Introduction
Autoimmune disorders are mediated by self-cells;
that is, induced by the immune response to autoantigens (1). Though common autoimmune disorders
have high incidence rates, the pathogeneses of these diseases
remain incompletely characterized (2). At present, to the best of our
knowledge, no drug has been identified to be sufficiently effective
for the treatment of autoimmune diseases, and clinical therapy is
largely based on symptomatic treatment and targeting the control of
disease progression (3–5). Therefore, exploring the pathogenesis
of these diseases is of great significance in developing effective
drugs and clinical strategies for the treatment of autoimmune
diseases.
Vitamin D3 is a derivative of cholesterol and the
most active form of Vitamin D3 is 1,25-dihydroxy-vitamin D3
[1,25(OH)2D3], formed by the action of hepatic
microsomal 25-hydroxylase and kidney mitochondrial 1α hydroxylase
(6). Previous studies have
demonstrated that abnormal levels of 1,25(OH)2D3 are
closely associated with various autoimmune diseases (7,8). At
present, studies investigating 1,25(OH)2D3 and immune
function have attracted much attention. It has been identified that
1,25(OH)2D3 blocked the differentiation and maturation
of dendritic cells (DCs) to promote T cell responses and
differentiation (9,10). 1,25(OH)2D3 also promotes
B lymphocyte-programmed death and inhibits the differentiation of
activated B cells into plasma cells and memory B lymphocytes, which
secrete immunoglobulins (11,12).
However, only a limited number of studies have examined the
macrophage polarization promoted by 1,25(OH)2D3.
It has been identified that 1,25(OH)2D3
promotes macrophage polarization: Macrophages are induced by tumor
necrosis factor α (TNF-α) or interleukin-10 (IL-10) (13,14),
and macrophage polarization to classically activated macrophages
(M1) may assist in virus clearance and participate in inflammatory
reactions; however, polarization of macrophages to alternatively
activated macrophages (M2) may promote tumor development and
inhibit inflammatory responses (15,16).
The polarization mechanism of macrophages has been
studied extensively. T-cell Immunoglobulin-mucin-3 (Tim-3) was
identified as a novel immunomodulatory protein (17). Tim-3 has the function of regulating
natural immune cells (18). At
present, previous studies have identified that Tim-3 may promote
macrophage polarization to M2; however, its mechanism has not been
fully described (19,20). Tim-3 belongs to the Tim protein
family (21). Previous studies
have suggested that Tim-3 is significantly upregulated in chronic
infectious diseases, including chronic Hepatitis C virus infection
(22), Human immunodeficiency
virus infection (23) and
tuberculosis (24) and
downregulated in autoimmune diseases, including multiple sclerosis
(25) and rheumatoid arthritis
(17). Therefore, Tim-3 is
considered to be included in a novel generation of immunomodulatory
targets.
In the present study, genetic interference
techniques were applied to explore the effect of
1,25(OH)2D3 on macrophages polarization, and its
mechanism. The expression levels of polarization-associated factors
with or without Tim-3 expression were detected. The results of the
present study enrich the current understanding of the mechanisms
underlying autoimmune diseases, and suggest alternative approaches
for disease treatment.
Materials and methods
Cells culture
The mouse macrophage RAW264.7 cell line was
purchased from the American Type Culture Collection (Rockville, MD,
USA). Cells were cultured in high-glucose Dulbecco's modified
Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) that contained 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.) with penicillin (100 U/ml)
and streptomycin sulfate (100 µg/ml) at 37°C in an incubator with
5% CO2. Culture reagents were purchased from Gibco
(Thermo Fisher Scientific, Inc.).
Cell transfection
The recombinant Tim-3 overexpression plasmid
(pTim-3), empty vector (pScramble) and small interfering (si)Tim-3
were purchased from Promega Corporation (Madison, WI, USA). The
sequences of siTim-3 were Sense 5′:GGUGUCUUAAUCCUUAAAU, and
Antisense 3′:CCACAGAAUUAGGAAUUUA. The cells were seeded at a
density of 1×104 cells/well into a 24-well plate. The
cells were then starved overnight by incubating them in DMEM
without FBS prior to cell transfection. When the confluence reached
60–70%, the cells were transfected with pTim-3 or siTim-3 (50 pmol)
using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the protocol of the manufacturer.
The time interval between transfection and subsequent
experimentation was 24 h. For screening the positive clones, the
RAW264.7 cell line with the stable knockdown or Tim-3
overexpression plasmid was cultured in the aforementioned complete
medium with 200 and 700 µg/ml G418 respectively, at 37°C for 24 h.
According to data from previous studies (26–28),
10−8 or 10−7 M 1,25(OH)2D3 was
used to treat the RAW264.7 cells for 3 days. Cell morphology was
observed using a laser confocal microscope, with magnifications
×100 and ×200 (Leica TCS SP8; Leica Microsystems, Wetzlar,
Germany).
ELISAs
ELISAs were used to measure the concentrations of
TNF-α, IL-6, IL-10 and transforming growth factor-β (TGF-β). The
kits were Mouse TNF-α Quantikine ELISA Kit (MTA00B, R&D
Systems, Inc., Minneapolis, MN, USA), Mouse IL-6 Quantikine ELISA
Kit (M6000B, R&D), Mouse IL-10 Quantikine ELISA Kit (M1000B,
R&D), and Mouse/Rat/Porcine/Canine TGF-beta 1 Quantikine ELISA
Kit (MB100B, R&D). Following washing, blocking solution was
added at 4°C and incubated for 2 h. Next, the primary antibody was
added at 4°C and incubated overnight. The secondary antibody was
then added and incubated for 1 h at room temperature. Horseradish
peroxidase was added dropwise for 30 min at room temperature, and
then tetramethylbenzidine was added for 10 min. The absorbance
value was measured at 450 nm using a microplate reader (ELX 800;
Bio-Tek Instruments, Winooski, VT, USA) and the concentration was
calculated following the standard curve.
Western blot analysis
The expression levels of Tim-3, nitric oxide
synthase, inducible (NOS2) and macrophage mannose receptor 1
(COD206) proteins were detected using western blot analysis. Cells
were lysed with liquid nitrogen and radioimmunoprecipitation assay
lysis buffer (Abmole Bioscience Inc., Houston, TX, USA). Next, the
cells were cleaved with 1% phenylmethyl sulfonyl fluoride mixed
with phosphatase inhibitors (Abmole Bioscience Inc) for 30 min at
4°C. The supernatant was collected using centrifugation at 13,000 ×
g at 4°C for 15 min. A standard curve was drawn using the BCA
method to determine the protein concentration. Then, proteins (20
µg/lane) were separated using 10% SDS-PAGE gels, and transferred to
polyvinylidene difluoride (PVDF) membranes (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) using a Trans-Blot Transfer Slot (Bio-Rad
Laboratories, Inc.). The PVDF membranes were then blocked with 5%
skimmed milk for 2 h at room temperature. Primary antibodies
against Tim-3 (cat. no. ab185703; 1:800; Abcam, Cambridge, MA,
USA), NOS2 (cat. no. 14-5920-80; 1:800; eBioscience; Thermo Fisher
Scientific, Inc.), CD206 (cat. no. 141703; 1:600; BioLegend, Inc.,
San Diego, CA, USA) were then added according to the manufacturers'
protocols. The samples were agitated at room temperature for 2 h
and then incubated at 4°C for 12 h. Rabbit anti-mouse IgG (cat. no.
ab99697; 1:9,000 Abcam), mouse anti-rabbit IgG (cat. no. BA1034;
1:1,000; Invitrogen; Thermo Fisher Scientific, Inc.) and goat
anti-mouse IgG (cat. no. ab6785; 1:8,000; Abcam) HRP-conjugated
secondary antibodies were added and incubated at room temperature
for 1.5 h. Chemiluminescence detection was performed using ECL
reagent (EMD Millipore, Billerica, MA, USA). The densitometric
analysis was performed using Bio-Rad ChemiDoc™ XRS+ System with
Image Lab™ Software version 4.1 (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR) analysis
RT-qPCR was performed to determine the mRNA levels
of Tim-3, NOS2 and COD206. The cells were triturated and lysed by
TRIzol® (Thermo Fisher Scientific, Inc.) at 0°C for 5
min. RNA was then extracted using CCl3 (Shanghai Aladdin
Biochemical Technology Co., Ltd., Shanghai, China) and dissolved in
diethyl pyrocarbonate-treated water (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). RNA concentration was measured using a UV
NanoDrop One Microvolume UV–Vis spectrophotometer (Thermo Fisher
Scientific, Inc., Wilmington, DE, USA). Reverse transcription
assays were performed on RNA samples to synthesize cDNA using a
reverse transcription kit (Takara Biotechnology, Co., Ltd., Dalian,
China). The samples were incubated at 37°C for 15 min, followed by
heat inactivation at 85°C for 15 sec. RT-qPCR experiments were
performed using the SYBR Prellix Ex Taq™ Real-Time PCR kit (Takara
Biotechnology, Co., Ltd.). qPCR was performed by activating the DNA
polymerase at 95°C for 5 min, which was followed by 40 cycles of a
two-step PCR (at 95°C for 10 sec and at 60°C for 30 sec) with a
final extension step at 75°C for 10 min. Samples were then held at
4°C. β-actin was used as an internal control. All primers were
purchased from Genewiz, Inc., (Suzhou, Jiangsu China), and are
summarized in Table I. The
2−ΔΔCq method was used to quantify the gene expression
levels (29).
 | Table I.Sequences of primers. |
Table I.
Sequences of primers.
| Primer name | Sequence
(5′-3′) | Product size,
bp |
|---|
| Tim-3-forward |
CTCCAAGAACCCTAACCACG |
|
| Tim-3-reverse |
AGCCCATGTGGAAATTTTTG | 288 |
| NOS2-forward |
CAGCTGGGCTGTACAAACCTT |
|
| NOS2-reverse |
CATTGGAAGTGAAGCGTTTCG | 245 |
| CD206-forward |
TTCGGACACCCATC-GGAATTT |
|
| CD206-reverse |
CACAAGCGCTGCGTGGAT | 262 |
|
β-actin-forward |
GCTGCGTGTGGCCCCTGAG |
|
|
β-actin-reverse |
ACGCAGGATGGCATGAGGGA | 252 |
Statistical analysis
All experimental data are presented as means ±
standard deviation. Statistical analysis was performed using SPSS
20 (IBM Corp., Armonk, NY, USA). The one-way analysis of variance
followed by Tukey's multiple comparison post-hoc test was performed
to analyze the differences among experimental groups.
P<0.05 was considered to indicate a statistically
significant difference.
Results
Confocal microscopy
The viable RAW264.7 macrophages were normal at
magnifications, ×100 and ×200 (Fig.
1).
1,25(OH)2D3 regulates the
secretion of inflammatory cytokines in RAW264.7 cell
It was identified that the levels of TNF-α and IL-6
secreted by RAW264.7 cells were decreased in the presence of
1,25(OH)2D3. However, the levels of IL-10 and TGF-β were
increased (P<0.01; Fig. 2A-D).
These differences were the most significant at concentrations of
10−7 M (P<0.05).
 | Figure 2.Effects of 1,25(OH)2D3 on
macrophage at 10−8 and 10−7 M concentrations.
RAW264.7 cells were cultured at different 1,25(OH)2D3
concentrations (0, 10−8 and 10−7 M). ELISAs
were used to detect the (A) IL-6, (B) IL-10, (C) TNF-α and (D)
TGF-β protein expression levels among different groups. (E) Tim-3,
(F) NOS2 and (G) CD206 protein levels were assessed by western blot
analysis. (H) Representative western blot image. (I) Tim-3, (J)
NOS2 and (K) CD206 mRNA levels were analyzed by reverse
transcription quantitative polymerase chain reaction assays.
*P<0.05 and **P<0.01 vs. control group; #P<0.05
and ##P<0.01 vs. 10−8 M group. Data are
presented as mean ± standard deviation (n=5).
1,25(OH)2D3, 1,25-dihydroxy-vitamin D3; IL, interleukin;
TNF-α, tumor necrosis factor α; TGF-β, transforming growth factor;
Tim-3, T-cell Immunoglobulin-mucin-3; NOS2, nitric oxide synthase,
inducible; CD206, macrophage mannose receptor 1. |
1,25(OH)2D3 upregulates
Tim-3 expression
To investigate the mechanism of how 1,25
(OH)2D3 affects the expression levels of inflammatory
factors, levels of Tim-3, and NOS2 and COD206, the markers of M1
and M2, were examined. The results indicated that the expression
level of Tim-3 was gradually increased in the 10−8 M and
10−7 M groups. However, whilst 1,25 (OH)2D3
did not affect the expressions of the NOS2 and COD206, it decreased
NOS2 mRNA levels (P<0.05; Fig.
2E-K). This observation indicated that 1,25(OH)2D3
promoted Tim-3 expression in a dose-dependent manner.
Effects of Tim-3 gene silencing on
cytokines
Following silencing of the Tim-3 gene, the levels of
TNF-α and IL-6 secreted by RAW264.7 cells were upregulated, while
the levels of IL-10 and TGF-β were downregulated (P<0.01;
Fig. 3A-D). Following the decrease
of the Tim-3 gene expression, the mRNA and protein expression
levels of NOS2 were observed increased compared with that of the NC
and control groups, while the expression levels of CD206 mRNA and
protein were significantly decreased (P<0.01; Fig. 3E-K). This phenomenon suggested that
silence of Tim-3 gene promoted the polarization of macrophages to
M1.
 | Figure 3.Effects of Tim-3 gene silencing on
macrophages. ELISAs were applied to detect the (A) IL-6, (B) IL-10,
(C) TNF-α and (D) TGF-β levels in the control, NC and siTim-3
groups. (E) Tim-3, (F) NOS2 and (G) CD206 protein levels were
assessed by western blot analysis. (H) Representative western blot
image. (I) Tim-3, (J) NOS2 and (K) CD206 mRNA levels were analyzed
by reverse transcription quantitative polymerase chain reaction
assays. **P<0.01 vs. control group; ##P<0.01 vs.
NC group. Data are presented as mean ± standard deviation (n=5).
IL, interleukin; TNF-α, tumor necrosis factor α; TGF-β,
transforming growth factor; Tim-3, T-cell Immunoglobulin-mucin-3;
NOS2, nitric oxide synthase, inducible; CD206, macrophage mannose
receptor 1; si, small interfering. |
Effects of overexpression of Tim-3 on
cytokines
The effects of the overexpression of Tim-3 on
cytokines were also examined. It was identified that the levels of
TNF-α and IL-6 in the pTim-3 group were decreased compared with
those in the NC group, and that the levels of IL-10 and TGF-β were
increased compared with those in the NC group (P<0.01; Fig. 4A-D). When Tim-3 was overexpressed,
the mRNA and protein expression levels of NOS2 were decreased,
while the expression levels of CD206 mRNA and protein were
increased (P<0.01; Fig. 4E-K).
This suggested that the overexpression of Tim-3 promoted the
polarization of macrophages to M2.
 | Figure 4.Effects of Tim-3 gene overexpression
on macrophages. The levels of (A) IL-6, (B) IL-10, (C) TNF-α and
(D) TGF-β in the control, NC and pTim-3 groups were detected using
ELISAs. (E) Tim-3, (F) NOS2 and (G) CD206 protein levels were
assessed by western blot analysis. (H) Representative western blot
image. (I) Tim-3, (J) NOS2 and (K) CD206 mRNA levels were analyzed
by reverse transcription quantitative polymerase chain reaction
assays. **P<0.01 vs. control group; ##P<0.01 vs.
NC group. Data are presented as mean ± standard deviation (n=5).
IL, interleukin; TNF-α, tumor necrosis factor α; TGF-β,
transforming growth factor; Tim-3, T-cell Immunoglobulin-mucin-3;
NOS2, nitric oxide synthase, inducible; CD206, macrophage mannose
receptor 1; pTim-3, overexpressing Tim-3 plasmid. |
Effects of 1,25(OH)2D3 on
cytokine under Tim-3 silencing or overexpression conditions
To additionally explore the effect of 1,25(OH)2D3 on
the polarization of macrophages, RAW264.7 cells were cultured in
the presence of 10−7 M 1,25(OH)2D3. The
expression levels of inflammatory factors, and M1 and M2 marker
proteins NOS2 and CD206, were detected.
It was identified that 1,25(OH)2D3 downregulated the
levels of TNF-α and IL-6 and upregulated the level of TGF-β in
siTim-3 cells. In addition, 1,25(OH)2D3 promoted the
expression of Tim-3 and increased the expression levels of CD206
protein in siTim-3 cells. However, 1,25(OH)2D3 decreased
the expression levels of NOS2. For RAW264.7 cells with stable
overexpression of Tim-3, 1,25(OH)2D3 was observed to additionally
decrease TNF-α and IL-6 levels and increase IL-10 and TGF-β levels,
compared to the Tim-3 overexpression group. In addition, 1,25
(OH)2D3 increased the level of Tim-3 and the expression
levels of CD206 in RAW264.7 cell with stable expression of Tim-3
gene (Fig. 5A-K). This indicated
that 1,25(OH)2D3 promoted the polarization of
macrophages to M2 and inhibited polarization of macrophages to M1
via upregulating the expression of Tim-3.
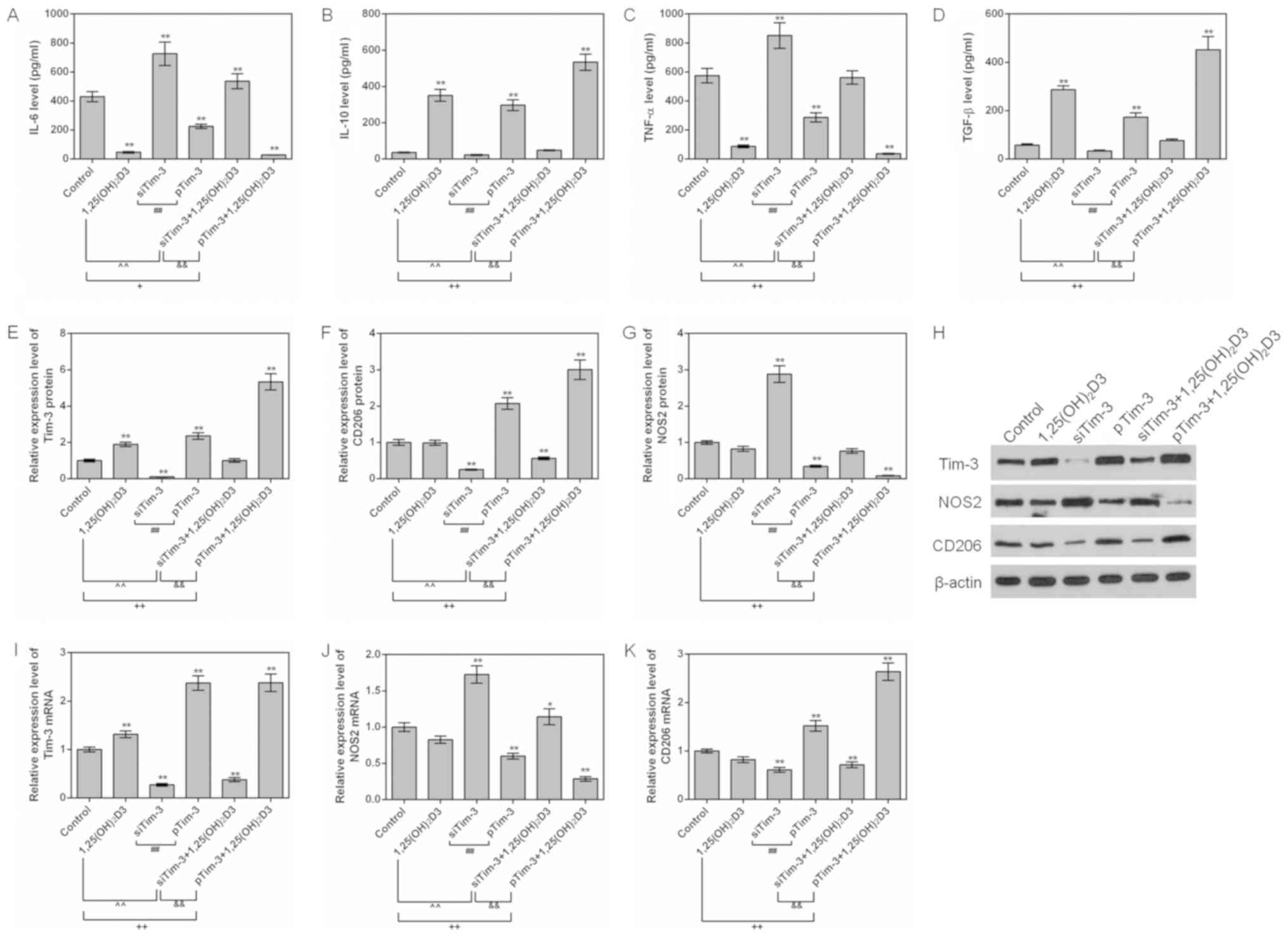 | Figure 5.Effects of 1,25(OH)2D3 on
macrophages with different Tim-3 expression levels. RAW264.7
macrophages with silenced Tim-3 expression and Tim-3 overexpression
were cultured with 10−7 M 1,25(OH)2D3. ELISAs
were used to assess the levels of (A) IL-6, (B) IL-10, (C) TNF-α
and (D) TGF-β in the control group. (E) Tim-3, (F) CD206 and (G)
NOS2 protein levels were assessed by western blot analysis. (H)
Representative western blot image. (I) Tim-3, (J) NOS2 and (K)
CD206 mRNA levels were analyzed by reverse transcription
quantitative polymerase chain reaction assays. *P<0.05 and
**P<0.01 vs. control group; +P<0.05;
++P<0.01; ##P<0.01;
&&P<0.01. Data are presented as mean ±
standard deviation (n=5). 1,25(OH)2D3,
1,25-dihydroxy-vitamin D3; IL, interleukin; TNF-α, tumor necrosis
factor α; TGF-β, transforming growth factor; Tim-3, T-cell
Immunoglobulin-mucin-3; NOS2, nitric oxide synthase, inducible;
CD206, macrophage mannose receptor 1; pTim-3, overexpressing Tim-3
plasmid; si, small interfering. |
Discussion
Macrophages may be induced to polarize to M1 or M2
subtypes by different cytokines, and also secrete different
cytokines to promote or suppress the immune response (30). Macrophage cells may also maintain
the homeostasis and serve an important role in the formation and
treatment of diseases (31).
It has been identified that Vitamin D3 has
immunomodulatory effects on chicken macrophages (32). Yin et al (33) also revealed that Vitamin D may
induce macrophage polarization by activating CYP27A1 to prevent
atherosclerosis in hypercholesterolemic animals. The present study
demonstrated that 1,25(OH)2D3 inhibited TNF-α and IL-6
and promoted the secretion of IL-10 and TGF-β. TNF-α induces
macrophage polarization to M1 and the secretion of TNF-α and IL-6.
IL-10 induces macrophage polarization to M2 and secretion of IL-10
and TGF-β (13,14). The results of the present study
indicated that 1,25(OH)2D3 decreased the level of NOS2
mRNA, but did not produce a significant effect on NOS2 protein
expression or CD206 protein and mRNA expression. However, the
expression of Tim-3 was increased by 1,25(OH)2D3. Tim-3
serves a role in modulating immunity (34), through inducing macrophage
polarization. This suggests that 1,25(OH)2D3 may
regulate macrophage polarization through Tim-3. Therefore, the
effect of 1,25(OH)2D3 on macrophages was investigated
through silencing or overexpressing Tim-3. The results demonstrate
that silencing of the Tim-3 gene may promote the secretion of TNF-α
and IL-6 but inhibit the secretion of IL-10 and TGF-β. In addition,
downregulation of Tim-3 increased the protein and mRNA levels of
NOS2 and decreased the protein and mRNA levels of the M2 marker
CD206. By contrast, overexpression of the Tim-3 gene exerted the
opposite effects This is consistent with previous data (17,22–25),
indicating that a high expression level of Tim-3 gene promotes
macrophage polarization to M2 and produces immunosuppressive
effects. Therefore, we hypothesized that 1,25(OH)2D3 may
induce cell polarization to M2 by upregulating Tim-3 expression and
exerting immunosuppressive effects.
The role of 1,25(OH)2D3 in macrophages
polarization was also explored. The data indicated that the
decreased levels of IL-6 and TNF-α and the increased IL-10 and
TGF-β were reversed in the presence of siTim-3. However, the effect
of 1,25(OH)2D3 was enhanced by the overexpression of
Tim-3. This suggested that 1,25(OH)2D3 produced an
immunosuppressive effect by promoting Tim-3 protein. It was
consistent with a previous study, in which 1,25(OH)2D3 was
identified to induce macrophage polarization to M2, leading to
immunosuppression (33).
However, the mechanism underlying the regulation of
Tim-3 protein levels by 1,25(OH)2D3 remains unclear. An
additional limitation of the present study is that RAW264.7 is a
cancerous macrophage cell line, as opposed to a primary cell line.
Therefore, additional investigation in primary cells or in
vivo experiments would strengthen the results of the present
study. Altogether, the mechanism for the regulation of M1/M2
conversion and inflammation by Tim-3 remains to be determined.
In summary, Tim-3 induces macrophage polarization to
M2. 1,25(OH)2D3 exhibited immunosuppressive effects by
upregulating Tim-3 protein levels. The present study suggested that
1,25(OH)2D3 may serve as an immunosuppressive drug and
that Tim-3 may be a potential target in treating autoimmune
diseases.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
SL and JC did all of the experiments, YL analyzed
the data, RY designed the study, and SL wrote the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Benros ME, Waltoft BL, Nordentoft M,
Ostergaard SD, Eaton WW, Krogh J and Mortensen PB: Autoimmune
diseases and severe infections as risk factors for mood disorders:
A nationwide study. JAMA Psychiatry. 70:812–820. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen SW, Zhong XS, Jiang LN, Zheng XY,
Xiong YQ, Ma SJ, Qiu M, Huo ST, Ge J and Chen Q: Maternal
autoimmune diseases and the risk of autism spectrum disorders in
offspring: A systematic review and meta-analysis. Behav Brain Res.
296:61–69. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Iyer A, Elsone L, Appleton R and Jacob A:
A review of the current literature and a guide to the early
diagnosis of autoimmune disorders associated with neuromyelitis
optica. Autoimmunity. 47:154–161. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nielsen PR, Benros ME and Dalsgaard S:
Associations between autoimmune diseases and
attention-deficit/hyperactivity disorder: A nationwide study. J Am
Acad Child Adolesc Psychiatry. 56:234–240.e231. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Andersson NW, Gustafsson LN, Okkels N,
Taha F, Cole SW, Munk-Jorgensen P and Goodwin RD: Depression and
the risk of autoimmune disease: A nationally representative,
prospective longitudinal study. Psychol Med. 45:3559–3569. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ota K, Dambaeva S, Kim MW, Han AR, Fukui
A, Gilman-Sachs A, Beaman K and Kwak-Kim J: 1,25-Dihydroxy-vitamin
D3 regulates NK-cell cytotoxicity, cytokine secretion, and
degranulation in women with recurrent pregnancy losses. Eur J
Immunol. 45:3188–3199. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Alhassan Mohammed H, Saboor-Yaraghi AA,
Mirshafiey A, Vahedi H, Shiri-Shahsavar MR and Mousavi Nasl
Khameneh A: Immunomodulatory and immunosuppressive roles of
1α,25(OH)2D3 in autoimmune diseases. Scand J Immunol. 85:95–103.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fitch N, Becker AB and HayGlass KT:
Vitamin D [1,25(OH)2D3] differentially regulates human innate
cytokine responses to bacterial versus viral pattern recognition
receptor stimuli. J Immunol. 196:2965–2972. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ferreira GB, Gysemans CA, Demengeot J, da
Cunha JP, Vanherwegen AS, Overbergh L, Van Belle TL, Pauwels F,
Verstuyf A, Korf H and Mathieu C: 1,25-Dihydroxyvitamin D3 promotes
tolerogenic dendritic cells with functional migratory properties in
NOD mice. J Immunol. 192:4210–4220. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ria F, Penna G and Adorini L: Th1 cells
induce and Th2 inhibit antigen-dependent IL-12 secretion by
dendritic cells. Eur J Immunol. 28:2003–2016. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sergeev IN, Pletsityi KD, Rusnak FI and
Spirichev VB: 1,25-Dihydroxyvitamin D3 receptors in lymphocytes and
T- and B-lymphocyte count in patients with glomerulonephritis. Vopr
Med Khim. 35:117–121. 1989.(In Russian). PubMed/NCBI
|
|
12
|
Geldmeyer-Hilt K, Heine G, Hartmann B,
Baumgrass R, Radbruch A and Worm M: 1,25-dihydroxyvitamin D3
impairs NF-kappaB activation in human naive B cells. Biochem
Biophys Res Commun. 407:699–702. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Davies LC, Jenkins SJ, Allen JE and Taylor
PR: Tissue-resident macrophages. Nat Immunol. 14:986–995. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wynn TA, Chawla A and Pollard JW:
Macrophage biology in development, homeostasis and disease. Nature.
496:445–455. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jablonski KA, Amici SA, Webb LM,
Ruiz-Rosado Jde D, Popovich PG, Partida-Sanchez S and
Guerau-de-Arellano M: Novel markers to delineate murine M1 and M2
macrophages. PLoS One. 10:e01453422015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cucak H, Grunnet LG and Rosendahl A:
Accumulation of M1-like macrophages in type 2 diabetic islets is
followed by a systemic shift in macrophage polarization. J Leukoc
Biol. 95:149–160. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee J, Park EJ, Noh JW, Hwang JW, Bae EK,
Ahn JK, Koh EM and Cha HS: Underexpression of TIM-3 and blunted
galectin-9-induced apoptosis of CD4+ T cells in rheumatoid
arthritis. Inflammation. 35:633–637. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kuchroo VK, Meyers JH, Umetsu DT and
DeKruyff RH: TIM family of genes in immunity and tolerance. Adv
Immunol. 91:227–249. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jiang X, Zhou T, Xiao Y, Yu J, Dou S, Chen
G, Wang R, Xiao H, Hou C, Wang W, et al: Tim-3 promotes
tumor-promoting M2 macrophage polarization by binding to STAT1 and
suppressing the STAT1-miR-155 signaling axis. Oncoimmunology.
5:e12112192016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun J, Huang Q, Li S, Meng F, Li X and
Gong X: miR-330-5p/Tim-3 axis regulates macrophage M2 polarization
and insulin resistance in diabetes mice. Mol Immunol. 95:107–113.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ying H, Kang Y, Zhang H, Zhao D, Xia J, Lu
Z, Wang H, Xu F and Shi L: MiR-127 modulates macrophage
polarization and promotes lung inflammation and injury by
activating the JNK pathway. J Immunol. 194:1239–1251. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Moorman JP, Wang JM, Zhang Y, Ji XJ, Ma
CJ, Wu XY, Jia ZS, Wang KS and Yao ZQ: Tim-3 pathway controls
regulatory and effector T cell balance during hepatitis C virus
infection. J immunol. 189:755–766. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tandon R, Giret MT, Sengupta D, York VA,
Wiznia AA, Rosenberg MG, Kallas EG, Ndhlovu LC and Nixon DF:
Age-related expansion of Tim-3 expressing T cells in vertically
HIV-1 infected children. PLoS One. 7:e457332012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qiu Y, Chen J, Liao H, Zhang Y, Wang H, Li
S, Luo Y, Fang D, Li G, Zhou B, et al: Tim-3-expressing CD4+ and
CD8+ T cells in human tuberculosis (TB) exhibit polarized effector
memory phenotypes and stronger anti-TB effector functions. PLoS
Pathog. 8:e10029842012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Koguchi K, Anderson DE, Yang L, O'Connor
KC, Kuchroo VK and Hafler DA: Dysregulated T cell expression of
TIM3 in multiple sclerosis. J Exp Med. 203:1413–1418. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Villaggio B, Soldano S and Cutolo M:
1,25-dihydroxyvitamin D3 downregulates aromatase expression and
inflammatory cytokine production in human macrophages. Clin Exp
Rheumatol. 30:934–938. 2012.PubMed/NCBI
|
|
27
|
Chambers ES, Suwannasaen D, Mann EH, Urry
Z, Richards DF, Lertmemongkolchai G and Hawrylowicz CM:
1α,25-dihydroxyvitamin D3 in combination with transforming growth
factor-β increases the frequency of Foxp3+ regulatory T
cells through preferential expansion and usage of interleukin-2.
Immunology. 143:52–60. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Karmali R, Hewison M, Rayment N, Farrow
SM, Brennan A, Katz DR and O'Riordan JL: 1,25(OH)2D3 regulates
c-myc mRNA levels in tonsillar T lymphocytes. Immunology.
74:589–593. 1991.PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
McNelis JC and Olefsky JM: Macrophages,
immunity, and metabolic disease. Immunity. 41:36–48. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Saeed S, Quintin J, Kerstens HH, Rao NA,
Aghajanirefah A, Matarese F, Cheng SC, Ratter J, Berentsen K, van
der Ent MA, et al: Epigenetic programming of monocyte-to-macrophage
differentiation and trained innate immunity. Science.
345:12510862014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shojadoost B, Behboudi S, Villanueva AI,
Brisbin JT, Ashkar AA and Sharif S: Vitamin D3 modulates the
function of chicken macrophages. Res Vet Sci. 100:45–51. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yin K, You Y, Swier V, Tang L, Radwan MM,
Pandya AN and Agrawal DK: Vitamin D protects against
atherosclerosis via regulation of cholesterol efflux and macrophage
polarization in hypercholesterolemic swine. Arterioscler Thromb
Vasc Biol. 35:2432–2442. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Anderson AC, Joller N and Kuchroo VK:
Lag-3, Tim-3, and TIGIT: Co-inhibitory receptors with specialized
functions in immune regulation. Immunity. 44:989–1004. 2016.
View Article : Google Scholar : PubMed/NCBI
|















