Introduction
Hepatocellular carcinoma (HCC) is the most
frequently occurring primary cancer of the liver and is the 5th,
and the 3rd leading cause of cancer-associated death worldwide
among women and men, respectively (1). These poor outcomes result from its
high recurrence following liver resection and its resistance to
systemic chemotherapy (2,3). As cases of HCC are frequently
identified at advanced stages of the disease, these patients are
only eligible for palliative care. Furthermore, as no effective
palliative chemotherapies are available, the prognosis of patients
with advanced HCC is poor (4).
Therefore, developing pharmacologically effective agents from
natural products with minimal toxicity or minimal adverse effects
has become an area of interest.
Epithelial-mesenchymal transition (EMT), the process
by which epithelial cancer cells lose their polarity and become
motile mesenchymal cells, has been implicated in carcinoma invasion
and metastasis (5). Downregulation
of epithelial (E)-cadherin and the acquisition of mesenchymal
markers including vimentin, are regarded as the key steps of EMT.
The zinc-finger proteins Snai1 (Snail) and Snai2 (Slug) are the
most prominent suppressors of E-cadherin transcription (6,7).
Matrix metalloproteinases (MMPs) are a family of
metalloendopeptidase that cleave the protein components of the
extracellular matrix (ECM) and the endothelial cell basement
membrane (8). MMPs serve a central
role in tumor EMT by the cleavage of components of the ECM and the
endothelial cell basement membrane. MMP2 and MMP9, members of the
MMP family, have been demonstrated in cancer tissues, and are
associated with the processes of tumor aggression, and metastasis
in human cancer (9). Furthermore,
a previous study reported that MMP2 and MMP9 could break down cell
surface associated molecules (10), and could cleave the E-cadherin
ectodomain near the plasma membrane into soluble E-cadherin
(11), which may be involved in
MMPs-enhanced invasion and metastatic potential of cancer
cells.
Matrine is an essential compound identified in the
traditional Chinese herb Sophora flavescens Ait. (12), which has been used to treat viral
hepatitis, liver cirrhosis, cardiac arrhythmia and skin
inflammations in China for centuries; furthermore, this herb does
not exhibit any obvious side-effects (13). Previously findings revealed that
matrine exerts anti-tumor effects on several tumor cell types,
including HCC, as well as pancreatic, gastric and breast cancer. In
addition, matrine was demonstrated to enhance the immune function
of patients and therefore improve the quality of life of patients
(14). Although matrine has been
used clinically to treat various types of cancer in recent years,
the therapeutic efficacy of matrine for EMT of HCC remains poorly
understood.
Consistent with the published literature, the
present study aimed to ascertain whether there were any alterations
in EMT in hepatoma cells as a result of being treated with matrine,
to investigate the mechanisms underlying the diminished EMT
activity of matrine through the phosphatidylinositol
3,4,5-trisphosphate 3-phosphatase and dual-specificity protein
phosphatase PTEN (PTEN)/protein kinase B (Akt) signaling pathway
and altered the expression of proteins involved in EMT, including
MMP2, MMP9, E-cadherin, vimentin, Slug and Snail. The aim of the
present study was to develop novel strategies for the treatment of
HCC.
Materials and methods
Chemicals and reagents
Matrine (purity >98%), MTT, Triton X-100 and
dimethyl sulfoxide (DMSO) were all purchased from Sigma-Aldrich
(Merck KGaA, Darmstadt, Germany). Anti-phosphorylated (p)-Akt (cat.
no. 66444-1-1g), anti-Akt (cat. no. 55230-1-AP), anti-PTEN (cat.
no. 22034-1-AP), anti-E-cadherin (cat. no. 20874-1-AP) and
anti-vimentin (cat. no. 10366-1-AP) antibodies were all purchased
from Wuhan Sanying Biotechnology (Wuhan, China). Anti-MMP2 (cat.
no. 40994), anti-MMP9 (cat. no. 3852), anti-Snail (cat. no. 3879),
anti-Slug (cat. no. 9585) antibodies and DyLight™ 594 Phalloidin
(cat. no. 12877) were all purchased from Cell Signaling Technology,
Inc. (Danvers, MA, USA). The anti-β-actin (cat. no. TA-09) antibody
was purchased from OriGene Technologies, Inc. (Beijing, China).
Radio immunoprecipitation assay (RIPA) lysis buffer and DAPI
staining solution were purchased from the Beyotime Institute of
Biotechnology (Shanghai, China).
Cell culture
Fetal bovine serum (FBS), penicillin/streptomycin
and trypsin were all purchased from Gibco (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Dulbecco's modified Eagle's
medium (DMEM) was purchased from Invitrogen (Thermo Fisher
Scientific, Inc.). The human hepatoma cell line Huh-7 was purchased
from the Shanghai Institute of Biochemistry and Cell Biology,
Chinese Academy of Science (Shanghai, China). Huh-7 cells were
cultured in DMEM supplemented with 100 U/ml penicillin, 100 µg/ml
streptomycin and 10% FBS and incubated at 37°C in a humidified
atmosphere containing 5% CO2.
Cell viability assays
The MTT assay was used to assess the cytotoxic
activity of matrine. Huh-7 cells were plated in 96-well culture
plates (1×104 cells/well) and allowed to adhere
overnight at room temperature, following which the cells were
treated with various concentrations of matrine (0, 0.2, 0.4, 0.8,
1.0, 1.2, 1.4, 1.6, 1.8, or 2.0 mg/ml). The cells were subsequently
incubated for 24 and 48 h then they were washed twice with PBS and
treated with 5 mg/ml MTT for 4 h. The cells were washed with PBS
and then solubilized with 150 µl DMSO. The optical density values
were measured at an absorbance of 490 nm using a Wellscan reader
(Thermo Labsystems, Santa Rosa, CA, USA). The percentage inhibition
of cellular proliferation was calculated using equation ‘the
inhibitory percentage (%) = [1 - A490(test)/A490(blank)]x100%’. The
assays were performed in triplicate in three independent
experiments.
Colony forming assay
Huh-7 cells in the exponential growth phase were
seeded into 6-well plates at a density of 8×102
cells/well. Following incubation of the cells for 12 h, they were
treated with 0, 0.8, or 1.6 mg/ml matrine for 24 h and then
continuously cultured in fresh medium at 37°C in a humidified
atmosphere containing 5% CO2. Following 10–15 days of
culture, the cells were first washed twice with PBS, then fixed
with methanol for 30 min at room temperature and finally stained
with 0.5% crystal violet for 30 min at room temperature. A colony
was defined as a unit of at least 50 cells and all visible colonies
were counted by light microscopy. The colony formation rate was
reported as a percentage and calculated as the number of
colonies/the number of seeded cells ×100. The assays were performed
in triplicate in three independent experiments.
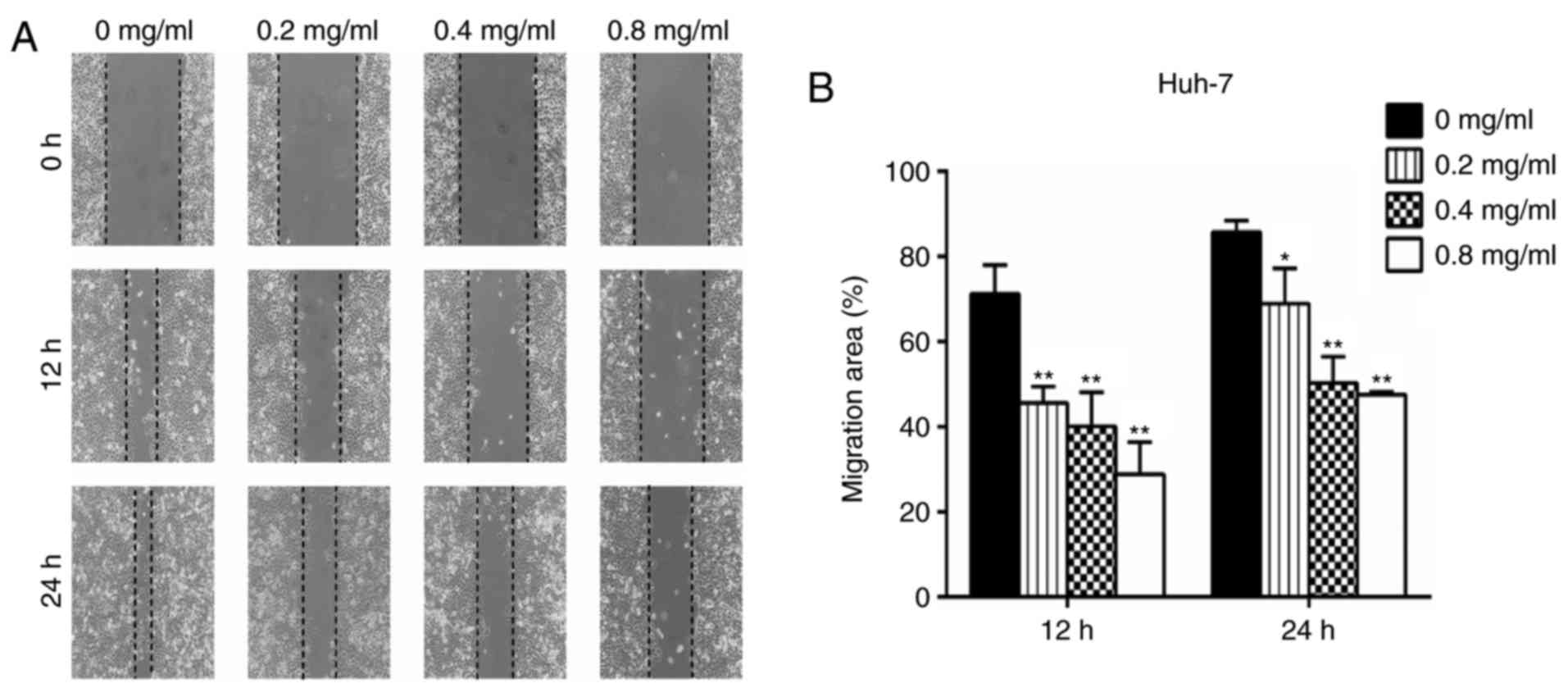
Wound healing assays
For the wound healing assays, Huh-7 cells were
seeded in 6-well plates at a density of 2×105 cells/well
and cultured for 24 h. Wounds were scratched with a 10 ml pipette
tip and then PBS was used to wash the wells and remove floating
cells prior to the addition of serum-free medium. The cells were
then treated with 0, 0.2, 0.4 or 0.8 mg/ml matrine for 0, 12 and 24
h. The migratory distance of the cells was measured and analyzed
using ImageJ 1.48 version (National Institutes of Health, Bethesda,
MD, USA). The tests were performed in triplicate in three
independent experiments.
In vitro invasion and migration
assays
Cell migration and invasion assays were performed
using Transwell chambers with 8-µm pores (Corning, Incorporated,
Corning, NY, USA). Huh-7 cells were pretreated with 0, 0.2, 0.4 or
0.8 mg/ml matrine for 24 h. For the migration assay,
1×104 cells/well in serum-free medium were seeded in the
upper chamber on a non-coated membrane. The lower chamber was
filled with medium containing 20% FBS, whereas the upper chamber
with a membrane pre-coated with Matrigel (BD Biosciences, Franklin
Lakes, NJ, USA) were used for the invasion assay. The cells were
subsequently incubated for 24 h at 37°C, those on the upper side of
the inserts were removed by swabbing with a cotton swab and the
cells on the bottom side of the filter were fixed with 100%
methanol for 30 min at room temperature and stained with 0.5%
crystal violet for 30 min at room temperature. The cells on the
lower surface were then counted and images were captured under an
inverted light microscope (Olympus Corporation, Tokyo, Japan). The
assays were performed in triplicate in three independent
experiments.
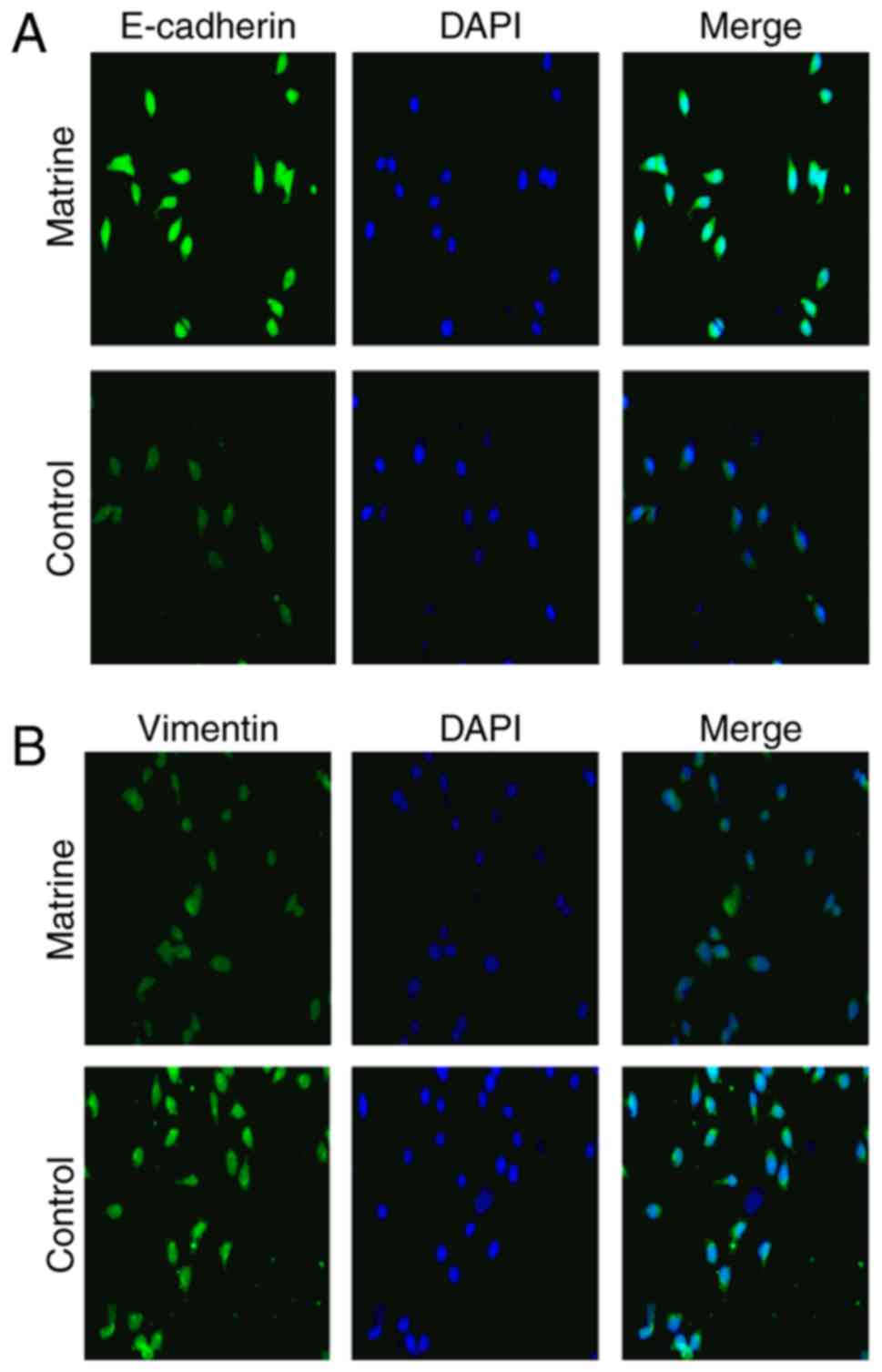
Immunofluorescence
Huh-7 cells were plated onto chamber slides at
2×104 cells per chamber for 12 h and stimulated with 0
and 0.8 mg/ml matrine for 24 h. Then, the cells were fixed with 4%
paraformaldehyde for 30 min at room temperature and subsequently
permeabilized with 0.25% Triton X-100 for 20 min at room
temperature. The slides were then treated with 0.5% bovine serum
albumin (BSA; Beyotime Institute of Biotechnology, Jiangsu, China)
in 0.1% Tween-20 for 30 min at room temperature, primary antibodies
targeting E-cadherin and vimentin were added at a 1:1,000 dilution
and incubated on the slides overnight at 4°C. Secondary antibodies
conjugated with the green fluorescent Alexa Fluor 594 dye (FITC
Conjugated Goat Anti-Rabbit IgG; cat. no. TA130021), were purchased
from OriGene Technologies, Inc. (Rockville, MD, USA), were added to
the slides at a dilution of 1:1,000 for 1 h at room temperature.
Then, the cells were washed with PBS and the coverslips were
mounted using an anti-fade mounting solution containing DAPI prior
to imaging the cells for 10 min at room temperature. The projected
cell area was evaluated using ImageJ software. The assays were
performed in triplicate in three independent experiments.
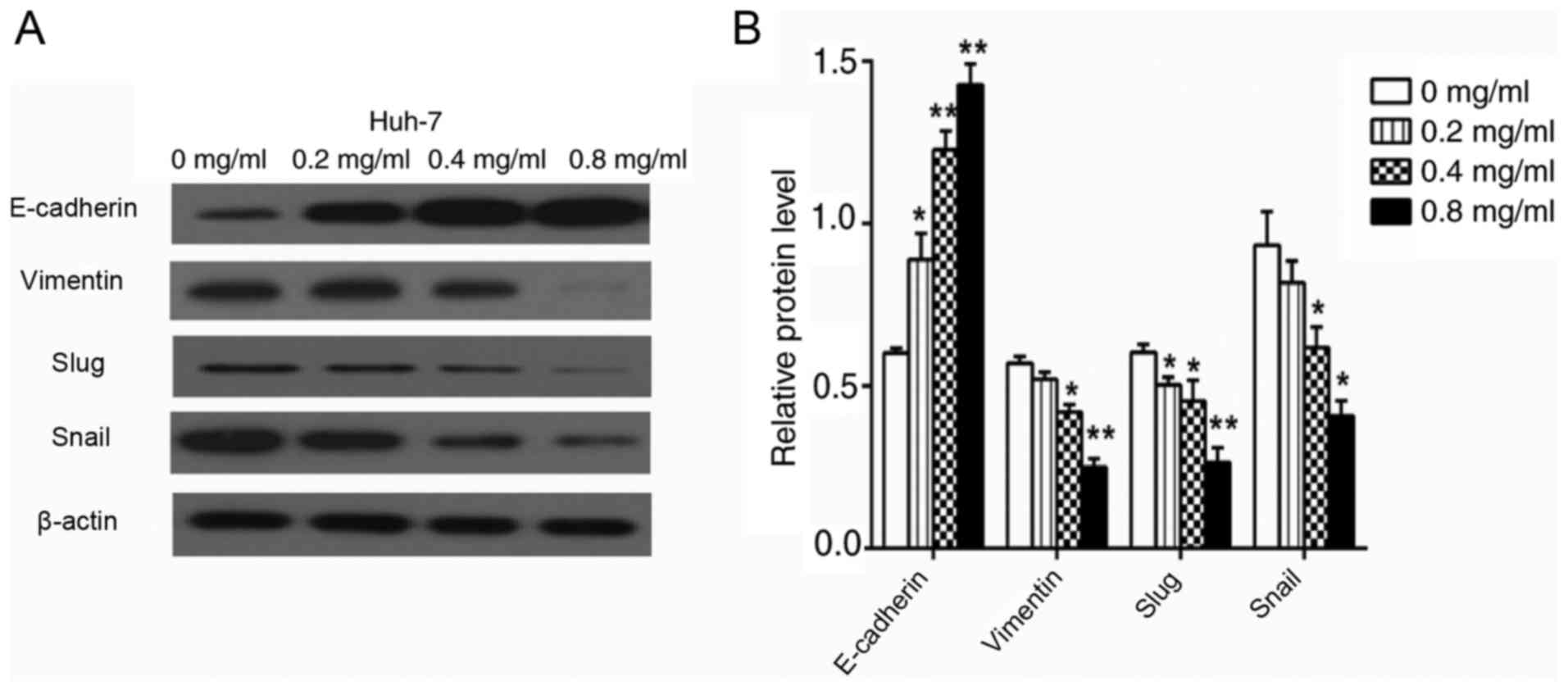
Western blot analysis
Huh-7 cells were planted in 6-well plates at
2×105 cells per well, treated with different
concentrations of matrine (0, 0.2, 0.4, or 0.8 mg/ml) for 36 h and
then lysed using a RIPA buffer. The lysates were centrifuged at 4°C
at 12,000 × g for 15 min and the protein concentration of the
remaining supernatants was measured using a bicinchoninic protein
assay kit (Beyotime Institute of Biotechnology). Samples of total
protein (20 µg) were separated using 12% SDS-PAGE. The separated
proteins were then transferred to polyvinylidene difluoride
membranes (EMD Millipore, Billerica, MA, USA) at 0.8
mA/cm2 for 2 h. Following blocking of these membranes
with a solution of 5% non-fat dry milk in buffer at 37°C for 1 h,
they were treated with antibodies targeting E-cadherin, vimentin,
Slug, Snail, PTEN, Akt, p-Akt and β-actin (all dilutions 1:1,000)
overnight at 4°C. Then, the membranes were washed and incubated
with horseradish peroxidase-conjugated (1:1,000) anti-mouse (cat.
no. ZB-5305) or (1:1,000) anti-rabbit antibodies (cat. no. ZB-5301;
Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd., Beijing,
China) at room temperature for 1 h. The protein bands on the
membranes were visualized using ECL detection reagents (P0018,
BeyoECL Plus; Beyotime Biotechnology, Jiangsu, China). In addition,
the antibodies targeting MMP2 and MMP9 (all dilutions 1:1,000) when
the cells were treated with matrine (0, 0.4 and 0.8 mg/ml) were
tested as described above. ImageJ software was used to perform the
densitometric analyses. The experiments were performed in
triplicate in three independent experiments.
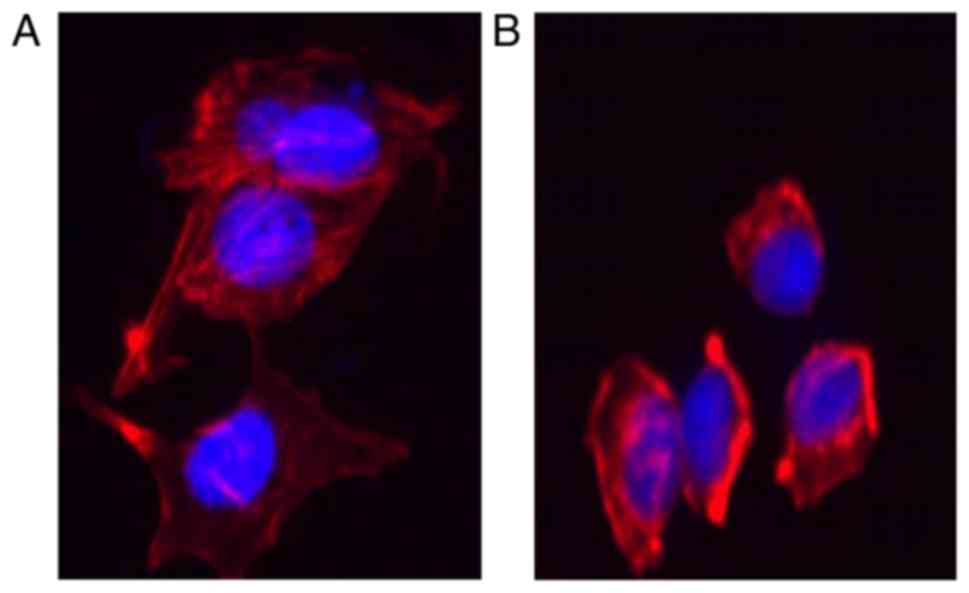
Actin staining of the
cytoskeleton
Huh-7 cells, which were treated with matrine (0, 0.8
mg/ml) were grown on coverslips and fixed with 4% fresh
paraformaldehyde for 10 min at room temperature, permeabilized with
0.1% Triton X-100 in PBS for 20 min at room temperature, and then
blocked with 5% BSA at room temperature for 1 h. Subsequently, the
cells were stained with Phalloidin for 2 h at room temperature in
the dark. Following being washed, the cells were counterstained
with DAPI for 10 min at room temperature. A fluorescence microscope
(Eclipse 90i; Nikon Corporation, Tokyo, Japan) was employed to
observe the distribution of filamentous (F)-actin. The assays were
performed in triplicate in three independent experiments.
Statistical analysis
Statistical analyses were performed using either the
Statistical Program for Social Sciences (SPSS) version 17.0 (SPSS,
Inc., Chicago, IL, USA) or the GraphPad Prism software package (v.
4.02; Graphpad, Inc., San Diego, CA, USA). Each experiment was
repeated at least three times. The values are expressed as the mean
± standard deviation. Either the Student's t-test or one-way
analysis of variance followed by the Newman-Keuls method were
performed to analyze the difference between two groups and multiple
groups, respectively. P<0.05 was considered to indicate a
statistically significant difference.
Results
Matrine inhibits the cell viability of
the human hepatoma cell line Huh-7
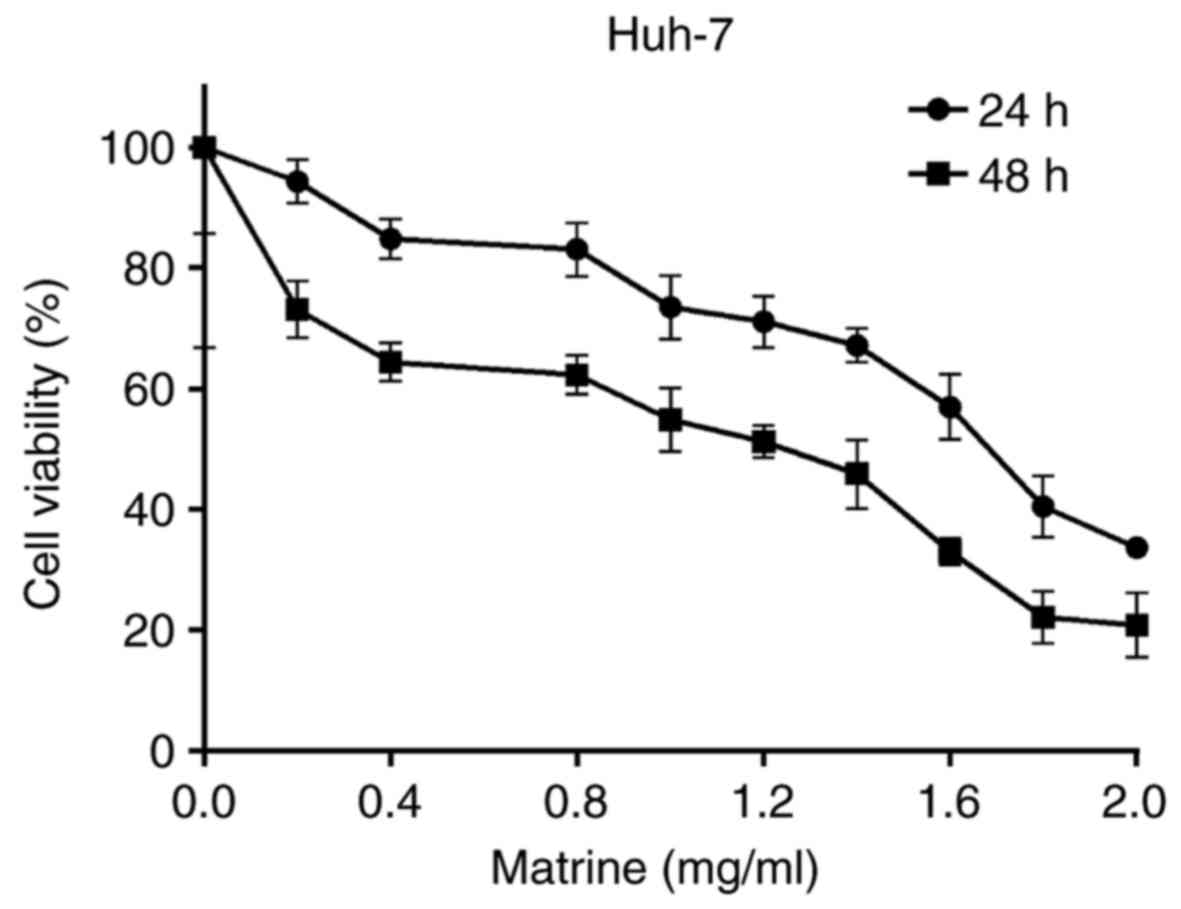
The effect of various concentrations of matrine (0
to 2 mg/ml) on the cell viability of Huh-7 cells following exposure
for 24 or 48 h is demonstrated in Fig.
1. At 1 mg/ml, matrine inhibited the cell viability of Huh-7
cells, whereas at concentrations lower than 1 mg/ml, the inhibitory
effect was not marked following 24 h. Therefore, concentrations
lower than 1 mg/ml were selected for the experiments associated
with invasion and metastasis.
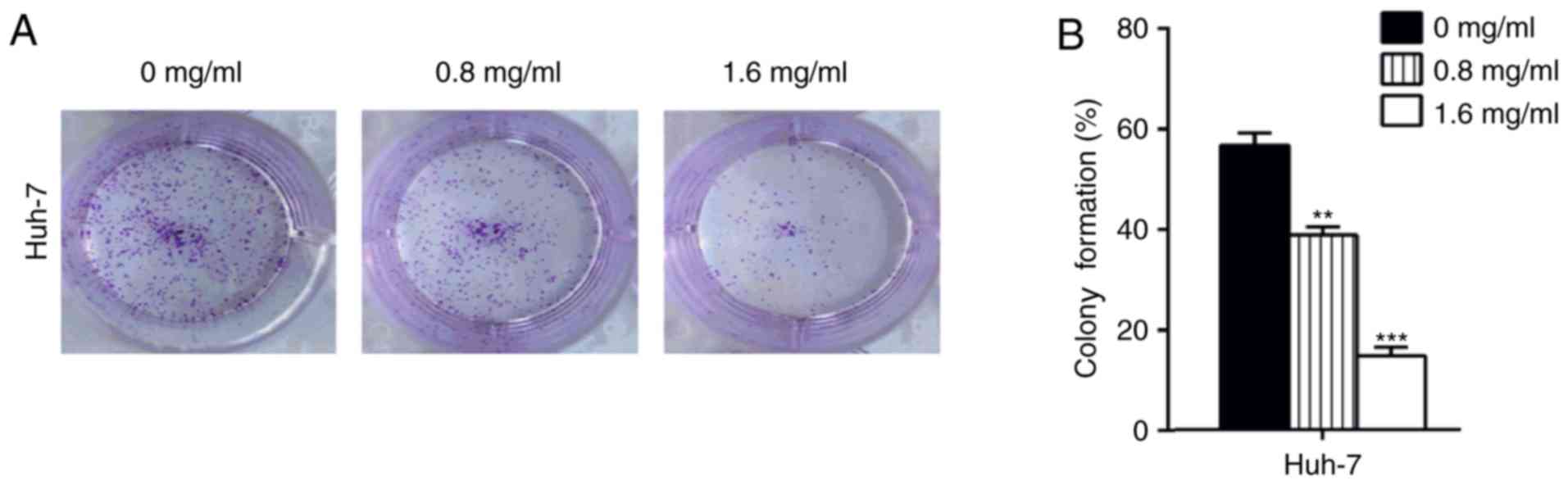
To investigate the long-term effects of matrine on
Huh-7 cells proliferation, a colony formation assay was performed.
Cells subjected to long-term matrine exposure exhibited
significantly reduced colony-forming abilities (P<0.01; Fig. 2). Furthermore, this suppression was
more significant in cells exposed to higher concentrations of
matrine (P<0.001). These results indicated that a matrine
concentration >1 mg/ml exerts a suppressive effect on the
proliferative abilities of Huh-7 cells.
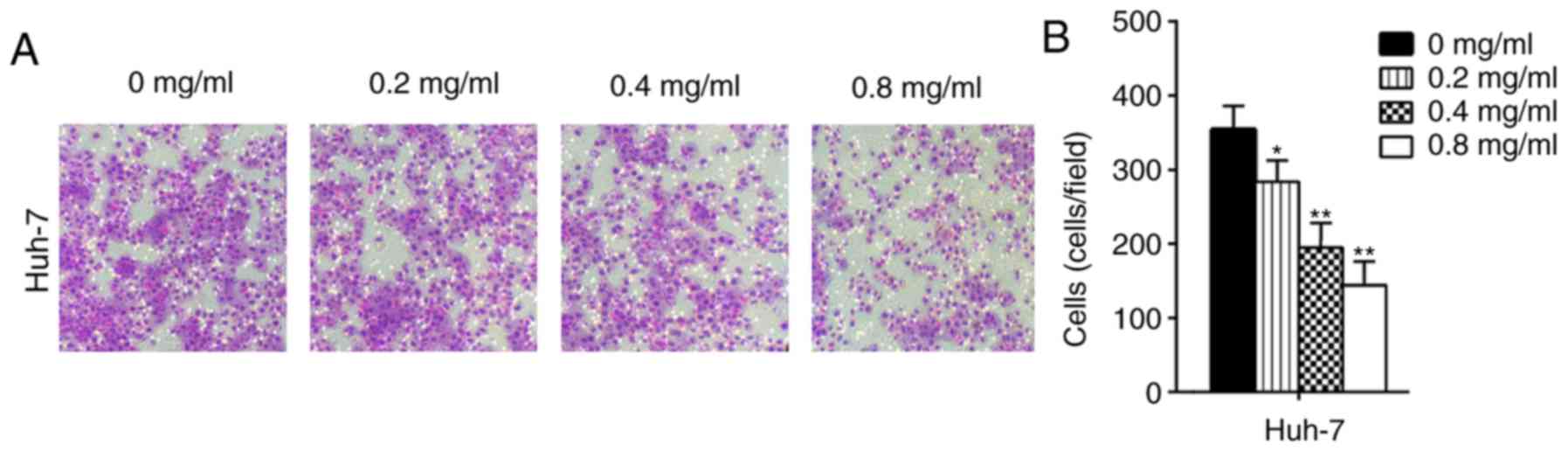
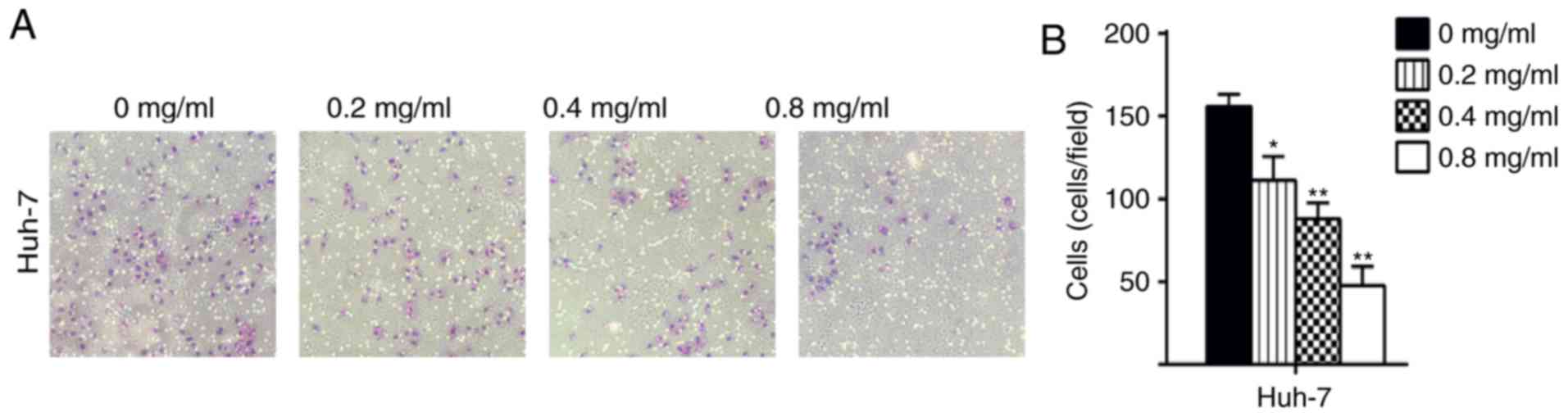
Matrine inhibits the migration and
invasion of Huh-7 cells
In the present study, the migratory ability of cells
treated with matrine was significantly decreased compared with the
control Huh-7 cells (P<0.05; Figs.
3 and 4). The results
demonstrated that matrine inhibited the invasion and migration of
Huh-7 cells in a concentration-dependent manner. In the wound
healing assays, the migration area (%) in the matrine group was
significantly increased compared with the control group (P<0.05;
Fig. 5). These experiments
illustrated that matrine may serve an essential role in regulating
the migratory and invasive abilities of a HCC cell line.
Matrine inhibits EMT by suppressing
vimentin and enhancing E-cadherin expression in human Huh-7 HCC
cells
Vimentin and E-cadherin have been reported to serve
important roles in EMT (15). To
determine how matrine induces EMT in Huh-7 cells, Huh-7 cells were
stimulated with 0.8 mg/ml matrine for 24 h (Fig. 6) and immunofluorescence was used to
measure the levels of vimentin and E-cadherin expressed in these
cells. It was observed that matrine induced the downregulation of
vimentin expression and the upregulation of E-cadherin expression;
these results indicate that matrine serves a critical role in
reducing EMT by decreasing vimentin expression and increasing
E-cadherin expression.
Matrine represses the cancer cells
exhibiting an EMT-like phenotypic alteration
Previous studies highlighted EMT as the mechanism by
which differentiated epithelial cells undergo notable morphological
alterations and acquire more motile and invasive capabilities
(16–19). Cytoskeletal reorganization is also
a characteristic of EMT and this feature was observed by F-actin
staining. As demonstrated in Fig
7A, the stress fibers in Huh-7 cells appeared as lamellipodia
and extensive parallel bundles, which were densely stained and
exhibited well-organized structures. In contrast, the lamellipodia
disappeared and the parallel bundles were disrupted in the cells
treated with matrine, which also demonstrated loosely organized
F-actin (Fig. 7B).
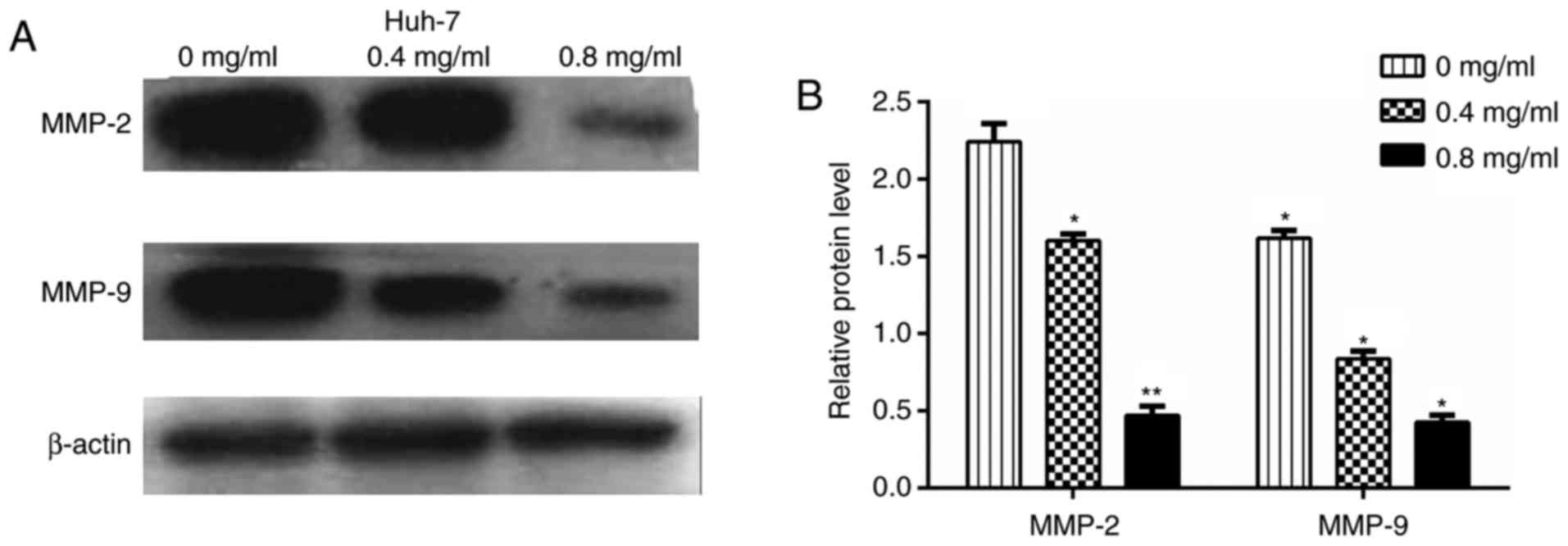
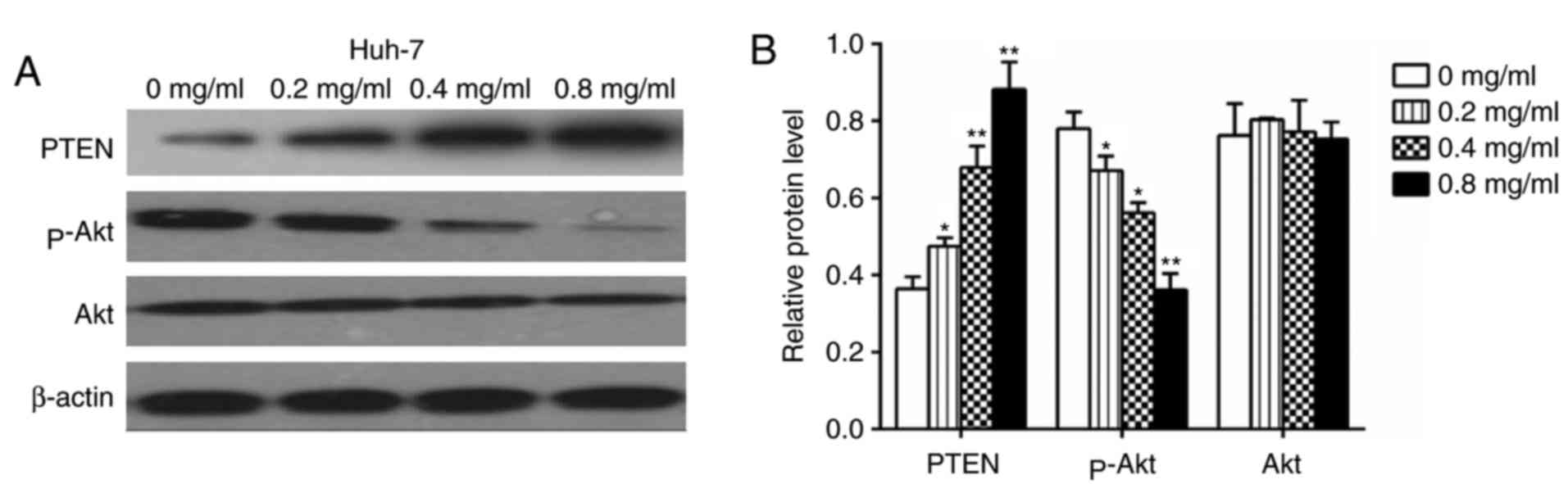
Matrine suppresses the expression of
Akt/p-Akt, vimentin, MMP2, MMP9, Slug and Snail and increases the
expression of PTEN and E-cadherin
Huh-7 cells treated with matrine for 24 h were
subjected to a western blotting assay to determine the expression
levels of vimentin, MMP2, MMP9, Slug, Snail and E-cadherin.
Figs. 8 and 9 revealed that compared with control
Huh-7 cells, matrine-treated cells demonstrated significantly
increased E-cadherin protein levels (P<0.01) and reduced
expression of vimentin, MMP2, MMP9, Slug and Snail in a
concentration-dependent manner. The effect of matrine on PTEN and
p-Akt expression in Huh-7 cells, was also investigated and the
western blot analysis demonstrated that matrine significantly
increased PTEN expression and significantly reduced the levels of
p-Akt in a concentration-dependent manner (P<0.05; Fig. 10).
Discussion
HCC is the most common liver malignancy and a major
health problem worldwide. Surgical resection and liver
transplantation, which are the current gold standards for the
treatment of HCC, are less than satisfactory due to metastasis and
high recurrence rates (20).
Matrine has been confirmed as a natural anti-tumor agent against
several types of cancer, including acute myeloid leukemia, prostate
cancer, lung cancer and human hepatoma (21–24).
Nevertheless, the anti-metastatic effect of matrine and its
associated mechanism(s) in HCC remained unclear. In the present
study, matrine was determined to inhibit the mobility and invasive
capability of hepatoma cells in vitro by modulating EMT via
the activation of the PTEN/Akt pathway. To the best of the authors'
knowledge, this is the first scientific study to report the
anti-metastatic effect of matrine on HCC.
In the present study, HCC cell line Huh-7 cells were
treated with matrine at concentrations lower than 1 mg/ml, which
exerted minimal effects on cell proliferation as confirmed by an
MTT assay. Notably, the cell morphology altered and a decrease in
lamellipodia, focal adhesion, and the stress fibers in cells
following treatment with matrine was observed. Based on this
observation, EMT-associated markers were analyzed. As anticipated,
the expression levels of mesenchymal markers were reduced and those
of epithelial markers were upregulated in the matrine-treated
cells. Transwell and wound healing assays were also used to analyze
the invasive and migratory abilities of hepatoma cells; these
properties were demonstrated to be suppressed. Therefore, these
data indicated that matrine may inhibit HCC invasion and metastasis
by inducing EMT, which is consistent with other studies that
focused on different cancer types (21,22).
Tumor metastasis and recurrence are two of the most
difficult challenges in curing hepatoma patients. To fully progress
to metastasis, carcinoma cells must complete multiple distinct
steps. An important biological process that has been demonstrated
to trigger cancer progression, metastasis and recurrence is EMT
(25). Indeed, tumor metastasis
has been associated with alterations in EMT markers (26), including decreased E-cadherin
expression (27), increased MMPs
expression (28) increased
vimentin expression (29),
increased collagen I expression (30), increased fibronectin expression
(31) and increased expression of
transcription factors from the Snail family (32). The phenomenon of EMT results in
epithelial cells losing their cell-cell adhesions, acquiring a
mesenchymal phenotype and also losing organized F-actin. This
causes cells to become more migratory and invasive, and in the case
of tumor cells, the ultimate consequence is metastatic spread
(27). In the present study, it
was observed that matrine enhanced E-cadherin expression and
reduced the expression of vimentin, Slug, Snail, MMP2 and MMP9. The
results suggested that the anti-metastatic effect of matrine on HCC
is associated with EMT.
EMT refers to a series of phenotypic and molecular
alterations that occur in non-cancerous cells over the course of
various steps of normal development as well as in cancer cells
(33). When EMT develops in
cancer, the patient's prognosis may be adversely affected (34). Therefore, an important property of
antineoplastic medicine is thought to be the successful repression
of EMT (35,36). Several phytochemicals, including
matrine, may offer a novel therapeutic approach for treating tumors
by repressing EMT (37–40). A previous study demonstrated that
the phosphoinositide 3-kinase (PI3K)/Akt pathway serves a
significant role in cell growth, metabolism, proliferation,
migration and apoptosis (41).
Furthermore, p-Akt is a well-known anti-apoptotic protein and the
primary downstream kinase of PI3K (42). Accordingly, the p-Akt levels in HCC
cells treated with matrine were investigated and the results
demonstrated that the p-Akt expression levels were reduced in a
concentration-dependent manner. To further clarify the mechanism
involved, the upstream regulatory factors of Akt, including PTEN
were measured. PTEN is tumor suppressor protein with protein
phosphatase and alkaline phosphatase activity and that can inhibit
the PI3K/Akt pathway (43). A
previous study confirmed that the p-Akt, p27 and pS6 expression
levels were increased in HCC tissues compared with the adjacent
non-tumor and normal liver tissues and the normal tissue
demonstrated elevated PTEN expression compared with the tumor
tissue (44). Furthermore, this
previous study demonstrated that over-expression of p-Akt, p27 and
pS6 was involved in poor differentiation, vascular invasion and
high tumor node metastasis stage of HCC, all of which were also
inversely proportional to PTEN expression. A previous study
demonstrated that silencing PTEN inhibited the cell growth
inhibition and apoptosis induced by matrine in M21 cells (45). These results suggested that PTEN
was required for the antitumor efficacy of matrine. In addition,
loss of PTEN function is the most commonly known genetic alteration
in the PI3-kinase cascade and is commonly associated with B-Raf
proto-oncogene mutations (45).
Furthermore, in line with recent findings, their results revealed
that HCC cells with downregulated p-Akt, p27 and pS6 expression
exhibited decreased invasion and metastasis (46). In addition, the induction of EMT by
transforming growth factor (TGF)-β is implicated in
hepatocarcinogenesis and HCC metastasis. These results demonstrated
that upregulation of HAb18G/cluster of differentiation (CD)147 is
stimulated by TGF-β and is coupled with the downregulation of
E-cadherin and the upregulation of neural-cadherin and vimentin.
HAb18G/CD147 expression is controlled by the
PI3K/Akt/phosphoinositide 3-kinase (GSK3b) cell survival signaling
pathway and is directly regulated by the transcription factor Slug.
One study revealed a novel effect of HAb18G/CD147 in mediating EMT
during the course of HCC progression and demonstrated that CD147 is
a Slug target in the TGF-β-PI3K/Akt-GSK3b-Snail-Slug-CD147
signaling cascade (47). Further
experiments are required to confirm the association of the
expression of the PTEN/p-Akt and the inhibitory effects of matrine.
Therefore, the present study investigated these downstream EMT
regulators of the PI3K/Akt pathway and reported that Snail and Slug
expression was downregulated in matrine-treated HCC cells. The
results demonstrated that the PTEN/PI3K/Akt pathway may serve as an
alternative mechanism underlying the effects of matrine.
In conclusion, the present study confirmed the
inhibitory function of matrine on the invasive and metastatic
abilities of HCC. Furthermore, decreases in vimentin expression and
increases in E-cadherin expression induced by matrine are
attributed to enhanced PTEN activity and inhibited Akt signaling.
This mechanism may contribute to the inhibitory effects of matrine
on invasion and metastasis in HCC. These results also revealed a
novel potential therapeutic application of matrine as an
anti-metastatic therapy for HCC.
Acknowledgements
The authors would like to thank Dr Pan for his
helpful advice regarding manuscript preparation. The present study
was performed in the First Affiliated Hospital of Harbin Medical
University Laboratory and the Shenzhen Longhua District Central
Hospital Laboratory.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
include in this published article.
Authors' contributions
YWW, BLC and SJZ conceived and designed the research
study. YWW, SJZ, JL and BBF performed the experiments and data
analysis described. YWW and JY prepared the figures and drafted the
manuscript. YWW was a major contributor in writing the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Berreta M, Rinaldi L, Di Benedetto F,
Lleshi A, De Re V, Facchini G, De Paoli P and Di Francia R:
Angiogenesis inhibitors for the treatment of hepatocellular
carcinoma. Front Pharmacol. 7:4282016.PubMed/NCBI
|
|
2
|
Germano D and Daniele B: Systemic therapy
of hepatocellular carcinoma: Current status and future
perspectives. World J Gastroenterol. 20:3087–3099. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ingle PV, Samsudin SZ, Chan PQ, Ng MK,
Heng LX, Yap SC, Chai AS and Wong AS: Development and novel
therapeutics in hepatocellular carcinoma: A review. Ther Clin Risk
Manag. 12:445–455. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Knox JJ, Qin R, Strosberg JR, Tan B,
Kaubisch A, El-Khoueiry AB, Bekaii-Saab TS, Rousey SR, Chen HX and
Erlichman C: A phase II trial of bevacizumab plus temsirolimus in
patients with advanced hepatocellular carcinoma. Invest New Drugs.
33:241–246. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chaffer CL, Juan San BP, Lim E and
Weinberg RA: EMT, cell plasticity and metastasis. Cancer Metastasis
Rev. 35:645–654. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou SL, Zhou ZJ, Hu ZQ, Li X, Huang XW,
Wang Z, Fan J, Dai Z and Zhou J: CXCR2/CXCL5 axis contributes to
epithelial-mesenchymal transition of HCC cells through activating
PI3K/Akt/GSK-3β\snail signaling. Cancer Lett. 358:124–35. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Darroli AA, Hink MA, DuBuc TQ, Teunisse
BJ, Goedhart J, Röttinger E and Postma M: Domain analysis of the
nematostella vectensis snail ortholog reveals unique nucleolar
localization that depends on the zinc-finger domains. Sci Rep.
5:121472015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Luo Z, Wang Q, Lau WB, Lau B, Xu L, Zhao
L, Yang H, Feng M, Xuan Y, Yang Y, et al: Tumor microenvironment:
The culprit for ovarian cancer metastasis? Cancer Lett.
377:174–182. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen CM, Hsieh SC, Lin CL, Lin YS, Tsal JP
and Hsieh YH: Alpha-mangostin suppresses the metastasis of human
renal carcinoma cells by targeting MEK/ERK expression and MMP9
transcription activity. Cell Physiol Biochem. 44:1460–1470. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kean MJ, Williams KC, Skaiski M, Myers D,
Burtnik A, Foster D and Coppolino MG: VAMP3, syntaxin-13 and SNAP23
are involved in secretion of matrix metalloproteinases, degradation
of the extracellular matrix and cell invasion. J Cell Sci.
122:4089–4098. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Horejs CM, Serio A, Purvis A, Gormley AJ,
Bertazzo S, Poliniewicz A, Wang AJ, DiMaggio P, Hohenester E and
Stevens MM: Biologically-active laminin-111 fragment that modulates
the epithelial-to-mesenchymal transiton in embryonic stem cells.
Proc Natl Acad Sci USA. 111:5908–5913. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Niu H, Zhang Y, Wu B, Zhang Y, Jiang H and
He P: Matrine induces the apoptosis of lung cancer cells through
downregulation of inhibitor of apoptosis proteins and the Akt
signaling pathway. Oncol Rep. 32:1087–1093. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang CY, Bai XY and Wang CH: Traditional
Chinese medicine: A treasured natural resource of anticancer drug
research and development. Am J Chin Med. 42:543–559. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ou X, Chen Y, Cheng X, Zhang X and He Q:
Potentiation of resveratrol-induced apoptosis by matrine in human
hepatoma HepG2 cells. Oncol Rep. 32:2803–2809. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ding Y, Li X, Hong D, Jiang L, He Y and
Fang H: Slience of MACC1 decreases cell migration and invasion in
human maliganant melanoma through inhibiting the EMT. Biosci
Trends. 10:258–264. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu S, Ye D, Xu D, Liao Y, Zhang L, Liu L,
Yu W, Wang Y, He Y, Hu J, et al: Autocrine epiregulin activates
EGFR pathway for lung metastasis via EMT in salivary adenoid cystic
carcinoma. Oncotarget. 7:25251–25263. 2016.PubMed/NCBI
|
|
17
|
Beerling E, Seinstra D, de Wit E, Kester
L, van der Velden D, Maynard C, Schäfer R, van Diest P, Voest E,
van Oudenaarden A, et al: Plasticity between epithelial and
mesenchymal states unlinks EMT from metastasis-enhancing stem cell
capacity. Cell Rep. 14:2281–2288. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ndo Goncalves N, Colombo J, Lopes JR,
Gelaleti GB, Moschetta MG, Sonehara NM, Hellmén E, Cde Zanon F,
Oliani SM and Zuccari DA: Effect of melatonin in epithelial
mesenchymal transition markers and invasive properties of breast
cancer stem cells of canine and human cell lines. PLoS One.
11:e01504072016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin SY, Lee YX, Yu SL, Chang GC and Chen
JJ: Phosphatase of regenerating liver-3 inhibits invasiveness and
proliferation in non-small cell lung cancer by regulating the
epithelial-mesenchymal transition. Oncotarget. 7:21799–21811.
2016.PubMed/NCBI
|
|
20
|
Reig M, Mariño Z, Perelló C, Iñarrairaegui
M, Ribeiro A, Lens S, Díaz A, Vilana R, Darnell A, Varela M, et al:
High rate of early tumor recurrence in patients with HCV-related
HCC undergoing interferon-free therapy. J Hepatol. 65:719–726.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu J, Hu G, Dong Y, Ma R, Yu Z, Jiang S,
Han Y, Yu K and Zhang S: Matrine induces Akt/mTOR signalling
inhibition-mediated autophagy and apoptosis in acute myeloid
leukaemia cells. J Cell Mol Med. 21:1171–1181. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang H, Du T, Xu G, Lai Y, Fan X, Chen X,
Li W, Yue F, Li Q, Liu L and Li K: Matrine suppresses invasion of
castration-resistant prostate cancer cells by downregulating
MMP-2/9 via NF-κM signaling pathway. Int J Oncol. 50:640–648. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu L, Wang G, Liu S, Wei J, Zhang S, Li M,
Zhou G and Wang L: Synthesis and biological evaluation of matrine
derivatives containing benzo-enpyrone structure as potent anti-lung
cancer agents. Sci Rep. 6:359182016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Y, Qi Y, Bai ZH, Ni CX, Ren QH, Xu WH,
Xu J, Hu HG, Qiu L, Li JZ, et al: A novel matrine derivate inhibits
differentiated human hepatoma cells and hepatic cancer stem-like
cells by suppressing PI3K/AKT signaling pathways. Acta Pharmacol
Sin. 38:120–132. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sato R, Semba T, Saya H and Arima Y:
Concise review: Stem cells and epithelial-mesenchymal transition in
cancer: Biological implications and therapeutic targets. Stem
Cells. 34:1997–2007. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bronsert P, Enderle-Ammour K, Bader M,
Timme S, Kuehs M, Csanadi A, Kayser G, Kohler I, Bausch D, Hoeppner
J, et al: Cancer cell invasion and EMT marker expression: A
three-dimensional study of the human cancer-host interface. J
Pathol. 234:410–422. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Canesin G, Cuevas EP, Santos V,
López-Menéndez C, Moreno-Bueno G, Huang Y, Csiszar K, Portillo F,
Peinado H, Lyden D and Cano A: Lysyl oxidase-like 2 (LOXL2) and E47
EMT factor: Novel partners in E-cadherin repression and early
metastasis colonization. Oncogene. 34:951–964. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Qu M, Yu J, Liu H, Ren Y, Ma C, Bu X and
Lan Q: The candidate tumor suppressor gene SLC8A2 inhibits
invasion, angiogenesis and growth of glioblastoma. Mol Cells.
40:761–772. 2017.PubMed/NCBI
|
|
29
|
Zhang J, Liu D, Feng Z, Mao J, Zhang C, Lu
Y, Li J, Zhang Q, Li Q and Li L: MicroRNA-138 modulates metastasis
and EMT in breast cancer cells by targeting vimentin. Biomed
Pharmacother. 77:135–141. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Islam SS, Mokhtari RB, El Hout Y, Azadi
MA, Alauddin M, Yeger H and Farhat WA: TGF-β1 induces EMT
reprogramming of porcine bladder urothelial cells into collagen
producing fibroblasts-like cells in a Smad2/Smad3-dependent manner.
J Cell Commun Signal. 8:39–58. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu YM, Chen ZJ, Liu H, Wei WD, Lu LL, Yang
XL, Liang WT, Liu T, Liu HL, Du J and Wang HS: Inhibition of ERRα
suppresses epithelial mesenchymal transition of triple negative
breast cancer cells by directly targeting fibronectin. Oncotarget.
6:25588–25601. 2015.PubMed/NCBI
|
|
32
|
Jin Y, Shenoy AK, Doernberg S, Chen H, Luo
H, Shen H, Lin T, Tarrash M, Cai Q, Hu X, et al: FBXO11 promotes
ubiquitination of the Snail family of transcription factors in
cancer progression and epidermal development. Cancer Lett.
362:70–82. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Verdone JE, Parsana P, Veltri RW and
Pienta KJ: Epithelial-mesenchymal transition in prostate cancer is
associated with quantifiable changes in nuclear structure.
Prostate. 75:218–224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Colangelo T, Fucci A, Votino C, Sabatino
L, Pancione M, Laudanna C, Binaschi M, Bigioni M, Maggi CA, Parente
D, et al: MicroRNA-130b promotes tumor development and is
associated with poor prognosis in colorectal cancer. Neoplasia.
15:1086–1099. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lv XQ, Qiao XR, Su L and Chen SZ: Honokiol
inhibits EMT-mediated motility and migration of human non-small
cell lung cancer cells in vitro by targeting c-FLIP. Acta Pharmacol
Sin. 37:1574–1586. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tong D, Liu Q, Liu G, Xu J, Lan W, Jiang
Y, Xiao H, Zhang D and Jiang J: Metformin inhibits
castration-induced EMT in prostate cancer by repressing
COX2/PGE2/STAT3 axis. Cancer Lett. 389:23–32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim EK, Choi EJ and Debnath T: Role of
phytochemicals in the inhibition of epithelial-mesenchymal
transition in cancer metastasis. Food Funct. 7:3677–3685. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kallifatidis G, Hoy JJ and Lokeshwar BL:
Bioactive natural products for chemoprevention and treatment of
castration-resistant prostate cancer. Semin Cancer Biol.
40–41:160–169. 2016. View Article : Google Scholar
|
|
39
|
Murden K, San KK, Martino A and Ezekiel U:
The effect of phytochemicals on a chemoresistant,
epithelial-mesenchymal transitioned, colorectal cancer cell line.
FASEB J. 30(1090): 22016.
|
|
40
|
Wang Z, Wu Y, Wang Y, Jin Y, Ma X, Zhang Y
and Ren H: Matrine inhibits the invasive properties of human glioma
cells by regulating epithelial-to-mesenchymal transition. Mol Med
Rep. 11:3682–3686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lim W, Yang C, Bazer FW and Song G:
Luteolin inhibits proliferation and induces apoptosis of human
placental choriocarcinoma cells by blocking the PI3K/AKT pathway
and regulating sterol regulatory element binding protein activity.
Biol Reprod. 95:822016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Winnay JN, Solheim MH, Dirice E, Sakaguchi
M, Noh HL, Kang HJ, Takahashi H, Chudasama KK, Kim JK, Molven A, et
al: PI3-kinase mutation linked to insulin and growth factor
resistance in vivo. J Clin Invest. 126:1401–1412. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jing X, Cheng W, Wang S, Li P and He L:
Resveratrol induces cell cycle arrest in human gastric cancer
MGC803 cells via the PTEN-regulated PI3K/Akt signaling pathway.
Oncol Rep. 35:472–478. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Su R, Nan H, Guo H, Ruan Z, Jiang L, Song
Y and Nan K: Associations of components of PTEN/AKT/mTOR pathway
with cancer stem cell markers and prognostic value of these
biomarkers in hepatocellular carcinoma. Hepatol Res. 46:1380–1391.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jin H, Sun Y, Wang S and Cheng X: Matrine
activates PTEN to induce growth inhibition and apoptosis in
V600EBRAF harboring melanoma cells. Int J Mol Sci. 14:16040–16057.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Du R, Liu Z, Hou X, Fu G, An N and Wang L:
Trichostatin A potentiates genistein-induced apoptosis and reverses
EMT in HEp2 cells. Mol Med Rep. 13:5045–5052. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ru NY, Wu J, Chen ZN and Bian H:
HAb18b18G/CD147 is involved in TGF-β-induced epithelial-mesenchymal
transition and hepatocellular carcinoma invasion. Cell Biol Int.
39:44–51. 2015. View Article : Google Scholar : PubMed/NCBI
|