Introduction
Breast cancer is one of the most common types of
malignant tumor in women. Previous studies demonstrated that the
rising incidence of breast cancer in Asian women is primarily due
to modifications of reproductive and lifestyle factors, including
reduced fertility, advancing age of the first pregnancy, decrease
in breastfeeding and increase of body mass index (1,2).
Furthermore, the age of the first pregnancy, late-onset menopause,
height and higher education level were positively associated with
high mammographic density (MD) (3). In contrast, age, positive family
history of breast cancer and longer durations of breastfeeding were
negatively associated with MD among Chinese women with an increased
risk of breast cancer (3).
MicroRNAs (miRNAs) are a group of short non-coding RNAs (4). A previous study demonstrated that
miRNAs were involved in the growth and development of breast
cancer, and may be used as biomarkers for cancer diagnosis and
prognosis (5). Therefore, it was
suggested that miRNAs may represent promising targets for treatment
of cancer.
miRNA-133a-3p (miR-133a-3p) is a member of the miRNA
family, and previous studies demonstrated that miR-133a-3p was
downregulated in multiple types of cancer, including prostate
cancer, colorectal cancer, ovarian cancer and breast cancer, and
that the downregulation of miR-133a-3p may lead to tumor growth,
inhibiting apoptosis in cancer cells (6–8). A
previous bioinformatics study identified that miR-133a-3p possessed
great diagnostic potential and its decreased expression level
suggested that it may represent a novel biomarker for bladder
cancer (9). Furthermore, a
previous study suggested that miR-133a-3p modulated the expression
of SUMO-specific protease 1 (SENP1) by binding to its
3′-untranslated region (3′UTR), resulting in its downregulation
(10). Additionally, miR-133a-3p
served a role in colorectal cancer (10). In the present study, it was
identified that miR-133a-3p may bind to the 3′UTR of dual
specificity phosphatase 1 (DUSP1) mRNA using bioinformatics
analysis. Overexpression of miR-133a-3p led to a significant
increase in the DUSP1 expression level. Chen et al (11) demonstrated that progesterone
receptors inhibited the proliferation of breast cancer cells by
inducing DUSP1 expression. Therefore, the present study suggested
that miR-133a-3p may increase the DUSP1 expression level, inducing
DUSP1 downstream genes, serving a role in the regulation of cancer
growth.
Puerarin is the principal bioactive agent isolated
from the root of Pueraria lobata (Willd.) Ohwi, which is
additionally termed Gegen in traditional Chinese medicine (12). Puerarin was involved in the
regulation of certain molecular targets, including miR-16, miR-214
and miR-106b (13–15), modulating cancer cell
proliferation, apoptosis and the cell cycle. Nonetheless, the
effect of puerarin on DUSP1 regulation remains to be investigated.
The present study identified a novel association between puerarin
and DUSP1; the effect of puerarin on breast cancer cells and its
underlying mechanism was investigated by bioinformatics and
cytological analyses.
Materials and methods
Cell culture
The human breast cancer cell HCC38 (Shanghai
Institute of Cell Biology, Shanghai, China) was cultured in
Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.), 100 mg/ml
streptomycin and 100 U/ml penicillin (Beyotime Institute of
Biotechnology, Haimen, China) at 37°C with 5% CO2.
Cell transfection
In total, 5 µl Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) solution was used for
the transfection of miRNA mimics and inhibitor of 20 µM:
miR-133a-3p mimics, 5′-UUUGGUCCCCUUCAACCAGCUG-3′; miR-133a-3p
inhibitor, 5′-GCUGGUUGAAGGGGACCAAAUU-3′. After 24 h of cell
transfection, follow-up experiments were carried out. HCC38 cells
were divided into four groups: Cells treated with 20 µM puerarin
(National Institutes for Food and Drug Control, Beijing, China)
group, miR-133a-3p mimics (Shanghai GenePharma Co., Ltd., Shanghai,
China) transfection group, 20 µM puerarin + miR-133a-3p mimics
transfection group and negative control (20 µM puerarin +
miR-133a-3p mimics control) transfection group. Cells were
transfected at 55–65% confluence. Cells were incubated under the
conditions of 5% CO2 atmosphere at 37°C.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
HCC38 cells were treated with 0, 10, 20 and 40 µM
puerarin (16) in 5%
CO2 atmosphere at 37°C for 24 h. The total RNA was
extracted from the four groups of cells using TRIzol®
reagent (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. A PrimeScript™ RT reagent kit
(Takara Bio, Inc., Otsu, Japan) was used to synthesize the cDNA
(37°C for 15 min; 85°C for 5 sec). The primer sequences are listed
in Table I. RT-qPCR was performed
using SYBR Premix Ex Taq (Takara Bio, Inc.) and the signal was
detected using the ABI-7500 system (Applied Biosystems; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol:
95°C for 30 sec, then 95°C for 5 sec, 56°C for 20 sec and 72°C for
30 sec for 40 cycles. Each RT-qPCR was repeated three times.
miR-133a-3p and DUSP1 mRNA relative expression levels were measured
using the 2−ΔΔCq method and GAPDH was used as reference
gene (17).
 | Table I.Primer sequences used for polymerase
chain reaction. |
Table I.
Primer sequences used for polymerase
chain reaction.
| Gene | Sense primer
(5′→3′) | Antisense primer
(5′→3′) |
|---|
| DUSP1 |
TCGCTGCGAAGGACATTTGGG |
AGCGACGCTTCCTGTAAACCC |
| miR-133a-3p |
GGGAGCCAAATGCTTTGCTAG |
CCCTCGGTTTACGAAACGATC |
| GAPDH |
CGGAGTCAACGGATTTGGTCGTAT |
AGCCTTCTCCATGGTGGTGAAGAC |
| U6 |
TGCGGGTGCTCGCTTCGGCAGC |
CCAGTGCAGGGTCCGAGGT |
Western blotting
The proteins from the four groups of HCC38 cells
were extracted using radioimmunoprecipitation lysis buffer
(Beyotime Institute of Biotechnology) and the protein concentration
was measured using a Bicinchoninic Acid Protein Assay kit (Beyotime
Institute of Biotechnology). The proteins were loaded at 20 µg/lane
and separated by 10% SDS-PAGE. Subsequently, proteins were
transferred onto polyvinylidene difluoride membranes (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). In total, 0.1% TBST was
used to block the transfer membranes, which were subsequently
incubated with 5% non-fat dry milk at 25°C for 1 h, and the primary
antibodies were incubated at 4°C overnight, and second antibody
were incubated for 1 h at room temperature. Densitometric analysis
by DAB visualization reagent (Beyotime Institute of Biotechnology)
was performed to detect protein expression level using ImageJ
software (version 1.49; National Institutes of Health, Bethesda,
MD, USA). The antibodies for cleaved caspase-9 (cat. no. 9505,
1:1,000), B-cell lymphoma 2 (Bcl-2; cat. no. 4223, 1:1,000), Bcl2
associated X (Bax; cat. no. 5023, 1:1,000), p38 (cat. no. 8690,
1:1,000), phosphorylated p38 (p-p38; cat. no. 4511, 1:1,000) and
β-actin (cat. no. 3700, 1:1,000) were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA), and DUSP1 antibody (cat. no.
ab61201, 1:1,000) was purchased from Abcam (Cambridge, UK), and the
secondary antibody (rabbit/mouse horseradish peroxidase-conjugated
anti-goat immunoglobulin G, (cat. no. A0208; 1:1,000) was purchased
from Beyotime Institute of Biotechnology.
Cell viability assay
Cell viability was detected by a Cell Counting Kit-8
(CCK-8; Dojindo Molecular Technologies, Inc., Kumamoto, Japan).
HCC38 cells were cultured in 90 µl complete medium, in 96-well
plates, 2×104 cells/well. Triplicate repetitions wells
were used for each group. Cells were incubated for 8 h and
subsequently treated with various doses of puerarin for 24, 48 and
72 h in 96-well plates at 37°C. Subsequently, 10 µl CCK-8 solution
and 90 µl FBS-free medium were mixed, and added to each well for
incubation for 10 min at 37°C. The absorbance was detected at 450
nm using a microplate reader (BioTek Instruments, Inc., Winooski,
VT, USA) to measure cell viability.
Cell apoptosis analysis by flow
cytometry (FCM)
HCC38 cell apoptosis was measured in the four groups
by FCM, using an Annexin V-fluorescein isothiocyanate (V-FITC)
apoptosis detection kit (Beyotime Institute of Biotechnology).
HCC38 cells were cultured, collected and washed with cold PBS
twice, and subsequently treated with puerarin for 48 h at 37°C.
Cells were treated with Annexin propidium iodide and V-FITC.
Subsequently the cells were centrifuged at 100 × g at 25°C for 5
min. Cell apoptosis was measured using a flow cytometer and FlowJo
software (version 10; FlowJo LLC, Ashland, OR, USA).
Caspase-3 activity analysis
Caspase-3 activity, measured to detect cell
apoptosis, was assessed using a caspase-3 colorimetric activity
assay kit (Beyotime Institute of Biotechnology) according to the
manufacturer's protocol. Cells were collected and washed twice with
cold PBS, and the cells were subsequently lysed with the buffer
provided in the kit. The caspase-3 substrate
N-Acetyl-Asp-Glu-Val-Asp-7-amido-4-Methylcoumarin was used to
incubate an equal amount of protein (100 µg) at 37°C for 2 h.
Caspase-3 activity was measured based on the absorbance at 405 nm
using a plate reader.
Luciferase reporter assay
The interaction between miR-133a-3p and the 3′UTR of
DUSP1 was predicted by TargetScanHuman version 7.1 (www.targetscan.org/vert_71/). The target sequence
was cloned in pcDNA3.1- Luciferase vector (Promega Corporation,
Madison, WI, USA) with firefly and Renilla luciferase using
XbaI. In total, two plasmids were generated: DUSP1 wild-type
(DUSP1-WT) containing the 3′UTR of DUSP1 and the predicted binding
site, and DUSP1-mutant (DUSP1-MT) containing a mutated binding site
in the 3′UTR. The mimics and vectors were co-transfected into HCC38
cells using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) at 37°C for 48 h. Subsequently, the
transfected HCC38 cells were analyzed using a dual-luciferase
reporter assay system (Promega Corporation). Each experiment was
replicated three times independently. Firefly luciferase activity
was normalized to Renilla luciferase activity.
Statistical analysis
Data are presented as the mean ± standard error.
Each experiment was replicated three times independently. All the
data were analyzed using SPSS (17.0; SPSS, Inc., Chicago, IL, USA).
Student's t-test or one-way analysis of variance followed by
Student-Newman-Keuls test was used to analyze the differences
between two or multiple groups, respectively. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-133a-3p expression level is
upregulated by puerarin and miR-133a-3p mimics
Previous studies identified that puerarin inhibited
the development of tumors, and miR-133a-3p served a role in
suppressing cancer (18). The aim
of the present study was to investigate the effect of puerarin on
the miR-133a-3p expression level in breast cancer. The present
results demonstrated that the miR-133a-3p expression level was
increased upon treatment with puerarin in HCC38 cells in a
dose-dependent manner, suggesting that puerarin may possess
antitumor activity (Fig. 1A). The
concentration of 20 µM puerarin demonstrated the best experimental
results. The effect of miR-133a-3p mimics and inhibitor is shown in
Fig. 1B; the miR-133a-3p
expression level was increased following treatment with puerarin
(20 µM), miR-133a-3p mimics and puerarin (20 µM) + miR-133a-3p
mimics compared with the NC group (Fig. 1C).
Puerarin and miR-133a-3p mimics
inhibit cell viability and promotes cell apoptosis
A CCK-8 assay was conducted to test the effect of
puerarin on the viability of breast cancer cells. Flow cytometry
results suggested that the apoptotic rates following treatment with
puerarin (20 µM), miR-133a-3p mimics and puerarin (20 µM) +
miR-133a-3p mimics were significantly increased compared with the
NC group (Fig. 2A and B;
P<0.05). Furthermore, the apoptotic rate of the puerarin (20 µM)
+ miR-133a-3p mimics group was increased compared with the puerarin
and miR-133a-3p mimics groups (Fig. 2A
and B). The CCK-8 assay suggested that puerarin inhibited the
viability of HCC38 cells in a time- and dose-dependent manner
(Fig. 2C). Furthermore, the cell
viability in the miR-133a-3p mimics, puerarin and puerarin +
miR-133a-3p mimics groups was significantly decreased compared with
the NC group (Fig. 2D;
P<0.01).
 | Figure 2.Puerarin promotes cell apoptosis and
inhibits cell viability of breast cancer. (A) HCC38 cell apoptotic
rate following treatment with 20 µM puerarin, miR-133a-3p mimics,
20 µM puerarin + miR-133a-3p mimics and NC group detected by a flow
cytometry assay. *P<0.05, **P<0.01 vs. NC. (B) Apoptosis rate
expressed as a percentage. (C) Cell viability following the
treatment with puerarin at various concentrations at 24, 48 and 72
h, detected by a CCK-8 assay. *P<0.05, **P<0.01 vs. 24 h. (D)
Cell viability detected using a CCK-8 assay in four treatment
groups. **P<0.01 vs. NC. NC, negative control; CCK-8, Cell
Counting Kit-8; miR, microRNA; PI, propidium iodide; FITC,
fluorescein isothiocyanate. |
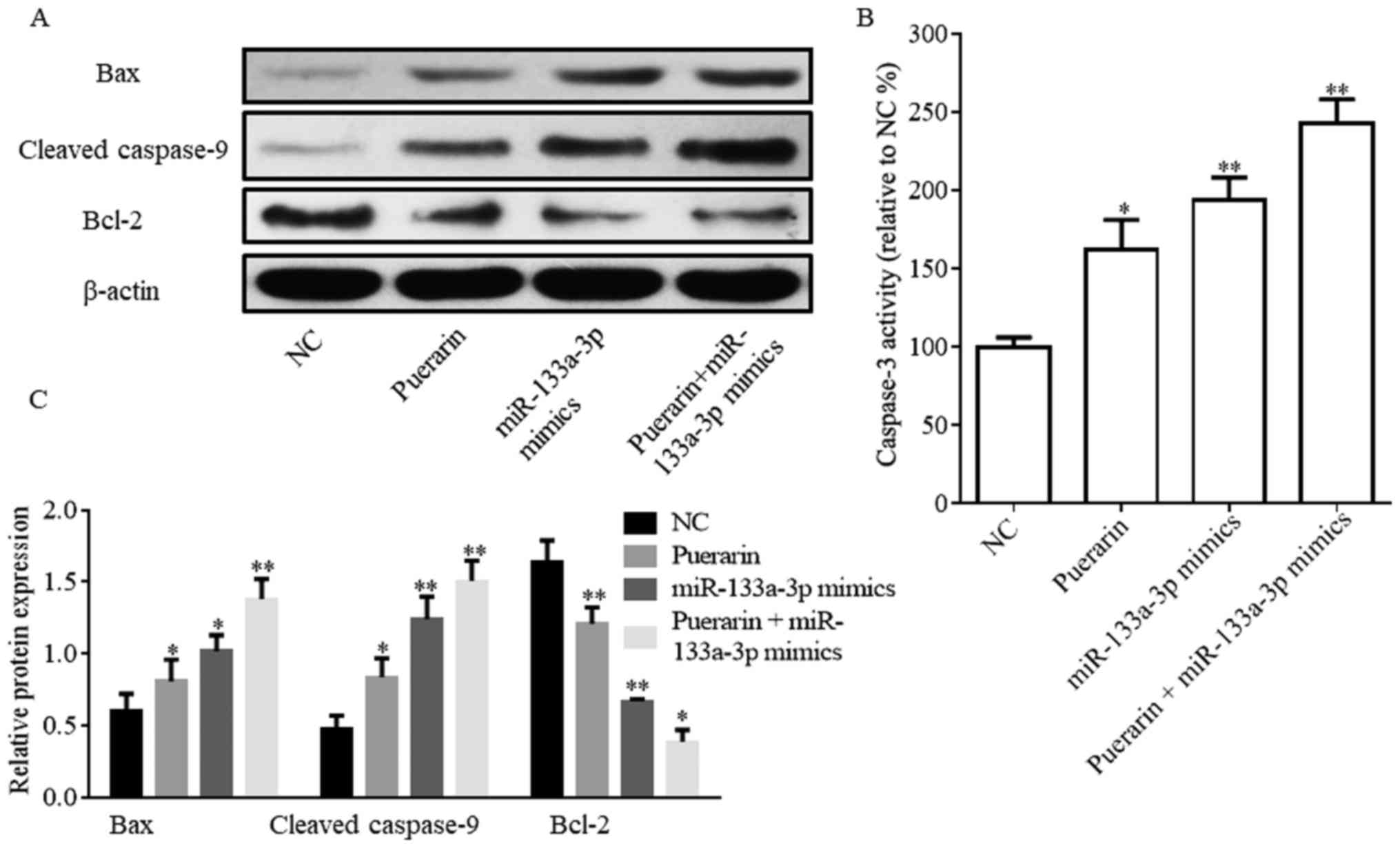
Protein expression levels of
apoptosis-associated proteins are regulated by puerarin and
miR-133a-3p mimics
The flow cytometry results suggested that puerarin
and miR-133a-3p mimics promoted HCC38 cell apoptosis. Therefore,
the expression levels of proteins associated with apoptosis and
caspase-3 activity were investigated (Fig. 3). The western blotting assay
suggested that Bax and cleaved caspase-9 expression levels were
significantly increased in the puerarin (20 µM), miR-133a-3p mimics
and puerarin (20 µM) + miR-133a-3p mimics groups compared with the
NC group; however, Bcl-2 expression exhibited the opposite trend
(Fig. 3A and B). Caspase-3
activity was significantly increased following the treatment with
puerarin and/or miR-133a-3p mimics (Fig. 3C).
DUSP1 is a direct target of
miR-133a-3p
A luciferase reporter gene assay was performed to
test whether DUSP1 was a direct target gene of miR-133a-3p.
Complementary binding was identified in the sequences of
miR-133a-3p and the DUSP1 3′UTR (Fig.
4A). The pcDNA 3.1-Luciferase vector was synthesized and
subsequently DUSP1 3′UTR-WT or MT sequences were inserted into the
plasmid. The present results demonstrated that the luciferase
activity of DUSP1 3′UTR-WT co-transfected with the miR-133a-3p
inhibitor was significantly decreased compared with the NC. In
addition, no significant difference was detected between the NC and
DUSP1 3′UTR-MT co-transfected with miR-133a-3p inhibitor (Fig. 4B). As miR-133a is positively
correlated with DUSP1 expression, inhibitor was used to perform
this experiment (Fig. 4C). The
expression of DUSP1 protein is also regulated by miR-133a-3p
(Fig. 4D). These present results
suggested that the sequence of DUSP1 3′UTR was complementary to the
miR-133A-3p sequence and that this site may be a direct target of
miR-133a-3p.
Puerarin and miR-133a-3p regulate the
DUSP1/p38 mitogen-activated protein kinase (MAPK) pathway
The western blotting assay suggested that puerarin
promoted the protein expression level of DUSP1 in a dose-dependent
manner (Fig. 5A). HCC38 cells were
transfected with the miR-133a-3p inhibitor and were subsequently
analyzed to examine the association between miR-133a-3p and DUSP1
(Fig. 5C and D). The DUSP1 mRNA
expression level was increased following treatment with puerarin;
however, miR-133a-3p inhibitor led to the opposite effect (Fig. 5B). The protein expression levels of
DUSP1 and MAPK were analyzed by western blotting. The present study
demonstrated that puerarin significantly increased the DUSP1 and
p38 protein expression levels, and decreased the p-p38 expression
level (Fig. 5C and D; P<0.05).
The miR-133a-3p inhibitor attenuated the effect of puerarin
(Fig. 5C and D). These present
results suggested that DUSP1 may serve tumor suppressive roles via
the MAPK pathway.
Discussion
Breast cancer is the most common invasive cancer
among women worldwide, accounting for 16% of all types of cancer
and 22.9% of the invasive types of cancer in women (19). Numerous previous studies
investigated the markers involved in breast cancer cell growth and
differentiation, which are essential processes for the formation
and development of breast cancer (20,21).
Cancer metastasis is the principal cause of the poor prognosis and
high mortality of breast cancer (22). At present, the treatment of breast
cancer is based on surgery and chemotherapy; however, the emergence
of drug resistance limits the efficacy of chemotherapy. The
efficacy of traditional Chinese medicine in cancer has been
investigated, and numerous Chinese herbs have been demonstrated to
inhibit cancer (23). The
combination of berberine and evodiamine suppressed the
proliferation of MCF-7 cells by inducing cell cycle arrest and
apoptosis, suggesting potential synergistic and combinatorial
applications of traditional Chinese medicine treatments (24). A previous study, including a total
of 1,853 patients with aggressive breast cancer, demonstrated that
Chinese herbal products contributed to a decrease in the risk of
invasive breast cancer (25). A
previous study suggested that puerarin regulated the expression
level of proteins involved in the mammalian target of
rapamycin/ribosomal protein S6 kinase β-1 signaling pathway, and
suggested that puerarin may represent a novel drug for the
treatment of bladder cancer (26).
The present study examined whether puerarin had an anticancer
effect on breast cancer and investigated its underlying mechanism
of action.
At present, anticancer therapy involving miRNAs is a
relatively novel field, and miRNA activity and function are
principal topics of investigation. Generally, miRNAs may serve as
oncogenes or tumor suppressor genes by regulating the target mRNA
or protein. SENP1 is one of the targets of miR-133a-3p, and the
miR-133a-3p/SENP1 axis was demonstrated to regulate cell
proliferation and the cell cycle in colorectal cancer (10). In addition, miR-133a-3p has been
identified as a tumor suppressor gene in breast cancer partly by
targeting LIM and SH3 domain protein 1 (27). The present study suggested, using a
bioinformatics analysis, that the DUSP1 gene may be one of the
targets of miR-133a-3p. Therefore, the present study suggested that
DUSP1, a gene involved in a number of tumors (28,29),
was part of the same pathway of miR-133a-3p in regulating
tumorigenesis. Furthermore, a previous study focused on the
development of antitumor drugs derived from miRNAs with potential
clinical implications (30). A
previous study demonstrated that pterostilbene decreased the
miR-19a expression level, affecting the phosphatase and tensin
homolog/RAC-α serine/threonine-protein kinase pathway, inhibiting
cell viability, cell cycle arrest and apoptosis, and promoting cell
invasion in hepatocellular carcinoma cells (31). In the present study, the effect of
puerarin on miR-133a-3p was investigated in breast cancer HCC38
cells. Furthermore, miR-133a-3p was suggested to serve a tumor
suppressive role by directly targeting DUSP1. Additionally, the
proteins downstream of DUSP1 were investigated.
As puerarin has the potential to suppress the growth
of tumor cells, research has focused on understanding the
biological activity of puerarin (32). Furthermore, the mechanism of
puerarin as an anticancer drug may involve the induction of tumor
suppressor genes or the inhibition of oncogenes. The present
results suggested that the viability of breast cancer cells HCC38
was suppressed by puerarin. Additionally, apoptosis of HCC38 was
promoted, and the present findings provide basis for further
investigating the effect of puerarin on breast cancer. In the
present study, puerarin promoted the expression of tumor suppressor
genes miR-133a-3p and DUSP1, suggesting that puerarin may affect
this pathway to serve antitumor effects. Furthermore, according to
the bioinformatics analysis, it was identified that miR-133a-3p and
DUSP1 genes interacted, based on the nucleotide sequence. The
expression of the DUSP1 gene was promoted by the upregulation of
miR-133a-3p. Xiao et al (33) demonstrated that miRNAs may promote
gene expression by targeting enhancers. DUSPs are a family of
proteins that function as negative regulators of MAPK activity in
mammalian cells (34). A previous
study suggested that silencing DUSP1 promoted the release of
pro-inflammatory cytokines by activating the MAPK signaling pathway
(35). The present study
identified that upregulation of DUSP1 expression led to an increase
of the p-p38 MAPK protein expression level.
Collectively, the present study identified that
puerarin decreased HCC38 cell viability and promoted apoptosis. The
mechanism underlying puerarin involved the upregulation of
miR-133a-3p expression. Notably, miR-133a-3p upregulation increased
the DUSP1 expression level, causing a decrease in the protein
expression level of p-p38. The impact of the downstream genes of
the MAPK pathway has not been investigated, representing a
limitation of the present study, and additional experiments are
required to address this aspect. The present results investigated
the molecular mechanisms of puerarin, a drug that may be used in
the future for improving the quality of life of the patients with
breast cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by The Nature and
Science Foundation of Jiangsu Province (China; grant no.
2016032A).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JC designed the present study. ZL, XR and JX
performed the experiments and wrote the paper. WX performed the
statistical analysis. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chia KS, Reilly M, Tan CS, Lee J, Pawitan
Y, Adami HO, Hall P and Mow B: Profound changes in breast cancer
incidence may reflect changes into a Westernized lifestyle: A
comparative population-based study in Singapore and Sweden. Int J
Cancer. 113:302–306. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Leung GM, Thach TQ, Lam TH, Hedley AJ, Foo
W, Fielding R, Yip PS, Lau EM and Wong CM: Trends in breast cancer
incidence in Hong Kong between 1973 and 1999: An age-period-cohort
analysis. Br J Cancer. 87:982–988. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sung H, Ren J, Li J, Pfeiffer RM, Wang Y,
Guida JL, Fang Y, Shi J, Zhang K, Li N, et al: Breast cancer risk
factors and mammographic density among high-risk women in urban
China. NPJ Breast Cancer. 4:32018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moss EG: MicroRNAs: Hidden in the genome.
Curr Biol. 12:R138–R140. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lv C, Li F, Li X, Tian Y, Zhang Y, Sheng
X, Song Y, Meng Q, Yuan S, Luan L, et al: miR-31 promotes mammary
stem cell expansion and breast tumorigenesis by suppressing Wnt
signaling antagonists. Nat Commun. 8:10362017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kojima S, Chiyomaru T, Kawakami K, Yoshino
H, Enokida H, Nohata N, Fuse M, Ichikawa T, Naya Y, Nakagawa M and
Seki N: Tumour suppressors miR-1 and miR-133a target the oncogenic
function of purine nucleoside phosphorylase (PNP) in prostate
cancer. Br J Cancer. 106:405–413. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dong Y, Zhao J, Wu CW, Zhang L, Liu X,
Kang W, Leung WW, Zhang N, Chan FK, Sung JJ, et al: Tumor
suppressor functions of miR-133a in colorectal cancer. Mol Cancer
Res. 11:1051–1060. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guo J, Xia B, Meng F and Lou G: miR-133a
suppresses ovarian cancer cell proliferation by directly targeting
insulin-like growth factor 1 receptor. Tumour Biol. 35:1557–1564.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gao L, Li SH, Tian YX, Zhu QQ, Chen G,
Pang YY and Hu XH: Role of downregulated miR-133a-3p expression in
bladder cancer: A bioinformatics study. OncoTargets Ther.
10:3667–3683. 2017. View Article : Google Scholar
|
|
10
|
Zhou GQ, Han F, Shi ZL, Yu L, Li XF, Yu C,
Shen CL, Wan DW, Zhu XG, Li R and He SB: miR-133a-3p targets
SUMO-specific protease 1 to inhibit cell proliferation and cell
cycle progress in colorectal cancer. Oncol Res. 26:795–800. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen CC, Hardy DB and Mendelson CR:
Progesterone receptor inhibits proliferation of human breast cancer
cells via induction of MAPK phosphatase 1 (MKP-1/DUSP1). J Biol
Chem. 286:43091–43102. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou YX, Zhang H and Peng C: Puerarin: A
review of pharmacological effects. Phytother Res. 28:961–975. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu X, Li S, Li Y, Cheng B, Tan B and Wang
G: Puerarin inhibits proliferation and induces apoptosis by
upregulation of miR-16 in bladder cancer cell line T24. Oncol Res.
26:1227–1234. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zeng X, Feng Q, Zhao F, Sun C, Zhou T,
Yang J and Zhan X: Puerarin inhibits TRPM3/miR-204 to promote
MC3T3-E1 cells proliferation, differentiation and mineralization.
Phytother Res. 32:996–1003. 2018. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yin M, Yuan Y, Cui Y, Hong X, Luo H, Hu X,
Tang M, Hescheler J and Xi J: Puerarin suppresses the self-renewal
of murine embryonic stem cells by inhibition of REST-miR-21
regulatory pathway. Cell Physiol Biochem. 37:527–536. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang WG, Liu XF, Meng KW and Hu SY:
Puerarin inhibits growth and induces apoptosis in SMMC-7721
hepatocellular carcinoma cells. Mol Med Rep. 10:2752–2758. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kang H, Zhang J, Wang B, Liu M, Zhao J,
Yang M and Li Y: Puerarin inhibits M2 polarization and metastasis
of tumor-associated macrophages from NSCLC xenograft model via
inactivating MEK/ERK 1/2 pathway. Int J Oncol. 50:545–554. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Donepudi MS, Kondapalli K, Amos SJ and
Venkanteshan P: Breast cancer statistics and markers. J Cancer Res
Ther. 10:506–511. 2014.PubMed/NCBI
|
|
20
|
Ciriello G, Gatza ML, Beck AH, Wilkerson
MD, Rhie SK, Pastore A, Zhang H, McLellan M, Yau C, Kandoth C, et
al: Comprehensive molecular portraits of invasive lobular breast
cancer. Cell. 163:506–519. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cancer Genome Atlas Network: Comprehensive
molecular portraits of human breast tumours. Nature. 490:61–70.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alkabban FM and Ferguson T: Cancer,
Breast. StatPearls Publishing; Treasure Island, FL: 2018
|
|
23
|
Xu SY, Huang X and Cheong KL: Recent
advances in marine algae polysaccharides: Isolation, structure, and
activities. Mar Drugs. 15:E3882017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Du J, Sun Y, Lu YY, Lau E, Zhao M, Zhou QM
and Su SB: Berberine and evodiamine act synergistically against
human breast cancer MCF-7 cells by inducing cell cycle arrest and
apoptosis. Anticancer Res. 37:6141–6151. 2017.PubMed/NCBI
|
|
25
|
Tsai YT, Lai JN, Lo PC, Chen CN and Lin
JG: Prescription of Chinese herbal products is associated with a
decreased risk of invasive breast cancer. Medicine (Baltimore).
96:e79182017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jiang K, Chen H, Tang K, Guan W, Zhou H,
Guo X, Chen Z, Ye Z and Xu H: Puerarin inhibits bladder cancer cell
proliferation through the mTOR/p70S6K signaling pathway. Oncol
Lett. 15:167–174. 2018.PubMed/NCBI
|
|
27
|
Sui Y, Zhang X, Yang H, Wei W and Wang M:
MicroRNA-133a acts as a tumour suppressor in breast cancer through
targeting LASP1. Oncol Rep. 39:473–482. 2018.PubMed/NCBI
|
|
28
|
Kesarwani M, Kincaid Z, Gomaa A, Huber E,
Rohrabaugh S, Siddiqui Z, Bouso MF, Latif T, Xu M, Komurov K, et
al: Targeting c-FOS and DUSP1 abrogates intrinsic resistance to
tyrosine-kinase inhibitor therapy in BCR-ABL-induced leukemia. Nat
Med. 23:472–482. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nettersheim D, Jostes S, Fabry M, Honecker
F, Schumacher V, Kirfel J, Kristiansen G and Schorle H: A signaling
cascade including ARID1A, GADD45B and DUSP1 induces apoptosis and
affects the cell cycle of germ cell cancers after romidepsin
treatment. Oncotarget. 7:74931–74946. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rukov JL and Shomron N: MicroRNA
pharmacogenomics: Post-transcriptional regulation of drug response.
Trends Mol Med. 17:412–423. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qian YY, Liu ZS, Zhang Z, Levenson AS and
Li K: Pterostilbene increases PTEN expression through the targeted
downregulation of microRNA-19a in hepatocellular carcinoma. Mol Med
Rep. 17:5193–5201. 2018.PubMed/NCBI
|
|
32
|
Wei HY, Zhang YJ and Zhao SZ: Puerarin
regulates neovascular glaucoma through pigment epitheliumderived
growth factorinduced NFκB signaling pathway. Mol Med Rep.
17:7866–7874. 2018.PubMed/NCBI
|
|
33
|
Xiao M, Li J, Li W, Wang Y, Wu F, Xi Y,
Zhang L, Ding C, Luo H, Li Y, et al: MicroRNAs activate gene
transcription epigenetically as an enhancer trigger. RNA Biol.
14:1326–1334. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Low HB and Zhang Y: Regulatory roles of
MAPK phosphatases in cancer. Immune Netw. 16:85–98. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang B, Li SL, Xie HL, Fan JW, Gu CW,
Kang C and Teng MJ: Effects of silencing the DUSP1 gene using
lentiviral vector-mediated siRNA on the release of proinflammatory
cytokines through regulation of the MAPK signaling pathway in mice
with acute pancreatitis. Int J Mol Med. 41:2213–2224.
2018.PubMed/NCBI
|