abundantly expressed in skeletal muscle and heart
(15,16). DKK-3 expression was decreased in
hypertrophic murine heart and human dilated cardiomyopathy heart
(5). In animal models, cardiac
dysfunction and remodeling induced by pressure overload can be
exacerbated by DKK-3 depletion and can be rescued by DKK-3
overexpression (5). Our previous
study indicated that DKK-3 served a cardioprotective role in the
pathology of CHF by inhibiting p38 mitogen-activated protein kinase
(p38) and c-JunN-terminal kinase (JNK) signaling pathways in an
apoptosis signal-regulating kinase 1 (ASK1)-dependent way (16). DKK-3 may be the downstream target
of curcumin owing to its similar cardioprotective effects in heart
failure. The present study was designed to investigate whether
curcumin exert cardiac protective effects by regulating DKK-3 and
try to clarify whether p38 and JNK pathways are involved in the
underlying molecular mechanisms.
Materials and methods
Animal preparation
A total of 40 adult New Zealand rabbits (male,
weighing 2–2.5 kg, ~12-week-old) were kept in the breeding facility
for one week prior to experimental use. Rabbits were singly housed
in animal holding cages at temperature (22±2°C) and humidity
(45–50%), with a 12-h light/dark cycle and unrestricted access to
food and water. Rabbits were randomized into four groups: Control
rabbits given with placebo (Con, n=10) or curcumin (Con-cur, n=10),
chronic heart failure rabbits fed with placebo (CHF, n=10) or
curcumin (CHF-cur, n=10). Placebo or curcumin were fed three days
prior to the pressure overload surgery and this continued until the
day of sacrifice. The dose of curcumin (Hebei Food Additive Co.,
Ltd. Shijiazhuang, Hebei, China) and placebo (Kangxin Co., Ltd.,
Shijiazhuang, Hebei, China) both were 100 mg/day/kg in capsules,
which were chosen with reference to a previous study (17). The placebo was made of starch and
was contained in an identical capsule to that of curcumin. All
procedures were according to the ‘Guide for the Care and Use of
Laboratory Animals’ published by the US National Institutes of
Health (NIH Publication no. 85-23; revised 1996) and approved by
the Institutional Animal Care and Use Committee at Renmin Hospital
of Wuhan University (Wuhan, China).
CHF production
CHF was induced by ten weeks volume overload and
eight weeks pressure overload as previously described with slight
modifications (18). Prior to each
surgery, all rabbits were anesthetized with pentobarbital sodium
(30 mg/kg) by intravenous injection. Volume overload was produced
by puncturing across the aortic valve through carotid artery
retrogradely with a catheter (external diameter 1.30 mm). The
catheter puncturing across the aortic valve was repeated until the
pulse pressure increased by ~100%. Two weeks following this,
pressure overload was produced by ligating the abdominal aorta
partly just above renal artery branch out. This resulted in
abdominal aorta stenosis by ~50% in CHF rabbits. Rabbits in the Con
and Con-cur groups underwent a sham operation, but neither aortic
valve regurgitation nor aorta stenosis was produced.
Cardiac function and structure
evaluation by echocardiography
Transthoracic echocardiography (10S transducer,
Vivid 7, GE Healthcare, Chicago, IL, USA) was performed at baseline
and eight weeks following the aorta stenosis surgery. Left
ventricular ejection fraction (LVEF), left ventricular fractional
shortening (LVFS), aortic diameter (AO), left ventricular posterior
wall thickness (LVPW) and interventricular septal thickness (IVS)
were assessed by echocardiography.
Heart weight and cardiac
histology
Following the echocardiography, all rabbits were
sacrificed and tissue samples from left ventricular (LV) were
prepared for the following analysis. LV samples were fixed in 4%
paraformaldehyde solution at room temperature for 24 h and then
were sliced into 5-µm sections transversely. Haematoxylin &
eosin (H&E), masson (G1006, Servicebio, Inc., Wuhan, China),
picrosirius red (PSR) (G1018, Servicebio, Inc.) staining and
terminal deoxynucleotidyl transferase-mediated dUTP nick end
labeling (TUNEL) assay (In Situ Cell Death Detection kit; cat. no.
11684817910; Roche Diagnostics, Basel, Switzerland) were performed.
In H&E staining, the slides were successively stained with
hematoxylin solution and eosin solution at room temperature for 5
min respectively. In masson staining, the slides were successively
stained with iron hematoxylin solution, acid ponceau fuchsin
solution and aniline blue solution at room temperature for 5 min
each respectively. In PSR staining, the slides were stained with
sirius red solution at room temperature for 10 min. In TUNEL
staining, slides of LV were first incubated with the TUNEL reaction
mixture containing terminal deoxynucleotidyl transferase (TdT) at
37°C for 60 min. Subsequently, diaminobenzidine was added to the
slides for 30 min and incubated in substrate solution for an
additional 15 min at 37°C. The cross-sectional area (CSA) of LV
myocytes was analyzed based on H&E staining, LV interstitial
collagen volume (%) was evaluated based on both Masson and PSR
staining by Image-Pro Plus 6.0 software (Media Cybernetics, Inc.,
Rockville, MD, USA). The cardiomyocyte apoptosis index was defined
as the proportion of apoptotic cardiomyocytes among the total
cardiomyocytes (apoptotic cardiomyocytes had deep-brown nuclei
while normal cardiomyocytes had blue nuclei).
Ultrastructure analysis
For cardiac ultrastructure analysis, LV samples were
immediately soaked in pre-cooled Ca2+-free phosphate
saline buffer (G1102; Servicebio, Inc.). The samples were
successively fixed with 2.5% glutaraldehyde and 1% osmium tetroxide
at 4°C for 2 h. Then the samples were dehydrated by ethanol of
gradient dilution (30, 50 and 70%) at 4°C for 10 min respectively,
later the samples were further dehydrated using an acetone of
gradient dilution (80, 90 and 100%) at room temperature for 10 min.
Samples were sliced into 50 nm section, embedded in the Epon resin
and then stained with uranyl acetate and lead citrate at 40°C for 4
h. Sections were imaged under a transmission electron microscope
(HT7700; Hitachi, Ltd., Tokyo, Japan).
Inflammation, hypertrophy and fibrosis
biomarkers analysis
Reactive oxygen species (ROS) production of LV
myocytes was detected by dihydroethidium (DHE; 10 mmol/l; D7008;
Sigma-Aldrich; Mecrk KGaA, Darmstadt, Germany) staining.
Cryosections (10-µm) were treated for 30 min at 37°C, and the
ethidium fluorescence (excitation/emission at 488/610 nm) was
examined by digital fluorescence microscopy (Eclipse Ti-SR; Nikon
Corporation, Tokyo, Japan). The relative fluorescence intensity was
analyzed by Image-Pro Plus 6.0 software (Media Cybernetics, Inc.).
Immunohistochemical staining was performed as described previously
(19). LV sections were incubated
overnight at 4°C with primary antibodies respectively as follows:
Mouse monoclonal anti-tumor necrosis factor-alpha (TNF-α) antibody
(1:200; cat. no. NB600-1422; Novus Biologicals, Ltd., Cambridge,
UK), mouse monoclonal anti-matrix metallopeptidase-2 (MMP-2)
antibody (1:200; cat. no. ab2462; Abcam, Cambridge, UK), mouse
monoclonal anti-matrix metallopeptidase-9 (MMP-9) antibody (1:200;
cat. no. ab58803; Abcam) and mouse monoclonal anti-sarcoplasmic
reticulum Ca2+ ATPase (SERCA2a) antibody (1:200; cat.
no. ab2817; Abcam). Following washes with PBS three times, sections
were incubated with the rabbit anti-mouse immunoglobulin (Ig)
M/horseradish peroxidase (HRP) secondary antibody (1:200; cat. no.
bs-0368R-HRP; BIOSS, Beijing, China). A total of five fields of
view (magnification, ×200) were selected randomly for each slide
under a light microscope (BX50, Olympus Corporation, Tokyo, Japan)
and the relative mean optical density of ROS or each target protein
was analyzed by Image-Pro Plus 6.0 software (Media Cybernetics,
Inc.).
Detection of DKK-3, p38, JNK and ASK1
expression
Immunohistochemical staining for DKK-3 was performed
as described above. Primary rabbit polyclonal anti-DKK-3 antibody
(1:200; cat. on. bs-2686R; BIOSS) and secondary antibody mouse
anti-rabbit IgM/HRP (1:200, bs-0369M-HRP, BIOSS) were used in the
staining. Western blotting was performed by using extracts from LV
tissue homogenates. Radio-immunoprecipitation assay lysis buffer
(G2002 Servicebio, Inc.) was used for protein extraction. The
protein concentration of the LV tissue homogenates was measured by
Pierce BCA Protein Assay kit (cat. no. 23225; Pierce; Thermo Fisher
Scientific, Inc. Waltham, MA, USA) according to the manufacturer's
protocol following centrifugation (4°C, 10 min, 15,000 × g).
SDS-PAGE gel wells were loaded with 50 µg of corresponding protein
for electrophoresis. The proteins were transferred to a
polyvinylidene fluoride membrane. The blots were incubated with the
primary antibodies overnight at 4°C as follows: Rabbit polyclonal
anti-DKK-3 antibody (1:1,000; cat. no. bs-2686R), rabbit polyclonal
anti-p38 antibody (1:1,000; cat. no. bs-0637R), rabbit polyclonal
anti-phospho-p38 (Thr180+Tyr182) antibody (p-p38; 1:1,000; cat. no.
bs-0636R), rabbit polyclonal anti-JNK (1:1,000; cat. no.
bs-10562R), rabbit anti-phospho-JNK (Thr183+Tyr185) antibody
(P-JNK; 1:1,000; cat. no. bs-1640R), rabbit polyclonal anti-ASK1
(1:1,000; cat. no. bs-1425R), rabbit anti-phospho-ASK1 (Ser1033)
antibody (P-ASK1; 1:1,000; cat. no. bs-5437R) and antibody against
GAPDH (1:5,000; cat. no. bs-2188R). Subsequently, the membrane was
incubated with the mouse anti-rabbit IgM/HRP secondary antibody
(1:5,000; cat. no. bs-0369M-HRP) for 1 h at room temperature. All
antibodies used in western blot analysis were purchased from BIOSS.
Visualization reagent (ECL; cat. no. G2014; Servicebio, Inc.) was
used in the western blotting. Membranes were analyzed by Quantity
One Imaging system (version 4.6.9; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The protein expression was normalized to that
of GAPDH.
Statistical analysis
Quantitative data are presented as the mean ±
standard deviation. Comparison between preoperative and
postoperative parameters was performed by matched Student's t-test.
Multi-group comparisons were examined via one-way analysis of
variance followed by a Student-Newman-Keuls post hoc test. The
statistical differences were examined by the statistical software
SPSS package v22.0 (IBM Corp., Armonk, NY, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Curcumin improves cardiac
function
The present study aimed to determine the effect of
curcumin on chronic heart failure and whether curcumin improved
cardiac function by observed symptoms and echocardiography. All
rabbits suffered from evident dyspnea and edema in the CHF group,
and six rabbits suffered from cough in the CHF-cur group. Based on
the echocardiography data (Table
I), LVEF and LVFS of the CHF group were significantly lower,
and AO, LVPW and IVS of the same group were significantly higher
compared with the Con group. The above results indicate that ten
weeks volume and eight weeks pressure overload produced CHF
successfully in the CHF and CHF-cur groups. Additionally, ten weeks
treatment with curcumin significantly alleviated the symptoms and
echocardiographic abnormalities in CHF-cur group (Table I).
 | Table I.Effects of curcumin on cardiac
function and morphology. |
Table I.
Effects of curcumin on cardiac
function and morphology.
| Group | Time | LVEF (%) | LVFS (%) | AO (mm) | LVPW (mm) | IVS (mm) |
|---|
| Con | Pre |
74.2±3.8 |
39.9±3.3 |
7.1±0.2 |
2.1±0.2 |
2.1±0.1 |
|
| Post |
74.0±3.4b |
39.8±2.9b |
7.1±0.3b |
2.2±0.1b |
2.2±0.2b |
| Con-cur | Pre |
71.4±8.9 |
38.0±7.4 |
7.1±0.6 |
2.2±0.4 |
2.1±0.3 |
|
| Post |
70.6±7.2 |
37.2±5.6 |
7.2±0.5 |
2.2±0.5 |
2.2±0.3 |
| CHF | Pre |
72.1±6.0 |
38.2±5.0 |
7.2±0.40 |
2.2±0.3 |
2.1±0.4 |
|
| Post |
32.1±8.8a,c |
14.1±4.5a,c |
10.3±1.5a,c |
3.1±0.4a,c |
4.1±0.8a,c |
| CHF-cur | Pre |
70.2±5.3 |
36.7±4.1 |
7.2±0.4 |
2.0±0.2 |
2.1±0.2 |
|
| Post |
48.3±12.9a,d |
23.0±7.2a,d |
8.7±0.5a,d |
2.6±0.3a,d |
3.1±0.3a,d |
Curcumin inhibits heart
remodeling
CHF is characterized by both cardiac dysfunction and
cardiac remodeling (20). The
effects of curcumin on cardiac remodeling was evaluated by cardiac
morphology, ultrastructure and histology. Heart/body weight ratio
of the CHF group was higher than the Con group, and curcumin
treatment decreased the ratio in the CHF-cur group compared with
the CHF group (Fig. 1A). H&E
staining demonstrated that average CSA of LV cardiomyocyte of CHF
group was higher than those of Con group significantly, and
curcumin ameliorated myocardial hypertrophy (Fig. 1B H&E and C). Compared with the
Con group, the ultrastructure of the CHF group demonstrated
disarranged myofibrils, swelled mitochondrial and diminished
crista. Curcumin partly alleviated the ultrastructure
disorganization in the CHF-cur group (Fig. 1B Ultrastructure). Marked cardiac
fibrosis detected by Masson or PSR staining was demonstrated in the
CHF group, and curcumin alleviated fibrosis in the CHF-cur group
(Fig. 1D MASSON, 1D PSR, 1E and
1F). Higher apoptosis of cardiomyocyte detected by TUNEL assay was
demonstrated in the CHF group, and curcumin ameliorated this effect
in the CHF-cur group (Fig. 1D
TUNEL and G).
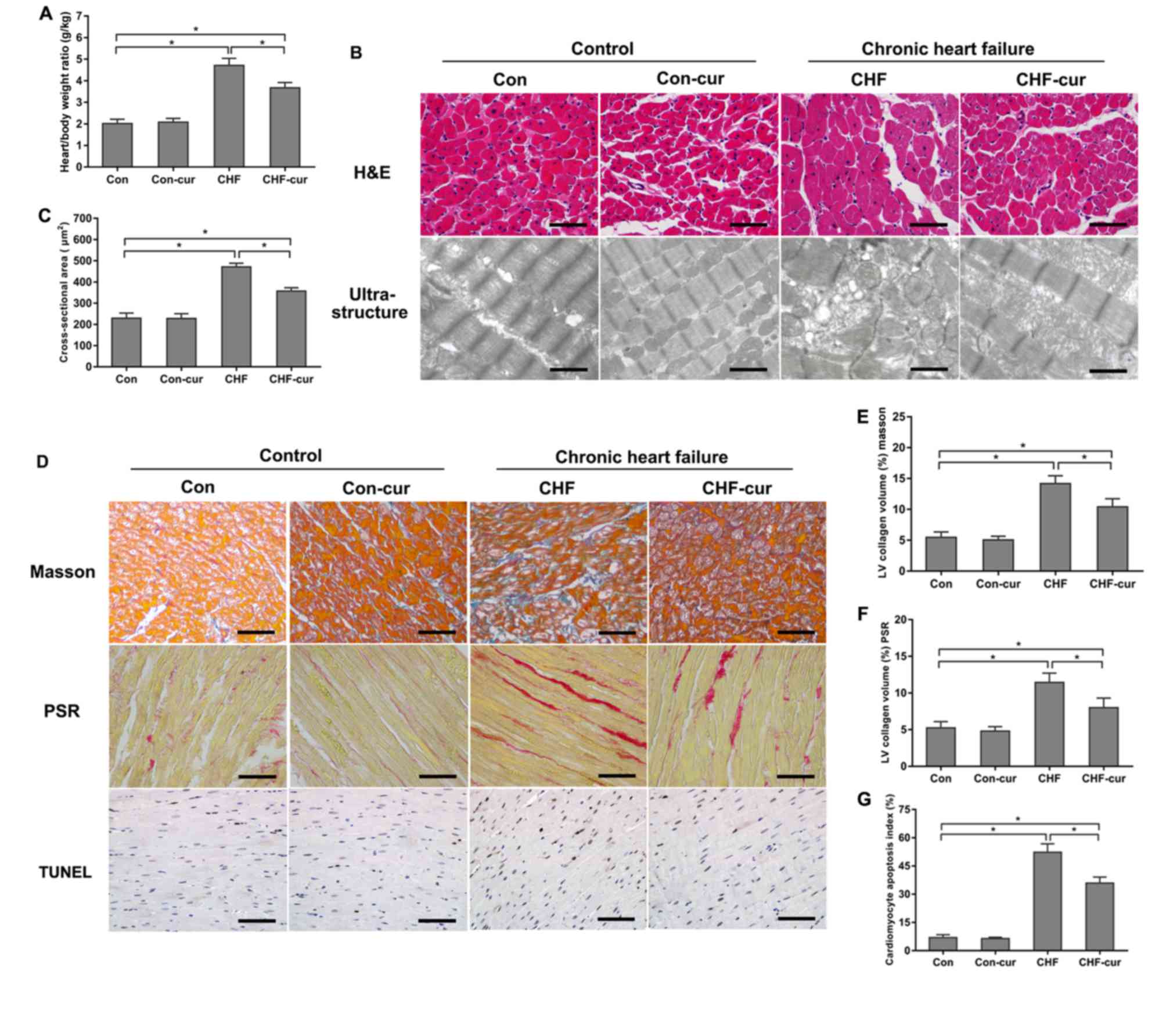 | Figure 1.Effects of curcumin on cardiac
morphology and histology. (A) Effects of curcumin on heart/body
weight ratio. (B) Representative images of H&E staining
(magnification, ×200; scale bar=50 µm) and ultrastructure of LVAW
cardiomyocytes (magnification, ×2,000; scale bar=3 µm). (C) Effects
of curcumin on LVAW cardiomyocytes cross-sectional area. (D)
Representative images of LVAW, measured by Masson, picrosirius red
staining and TUNEL assay (magnification, ×200; scale bar=50 µm).
Quantification of LVAW collagen volume (%) detected by (E) masson
staining, (F) picrosirius red staining. (G) Quantification of
cardiomyocyte apoptosis index detected by TUNEL assay. All data are
expressed as the mean ± stadard deviation, n=10. *P<0.05. CHF,
chronic heart failure; Con, control; Cur, curcumin; LVAW, left
ventricular posterior wall thickness; TUNEL, terminal
deoxynucleotidyl transferase-mediated dUTP nick end labeling;
H&E, haematoxylin & eosin. |
Curcumin regulates the expression of
certain CHF molecular biomarkers
Curcumin was demonstrated to improve cardiac
function and inhibit heart remodeling. The effects of curcumin on
the biomarkers above, including ROS, TNF-α, MMP-2 and MMP-9, were
investigated. In the sample images of DHE staining, red granules in
cytoplasm and cell nucleus indicated positive staining of ROS
(Fig. 2A ROS) and yellow granules
in cytoplasm indicated the positive staining of the target proteins
in the sample images of immunohistochemical staining (Fig. 2A TNF-α, 2D MMP-2, 2D MMP-9 and 2G
SERCA2a). Quantitative analysis of the DHE staining and
immunohistochemical staining indicated that expression of ROS,
TNF-α, MMP-2 and MMP-9 were increased in the CHF group compared
with the Con group but these proteins were decreased in the CHF-cur
group (Fig. 2A-F). Compared with
the Con group, the expression of SERCA2a decreased in the CHF
group, and curcumin increased its expression in the CHF-cur group
(Fig. 2G and H).
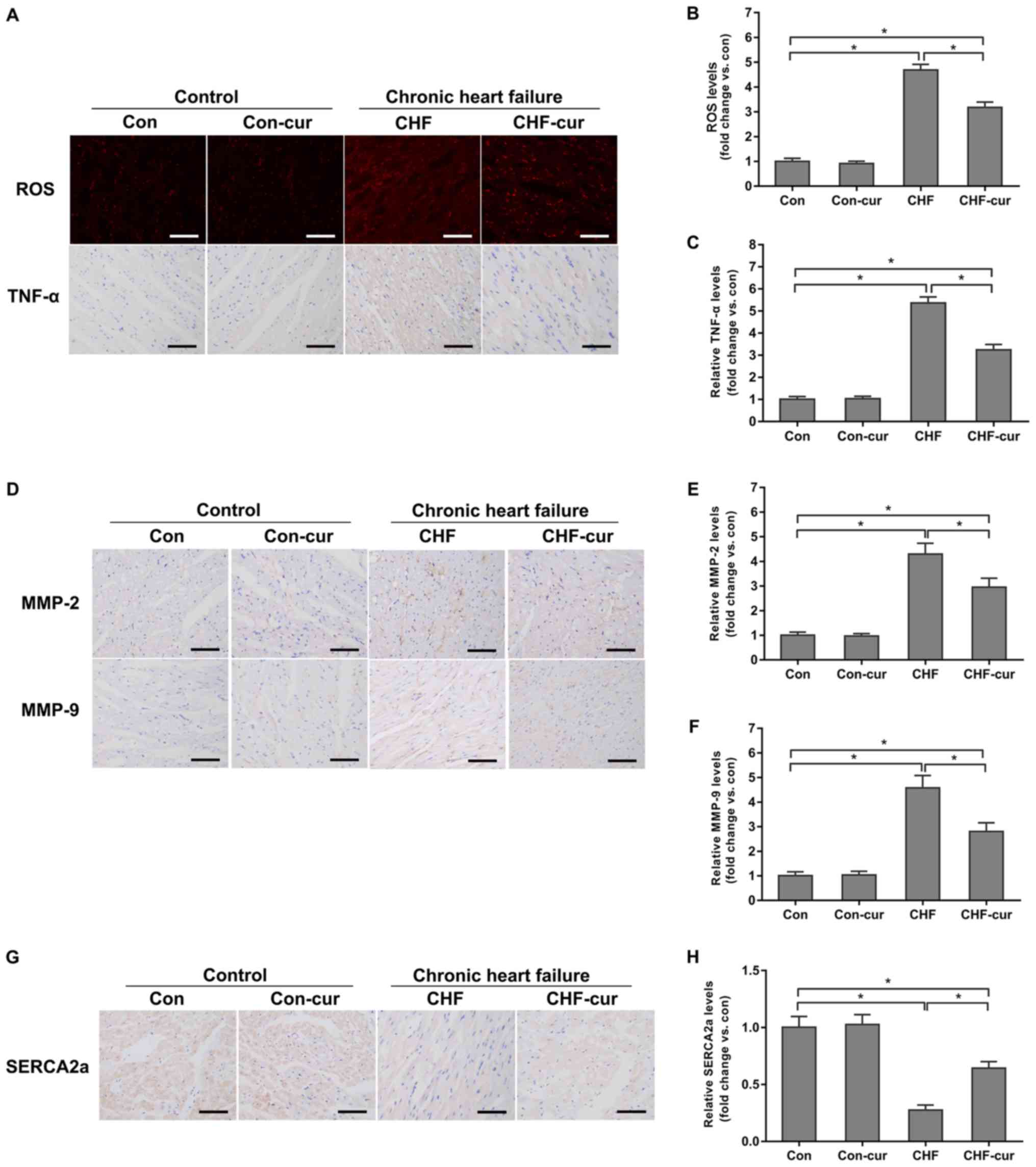 | Figure 2.Effects of curcumin on the expression
of CHF molecular biomarkers. (A) Representative images of ROS and
TNF-α staining and quantification of (B) ROS and of (C) TNF-α
expression levels, as measured by DHE (ROS) and immunohistochemical
staining (TNF-α) in LVAW cardiomyocytes. (D) Representative images
of MMP-2 and MMP-9 and quantification of (E) MMP-2 and (F) MMP-9
expression levels, as measured by immunohistochemical staining in
LVAW cardiomyocytes. (G) Representative images of SERCA2a and (H)
quantification of its expression levels, as measured by
immunohistochemical staining in LVAW cardiomyocytes. Magnification,
×200. Scale bar=50 µm. All data are expressed as the mean ± stadard
deviation, n=10. *P<0.05. CHF, chronic heart failure; Con,
control; Cur, curcumin; LVAW, left ventricular posterior wall
thickness; MMP, matrix metalloproteinase; ROS, reactive oxygen
species; SERCA, sarcoplasmic reticulum Ca2+ ATPase; TNF,
Tumor necrosis factor; DHE, dihydroethidium. |
Curcumin increases expression of
DKK-3
Previous studies demonstrated that DKK-3
down-regulation contributed to the stress-induced or
infarction-induced dysfunction and remodeling in CHF (5,16).
The influence of curcumin on DKK-3 was further investigated. In the
sample images of the immunohistochemical staining (Fig. 3A), yellow granules in cytoplasm
indicated the expression of DKK-3. As illustrated in the images
(Fig. 3A), DKK-3 was mostly
expressed in cytoplasm and its expression was higher in the Con
group compared with in the CHF group and CHF-cur group.
Quantitative analysis of relative DKK-3 expression levels based on
immunohistochemical staining indicated that DKK-3 was reduced in
the CHF group compared with the Con group but it was increased in
the CHF-cur group (Fig. 3A and B).
The western blot analysis indicated that expression levels of DKK-3
were lower in the CHF compared with the Con group, while these were
partially restored by curcumin treatment in CHF-cur group (Fig. 3C and D), which were in accordance
to the immunohistochemistry staining results.
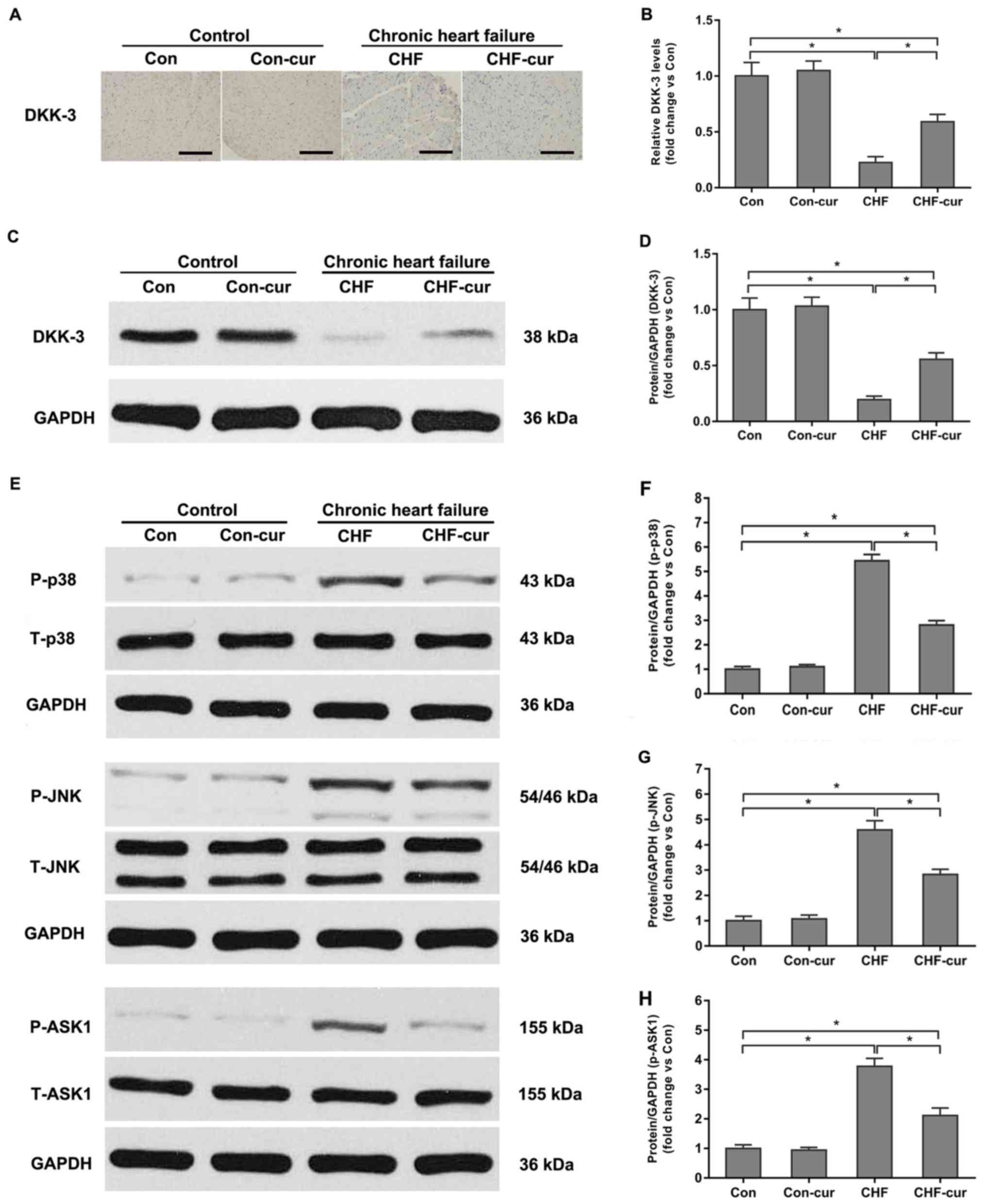 | Figure 3.Effects of curcumin on DKK-3, p38,
JNK and ASK1 protein expression levels. (A) Representative images
of DKK-3 and (B) quantification of its expression levels in LVAW,
as evaluated by immunohistochemical staining. (C) Representative
images of DKK-3 and (D) quantification of its expression, as
evaluated by western blot analysis. (E) Representative images of
p-p38, p-JNK and p-ASK1 and quantification of (F) p-p38; (G) p-JNK
and (H) p-ASK1 expression levels, as measured by western blot
analysis. Magnification, ×200. Scale bar=50 µm. All data are
expressed as the mean ± stadard deviation, n=10. *P<0.05. ASK1,
apoptosis signal-regulating kinase 1; CHF, chronic heart failure;
Con, control; Cur, curcumin; DKK-3, Dickkopf-related protein 3;
LVAW, left ventricular posterior wall thickness; p, phosphorylated;
T, total; p38, p38 mitogen-activated protein kinase; JNK, c-Jun
N-terminal kinase. |
Dickkopf-3 upregulation is involved in
the cardioprotective effects of curcumin on CHF rabbits
DKK-3 was demonstrated to be an inhibitor of p38 and
JNK signaling pathways (5,16). In order to reveal the signaling
mechanisms involved in the regulation of curcumin, the influence of
curcumin on DKK-3, p38 and JNK was investigated. Compared with the
Con group, p-p38 and P-JNK were significantly higher in the CHF
group. Curcumin decreased p-p38 and P-JNK expression levels in
CHF-cur group compared with the CHF group (Fig. 3E-G). This indicated that p38 and
JNK signaling pathways were significantly activated during CHF and
curcumin partially inhibited the p38 and JNK signaling pathways
activation in the CHF-cur group. In addition, ASK1 is regarded as
the upstream component of the p38 and JNK signaling pathways and
ASK1 phosphorylation is largely responsible for the increase of p38
and JNK phosphorylation (16). A
previous study reported that DKK-3 could protect against cardiac
remodeling induced by myocardial infarction or pressure overload
via negatively regulating the p38 and JNK signaling pathways in an
ASK1-dependent way (5,16). Therefore, the effects of curcumin
on ASK1 were further investigated. Corresponding to the p38 and JNK
activation, ASK1 was activated in CHF and curcumin partly inhibited
its activation in CHF-cur group (Fig.
3E and H).
Discussion
Rabbit and human hearts are anatomically and
functionally similar, and therefore CHF is commonly modeled in
rabbits (18,21). Curcumin is a potential candidate
for a novel anti-cancer drug and the cardioprotective effects of
curcumin on heart diseases have been reported, but the underlying
mechanisms have not been revealed fully (9). In the present study, a CHF model was
successfully induced by ten weeks volume and eight weeks pressure
overload. Ten weeks treatment with curcumin improved cardiac
performance, which was indicated by the heart/body weight ratio and
echocardiography parameters. Cardiac remodeling was also alleviated
by curcumin treatment in the CHF-cur rabbits, including myocardial
hypertrophy, fibrosis, apoptosis and ultrastructure
disorganization. Curcumin also decreased the expression of some CHF
molecular biomarkers. The present study demonstrated that curcumin
upregulated DKK-3 expression which may lead to inhibition of p38
and JNK signaling pathways and this may be a potential underlying
mechanism by which curcumin exerted cardiac protective effects, but
this needs to be confirmed in future studies.
In previous studies, curcumin was reported to exert
anti-inflammatory and anti-oxidant effects on several kinds of
diseases (9,22). Cardiac remodeling, including
cardiac hypertrophy, fibrosis and apoptosis, is partially
attributable to overactive inflammatory response and excessive ROS
release in CHF (6). When heart
suffers from prolonged pressure overload or myocardial infarction,
intracellular ROS can activate multiple signaling pathways,
including p38 and JNK signaling pathways which may upregulate the
expression of TNF-α in myocytes (23). TNF-α in turn increases ROS
production by uncoupling mitochondrial respiration in a positive
feedback mechanism (6). This
pathological process may accelerate the cardiac remodeling through
multifaceted impaction (24,25).
DKK-3 was also demonstrated to suppress inflammation following
myocardial infarction and promote cell survival by reducing the
expression of the superoxide during oxidative stress, but the
effects of curcumin on DKK-3 has not been clarified (16,26).
The present study demonstrated that curcumin increased DKK-3
expression and the increased DKK-3 suppressed inflammatory
reactions, and reduced ROS via inhibiting p38 and JNK signaling
pathways in rabbits with CHF.
Curcumin has been reported to serve therapeutic
roles on renal and hepatic fibrosis, but rarely on cardiac fibrosis
(27–29). An excessive deposition of
extracellular matrix (ECM) molecules is another determining factor
for cardiac remodeling, especially for myocardial fibrosis
(30,31). Previous studies demonstrated that
the deposition of ECM positively associates with MMP-2 and MMP-9
levels while tissue inhibitors of metalloproteinases alleviate this
deposition (31,32). Upregulation of MMP-2 and MMP-9 in
cardiac fibroblasts is associated with the activation of p38 and
JNK signaling pathways (33,34).
Inducing excessive deposition of ECM, p38 and JNK signaling
pathways activation accelerates cardiac fibrosis by promoting
migration of inflammatory cells and vascular regeneration through
upregulating MMPs (35). A
previous study reported that decreased DKK-3 expression leads to
up-regulation of MMPs in normal prostate epithelial cells and acts
as an inhibitor of p38 and JNK signaling pathways (36). The present study demonstrated that
curcumin increased DKK-3 expression and downregulated MMP-2 and
MMP-9 possibly by inhibiting p38 and JNK pathways in CHF
rabbits.
It has been previously demonstrated that CHF is
characterized by dysregulation of SERCA2a, the major
Ca2+ pump in the cardiomyocytes (37,38).
SERCA2a can regulate excitation-contraction coupling in
cardiomyocytes (38,39). SERCA2a can be regulated precisely
according to the physiological condition of the heart (40,41).
SERCA2a was identified to be lower in cardiomyocytes of CHF models
and an increase of SERCA2a expression could improve cardiac
function (42,43). SERCA2a expression was demonstrated
to be downregulated by activated p38 and JNK signaling pathways in
cultured neonatal and mature cardiomyocytes (44,45).
The association between DKK-3 and SERCA2a, as well as the effects
of curcumin on SERCA2a, have rarely been studied (46). In the present study, p38 and JNK
signaling pathways were inhibited while both DKK-3 and SERCA2a
expression levels were upregulated in the CHF-cur group compared
with the CHF group. This indicates that curcumin enhanced DKK-3
expression, which in turn may up-regulate SERCA2a via inhibition of
p38 and JNK signaling pathways in CHF rabbits.
It was reported that curcumin exerts
anti-inflammatory effects via inhibition of the p38 and JNK
pathways in HaCaT cells (47).
Activation of p38 and JNK pathways is also involved in the
pathology of CHF (33,48). In addition, ASK1 phosphorylation is
mainly responsible for the activation of p38 and JNK pathways, and
a previous study demonstrated that DKK-3 inhibited them in an
ASK1-dependent manner (16).
Cardiac hypertrophy and fibrosis were exacerbated by DKK-3 knockout
while ameliorated by DKK-3 overexpression (5). In the present study, p38, JNK and
ASK1 kinases were all activated in the CHF group, accompanied by
the downregulation of DKK-3 and were partially restored by curcumin
in the CHF-cur group compared with the CHF group. Yao et al
(17) reported that curcumin may
improve left ventricular function and remodeling in pressure
overloaded rabbits by suppressing myocardial expression of TNF-a
and MMP-2. Jeong et al (49) demonstrated that curcumin could
attenuate cardiac remodeling in myocardial ischemia/reperfusion
(I/R) injury by inhibiting p38 and JNK pathways in vivo. In
addition, curcumin pretreatment was also demonstrated to protect
cardiac cells against I/R injury by reducing oxidative stress and
inhibiting the phosphorylation of JNK in vitro, as
demonstrated by Fiorillo et al (50). The present results are in
accordance with previous studies and could serve as additional
evidence for the cardioprotective potentials of curcumin in cardiac
disease. Furthermore, a potential underlying mechanism was revealed
in which curcumin may inhibit p38 and JNK signaling pathways in an
ASK1-dependent way via up-regulation of DKK-3 in a rabbit model of
CHF. However, there are limitations in the present study for
identifying whether knock-out of the DKK-3 gene affects the
cardioprotective effect of curcumin and it would be worth further
investigation by future studies. The present study proved that
DKK-3 upregulation mediates the cardioprotective effects of
curcumin on chronic heart failure. Further studies should verify
whether knock-out of the DKK-3 gene affects the cardioprotective
effect of curcumin and overexpression experiments are needed in
order to validate the present findings. Further studies with
cardiospecific DKK-3 knockout and overexpression experiments are
needed in order to validate the present findings.
Acknowledgements
Not applicable.
Funding
The present study was supported by National Natural
Science Foundation of China (grant. nos. 81570460, 81500654,
81170767, 81571376 and 81500668); Natural Science Foundation of
Hubei Province (grant. no. 2016CFB392); Hubei Province health and
family planning scientific research project (grant. no.
WJ2017Z003).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Author's contributions
QC designed the study and drafted this manuscript;
MB directed the design and implementation of this research and is
the guarantor of the manuscript; JZ and LG participated in the
design of this study, and reviewed and edited this manuscript; YZ
and MD collected the relative data and were responsible for the
data integrity. All authors approved the final version to be
published.
Ethics approval and consent to
participate
All procedures were according to the ‘Guide for the
Care and Use of Laboratory Animals’ published by the US National
Institutes of Health (NIH Publication no. 85-23, revised 1996) and
approved by the Institutional Animal Care and Use Committee at
Renmin Hospital of Wuhan University (Wuhan, China).
Consent for publication
Not applicable.
Competing interests
Authors declare they have no competing
interests.
References
|
1
|
Cook C, Cole G, Asaria P, Jabbour R and
Francis DP: The annual global economic burden of heart failure. Int
J Cardiol. 171:368–376. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Benjamin EJ, Blaha MJ, Chiuve SE, Cushman
M, Das SR, Deo R, de Ferranti SD, Floyd J, Fornage M, Gillespie C,
et al: Heart disease and stroke statistics-2017 update: A report
from the american heart association. Circulation. 135:e146–e603.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Josiak K, Jankowska EA, Piepoli MF,
Banasiak W and Ponikowski P: Skeletal myopathy in patients with
chronic heart failure: Significance of anabolic-androgenic
hormones. J Cachexia Sarcopenia Muscle. 5:287–296. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Koitabashi N and Kass DA: Reverse
remodeling in heart failure-mechanisms and therapeutic
opportunities. Nat Rev Cardiol. 9:147–157. 2012. View Article : Google Scholar
|
|
5
|
Zhang Y, Liu Y, Zhu XH, Zhang XD, Jiang
DS, Bian ZY, Zhang XF, Chen K, Wei X, Gao L, et al: Dickkopf-3
attenuates pressure overload-induced cardiac remodelling.
Cardiovasc Res. 102:35–45. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hofmann U and Frantz S: How can we cure a
heart ‘in flame’? A translational view on inflammation in heart
failure. Basic Res Cardiol. 108:3562013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rahman AF, Angawi RF and Kadi AA: Spatial
localisation of curcumin and rapid screening of the chemical
compositions of turmeric rhizomes (Curcuma longa Linn.) using
direct analysis in real time-mass spectrometry (DART-MS). Food
Chem. 173:489–494. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jankun J, Wyganowska-Swiatkowska M,
Dettlaff K, Jelinska A, Surdacka A, Watrobska-Swietlikowska D and
Skrzypczak-Jankun E: Determining whether curcumin
degradation/condensation is actually bioactivation (Review). Int J
Mol Med. 37:1151–1158. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Parada E, Buendia I, Navarro E, Avendano
C, Egea J and Lopez MG: Microglial HO-1 induction by curcumin
provides antioxidant, antineuroinflammatory, and glioprotective
effects. Mol Nutr Food Res. 59:1690–1700. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhu X, Li Q, Chang R, Yang D, Song Z, Guo
Q and Huang C: Curcumin alleviates neuropathic pain by inhibiting
p300/CBP histone acetyltransferase activity-regulated expression of
BDNF and cox-2 in a rat model. PloS One. 9:e913032014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee JY, Lee YM, Chang GC, Yu SL, Hsieh WY,
Chen JJ, Chen HW and Yang PC: Curcumin induces EGFR degradation in
lung adenocarcinoma and modulates p38 activation in intestine: The
versatile adjuvant for gefitinib therapy. PloS One. 6:e237562011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Morimoto T, Sunagawa Y, Kawamura T, Takaya
T, Wada H, Nagasawa A, Komeda M, Fujita M, Shimatsu A, Kita T and
Hasegawa K: The dietary compound curcumin inhibits p300 histone
acetyltransferase activity and prevents heart failure in rats. J
Clin Invest. 118:868–878. 2008.PubMed/NCBI
|
|
13
|
Li HY, Yang M, Li Z and Meng Z: Curcumin
inhibits angiotensin II-induced inflammation and proliferation of
rat vascular smooth muscle cells by elevating PPAR-gamma activity
and reducing oxidative stress. Int J Mol Med. 39:1307–1316. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ochiai K, Watanabe M, Ueki H, Huang P,
Fujii Y, Nasu Y, Noguchi H, Hirata T, Sakaguchi M, Huh NH, et al:
Tumor suppressor REIC/Dkk-3 interacts with the dynein light chain,
Tctex-1. Biochem Biophys Res Commun. 412:391–395. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Veeck J and Dahl E: Targeting the Wnt
pathway in cancer: The emerging role of Dickkopf-3. Biochim Biophys
Acta. 1825:18–28. 2012.PubMed/NCBI
|
|
16
|
Bao MW, Cai Z, Zhang XJ, Li L, Liu X, Wan
N, Hu G, Wan F, Zhang R, Zhu X, et al: Dickkopf-3 protects against
cardiac dysfunction and ventricular remodelling following
myocardial infarction. Basic Res Cardiol. 110:252015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yao QH, Wang DQ, Cui CC, Yuan ZY, Chen SB,
Yao XW, Wang JK and Lian JF: Curcumin ameliorates left ventricular
function in rabbits with pressure overload: Inhibition of the
remodeling of the left ventricular collagen network associated with
suppression of myocardial tumor necrosis factor-alpha and matrix
metalloproteinase-2 expression. Biol Pharm Bull. 27:198–202. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nikolaidou T, Cai XJ, Stephenson RS, Yanni
J, Lowe T, Atkinson AJ, Jones CB, Sardar R, Corno AF, Dobrzynski H,
et al: Congestive heart failure leads to prolongation of the PR
interval and atrioventricular junction enlargement and ion channel
remodelling in the rabbit. PloS One. 10:e01414522015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mukherjee R, Mingoia JT, Bruce JA, Austin
JS, Stroud RE, Escobar GP, McClister DM Jr, Allen CM, Alfonso-Jaume
MA, Fini ME, et al: Selective spatiotemporal induction of matrix
metalloproteinase-2 and matrix metalloproteinase-9 transcription
after myocardial infarction. Am J Physiol Heart Circ Physiol.
291:H2216–H2228. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dhalla NS, Saini-Chohan HK,
Rodriguez-Leyva D, Elimban V, Dent MR and Tappia PS: Subcellular
remodelling may induce cardiac dysfunction in congestive heart
failure. Cardiovasc Res. 81:429–438. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang F, Dang Y, Li Y, Hao Q, Li R and Qi
X: Cardiac Contractility modulation attenuate myocardial fibrosis
by inhibiting TGF-β1/Smad3 signaling pathway in a rabbit model of
chronic heart failure. Cell Physiol Biochem. 39:294–302. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Soetikno V, Sari FR, Lakshmanan AP,
Arumugam S, Harima M, Suzuki K, Kawachi H and Watanabe K: Curcumin
alleviates oxidative stress, inflammation, and renal fibrosis in
remnant kidney through the Nrf2-keap1 pathway. Mol Nutr Food Res.
57:1649–1659. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hori M and Nishida K: Oxidative stress and
left ventricular remodelling after myocardial infarction.
Cardiovasc Res. 81:457–464. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Munzel T, Gori T, Keaney JF Jr, Maack C
and Daiber A: Pathophysiological role of oxidative stress in
systolic and diastolic heart failure and its therapeutic
implications. Eur Heart J. 36:2555–2564. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Deo SH, Fisher JP, Vianna LC, Kim A,
Chockalingam A, Zimmerman MC, Zucker IH and Fadel PJ: Statin
therapy lowers muscle sympathetic nerve activity and oxidative
stress in patients with heart failure. Am J Physiol Heart Circ
Physiol. 303:H377–H385. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qui S, Kano J and Noguchi M: Dickkopf 3
attenuates xanthine dehydrogenase expression to prevent oxidative
stress-induced apoptosis. Genes Cells. 22:406–417. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou X, Zhang J, Xu C and Wang W: Curcumin
ameliorates renal fibrosis by inhibiting local fibroblast
proliferation and extracellular matrix deposition. J Pharmacol Sci.
126:344–350. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang F, Zhang Z, Chen L, Kong D, Zhang X,
Lu C, Lu Y and Zheng S: Curcumin attenuates angiogenesis in liver
fibrosis and inhibits angiogenic properties of hepatic stellate
cells. J Cell Mol Med. 18:1392–1406. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang ME, Chen YC, Chen IS, Hsieh SC, Chen
SS and Chiu CH: Curcumin protects against thioacetamide-induced
hepatic fibrosis by attenuating the inflammatory response and
inducing apoptosis of damaged hepatocytes. J Nutr Biochem.
23:1352–1366. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bonnans C, Chou J and Werb Z: Remodelling
the extracellular matrix in development and disease. Nat Rev Mol
Cell Biol. 15:786–801. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vanhoutte D, van Almen GC, Van Aelst LN,
Van Cleemput J, Droogne W, Jin Y, Van de Werf F, Carmeliet P,
Vanhaecke J, Papageorgiou AP and Heymans S: Matricellular proteins
and matrix metalloproteinases mark the inflammatory and fibrotic
response in human cardiac allograft rejection. Eur Heart J.
34:1930–1941. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Takawale A, Sakamuri SS and Kassiri Z:
Extracellular matrix communication and turnover in cardiac
physiology and pathology. Compr Physiol. 5:687–719. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dodd T, Jadhav R, Wiggins L, Stewart J,
Smith E, Russell JC and Rocic P: MMPs 2 and 9 are essential for
coronary collateral growth and are prominently regulated by p38
MAPK. J Mol Cell Cardiol. 51:1015–1025. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liang KC, Lee CW, Lin WN, Lin CC, Wu CB,
Luo SF and Yang CM: Interleukin-1beta induces MMP-9 expression via
p42/p44 MAPK, p38 MAPK, JNK, and nuclear factor-kappaB signaling
pathways in human tracheal smooth muscle cells. J Cell Physiol.
211:759–770. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cuenda A and Rousseau S: p38 MAP-kinases
pathway regulation, function and role in human diseases. Biochim
Biophys Acta. 1773:1358–1375. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Romero D, Al-Shareef Z, Gorrono-Etxebarria
I, Atkins S, Turrell F, Chhetri J, Bengoa-Vergniory N, Zenzmaier C,
Berger P, Waxman J and Kypta R: Dickkopf-3 regulates prostate
epithelial cell acinar morphogenesis and prostate cancer cell
invasion by limiting TGF-β-dependent activation of matrix
metalloproteases. Carcinogenesis. 37:18–29. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Brini M and Carafoli E: Calcium pumps in
health and disease. Physiol Rev. 89:1341–1378. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Roe AT, Frisk M and Louch WE: Targeting
cardiomyocyte Ca2+ homeostasis in heart failure. Curr Pharm Des.
21:431–448. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Inesi G, Prasad AM and Pilankatta R: The
Ca2+ ATPase of cardiac sarcoplasmic reticulum: Physiological role
and relevance to diseases. Biochem Biophys Res Commun. 369:182–187.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cutler MJ, Wan X, Plummer BN, Liu H,
Deschenes I, Laurita KR, Hajjar RJ and Rosenbaum DS: Targeted
sarcoplasmic reticulum Ca2+ ATPase 2a gene delivery to restore
electrical stability in the failing heart. Circulation.
126:2095–2104. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Huang CL: SERCA2a stimulation by
istaroxime: A novel mechanism of action with translational
implications. Br J Pharmacol. 170:486–488. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Butter C, Rastogi S, Minden HH, Meyhofer
J, Burkhoff D and Sabbah HN: Cardiac contractility modulation
electrical signals improve myocardial gene expression in patients
with heart failure. J Am Coll Cardiol. 51:1784–1789. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gwathmey JK, Yerevanian A and Hajjar RJ:
Targeting sarcoplasmic reticulum calcium ATPase by gene therapy.
Hum Gene Ther. 24:937–947. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Scharf M, Neef S, Freund R, Geers-Knorr C,
Franz-Wachtel M, Brandis A, Krone D, Schneider H, Groos S, Menon
MB, et al: Mitogen-activated protein kinase-activated protein
kinases 2 and 3 regulate SERCA2a expression and fiber type
composition to modulate skeletal muscle and cardiomyocyte function.
Mol Cell Biol. 33:2586–2602. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ritchie MF, Zhou Y and Soboloff J:
Transcriptional mechanisms regulating Ca(2+) homeostasis. Cell
calcium. 49:314–321. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sumbilla C, Lewis D, Hammerschmidt T and
Inesi G: The slippage of the Ca2+ pump and its control by anions
and curcumin in skeletal and cardiac sarcoplasmic reticulum. J Biol
Chem. 277:13900–13906. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cho JW, Lee KS and Kim CW: Curcumin
attenuates the expression of IL-1beta, IL-6, and TNF-alpha as well
as cyclin E in TNF-alpha-treated HaCaT cells; NF-kappaB and MAPKs
as potential upstream targets. Int J Mol Med. 19:469–474.
2007.PubMed/NCBI
|
|
48
|
Kaikkonen L, Magga J, Ronkainen VP,
Koivisto E, Perjes A, Chuprun JK, Vinge LE, Kilpio T, Aro J, Ulvila
J, et al: p38α regulates SERCA2a function. J Mol Cell Cardiol.
67:86–93. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Jeong CW, Yoo KY, Lee SH, Jeong HJ, Lee CS
and Kim SJ: Curcumin protects against regional myocardial
ischemia/reperfusion injury through activation of RISK/GSK-3β and
inhibition of p38 MAPK and JNK. J Cardiovasc Pharmacol Ther.
17:387–394. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fiorillo C, Becatti M, Pensalfini A,
Cecchi C, Lanzilao L, Donzelli G, Nassi N, Giannini L, Borchi E and
Nassi P: Curcumin protects cardiac cells against
ischemia-reperfusion injury: Effects on oxidative stress,
NF-kappaB, and JNK pathways. Free Radic Biol Med. 45:839–846. 2008.
View Article : Google Scholar : PubMed/NCBI
|














