Introduction
Alzheimer's disease (AD), which is caused by the
loss of neurons and synapse degeneration, is clinically
characterized by changes in cognitive function (1), which affects >47 million people
worldwide (2). Due to its complex
pathological mechanism, its therapeutic targets have not yet been
identified (3). A widely accepted
hypothesis is that oxidative stress serves an important role in the
progression of AD as amyloid β (Aβ) aggregation has been observed
and often accompanied with neurotoxic reactive oxygen species (ROS)
production (4). Glutamate, a
neurotransmitter in the central nervous system, contributes to this
process by causing the accumulation intracellular ROS (5). ROS overproduction and Aβ aggregation
are not only responsible for synaptic dysfunction, but also cause
mitochondria-mediated apoptosis (6).
Until now, the agents applied in clinics have failed
to successfully treat patients with AD. It has been reported that
antioxidants can help maintain cellular redox homeostasis, which
may have beneficial effects on AD (7). Natural products constitute a large
and underappreciated source of candidate agents for AD treatment
(8,9). By modulating mitochondria-mediated
apoptotic signaling, Sparassis crispa polysaccharide has
been demonstrated to successfully protect PC12 cells against
L-glutamic acid (L-Glu) damage (10). In an AlCl3- and
D-galactose (D-gal)-induced AD mouse model, which exhibits AD-like
behaviors, Hericium erinaceus alleviated AD-like symptoms
via the modulation of neurotransmitter levels and apoptosis
associated with mitochondrial function (11). Scutellarin (SC), mainly extracted
from the Chinese herb Erigeron breviscapus (vant.), has been
reported to possess various pharmacological benefits, particularly
for neurological disorders (12).
SC can attenuate neuronal injury by restoring the imbalance of
excitatory vs. inhibitory amino acids and reducing oxidative stress
(12). In addition, SC has been
demonstrated to protect rats against ischemic injury mainly via the
downregulation of endogenous metabolite levels (13).
Based on previous research, the present study aimed
to investigate the protective effects of SC against L-Glu-induced
HT22 cell damage, and AlCl3- and D-gal-induced AD-like
behavior. The results indicated that SC enhanced cell viability,
reduced cell apoptosis, restored mitochondrial function, and
regulated anti- and pro-apoptotic protein expression in
L-Glu-induced HT22 cells. In the AD mouse model in the present
study, SC successfully improved behavior, physiological and
biochemical indices. The data suggested that SC may be used as an
adjuvant therapy in AD and exerts its effects via the regulation of
oxidative stress-induced mitochondrial-mediated apoptosis.
Materials and methods
Cell culture
The mouse hippocampal neuronal HT22 cell line (cat.
no. BNCC-337709; Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences, Shanghai, China), was cultured in
Dulbecco's modified Eagle's medium (DMEM; Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (Invitrogen; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 100 µg/ml streptomycin (Invitrogen; Thermo Fisher
Scientific, Inc.) in a humidified atmosphere containing 5%
CO2 at 37°C.
Cell viability, lactate dehydrogenase
(LDH) release and caspase-3 activity analysis
HT22 cells were seeded into 96-well plates (6,000
cells/100 µl/well), pre-treated with 5 and 15 µM SC (CAS no.
27740-01-8; Shanghai YuanYe Biotechnology Co. Ltd., Shanghai,
China), respectively for 3 h, and then exposed to 25 mM L-Glu
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for a further 24 h
at 37°C. The cell culture medium was collected to measure the
release of LDH using an in vitro Toxicology Assay kit
[Cytotoxicity Detection Kit (LDH); Sigma-Aldrich; Merck KGaA],
according to a previous study (14). A MTT assay (Sigma-Aldrich; Merck
KGaA) was used to detect cell viability as previously described
(15).
A total of 5×105 HT22 cells were seeded
into 6-well plates, pre-treated with 5 and 15 µM SC for 3 h, and
then exposed to 25 mM L-Glu for a further 24 h as aforementioned.
Caspase-3 activity was analyzed using a Caspase-3 activity assay
kit (cat. no. G015; Nanjing Jiancheng Bioengineering Institute,
Nanjing, China).
Cell apoptosis assay
A total of 5×105 of HT22 cells were
seeded into 6-well plates, pre-treated with 5 and 15 µM SC for 3 h,
and then exposed to 25 mM L-Glu for a further 24 h as
aforementioned. Cells were collected and stained using a Dead Cell
Kit (propidium iodide and Annexin V; cat. no. MCH100105; EMD
Millipore, Billerica, MA, USA) for 15 min at 25°C in the dark. The
Muse® Cell Analyzer from EMD Millipore was used to
analyze cell fluorescence.
Assessment of intracellular ROS levels
and dissipation of mitochondrial membrane potential (MMP)
A total of 5×105 HT22 cells were seeded
into 6-well plates, pre-treated with 5 and 15 µM SC for 3 h, and
then exposed to 25 mM L-Glu for a further 12 h, which was
determined according to the results of preliminary experiments.
Treated cells were stained with 5 µM 2,7-dichlorofluorescein
diacetate (Sigma-Aldrich; Merck KGaA) for 20 min at room
temperature in the dark, and the intracellular ROS levels were
observed using a fluorescence microscope (magnification, ×200; Axio
Observer Z1; Carl Zeiss AG, Oberkochen, Germany). Quantitative data
analysis was performed using ImageJ software version 1.46 (National
Institutes of Health, Bethesda, MD, USA) and data were expressed as
the green fluorescence intensity.
The treated cells were also stained with 2 µM JC-1
(Sigma-Aldrich; Merck KGaA) for 15 min at room temperature in the
dark. Alterations in fluorescence were detected using a
fluorescence microscope (magnification, ×200; Axio Observer Z1;
Carl Zeiss AG). The data were analyzed with ImageJ software and
expressed as the ratio of red to green fluorescence intensity.
Western blot analysis
A total of 5×105 HT22 cells were seeded
into 6-well plates, pre-treated with 5 and 15 µM SC for 3 h, and
then exposed to 25 mM L-Glu for a further 24 h as aforementioned.
Treated cells were lysed with radioimmunoprecipitation assay buffer
(Sigma-Aldrich; Merck KGaA) containing 1% protease inhibitor
cocktail (Sigma-Aldrich; Merck KGaA). Following the determination
of protein concentration using a bicinchoninic acid assay protein
kit (EMD Millipore), 40 µg protein was loaded onto the gel and
separated by 12% SDS-PAGE. The separated proteins were
electrophoretically transferred onto 0.45 µm nitrocellulose
membranes (Bio Basic, Inc., Markham, ON, Canada). The membranes
were then blocked using 5% bovine serum albumin (Beijing Dingguo
Changsheng Biotechnology Co., Ltd., Beijing, China) at 4°C for 4 h.
The membranes were incubated with primary antibodies at a dilution
of 1:2,000 for 12 h at 4°C: Cleaved caspase-3 (cat. no. 9661),
B-cell lymphoma 2 (Bcl-2; cat. no. 3498), B-cell lymphoma-extra
large (Bcl-xL; cat. no. 2764), Bcl-2-associated X protein (Bax;
cat. no. 14796) and GAPDH (cat. no. 5174), and were all Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). Following three washes with
Tris-buffered saline containing 0.1% Tween-20, the membranes were
incubated with horseradish peroxidase-conjugated anti-rabbit IgG
secondary antibodies (cat. no. 7074; Santa Cruz Biotechnology,
Inc.) at a dilution of 1:3,000 for 3 h at 25°C. Enhanced
chemiluminescence kit (GE Healthcare Life Sciences, Little
Chalfont, UK) was used to visualize the bands and ImageJ software
was used to quantify the band intensity. Assays were performed in
triplicate.
AD mouse model and treatment
The Institution Animal Ethics Committee of Jilin
University approved the experimental protocol (approval no.
20170012) conducted in the present study. A total of 42 Balb/c male
mice (10-weeks-old, 20–24 g) were purchased from the Norman Bethune
University of Medical Science, Jilin University (Changchun, China)
and housed in cages in a temperature-controlled room at 23±1°C with
40–60% humidity and a 12/12 h light/dark cycle, as well as
sufficient access to water and food.
Of the 42 mice, 30 were subcutaneously injected 120
mg/kg D-gal and orally treated with 20 mg/kg AlCl3 once
per day for 8 weeks to establish an AD mice model similar to a
previous study (16). At the fifth
week, only 24 mice were retained for further experimentation, as
the remaining 6 mice failed to exhibit AD-associated behaviors in
the water maze test, and these 24 mince were then randomly divided
into two groups, which were either treated orally with normal
saline solution (n=12) or 20 mg/kg of SC (n=12) for 4 weeks. Normal
mice (n=12) treated orally with normal saline solution for 8 weeks,
served as the control group. From week 9, behavioral tests,
including autonomic activity and a Morris water maze tests, were
performed according to a previous study without modifications
(16). Orbital blood (300 µl) was
sampled via peritoneal injection following anesthesia with 400
mg/kg of 10% chloral hydrate. Following euthanasia by injection
with 150 mg/kg 1.5% pentobarbital, whole brain tissues were
collected.
Determination of acetylcholine (Ach),
choline acetyltransferase (ChAT), superoxide dismutase (SOD), ROS
and Aβ1–42 levels in the brain and/or serum
The levels of Ach [cat. nos. A105-1 (for brain
tissues) and A105-2 (for serum); Nanjing Jiancheng Bioengineering
Institute, Nangjing, China], ChAT [cat. nos. A079-1 (for brain
tissues) and A079-2 (for serum); Nanjing Jiancheng Bioengineering
Institute], Aβ1-42 (cat. no. E-20118; Shanghai Yuanye Biological
Technology Co., Ltd., Shanghai, China) and SOD (cat. no. A001-3;
Shanghai Yuanye Biological Technology Co., Ltd.) in the serum and
brain tissues, as well as ROS (cat. no. E-20634; Shanghai Yuanye
Biological Technology Co., Ltd., Shanghai, China) in brain tissues
alone, were measured using ELISA kits.
Immunohistochemistry
Following collection, the brain tissues were fixed
in 10% neutral formalin at room temperature for 48 h, and were then
desiccated and embedded in paraffin (7 µm thick). The sections were
then soaked in a descending alcohol series and then washed with
deionized water and PBS for 5 min each. Following this, the
paraffin sections were soaked in 0.01 M boiled citrate buffer at
95°C for 20 min in order to perform antigen retrieval. The protein
expression levels of Aβ1–42 and phosphorylated (p)-Tau in brain
tissue were detected via immunohistochemical analysis. Briefly, the
slides of brain samples were heated in citrate buffer for 15 min to
retrieve antigens, incubated with 3% hydrogen peroxide for 10 min
at room temperature to block endogenous peroxidase and by blocked
with 2% goat serum (Histostain™-Plus Kits; cat. no. SP-0023;
Nanjing Jiancheng Bioengineering Institute) dissolved in PBS
(16). The slides were the
incubated with antibodies against Aβ1–42 (1:200; cat. no. bs-0877R;
Beijing Biosynthesis Biotechnology Co., Ltd., Beijing, China) and
p-Tau (1:200; cat. no. bs-2392R; Beijing Biosynthesis Biotechnology
Co., Ltd.) for 12 h at 4°C. After washing with PBS, the slides were
incubated with a horseradish peroxidase-conjugated secondary
antibody (1:2,000; cat. no. sc-7074; Santa Cruz Biotechnology,
Dallas, TX, USA) at 25°C for 4 h. The slides were visualized with
3,3′-diaminobenzidine for 3 min at room temperature and Mayer's
hematoxylin for 3 min at room temperature, and were dehydrated with
ethanol and xylene. Finally, images were captured using an Olympus
IX73 light microscope (magnification, ×100; Olympus, Tokyo, Japan).
The intensity of Aβ1–42 and p-Tau expression was analyzed using
ImageJ software.
Statistical analysis
All experimental data are presented as the mean ±
standard deviation. Statistical data were analyzed using one-way
analysis of variance followed by a Duncan's multiple range post hoc
test. Statistical analysis was conducted using SPSS v.16.0 software
(SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
SC protects HT22 cells against
L-Glu-induced damage
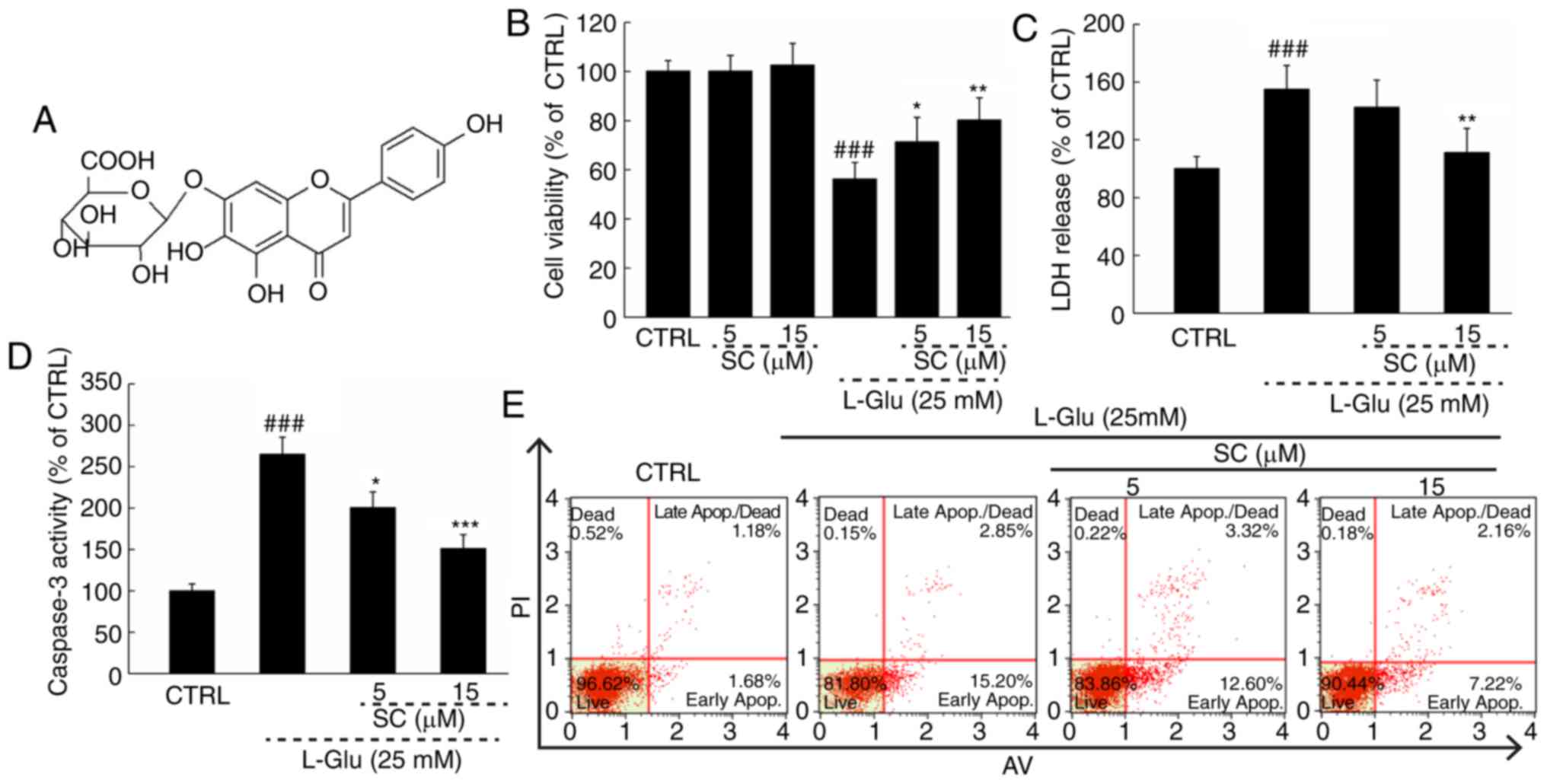
Normal HT22 cells treated with SC (Fig. 1A) alone exhibited no significant
alterations in cell viability compared with in the control;
however, a significant decrease in cell viability was observed in
response to L-Glu (P<0.01; Fig.
1B). Treating L-Glu-damaged cells with SC improved cell
viability by >12% (P<0.05; Fig.
1B). Pre-treatment with 15 µM SC significantly reduced LDH
release by 43.9% (P<0.01; Fig.
1C) and caspase-3 activity by 98.7% (P<0.001; Fig. 1D) compared with L-Glu-treated HT22
cells. In addition, HT22 cells exposed to 25 mM L-Glu exhibited an
apoptosis rate of 18.1% (Fig. 1E).
The cell apoptosis was reduced to 9.4% in cells pre-treated with 15
µM SC (Fig. 1E).
SC reduces ROS accumulation, restores
MMP dissipation, and regulates anti- and pro-apoptotic protein
expression
In L-Glu-treated HT22 cells, levels of intracellular
ROS accumulation were indicated by enhanced green fluorescence;
significant ROS accumulation was observed in response to L-Glu
treatment compared with in the control (Fig. 2A). Dissipation of MMP was also
observed, as indicated by a change of red to green fluorescence
(Fig. 2B). Pre-treatment with 5
and 15 µM SC significantly suppressed the accumulation of ROS
(P<0.001; Fig. 2A) and restored
MMP dissipation compared with in the L-Glu-treated HT22 cells
(P<0.001; Fig. 2B).
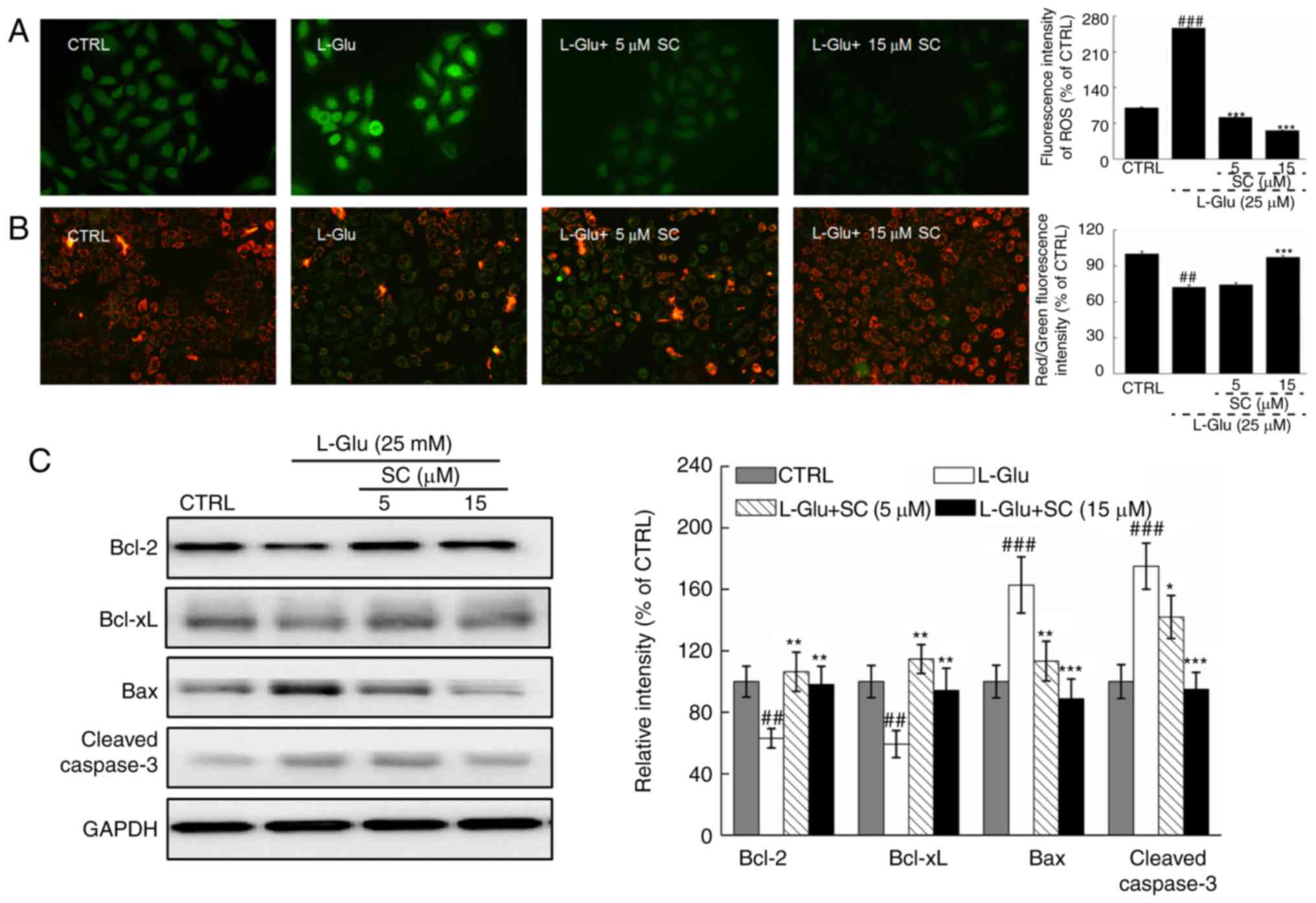 | Figure 2.SC reduces ROS accumulation, restores
MMP dissipation, and regulates anti- and pro-apoptotic protein
expression. HT22 cells were pre-incubated with 5 and 15 µM SC for 3
h, and exposed to L-Glu for another 12 h. (A) Representative
immunofluorescent images of HT22 stained with
2,7-dichlorofluorescein diacetate to visualize intracellular ROS
(magnification, ×200; n=6). SC suppressed the accumulation of ROS,
as demonstrated by reductions in green fluorescence intensity. (B)
Immunofluorescence staining with JC-1 revealed that SC restored the
dissipation of MMP (magnification, ×200; n=6). The quantitative
data of the ratio of red to green fluorescence intensity is
presented on the right. (C) Following treatment with SC, cells were
treated with L-Glu for another 24 h. Western blot analysis
demonstrated that 15 µM SC was able to restore the expression of
anti-apoptotic (Bcl-2 and Bcl-xL) and pro-apoptotic (Bax and
cleaved caspase-3) proteins to normal levels. Quantification data
were normalized to GAPDH and expressed as a percentage of CTRL
(n=3). ##P<0.01, ###P<0.001 vs. CTRL;
*P<0.05, **P<0.01, ***P<0.001 vs. L-Glu. Bax,
Bcl-2-associated X protein; Bcl-2, B-cell lymphoma 2; Bcl-xL,
B-cell lymphoma-extra large; CTRL, control; L-Glu, L-glutamic acid;
MMP, mitochondrial membrane potential; ROS, reactive oxygen
species; SC, scutellarin. |
L-Glu-damaged cells revealed low protein expression
levels of Bcl-2 and Bcl-xL, but high protein expression levels of
Bax and cleaved caspase-3 compared with in control HT22 cells
(P<0.01; Fig. 2C).
Pre-incubation with SC resulted in >35.1 and 33.8% increase in
Bcl-2 and Bcl-xL expression levels, respectively (P<0.01;
Fig. 2C). Conversely, Bax
(P<0.01) and cleaved caspase-3 (P<0.05; Fig. 2C) expression levels were reduced by
>48.7 and 33.6%, respectively. These results suggested that SC
protected HT22 cells against L-Glu-induced apoptosis via regulation
of mitochondrial function.
SC alleviates behavioral symptoms in
AD mice
Behavioral testing was performed in a D-gal plus
AlCl3 mouse model of AD to investigate the therapeutic
effects of SC on AD. Compared with non-treated AD mice, SC
significantly enhanced the number of vertical movements by 44.6%
(P<0.01; Fig. 3A) and
horizontal movements by 35.2% (P<0.01; Fig. 3B) in an autonomic activity
test.
 | Figure 3.SC alleviates the clinical features
of AD in vivo. In an AlCl3 plus D-gal-induced
mouse model of AD, 4-week SC administration enhanced (A) vertical
and (B) horizontal movements in an autonomous activity test (n=12).
(C) SC treatment decreased the escape latency time in a water maze
test (n=12). (D) SC enhanced the serum levels of Aβ1-42, but
reduced the levels of Aβ1–42 in brain lysate as determined via
ELISA (n=12). Immunohistochemistry revealed that SC reduced (E)
Aβ1–42 and (F) p-Tau expression levels in the hippocampus (n=6).
The quantitative data were expressed as percentage protein density
of CTRL. Data are expressed as the mean ± standard deviation.
#P<0.05, ##P<0.01,
###P<0.001 vs. CTRL; *P<0.05, **P<0.01 and
***P<0.001 vs. AlCl3 plus D-gal. Aβ1-42, amyloid
β1-42; AD, Alzheimer's disease; AlCl3, aluminum
chloride; CTRL, control; D-gal, D-galactose; p-Tau,
phosphorylated-Tau; SC, scutellarin. |
The water maze test can be applied to evaluate the
learning and memory abilities of animals, and is commonly performed
in AD research (17). Compared
with in heathy mice, the escape latency time was significantly
increased by 56.8% in AD mice (P<0.01; Fig. 3C). Administration of SC for 4 weeks
led to a 26.3% reduction in escape latency time compared with in AD
mice (P<0.05; Fig. 3C). The
attenuation of AD-associated symptoms following treatment with SC
further suggested that SC exhibits therapeutic effects in AD in
mice models.
SC regulates Aβ1–42 expression levels
and p-Tau aggregation
Aβ1-42, which has strong aggregating properties, has
been reported to be the key component of amyloid plaques (16). Following 4-week SC administration,
the serum Aβ1–42 concentrations were increased by 48.1% compared
with non-treated AD mice (P<0.05; Fig. 3D), whereas Aβ1–42 concentrations in
brain were reduced by 19.6% (P<0.05; Fig. 3D). The results of
immunohistochemical analysis indicated that, compared with in the
nontreated SD group, SC significantly reduced the Aβ1–42 levels in
brain, particularly in the region of the hippocampus (P<0.001;
Fig. 3E).
Compared with control mice, the expression levels of
p-Tau were significantly enhanced in the hippocampus of AD mice;
however, SC administration significantly reduced the levels of
p-Tau (P<0.001; Fig. 3F). The
attenuation of effects associated with AD following treatment with
SC may be due to resultant decreased levels of Aβ1–42 and
p-Tau.
SC regulates Ach, ChAT, SOD and
ROS
Compared with in control mice, the levels of SOD,
Ach and ChAT in the serum and brain lysate samples were
significantly lower in AD mice (P<0.05), whereas significantly
increased levels of ROS were detected in the brain lysates of AD
mice (P<0.01; Fig. 4A-D). For
factors associated with oxidative stress, SC treatment
significantly enhanced the concentration of SOD in the serum and
brain lysate (P<0.05; Fig. 4B),
and reduced the levels of ROS in the brain lysate of AD mice
(P<0.01; Fig. 4A) compared with
in non-treated AD mice. In terms of cholinergic function, SC
significantly increased the levels of Ach (P<0.01; Fig. 4C) and ChAT (P<0.01; Fig. 4D) in the serum and brains of AD
mice compared with in the non-treated AD groups. The results
suggested that regulation of oxidative stress and cholinergic
function are associated with protective effects exhibited by SC
against AD.
 | Figure 4.SC regulates ROS, SOD, Ach and ChAT
in vivo. In an AlCl3 plus D-gal-induced
Alzheimer's disease mouse model, 4-week SC administration (A)
reduced ROS levels in brain lysate, and enhanced the levels of (B)
SOD, (C) Ach and (D) ChAT in the serum and brain lysate, as
measured by ELISA. Data are expressed as the mean ± standard
deviation (n=12). #P<0.05, ##P<0.01,
###P<0.001 vs. CTRL; *P<0.05, **P<0.01 vs.
AlCl3 plus D-gal. Aβ1-42, amyloid β1-42; Ach,
acetylcholine; AlCl3, aluminum chloride; ChAT, choline
acetyltransferase; CTRL, control; D-gal, D-galactose; ROS, reactive
oxygen species; SC, scutellarin; SOD, superoxide dismutase. |
Discussion
The present study revealed the protective effects of
SC against L-Glu-induced HT22 cell damage, as demonstrated by the
amelioration of mitochondrial-mediated apoptosis. SC restored the
MMP dissipation and suppressed the accumulation of intracellular
ROS in L-Glu-damaged HT22 cells. Glutamate inhibits cysteine
uptake, leading to necrosis and apoptosis due to the disequilibrium
between oxidation and antioxidation, which affects ROS levels
(18). High levels of ROS helps
open mitochondrial permeability transition pores, resulting in the
dissipation of MMP (19). A short
feedback loop has been proposed between ROS levels and
mitochondrial function, whereby enhanced cytoplasm ROS levels may
lead to the dissipation of MMP and subsequent release of
mitochondrial ROS as well as other pro-apoptotic molecules, such as
caspase-3, from the mitochondria to the cytoplasm (20). In addition, SC enhanced the
expression levels of Bcl-2 and Bcl-xL, and reduced the levels of
Bax in L-Glu-treated HT22 cells. Bcl-2 family members are present
in the membrane of mitochondria and directly influence
mitochondrial function by regulating the opening of mitochondrial
permeability transition pores (21). These findings suggested that
SC-mediated neuroprotection against L-Glu-induced neurotoxicity in
HT22 cells may be associated with oxidative stress-induced
mitochondrial-mediated apoptosis.
In AD mice, long-term D-gal injections cause ROS
accumulation and mitochondrial dysfunction (22). Polyunsaturated fatty acids in the
brain can be easily damaged by oxidative stress (23), which in turn affects learning and
memory abilities (24). In
addition, aluminum can form a complex with Aβ that can more readily
access the brain and is responsible for cerebrovascular
dysregulation (25). Aβ
aggregation in the brain, serving as a hallmark of spatial memory
deficit and cognitive dysfunction, can also cause imbalance to the
oxidative stress system by inducing the overproduction of ROS
during the progression of AD (26). However, the clearance of Aβ
aggregation in the brains of patients with AD can increase Aβ
levels in peripheral blood (27).
The aggregation of Aβ may also further initiate the Tau pathology,
contributing to the formation of neurofibrillary tangles (16). In the present study, mice with AD
induced by AlCl3 and D-gal revealed that SC enhanced the
levels of SOD in the serum and brain lysate, reduced the levels of
ROS in brain lysate, increased the concentration of Aβ1–42 in the
serum, and suppressed Aβ1–42 aggregation and Tau protein deposition
in the hippocampus area. SOD, an endogenous antioxidant, has been
reported as the first-line of defense against oxidative damage
(28). Combined with the in
vitro results, SC-induced improvement in the cognitive
performance of AD mice may be, at least partially, associated with
its modulation of the balance of oxidative stress, which suppressed
cell apoptosis.
In the present study, it was also revealed that SC
significantly enhanced the levels of Ach and ChAT in the serum and
brain lysate of AD mice. Impaired cholinergic function has been
noted in patients with AD, and in particular, low levels of Ach and
ChAT have been reported to contribute to cognitive and memory
decline (29). Ach, dynamically
controlled by AchE, is responsible for the storage and recovery of
long-term memory (28). Previous
studies demonstrated that Armillaria mellea and Amanita
caesarea can improve AD-like behaviors via enhancing the levels
of Ach and ChAT in the serum and brains of AlCl3- and
D-gal induced AD mice (15,17).
The data in the present study suggested that SC-mediated protection
against AD may be associated with its regulation of cholinergic
function. However, the association between cholinergic transmitters
and oxidative stress remains unknown and further investigation is
required.
In conclusion, SC exhibited neuroprotective effects
against L-Glu-induced neurotoxicity in HT22 cells and alleviated
AD-like behaviors in mice, which may largely be attributable to its
antioxidation and antiapoptotic properties. These findings support
SC as a potential agent for the prevention and/or adjuvant therapy
for AD.
Acknowledgements
The authors would like to thank Professor Yang Liu
for her revision of the manuscript.
Funding
The present study was supported by the Project from
Health and Family Planning Commission Project from Jiangsu Province
(grant no. H201536), Scientific Research Program of the Affiliated
Hospital of Jiangsu University in China (grant no. jdfyRC-2015004)
and the Science and Technology Project of ‘13th Five-Year Plan’ of
Department of Education in Jilin Province of China (grant no.
JJKH20170822KJ).
Availability of data and materials
All data generated and analyzed during the present
study are included in this published article.
Authors' contributions
XW and LK designed the experiments; XH, ST, JH, MD
and XS performed the experiments and processed data; XH, XW and LK
wrote the manuscript; MD, XW and LK revised the manuscript.
Ethics approval and consent to
participate
Institution Animal Ethics Committee of Jilin
University approved the experimental protocol (approval no.
20170012).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rygiel K: Novel strategies for Alzheimer's
disease treatment: An overview of anti-amyloid beta monoclonal
antibodies. Indian J Pharmacol. 48:629–636. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xu T, Niu C, Zhang X and Dong M:
β-Ecdysterone protects SH-SY5Y cells against β-amyloid-induced
apoptosis via c-Jun N-terminal kinase- and Akt-associated
complementary pathways. Lab Invest. 98:489–499. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rosello A, Warnes G and Meier UC: Cell
death pathways and autophagy in the central nervous system and its
involvement in neurodegeneration, immunity and central nervous
system infection: To die or not to die-that is the question. Clin
Exp Immunol. 168:52–57. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Daulatzai MA: Cerebral hypoperfusion and
glucose hypometabolism: Key pathophysiological modulators promote
neurodegeneration, cognitive impairment and Alzheimer's disease. J
Neurosci Res. 95:943–972. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Luo P, Fei F, Zhang L, Qu Y and Fei Z: The
role of glutamate receptors in traumatic brain injury: Implications
for postsynaptic density in pathophysiology. Brain Res Bull.
85:313–320. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hu JF, Chu SF, Ning N, Yuan YH, Xue W,
Chen NH and Zhang JT: Protective effect of (−)clausenamide against
Abeta-induced neurotoxicity in differentiated PC12 cells. Neurosci
Lett. 483:78–82. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhao Y and Zhao B: Natural antioxidants in
prevention and management of Alzheimer's disease. Front Biosci
(Elite Ed). 4:794–808. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang D, Tan QR and Zhang ZJ:
Neuroprotective effects of paeoniflorin, but not the isomer
albiflorin, are associated with the suppression of intracellular
calcium and calcium/calmodulin protein kinase II in PC12 cells. J
Mol Neurosci. 51:581–590. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang D, Guo TQ, Wang ZY, Lu JH, Liu DP,
Meng QF, Xie J, Zhang XL, Liu Y and Teng LS: ERKs and
mitochondria-related pathways are essential for glycyrrhizic
acid-mediated neuroprotection against glutamate-induced toxicity in
differentiated PC12 cells. Braz J Med Biol Res. 47:773–779. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hu S, Wang D, Zhang J, Du M, Cheng Y, Liu
Y, Zhang N, Wang D and Wu Y: Mitochondria related pathway is
essential for polysaccharides purified from sparassis crispa
mediated neuro-protection against glutamate-induced toxicity in
differentiated PC12 cells. Int J Mol Sci. 17(pii): E1332016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang J, An S, Hu W, Teng M, Wang X, Qu Y,
Liu Y, Yuan Y and Wang D: The Neuroprotective properties of
hericium erinaceus in glutamate-damaged differentiated PC12 cells
and an Alzheimer's disease mouse model. Int J Mol Sci. 17(pii):
E18102016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tang H, Tang Y, Li N, Shi Q, Guo J, Shang
E and Duan JA: Neuroprotective effects of scutellarin and
scutellarein on repeatedly cerebral ischemia-reperfusion in rats.
Pharmacol Biochem Behav. 118:51–59. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tang H, Tang Y, Li NG, Lin H, Li W, Shi Q,
Zhang W, Zhang P, Dong Z and Shen M: Comparative metabolomic
analysis of the neuroprotective effects of scutellarin and
scutellarein against ischemic insult. PLoS One. 10:e01315692015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang D, Lu J, Liu Y, Meng Q, Xie J, Wang Z
and Teng L: Liquiritigenin induces tumor cell death through
mitogen-activated protein kinase-(MPAKs-) mediated pathway in
hepatocellular carcinoma cells. BioMed Res Int.
2014:9653162014.PubMed/NCBI
|
|
15
|
An S, Lu W, Zhang Y, Yuan Q and Wang D:
Pharmacological basis for use of Armillaria mellea polysaccharides
in Alzheimer's disease: Antiapoptosis and antioxidation. Oxid Med
Cell Longev. 2017:41845622017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao JM, Li L, Chen L, Shi Y, Li YW, Shang
HX, Wu LY, Weng ZJ, Bao CH and Wu HG: Comparison of the analgesic
effects between electro-acupuncture and moxibustion with visceral
hypersensitivity rats in irritable bowel syndrome. World J
Gastroenterol. 23:2928–2939. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li Z, Chen X, Lu W, Zhang S, Guan X, Li Z
and Wang D: Anti-oxidative stress activity is essential for Amanita
caesarea mediated neuroprotection on glutamate-induced apoptotic
HT22 cells and an Alzheimer's disease mouse model. Int J Mol Sci.
18(pii): E16232017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brown DI and Griendling KK: Regulation of
signal transduction by reactive oxygen species in the
cardiovascular system. Circ Res. 116:531–549. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Circu ML and Aw TY: Reactive oxygen
species, cellular redox systems, and apoptosis. Free Radic Biol
Med. 48:749–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lemasters JJ, Qian T, Trost LC, Herman B,
Cascio WE, Bradham CA, Brenner DA and Nieminen AL: Confocal
microscopy of the mitochondrial permeability transition in necrotic
and apoptotic cell death. Biochem Soc Symp. 66:205–222. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gross A and Katz SG: Non-apoptotic
functions of BCL-2 family proteins. Cell Death Differ.
24:1348–1358. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qu Z, Zhang J, Yang H, Huo L, Gao J, Chen
H and Gao W: Protective effect of tetrahydropalmatine against
d-galactose induced memory impairment in rat. Physiol Behav.
154:114–125. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang T, Sun Q and Chen S: Oxidative
stress: A major pathogenesis and potential therapeutic target of
antioxidative agents in Parkinson's disease and Alzheimer's
disease. Prog Neurobiol. 147:1–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nemeth M, Millesi E, Wagner KH and Wallner
B: Sex-specific effects of diets high in unsaturated fatty acids on
spatial learning and memory in guinea pigs. PLoS One.
10:e01404852015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Banks WA, Niehoff ML, Drago D and Zatta P:
Aluminum complexing enhances amyloid beta protein penetration of
blood-brain barrier. Brain Res. 1116:215–221. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim DI, Lee KH, Gabr AA, Choi GE, Kim JS,
Ko SH and Han HJ: Abeta-induced Drp1 phosphorylation through Akt
activation promotes excessive mitochondrial fission leading to
neuronal apoptosis. Biochim Biophys Acta. 1863:2820–2834. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fei M, Jianghua W, Rujuan M, Wei Z and
Qian W: The relationship of plasma Aβ levels to dementia in aging
individuals with mild cognitive impairment. J Neurol Sci.
305:92–96. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Islam MT: Oxidative stress and
mitochondrial dysfunction-linked neurodegenerative disorders.
Neurol Res. 39:73–82. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Farkas E and Luiten PG: Cerebral
microvascular pathology in aging and Alzheimer's disease. Prog
Neurobiol. 64:575–611. 2001. View Article : Google Scholar : PubMed/NCBI
|