Introduction
Osteoporosis is a bone metabolic disorder,
characterized by low bone mineral density (BMD) and
microarchitectural deterioration with increased bone fragility and
subsequent susceptibility to fractures (1). It has been reported that osteoporosis
is induced by an imbalance between bone resorption by osteoclasts
and bone deposition by osteoblasts (2). Postmenopausal osteoporosis (PMOP) is
a major public health concern worldwide that frequently presents in
postmenopausal women due to the estrogen deficiency and continuous
calcium loss that occurs with aging (3). A proactive approach that identifies
patients at high risk of developing PMOP is recommended to prevent
bone loss (4).
With the advancement of high-throughput
technologies, gene microarray analysis has become an effective
method for identifying differentially expressed genes (DEGs) and,
therefore, potential biomarkers in various diseases. Multiple
studies (5–7) have utilized gene microarray analysis
to identify key genes in the pathogenesis of PMOP. Integrated
multiple gene microarray analysis may contribute to the
identification of more accurate gene biomarkers.
The present study aimed to develop accurate
biomarkers and provide clues for exploring the underlying mechanism
of PMOP. By integrating multiple microarray analysis in this
present study, DEGs between PMOP patients and normal controls were
identified. Based on these DEGs, the optimal gene combination with
the greatest diagnostic value for PMOP was determined. Functional
annotation and protein-protein interaction (PPI) network
constructions were also performed to explore the biological
functions of DEGs. These findings will help elucidate the mechanism
underlying PMOP development and uncover novel diagnostic
biomarkers.
Materials and methods
Microarray expression profiling
Microarray datasets of PMOP and normal controls were
downloaded from the Gene Expression Omnibus (GEO) database
(http://www.ncbi.nlm.nih.gov/geo). All
datasets that contained whole-genome mRNA expression profiles
between PMOP patient and control blood samples were enrolled in the
current study. The datasets were scale normalized.
Identification of DEGs between PMOP
patients and normal controls
MetaMA is an R package-implementing meta-analysis
approach for microarray data (8).
Data from multiple microarray analyses were combined by metaMA
(inverse normal method), and individual P-values were obtained. In
the integrated analysis performed in the present study, DEGs
between PMOP patients and normal controls were identified at a
P-value of <0.05. Hierarchical clustering analyses of mRNAs were
conducted with ‘pheatmap’ package in R (version 3.3.3; www.r-project.org).
Identification of optimal diagnostic
gene biomarkers for PMOP
The LASSO algorithm was applied with the glmnet
package (https://cran.r-project.org/web/packages/glmnet/) in
order to reduce the dimensions of the data (9). The Boruta algorithm (https://cran.r-project.org/web/packages/Boruta/)
employs a wrapper approach, built around a random forest classifier
(10). This algorithm is used to
compare the relevance of the features to those of the random probes
(11). The scale-standardized
datasets were merged, the DEGs between PMOP patients and normal
controls were retained for feature selection, and gene biomarkers
for PMOP were identified with the LASSO and Boruta algorithms.
Furthermore, the optimal gene biomarkers for PMOP were identified
by overlapping biomarkers derived from these two algorithms.
Hierarchical clustering analysis of these shared gene biomarkers,
obtained by LASSO and Boruta algorithms, was conducted with the R
package ‘pheatmap’ (R version 3.3.3).
Based on these optimal gene biomarkers, several
classification models, including support vector machine (SVM),
decision tree and random forest models, were established to further
identify the diagnostic value of these biomarkers in PMOP. An SVM
model was established with an ‘e1071’ package (https://cran.r-project.org/web/packages/e1071/index.html).
A decision tree model was established with the ‘rpart’ package
(https://cran.r-project.org/web/packages/rpart/). A
random forest model was established with the ‘randomForests’
package (https://cran.r-project.org/web/packages/randomForest/).
These three classification models were compared by the average
misjudgment rates of their 10-fold cross validations. The
diagnostic ability of the three classification models was assessed
by calculating the receiver operating characteristic curve, and
measuring the area under the curve (AUC), accuracy, sensitivity and
specificity.
PMOP-specific PPI network
To further investigate the biological functions of
these optimal gene biomarkers, a PPI network was constructed with
the BioGRID (also known as Biological General Repository for
Interaction Datasets; http://thebiogrid.org/) and Cytoscape (http://www.cytoscape.org). Nodes and edges in the PPI
network represented the proteins and the interactions between two
proteins, respectively.
Functional annotation
Based on the PMOP-specific PPI network, the proteins
that integrated with proteins encoded by the optimal gene
biomarkers were identified. Gene Ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway enrichment
analyses of all gene biomarkers and other DEGs that encode proteins
within the PMOP-specific PPI network were conducted with the online
software GeneCodis (http://genecodis.cnb.csic.es/analysis). Differences
(using the Benjamini and Hochberg method) were defined as
statistically significant when the false discovery rate (FDR) was
<0.05.
Results
DEGs between PMOP patients and normal
controls
Three datasets, including GSE56815, GSE13850 and
GSE7429, were downloaded from the GEO database [(Table I) (12)]. Based on these three datasets,
1,320 DEGs (710 upregulated DEGs and 613 downregulated DEGs) with
FDR<0.05 were identified between PMOP patients and normal
controls. Hierarchical clustering analysis of the top 100 DEGs
between PMOP patients and normal controls is presented in Fig. 1.
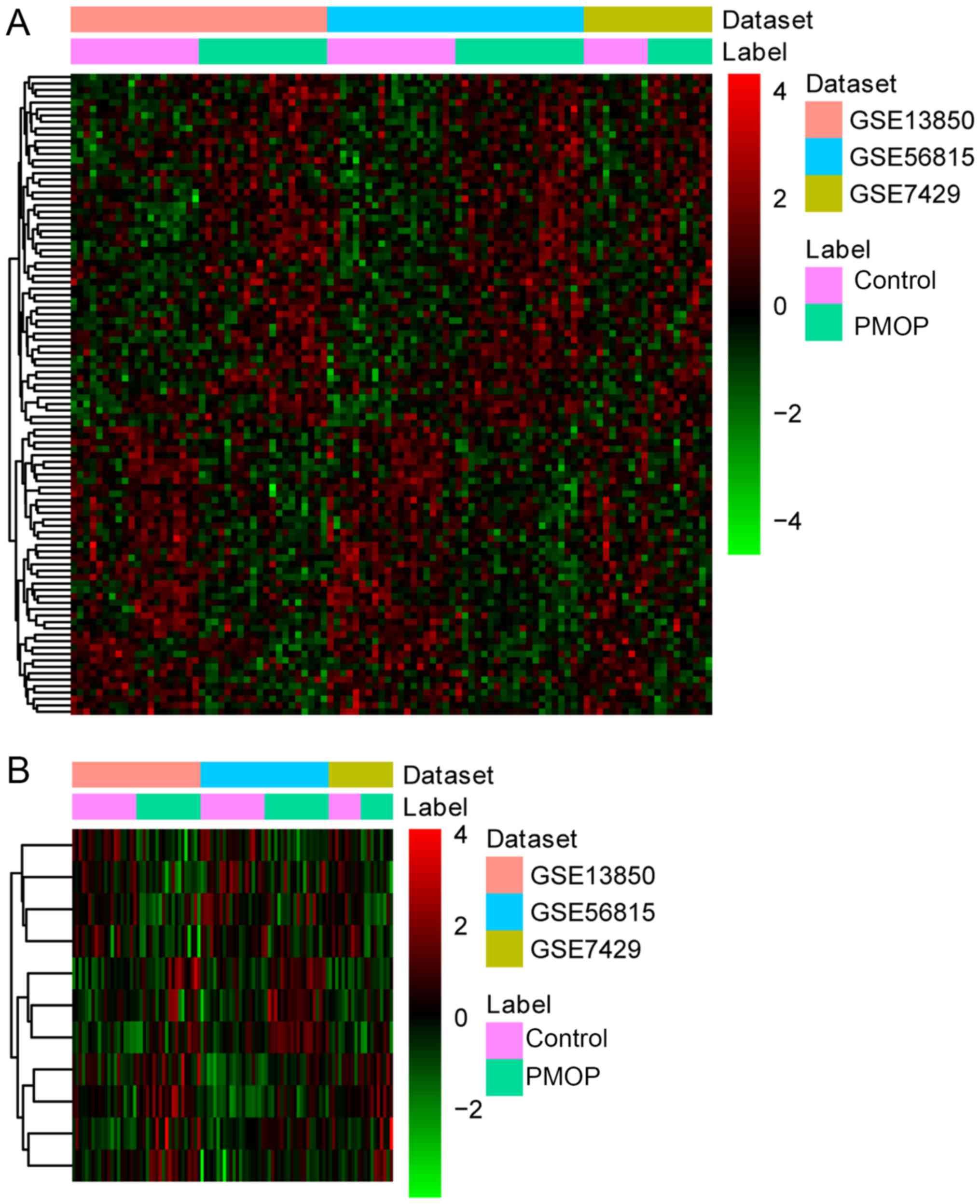 | Figure 1.Hierarchical clustering analysis of
DEGs between PMOP patients and normal controls. (A) Analysis of top
100 DEGs between PMOP patients and normal controls. (B) Analysis of
11 shared gene biomarkers for PMOP obtained by both the LASSO and
Boruta algorithms. Rows and columns represent DEGs and samples,
respectively. The color scale represents the expression levels.
DEGs, differentially expressed genes; PMOP, postmenopausal
osteoporosis. BMD, bone mineral density; DEGs, ESR1, estrogen
receptor 1; GEO, Gene Expression Omnibus; GO, Gene Ontology; KEGG,
Kyoto Encyclopedia of Genes and Genomes; NE, norepinephrine; PPI,
protein-protein interaction; RANKL, receptor activator of NF-κB
ligand; SVM, support vector machine. |
 | Table I.Datasets used in the present
study. |
Table I.
Datasets used in the present
study.
| GEO ID | Sample | Country | Year | First author | PMOP to control
ratio |
|---|
| GSE56815 | Blood | USA | 2016 | Liu | 20:20 |
| GSE13850 | Blood | USA | 2009 | Xiao | 20:20 |
| GSE7429 | Blood | USA | 2008 | Xiao | 10:10 |
Identification of optimal diagnostic
gene biomarkers for PMOP
A total of 31 and 32 gene biomarkers were identified
with the LASSO and Boruta algorithms, respectively. Furthermore, 11
shared mRNA biomarkers for PMOP were identified by overlapping the
biomarkers derived from these two algorithms (Table II and Fig. 2). These 11 mRNA biomarkers included
dehydrogenase E1 and transketolase domain containing 1 (DHTKD1),
osteoclast stimulating factor 1 (OSTF1), G protein-coupled receptor
87 (GPR87), GPR116 (also known as adhesion G protein-coupled
receptor F5), BCL2 interacting killer (BIK), adrenoceptor β1
(ADRB1), neogenin 1 (NEO1), RB binding protein 4 (RBBP4), cylicin 2
(CYLC2), EF-hand calcium binding domain 1 (EFCAB1) and DEAH-box
helicase 35 (DHX35). Hierarchical clustering analysis of these 11
mRNA biomarkers was performed.
 | Figure 2.Expression levels of 11 gene
biomarkers in the blood samples of postmenopausal osteoporosis
patients and normal controls. Gene expression levels of (A) EFCAB1,
(B) DHTKD1, (C) OSTF1, (D) GPR116, (E) BIK, (F) ADRB1, (G) NEO1,
(H) RBBP4, (I) GPR87, (J) CYLC2 and (K) DHX35 are shown.
***P<0.001 vs. the normal control group. EFCAB1, EF-hand calcium
binding domain 1; DHTKD1, dehydrogenase E1 and transketolase domain
containing 1; OSTF1, osteoclast stimulating factor 1; GPR, G
protein-coupled receptor; BIK, BCL2 interacting killer; ADRB1,
adrenoceptor β1; NEO1, neogenin 1; RBBP4, RB binding protein 4;
CYLC2, cylicin 2; DHX35, DEAH-box helicase 35. |
| Table II.A total of 11 shared gene biomarkers
for postmenopausal osteoporosis, obtained using the LASSO and
Boruta algorithms. |
Table II.
A total of 11 shared gene biomarkers
for postmenopausal osteoporosis, obtained using the LASSO and
Boruta algorithms.
| Gene ID | Gene symbol | Combined ES | P-value | FDR | Regulation |
|---|
| 79645 | EFCAB1 | −1.16 |
4.51×10−8 |
2.80×10−4 | Down |
| 55526 | DHTKD1 | 1.07 |
4.61×10−7 |
1.45×10−3 | Up |
| 26578 | OSTF1 | 1.05 |
1.99×10−6 |
3.38×10−3 | Up |
| 221395 | GPR116 | 0.98 |
2.58×10−6 |
3.38×10−3 | Up |
| 638 | BIK | 0.92 |
9.30×10−6 |
8.69×10−3 | Up |
| 153 | ADRB1 | 0.89 |
1.97×10−5 |
1.27×10−2 | Up |
| 4756 | NEO1 | −0.85 |
5.61×10−5 |
2.09×10−2 | Down |
| 5928 | RBBP4 | 0.81 |
7.70×10−5 |
2.32×10−2 | Up |
| 53836 | GPR87 | −0.80 |
1.20×10−4 |
2.61×10−2 | Down |
| 1539 | CYLC2 | −0.74 |
3.42×10−4 |
4.12×10−2 | Down |
| 60625 | DHX35 | 0.73 |
5.64×10−4 |
5.08×10−2 | Up |
The SVM, decision tree and random forest models were
established with these 11 mRNA biomarkers, and the accuracy of
these three models was 93, 78 and 94%, respectively. The AUC of
SVM, decision tree and random forest models was 0.975, 0.799 and
0.975, respectively. In addition, the sensitivity and specificity
of the SVM model were 92 and 100%, respectively (Fig. 3A). The sensitivity and specificity
of the decision tree model were 70 and 88%, respectively (Fig. 3B). Finally, the sensitivity and
specificity of the random forest model were 90 and 100%,
respectively (Fig. 3C).
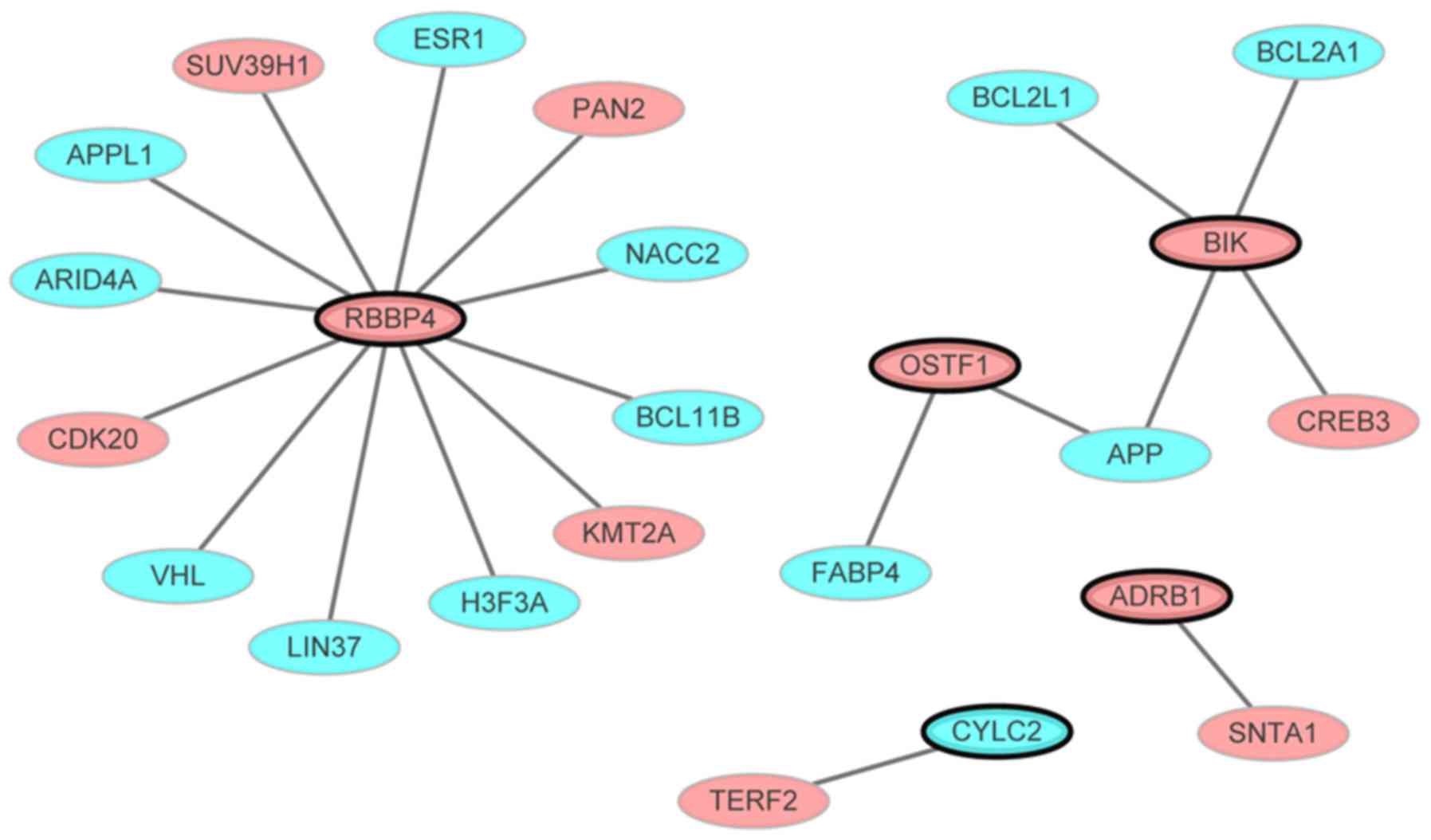
PMOP-specific PPI network
The PMOP-specific PPI network consisted of 24 nodes
and 20 edges (Fig. 4). Out of the
11 genes not all interacted with other differentially expressed
genes. Only the gene that interacted with other differentially
expressed genes (even though one gene) in the PPI network were
presented. Nodes and edges represented the proteins and the
interactions between two proteins, respectively. The red and blue
ellipses represented the proteins encoded by up- and downregulated
DEGs between PMOP patients and normal controls, respectively. RBBP4
(degree=12) was the hub gene of this PMOP-specific PPI network.
Functional annotation
Based on the functional annotations of these 11
biomarkers and DEGs that encode proteins of the PMOP-specific PPI
network, prostate epithelial cord elongation (FDR<0.05),
estrogen response element binding (FDR<0.05) and nucleosome
remodeling deacetylase complex (FDR<0.05) were the most
significant GO terms. Furthermore, endocrine and other
factor-regulated calcium reabsorption (FDR=2.23×10−5),
was a significantly enriched pathway in PMOP; Estrogen receptor 1
(ESR1) was revealed to be upregulated within this pathway (Table III).
| Table III.Top 10 significantly GO terms and
KEGG pathways in postmenopausal osteoporosis. |
Table III.
Top 10 significantly GO terms and
KEGG pathways in postmenopausal osteoporosis.
| A, GO terms |
|---|
|
|---|
| ID | Term | FDR | Genes |
|---|
| Biological
process |
|
GO:0060523 | Prostate epithelial
cord elongation | <0.05 | ESR1 |
|
GO:0031649 | Heat
generation | <0.05 | ADRB1 |
|
GO:0045986 | Negative regulation
of smooth muscle contraction | <0.05 | ADRB1 |
|
GO:0002025 | Vasodilation by
norepinephrine-epinephrine involved in regulation of systemic
arterial blood pressure | <0.05 | ADRB1 |
|
GO:0031077 | Post-embryonic
camera-type eye development | <0.05 | BCL11B |
|
GO:0048386 | Positive regulation
of retinoic acid receptor signaling pathway | <0.05 | ESR1 |
|
GO:0003382 | Epithelial cell
morphogenesis | <0.05 | BCL11B |
|
GO:0051124 | Synaptic growth at
neuromuscular junction | <0.05 | APP |
|
GO:0060750 | Epithelial cell
proliferation involved in mammary gland duct elongation | <0.05 | ESR1 |
|
GO:0046878 | Positive regulation
of saliva secretion | <0.05 | ADRB1 |
| Molecular
function |
|
GO:0034056 | Estrogen response
element binding | <0.05 | ESR1 |
|
GO:0051400 | BH domain
binding | <0.05 | BIK |
|
GO:0051380 | Norepinephrine
binding | <0.05 | ADRB1 |
|
GO:0004535 | Poly(A)-specific
ribonuclease activity | <0.05 | PAN2 |
|
GO:0051434 | BH3 domain
binding | <0.05 | BCL2L1 |
|
GO:0051425 | PTB domain
binding | <0.05 | APP |
|
GO:0004939 | β-adrenergic
receptor activity | <0.05 | ADRB1 |
|
GO:0004591 | Oxoglutarate
dehydrogenase (succinyl-transferring) activity | <0.05 | DHTKD1 |
|
GO:0004940 | β1-adrenergic
receptor activity | <0.05 | ADRB1 |
|
GO:0031798 | Type 1 metabotropic
glutamate receptor binding | <0.05 | ESR1 |
| Cellular
component |
|
GO:0016581 | NuRD complex | <0.05 | APPL1, RBBP4 |
|
GO:0097136 | Bcl-2 family
protein complex | <0.05 | BCL2L1 |
|
GO:0033150 | Cytoskeletal
calyx | <0.05 | CYLC2 |
|
GO:0033553 | rDNA
heterochromatin | <0.05 | SUV39H1 |
|
GO:0033186 | CAF-1 complex | <0.05 | RBBP4 |
|
GO:0044429 | Mitochondrial
part | <0.05 | BCL2L1, ESR1 |
|
GO:0030870 | Mre11 complex | <0.05 | TERF2 |
|
GO:0030891 | VCB complex | <0.05 | VHL |
|
GO:0001740 | Barr body | <0.05 | H3F3A |
|
GO:0016589 | NURF complex | <0.05 | RBBP4 |
|
| B, KEGG
pathways |
|
| ID | Pathway | FDR | Genes |
|
| KEGG:05200 | Pathways in
cancer | <0.05 | BCL2L1, VHL,
APPL1 |
| KEGG:04962 |
Vasopressin-regulated water
reabsorption | <0.05 | CREB3 |
| KEGG:04961 | Endocrine and other
factor-regulated calcium reabsorption | <0.05 | ESR1 |
| KEGG:00310 | Lysine
degradation | <0.05 | SUV39H1 |
| KEGG:05014 | Amyotrophic lateral
sclerosis (ALS) | <0.05 | BCL2L1 |
| KEGG:05210 | Colorectal
cancer | <0.05 | APPL1 |
| KEGG:03018 | RNA
degradation | <0.05 | PAN2 |
| KEGG:03320 | PPAR signaling
pathway | <0.05 | FABP4 |
| KEGG:05211 | Renal cell
carcinoma | <0.05 | VHL |
| KEGG:05212 | Pancreatic
cancer | <0.05 | BCL2L1 |
Discussion
PMOP increases the risk of fragility fractures in
postmenopausal women, and imposes a significant burden on patients'
families and society. Previous studies have indicated that
identification of patients at high risk of developing PMOP can
contribute to the prevention of bone loss (4,13).
In the present study, 1,320 DEGs between PMOP
patients and normal controls were identified with integrated
microarray analysis. The gene biomarkers for PMOP were further
identified with the LASSO and Boruta algorithms. An 11-gene
combination (EFCAB1, DHTKD1, OSTF1, GPR116, BIK, ADRB1, NEO1,
RBBP4, GPR87, CYLC2 and DHX35) was revealed as an optimal biomarker
for PMOP with feature selection and classification procedures using
SVM, decision tree and random forest models. Based on the random
forest model, the 11-gene combination achieved a 94% prediction
accuracy in distinguishing patients with PMOP from normal controls,
with 90% sensitivity and 100% specificity. The results obtained
using the other two models (SVM and decision tree) further
supported this finding.
Among these 11 genes, CYLC2 has previously been
shown to be upregulated in B cells from postmenopausal women with
low BMD compared with that in postmenopausal women with high BMD
(14). In addition, CYLC2 is
involved in the structural component of the cytoskeleton. A
previous study indicated that the structural component of the
cytoskeleton is associated with PMOP, which may suggest a potential
role of CYLC2 in PMOP (14).
Three DEGs identified in the present study, namely
OSTF1, ADRB1 and NEO1, have previously been reported to be
associated with the balance between bone formation and bone
resorption (15–19). OSTF1 is an intracellular protein
that is highly expressed in osteoclasts, which indirectly enhances
osteoclast formation and bone resorption (15). ADRB1 belongs to the family of
guanine nucleotide-binding regulatory protein-coupled receptors,
which regulate the physiological effects of the hormone epinephrine
and the neurotransmitter norepinephrine (NE) (16). The β-adrenergic system is also
involved in leptin-dependent central regulation of bone turnover
(17,18). Intraosseous sympathetic nerve
fibers can be activated and release NE via leptin stimulation
(17). Adrenergic receptors
expressed on osteoblasts bind to the released NE and result in
suppression of bone formation. In addition, β-adrenergic-stimulated
production of the receptor activator of nuclear factor (NF)-κB
ligand by osteoblasts may contribute to a negative bone mineral
balance (19). In the present
study, OSTF1 and ADRB1 were upregulated in the blood of patients
with PMOP compared with that of normal controls. Furthermore, NEO1
encodes a cell surface protein that belongs to the immunoglobulin
superfamily, and has been speculated to serve roles in cell growth
and differentiation and in cell-cell adhesion. A previous study
reported abnormal chondrocyte maturation and endochondral bone
growth in NEO1 knockout mice (20). Additionally, the association
between NEO1 and bone mass was identified by high-throughput
screening of mouse gene knockouts (21). To the best of our knowledge, the
present study is the first to reveal a downregulation of NEO1 in
the blood of patients with PMOP.
Estrogen deficiency is a pivotal cause of
postmenopausal bone loss (22).
RBBP4 is an estrogen-associated gene, which was included in the
11-gene combination described in the present study. It is also a
chromatin remodeling factor that encodes a ubiquitously expressed
nuclear protein that belongs to a highly conserved subfamily of
WD-repeat proteins (23). RBBP4
has been reported to be involved in the chromatin remodeling and
transcriptional repression associated with histone deacetylation
(24). An upregulation in the
expression of RBBP4 was detected in the tibia callus of
estrogen-deficient rats (25). To
the best of our knowledge, the present study is the first to reveal
an upregulation of RBBP4 in the blood of patients with PMOP.
Furthermore, as the hub protein of the PMOP-specific PPI network,
RBBP4 was integrated with ESR1, a well-known PMOP-associated gene,
which was revealed to be enriched in the endocrine and other
factor-regulated calcium reabsorption pathway (KEGG ID: 04961).
ESR1 is expressed on cells that contribute to bone formation, such
as osteoblasts, osteocytes and osteoclasts. It also increases the
formation and function of osteoblasts and reduces bone resorption
activities (26). Therefore, the
RBBP4-ESR1 interaction may serve a key role in PMOP.
To the best of our knowledge, no previous study has
reported the association between PMOP and the six other genes
described in the current study, including DHTKD1, GPR87, GPR116,
BIK, EFCAB1 and DHX35. DHTKD1 is a nuclear gene that is involved in
mitochondrial lysine metabolism and adenosine triphosphate
production (27,28). DHTKD1 has also been demonstrated to
link mitochondrial dysfunction and eosinophilic esophagitis
(29). Kim and Lee (30) indicated that mitochondrial
dysfunction may be a potential pathophysiological mechanism of
PMOP, which suggested that DHTKD1 may regulate mitochondrial
dysfunction in PMOP. In addition, GPR87 is a cell surface G
protein-coupled receptor that has been reported to be overexpressed
in various types of cancer (31,32),
and it serves a critical oncogenic role in pancreatic cancer
progression by activating the NF-κB signaling pathway (33). GPR116 is a member of the G
protein-coupled receptor family predominantly expressed in the
alveolar type II epithelial cells of the lung. Since NF-κB
signaling pathway is an important mediator in osteoblast
differentiation, it can be speculated that both GPR87 and GPR116
may serve a role in PMOP by regulating the NF-κB signaling pathway.
Another identified gene, BIK, is a member of the BH3-only Bcl-2
family of pro-apoptotic proteins, which is suppressed in various
types of cancer (34). Methylated
BIK was identified in the bone marrow of patients with multiple
myeloma, and dysregulated BIK expression was observed in
hematopoietic cell fractions of patients with myelodysplastic
syndrome, highlighting the importance of BIK in bone disease
(34,35). Furthermore, the gene DHX35 is a
putative RNA helicase, and its variants have been reported to be
involved with facial morphology, thyroid cancer and colorectal
cancer (36–38) Finally, DNA methylation of EFCAB1
was demonstrated to be involved in multi-organ carcinogenesis
(39). However, further research
is required to explore the roles of DHX35 and EFCAB1 in PMOP.
In conclusion, the present study identified 11 genes
that were significantly associated with PMOP and provided clues for
exploring the molecular mechanism of PMOP. Three of the identified
genes (OSTF1, ADRB1 and NEO1) were speculated to be involved in
PMOP by regulating the balance between bone formation and bone
resorption, while two genes (GPR87 and GPR116) may regulate the
NF-κB signaling pathway. RBBP4 and DHTKD1 may also be potential
regulators of PMOP via interacting with ESR1 and regulating
mitochondrial dysfunction, respectively. Furthermore, the
constituents of this 11-gene combination may serve as potential
biomarkers for PMOP. However, biological investigations and
validation with a larger sample size are lacking, and are
considered to be limitations of the present study. Further
investigations are required to validate the diagnostic abilities of
this gene combination for PMOP prior to its clinical
application.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CY and XS made substantial contributions to the
study conception and design. CY, JR, BL, CJ, CM, CC and YS
collected and analyzed the data. CY, JR, BL and CJ interpreted the
data. All authors were involved in drafting the manuscript and
provided final approval of this manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
BMD
|
bone mineral density
|
|
DEGs
|
differentially expressed genes
|
|
ESR1
|
estrogen receptor 1
|
|
GEO
|
Gene Expression Omnibus
|
|
GO
|
Gene Ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
NE
|
norepinephrine
|
|
PMOP
|
postmenopausal osteoporosis
|
|
PPI
|
protein-protein interaction
|
|
RANKL
|
receptor activator of NF-κB ligand
|
|
SVM
|
support vector machine
|
References
|
1
|
Kanis JA, McCloskey EV, Johansson H, Oden
A, Melton LJ III and Khaltaev N: A reference standard for the
description of osteoporosis. Bone. 42:467–475. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Paschalis EP, Gamsjaeger S, Hassler N,
Fahrleitner-Pammer A, Dobnig H, Stepan JJ, Pavo I, Eriksen EF and
Klaushofer K: Vitamin D and calcium supplementation for three years
in postmenopausal osteoporosis significantly alters bone mineral
and organic matrix quality. Bone. 95:41–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Meng J, Zhang D, Pan N, Sun N, Wang Q, Fan
J, Zhou P, Zhu W and Jiang L: Identification of mir-194-5p as a
potential biomarker for postmenopausal osteoporosis. PeerJ.
3:e9712015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sanders S and Geraci SA: Osteoporosis in
postmenopausal women: Considerations in prevention and treatment:
(women's health series). Southern Med J. 106:698–706. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yan B, Li J and Zhang L: Identification of
B cells participated in the mechanism of postmenopausal women
osteoporosis using microarray analysis. Int J Clin Exp Med.
8:1027–1034. 2015.PubMed/NCBI
|
|
6
|
Liu Y, Wang Y, Yang N, Wu S, Lv Y and Xu
L: In silico analysis of the molecular mechanism of postmenopausal
osteoporosis. Mol Med Rep. 12:6584–6590. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ma M, Luo S, Zhou W, Lu L, Cai J, Yuan F
and Yin F: Bioinformatics analysis of gene expression profiles in B
cells of postmenopausal osteoporosis patients. Taiwan J Obstet
Gynecol. 56:165–170. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Marot G, Foulley JL, Mayer CD and
Jaffrézic F: Moderated effect size and P-value combinations for
microarray meta-analyses. Bioinformatics. 25:2692–2699. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Friedman J, Hastie T and Tibshirani R:
Regularization paths for generalized linear models via coordinate
descent. J Stat Softw. 33:1–22. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Breiman L: Random forests. Machine
Learning. 45:5–32. 2001. View Article : Google Scholar
|
|
11
|
Stoppiglia H, Dreyfus G, Dubois R and
Oussar Y: Ranking a random feature for variable and feature
selection. J Mach Learn Res. 3:1399–1414. 2003.
|
|
12
|
Xiao P, Chen Y, Jiang H, Liu YZ, Pan F,
Yang TL, Tang ZH, Larsen JA, Lappe JM, Recker RR and Deng HW: In
vivo genome-wide expression study on human circulating B cells
suggests a novel esr1 and mapk3 network for postmenopausal
osteoporosis. J Bone Miner Res. 23:644–654. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tella SH and Gallagher JC: Prevention and
treatment of postmenopausal osteoporosis. J Steroid Biochem Mol
Biol. 155–170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma M, Chen X, Lu L, Yuan F, Zeng W, Luo S,
Yin F and Cai J: Identification of crucial genes related to
postmenopausal osteoporosis using gene expression profiling. Aging
Clin Exp Res. 28:1067–1074. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vermeren M, Lyraki R, Wani S, Airik R,
Albagha O, Mort R, Hildebrandt F and Hurd T: Osteoclast stimulation
factor 1 (ostf1) knockout increases trabecular bone mass in mice.
Mamm Genome. 28:498–514. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang Y, Lin Y, Zhao H, Guo Q, Yan C and
Lin N: Revealing the effects of the herbal pair of euphorbia kansui
and glycyrrhiza on hepatocellular carcinoma ascites with
integrating network target analysis and experimental validation.
Int J Biol Sci. 12:594–606. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Takeda S, Elefteriou F, Levasseur R, Liu
X, Zhao L, Parker KL, Armstrong D, Ducy P and Karsenty G: Leptin
regulates bone formation via the sympathetic nervous system. Cell.
111:305–317. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Karsenty G: Leptin controls bone formation
through a hypothalamic relay. Recent Prog Horm Res. 56:401–415.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Elefteriou F, Ahn JD, Takeda S, Starbuck
M, Yang X, Liu X, Kondo H, Richards WG, Bannon TW, Noda M, et al:
Leptin regulation of bone resorption by the sympathetic nervous
system and cart. Nature. 434:514–520. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou Z, Xie J, Lee D, Liu Y, Jung J, Zhou
L, Xiong S, Mei L and Xiong WC: Neogenin regulation of bmp-induced
canonical smad signaling and endochondral bone formation. Dev cell.
19:90–102. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Brommage R, Liu J, Hansen GM, Kirkpatrick
LL, Potter DG, Sands AT, Zambrowicz B, Powell DR and Vogel P:
High-throughput screening of mouse gene knockouts identifies
established and novel skeletal phenotypes. Bone Res. 2:140342014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sharma D, Larriera AI, Palacio-Mancheno
PE, Gatti V, Fritton JC, Bromage TG, Cardoso L, Doty SB and Fritton
SP: The effects of estrogen deficiency on cortical bone
microporosity and mineralization. Bone. 110:1–10. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang X, Yao F, Liang X, Zhu X, Zheng R,
Jia B, Hou L and Zou X: Cloning and expression of
retinoblastoma-binding protein 4 gene in embryo diapause
termination and in response to salinity stress from brine shrimp
artemia sinica. Gene. 591:351–361. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Balboula AZ, Stein P, Schultz RM and
Schindler K: Rbbp4 regulates histone deacetylation and bipolar
spindle assembly during oocyte maturation in the mouse. Biol
Reprod. 92:1052015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Saul D, Ninkovic M, Komrakova M, Wolff L,
Simka P, Gasimov T, Menger B, Hoffmann DB, Rohde V and Sehmisch S:
Effect of zileuton on osteoporotic bone and its healing, expression
of bone, and brain genes in rats. J Appl Physiol. 124:118–130.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shalan NA, Mustapha NM and Mohamed S: Noni
leaf and black tea enhance bone regeneration in estrogen-deficient
rats. Nutrition. 33:42–51. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu W, Zhu H, Gu M, Luo Q, Ding J, Yao Y,
Chen F and Wang Z: Dhtkd1 is essential for mitochondrial biogenesis
and function maintenance. FEBS Lett. 587:3587–3592. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Danhauser K, Sauer S, Haack TB, Wieland T,
Staufner C, Graf E, Zschocke J, Strom TM, Traub T, Okun JG, et al:
Dhtkd1 mutations cause 2-aminoadipic and 2-oxoadipic aciduria. Am J
Hum Genet. 91:1082–1087. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sherrill JD, Kc K, Wang X, Wen T,
Chamberlin A, Stucke EM, Collins MH, Abonia JP, Peng Y, Wu Q, et
al: Whole-exome sequencing uncovers oxidoreductases dhtkd1 and
ogdhl as linkers between mitochondrial dysfunction and eosinophilic
esophagitis. JCI Insight. 3:2018. View Article : Google Scholar
|
|
30
|
Kim JH and Lee DC: Mitochondrial DNA copy
number in peripheral blood is associated with femoral neck bone
mineral density in postmenopausal women. J Rheumatol. 39:1465–1472.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Glatt S, Halbauer D, Heindl S, Wrnitznig
A, Kozina D, Su KC, Puri C, Garin-Chesa P and Sommergruber W:
Hgpr87 contributes to viability of human tumor cells. Int J Cancer.
122:2008–2016. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Y, Scoumanne A and Chen X: G
protein-coupled receptor 87: A promising opportunity for cancer
drug discovery. Mol Cell Pharmacol. 2:111–116. 2010.PubMed/NCBI
|
|
33
|
Wang L, Zhou W, Zhong Y, Huo Y, Fan P,
Zhan S, Xiao J, Jin X, Gou S, Yin T, et al: Overexpression of g
protein-coupled receptor gpr87 promotes pancreatic cancer
aggressiveness and activates nf-κb signaling pathway. Mol Cancer.
16:612017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hatzimichael E, Dasoula A, Kounnis V,
Benetatos L, Lo Nigro C, Lattanzio L, Papoudou-Bai A, Dranitsaris
G, Briasoulis E and Crook T: Bcl2-interacting killer cpg
methylation in multiple myeloma: A potential predictor of
relapsed/refractory disease with therapeutic implications. Leuk
Lymphoma. 53:1709–1713. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Langemeijer SM, Mariani N, Knops R,
Gilissen C, Woestenenk R, de Witte T, Huls G, van der Reijden BA
and Jansen JH: Apoptosis-related gene expression profiling in
hematopoietic cell fractions of MDS patients. PLoS One.
11:e01655822016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hoff AM, Johannessen B, Alagaratnam S,
Zhao S, Nome T, Løvf M, Bakken AC, Hektoen M, Sveen A, Lothe RA and
Skotheim RI: Novel rna variants in colorectal cancers. Oncotarget.
6:36587–36602. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Figlioli G, Kohler A, Chen B, Elisei R,
Romei C, Cipollini M, Cristaudo A, Bambi F, Paolicchi E, Hoffmann
P, et al: Novel genome-wide association study-based candidate loci
for differentiated thyroid cancer risk. J Clin Endocrinol Metab.
99:E2084–E2092. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cha S, Lim JE, Park AY, Do JH, Lee SW,
Shin C, Cho NH, Kang JO, Nam JM, Kim JS, et al: Identification of
five novel genetic loci related to facial morphology by genome-wide
association studies. BMC Genomics. 19:4812018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ohara K, Arai E, Takahashi Y, Ito N,
Shibuya A, Tsuta K, Kushima R, Tsuda H, Ojima H, Fujimoto H, et al:
Genes involved in development and differentiation are commonly
methylated in cancers derived from multiple organs: A
single-institutional methylome analysis using 1007 tissue
specimens. Carcinogenesis. 38:241–251. 2017.PubMed/NCBI
|