Introduction
In recent years, the incidence of diabetes and
associated mortalities have exhibited a trend of rapid growth
worldwide (1). Diabetic
cardiomyopathy (DCM) is a major complication of diabetes. The major
pathological changes involved are myocardial hypertrophy, apoptosis
and myocardial interstitial fibrosis (2). Of these, myocardial fibrosis is the
major pathological feature, and may eventually induce cardiac
remodeling, cardiac dilatation, cardiac dysfunction, arrhythmia and
congestive heart failure (3).
According to recent data (4),
cardiac fibroblasts (CFs) are highly activated during diabetes,
which results in a disorder of the dynamic balance of cardiac
extracellular matrix synthesis and deposition, along with the
excessive deposition of collagen (Col), thus leading to myocardial
fibrosis and cardiac dysfunction (5,6).
However, the exact mechanisms underlying myocardial fibrosis in DCM
remain elusive.
The calcium-sensitive receptor (CaSR) is a member of
the C family of the G protein coupling receptor superfamily and is
widely expressed in prokaryotic and eukaryotic cells. CaSR is
involved in regulating the homeostasis of calcium and other metal
ions, proliferation, differentiation, chemotaxis, apoptosis, gene
expression, membrane potential, and aging (7,8).
Studies by our group and other research groups have
demonstrated that the CaSR is involved in myocardial
ischemia/reperfusion injury, myocardial infarction and pulmonary
hypertension (9,10). A recent study by our group
indicated that the expression of CaSR in the myocardium tissues of
diabetic rats, and CFs treated with high concentrations of glucose
was significantly increased (11).
However, whether CaSR participates in diabetic myocardial fibrosis
has remained elusive.
In the present study, a rat model of type 1 diabetes
(T1D) and CFs cultured under high-glucose (HG) conditions were
subjected to treatment with a CaSR agonist or inhibitor in order to
explore the functional role of CaSR in diabetic myocardial fibrosis
and the underlying mechanisms.
Materials and methods
Animal experimental protocol
Male Wistar rats (weight, 250±50 g; age, 8 weeks)
were provided by the Animal Research Institute of Harbin Medical
University (HMU; Harbin, China). The study was approved by the HMU
Medical Science Ethics Committee. All rats were maintained under a
12-h light/dark cycle and fed with standard chow and clean water
ad libitum. The rats were randomly divided into four groups
(n=8/group): i) control group: age-matched non-diabetic Wistar rats
were injected with citric acid-citrate sodium buffer; ii) T1D
group: intraperitoneal injection of streptozotocin (STZ; 60 mg/kg;
Sigma-Aldrich; Merck KGaA) (12);
iii) T1D+R568 group: T1D rats were subcutaneously injected with
R568 (10 µmol/kg/day in saline); iv) T1D+Calhex231 group: T1D rats
were subcutaneously injected with Calhex231 (10 µmol/kg/day in
saline). Rats in the four groups were sacrificed after 12 weeks and
a range of indices were assessed.
Isolation and incubation of neonatal
rat CFs
Neonatal rat CFs were isolated from the hearts of
1–3-day-old Wistar rats. In brief, the heart was quickly removed,
immediately placed in D-Hank's solution, cut into pieces (0.5
mm3) and digested with trypsin (Beyotime Institute of
Biotechnology) for 8 min. The digestion was then terminated by
adding Dulbecco's modified Eagle's medium (DMEM). After the same
process had been repeated 8 times, cells were collected by
centrifugation at 800 × g for 10 min and a temperature of 4°C.
After 2 h of incubation, the unattached cells were discarded; the
attached cells (CFs) were plated in a petri dish and maintained at
37°C in a 5% CO2 humidified incubator in DMEM containing
10% fetal bovine serum and 1% penicillin-streptomycin. The media
was changed two times/week. To ensure the purity of the CFs, the
cells were passaged for three generations and then CFs were treated
with HG (40 mM), the CaSR agonist R568 (5 µM), the CaSR inhibitor
Calhex231 (3 µM), the transforming growth factor (TGF)-β-type I/II
receptor (TβRI/II) kinase inhibitor LY2109761 (20 µM) or
TGF-β1 (5, 10, 20 ng/ml; Shanghai San Shu Biotechnology,
Ltd.).
Analysis of serum and culture
media
Blood samples obtained from the aorta were
centrifuged at 1,200 × g for 20 min and serum was stored at −80°C
until assayed. Random serum insulin levels were determined using a
commercially available ultrasensitive ELISA kit (cat. no. 201804A;
Beyotime Institute of Biotechnology). Serum levels of
triacylglycerol (TG) and total cholesterol (TC) were analyzed using
a standard biochemistry panel (cat. no. 201812AS/201812ED; Beyotime
Institute of Biotechnology). Blood glucose in blood samples from
the tail vein was measured using a blood glucose analyzer
(ACCU-CHEK; Roche Diagnostics GmbH).
Culture media of the CFs were collected to determine
TGF-β1, Col-I/III and matrix metallopeptidase (MMP)-2/9
levels (Wuhan Boster Biological Technology, Ltd.). These cytokines
were determined by ELISA using commercially available detection
kits (cat. nos. EK0513, EK0411, EK0424, EK0467, EK0459, EK0465),
following the supplier's protocols.
Hematoxylin and eosin (H&E),
Masson trichrome and Sirius red staining
The rats were sacrificed with 200 mg/kg
pentobarbital sodium by intraperitoneal injection and the heart was
quickly removed and washed with phosphate buffer. The cardiac
tissue was fixed in 10% buffered formaldehyde at 4°C for 12 h,
embedded in paraffin, sliced at 4 mm and used for morphological
assessment. Pathological changes in cardiac tissues were observed
by H&E, paraffin sections were stained with 0.5% hematoxylin at
room temperature for 10 min and 0.5% eosin for 3 min. The extent of
myocardial fibrosis was determined by Masson trichrome (1%) and
Sirius red staining (0.5%); sections were stained for 10 min at
55°C and for 30 min at room temperature, respectively. Analysis was
conducted with an optical microscopy (BX61; Olympus
Corporation).
Western blot analysis
The rat hearts and CF cells were homogenized in 0.5
ml radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology) prior to transfer to small tubes and rotation for 1
h at 4°C. Solubilized proteins were collected after centrifugation
at 3,000 × g for 30 min at 4°C. The supernatant was then collected
and stored at −80°C. The protein concentration of each sample was
quantified using a Bicinchoninic Acid Protein Assay kit (Beyotime
Institute of Biotechnology). Protein lysates (20 µg/lane) from
cells of each group were separated by SDS-PAGE (12.5%) and
electro-transferred onto polyvinylidene difluoride membranes (EMD
Millipore). Non-specific proteins on membranes were blocked with 5%
non-fat dried milk for 2 h at room temperature. Subsequently, the
membranes were incubated overnight at 4°C with the following
primary antibodies (1:1,000 dilution): CaSR (cat. no. sc-47741),
TGF-β1 (cat. no. sc-130348); TβRI/II (cat. nos.
sc-518045 and sc-17799); and Col-I/III (cat. nos. sc-59772 and
sc-271249; all Santa Cruz Biotechnology, Inc.); Smad2 and
phosphorylated (p)-Smad2 (cat. nos. 5339 and 18338; Cell Signaling
Technology, Inc.); protein kinase C (PKC; cat. no. 2056); p-PKC
(cat. no. 2060); p38 (cat. no. 9212); p-p38 (cat. no. 9215); and
matrix metallopeptidase (MMP)2/9 (cat. nos. 4022 and 3852; all Cell
Signaling Technology, Inc.); and β-actin (cat. no. sc-69879) and
β-tubulin (cat. no. sc 55529; Santa Cruz Biotechnology, Inc.). The
membranes were then incubated with horseradish
peroxidase-conjugated anti-mouse/anti-rabbit immunoglobulin G
antibodies (cat. nos. bs-0295M-HRP and bs-0296R-HRP; BIOSS) at a
1:5,000 dilution for 1 h at room temperature. The specific complex
was visualized using an enhanced chemiluminescence plus western
blot detection system (Immobilon Western HRP; EMD Millipore). The
relative intensities of protein bands were finally quantified using
a Bio-Rad Chemi EQ densitometer (Bio-Rad Laboratories, Inc.) and
the band density was semi-quantified using AlphaView v3.2.2
software (ProteinSimple; Bio-Techne).
Detection of cell proliferation and
migration by Cell Counting Kit (CCK)-8 and a scratch wound repair
assay
CFs were plated onto 96-well plates at a density of
2×103 cells/well. After 12 h, the medium was replaced
with an antibiotic-free medium and the wells were divided into four
groups: control (5.5 mM), HG (40 mM), HG+R568 (40 mM + 5 µM), and
HG+Calhex231 (40 mM + 3 µM), with five replications/group. Finally,
cell proliferation was detected according to the CCK-8 assay kit
(cat. no. AR1199, Wuhan Boster Biological Technology, Ltd.)
instructions (absorbance at 450 nm).
Rat CFs treated with HG (40 mM), R568 (5 µM) or
Calhex231 (3 µM) were then subjected to scratch assays as
previously described (13). Images
were captured at 0 and 24 h after scratching using phase-contrast
microscopy.
Measurement of intracellular
Ca2+ using Fluo-3/AM probes
CFs treated with HG (40 mM), R568 (5 µM) or
Calhex231 (3 µM) were stained using 5 mM Fluo-3/AM (cat. no.
ab145254; Abcam) for 30 min at 37°C in the dark. Subsequently, the
cells were washed with Ca2+-free Tyrode's solution to
remove residual dye. The fluorescence of Ca2+ was then
measured by fluorescence microscopy (BX61; Olympus Corporation).
The excitation wavelength was 488 nm and the emission wavelength
was 530 nm.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
To determine the mRNA expression of Col-I, Col-III,
MMP-2 and MMP-9, total RNA was extracted from CFs using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Complementary DNA
was synthesized from 2 µg total RNA by using a Superscript Reverse
Transcription kit (TaqMan™; cat. no. 4366596; Thermo Fisher
Scientific, Inc.). For the quantification of the mRNA levels of
Col-I/III and MMP-2/9, PCR was performed using a SYBR Green PCR
Reagent kit (Applied Biosystems; Thermo Fisher Scientific, Inc.) in
a Light Cycler® 480 Sequence Detection System (Roche
Applied Science). β-actin served as the internal control and the
relative expression was determined. The primer sequences used in
the present study were as follows: Col-I forward,
5′-ATGTTCAGCTTTGTGGAC-3′ and reverse, 5′-GGATGCCATCTTGTCCAG-3′;
Col-III forward, 5′-CAAAGGAGAGCCAGGAGCAC-3′ and reverse,
5′-CTCCAGGCGAACCATCTTTG-3′; MMP-2 forward,
5′-TCAAATCGGACTGGCTGGGC-3′ and reverse, 5′-AACCAGGCCTCTTCACGTCC-3′;
MMP-9 forward, 5′-GAGGGGGAGGAGCTAGTTTGCC-3′ and reverse,
5′-AAGGACAGCGTGCAGAGAGGG-3′; β-actin forward,
5′-CCGGCTTCGCGGGCGACG-3′ and reverse, 5′-TCCCGGCCAGCCAGGTCC-3′.
Statistical analyses
All experiments were performed at least three times
independently. All data are expressed as the mean ± standard error
of the mean. Statistical analysis was performed by a two-tailed
Student's t-test or one-way analysis of variance, followed by the
Bonferroni multiple comparisons test using SPSS 18.0 software (SPSS
Inc.). P<0.05 was considered to indicate a statistically
significant difference.
Results
Successful modeling of T1D
cardiomyopathy
Blood glucose at weeks 2, 4, 8 and 12, along with
insulin levels, the ratio of heart weight to body weight (HW/BW),
as well as serum TGs and TCs on week 12 were determined. The
results indicated that compared with the control group, the blood
glucose levels at each time-point were higher, the insulin level
was significantly decreased, and TG and TC were significantly
increased in the T1D group, T1D+R568 group and T1D+Calhex231 group.
Notably, compared with that in the control group, HW/BW was
significantly increased in the T1D and T1D+R568 groups, but
compared with that in the T1D group, it was significantly decreased
in the T1D+Calhex231 group (Fig.
1).
 | Figure 1.Successful modeling of type 1
diabetic cardiomyopathy. (A) Random blood glucose was assessed at
weeks 2, 4, 8 and 12. Other indicators were analyzed at week 12
after successful modeling. Blood glucose, TG, TC and insulin levels
in the serum. (B) HW/BW. *P<0.05, **P<0.01, ***P<0.001,
****P<0.0001 vs. the Control; #P<0.05 vs. T1D rats
(n=8). TG, triacylglycerol; TC, total cholesterol; HW/BW, heart
weight to body weight ratio; T1D, type 1 diabetes. |
Masson staining and Sirius red staining revealed
large amounts of collagen deposition in the interstitial and
perivascular areas, particularly in denatured and necrotic areas;
H&E staining further indicated that the cardiac myocytes were
disordered and hypertrophic in the T1D group, T1D+R568 group and
T1D+Calhex231 group (Fig. 2A).
 | Figure 2.(A) Effects of CaSR on collagens and
morphological changes of myocardial tissue in rats. Masson's
trichrome staining (collagen deposition displayed in blue) and
Sirius red staining (collagen fibers displayed in red).
Representative images of H&E staining examined by transmission
electron microscopy. (B) Representative western blots for the
detection of CaSR, Col-I, Col-III and TGF-β1 in
comparison with β-actin expression in rats. *P<0.05,
**P<0.01, ****P<0.0001 vs. Controls; #P<0.05,
##P<0.01, ###P<0.001 vs. T1D rats
(n=8). TGF, transforming growth factor; T1D, type 1 diabetes; CaSR,
calcium-sensing receptor; Col, collagen. |
Effects of CaSR on collagen in T1D
rats
At week 12, western blot analysis of cardiac tissue
homogenate indicated that compared with that in the control group,
the expression of Col-I, Col-III and TGF-β1 was
increased in the T1D group and T1D+R568 groups, and the expression
of CaSR was significantly increased in the T1D+R568 group. However,
the opposite results were observed in the T1D+Calhex231 group
(Fig. 2B).
Effects of CaSR on proliferation and
migration of CFs
The effect of osmotic control was detected using
mannitol (40 mM), and the results revealed that mannitol had no
effect on the viability of cardiac fibroblasts (data not shown),
however, CFs treated with high glucose (40 mM) exhibited an
increase in viability in a time-dependent manner (data not shown).
These results indicated that the effect of high glucose on
viability may not be attributed to high osmolarity and treatment
with 40 mM glucose was considered as the high glucose group in
subsequent experiments.
The results of the scratch wound repair assay
indicated that the rate of CF migration was higher in the HG group
and the HG+R568 group compared with that in the control group,
while the migration was significantly reduced in the HG+Calhex231
group (Fig. 3A). The proliferation
(cell viability) of CFs at 24 h was detected using CCK-8 assays.
Compared with that in the control group, the cell proliferation was
greater in the HG group and the HG+R568 group, but was
significantly decreased in the HG+Calhex231 group (Fig. 3B).
Effects of CaSR on intracellular
calcium concentration and the amount of TGF-β1 secreted
by CFs
To further study the mechanism of diabetic
myocardial fibrosis, cytosolic Ca2+ was determined with
Fluo-3/AM staining and assessment of the fluorescence intensity.
The results indicated that the fluorescence intensity was higher in
the HG and HG+R568 groups, and was lower in the HG+Calhex231 group
(Fig. 4A). The protein levels of
p-PKC and p-P38 were significantly increased in the HG and HG+R568
groups. However, the opposite result was obtained in the Calhex231
group (Fig. 4B). The expression of
TGF-β1 was higher in the HG and HG+R568 groups but was
significantly lower in the HG+Calhex231 group (Fig. 4C).
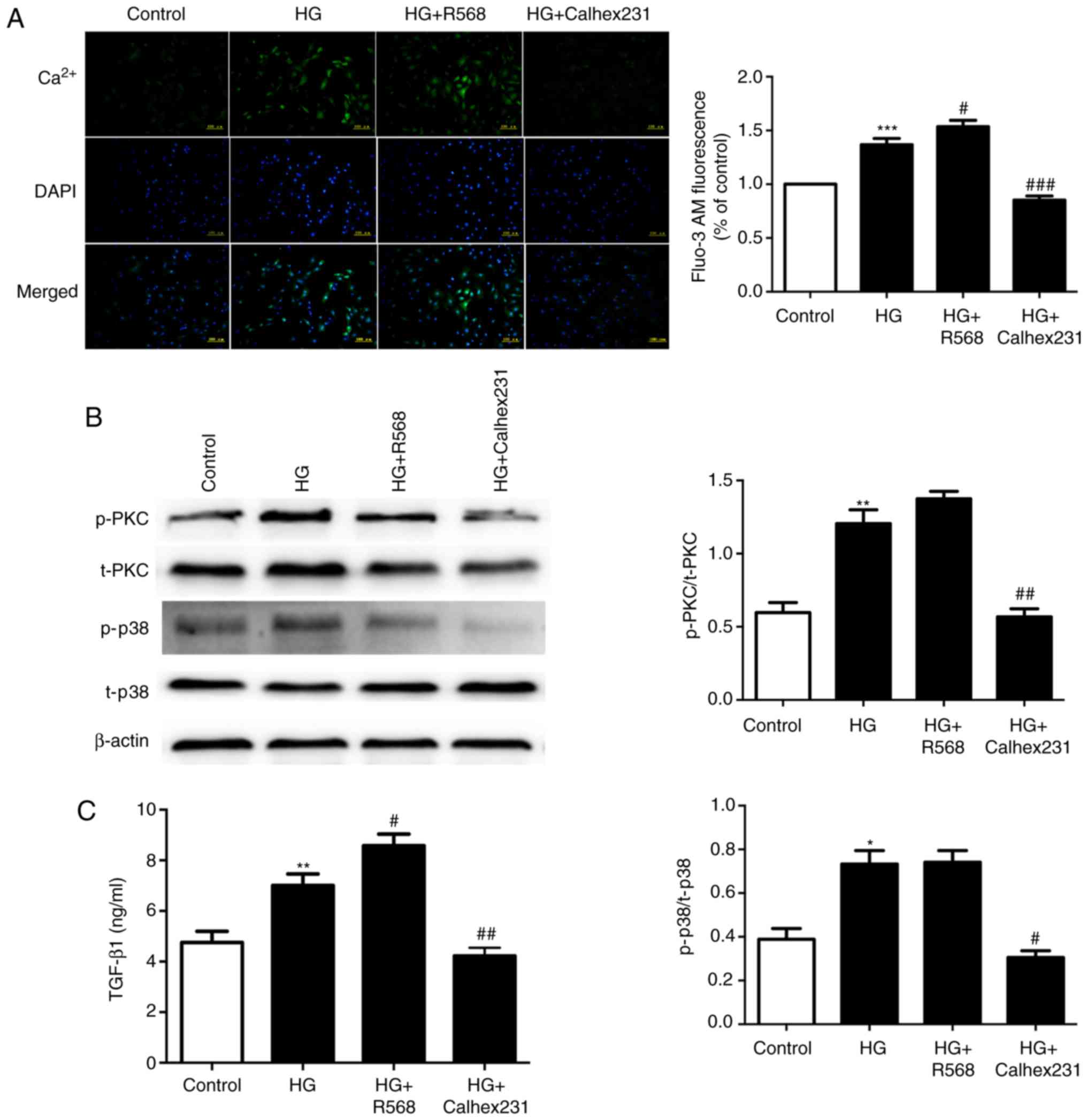 | Figure 4.Measurement of intracellular
Ca2+ in CFs, the level of p-PKC/p38 and the expression
of TGF-β1 in the culture medium. CFs were cultured for
24 h at 37°C in a control group (5.5 mM), HG group (40 mM), HG+R568
(5 µM) group and HG+Calhex231 (3 µM) group. (A) Cytosolic
Ca2+ was stained with Fluo-3/AM and assessed by
measuring the fluorescence intensity. (B) The protein levels of
p-PKC and p-p38 were evaluated by western blot analysis. (C) The
expression of TGF-β1 in the culture supernatants of the
CFs was determined using ELISA. *P<0.05, **P<0.01,
***P<0.001 vs. the Control; #P<0.05,
##P<0.01, ###P<0.001 vs. HG (n≥16).
TGF, transforming growth factor; p-PKC, phosphorylated protein
kinase C; HG, high glucose; CFs, cardiac fibroblasts. |
Signaling pathways associated with
myocardial fibrosis
Western blot analysis indicated that the protein
levels of TGF-β1, TβRI, TβRII and p-Smad2 were
significantly upregulated in the HG and R568 groups, and were
markedly downregulated in the HG+Calhex231 and HG+R568+LY2109761
groups compared to the HG and R568 groups, except for the
expression of TGF-β1 in the LY2109761 group (Fig. 5).
 | Figure 5.Effects of TβRI/II kinase inhibitor
(LY2109761) on TGF-β1/Smads pathway in CFs. CFs were
cultured for 48 h at 37°C in an HG group (40 mM), R568 (5 µM),
HG+Calhex231 (3 µM) and HG+R568+LY2109761 (20 µM) group.
Representative western blots of TGF-β1, TβRI, TβRII in
comparison with β-actin expression and p-Smad2 in comparison with
t-Smad2 expression in CFs are provided. **P<0.01, ***P<0.001,
****P<0.0001 vs. the Control; #P<0.05,
##P<0.01, ###P<0.001 vs. HG;
@@P<0.01 vs. R568 (n≥3). HG, high glucose; p-/t-Smad,
phosphorylated/total Smad; TGF, transforming growth factor; TβRI,
TGF-β-type I receptor; CFs, cardiac fibroblasts. |
Effects of TGF-β1 on
extracellular matrix (ECM) of CFs
Since excessive ECM is the major cause of myocardial
fibrosis, the changes in the mRNA and the protein levels of Col-I,
Col-III, MMP-2 and MMP-9 after treatment with HG (40 mmol) and
TGF-β1 (0, 5, 10 or 20 ng/ml) were assessed (Fig. 6A and B). Furthermore, the release
of Col-I, Col-III, MMP-2 and MMP-9 was determined by ELISA. HG and
TGF-β1 (5, 10 and 20 ng/ml) stimulation caused a
significant increase in the production of these factors by CFs
compared to the control and HG groups (Fig. 6C).
 | Figure 6.Effects of TGF-β1 in
culture medium on the changes of extracellular matrix components.
Cardiac fibroblasts were treated with HG (40 mM) in the presence or
absence of TGF-β1 (5, 10, 20 ng/ml) for 48 h at 37°C.
(A) After different treatments, the protein expression of Col-I,
Col-III, MMP-2 and MMP-9 was evaluated by western blot analysis.
(B) The changes in the mRNA levels of Col-I, Col-III, MMP-2 and
MMP-9 were measured by RT-qPCR, and the mRNA levels were normalized
to β-actin, which was used as the housekeeping gene. (C) The cell
supernatants were collected for the determination of Col-I,
Col-III, MMP2 and MMP9 expression using ELISA. *P<0.05,
**P<0.01 vs. the Control; #P<0.05,
##P<0.01, ###P<0.001 vs. HG (n≥8). Col,
collagen; SMA, smooth muscle actin; MMP, matrix metallopeptidase;
TGF, transforming growth factor; HG, high glucose. |
Discussion
Diabetes is a metabolic disease characterized by
hyperglycemia due to impaired insulin secretion or insulin
resistance. Persistent hyperglycemia and metabolic disorders may
lead to the impairment of tissues and organs, particularly the
cardiovascular system, nervous system and kidneys. DCM is a heart
disease independent of congenital heart disease, coronary heart
disease, and valvar heart disease, and is also a significant cause
of the increased mortality in patients with diabetes (14).
Myocardial tissue mainly consists of cardiomyocytes
and non-cardiomyocytes. CFs account for 90% of the total
non-cardiomyocytes and are not only the structural scaffolds of the
heart, but also link myocardial cells, endothelial cells and blood
vessels (15). CFs are involved in
maintaining homeostasis and the remodeling of ECM,
electrophysiological activity and the production of cell growth
factors (16). Col-I and -III are
two major components of the ECM, and have an important role in
maintaining the structure and function of the heart (17).
According to the existing literature, HG levels may
stimulate the proliferation of fibroblasts, promote myofibroblast
trans-differentiation and activate the transcription and secretion
of ECM proteins via the activation of angiotensin II, TGF-β, the
extracellular signal-regulated kinase signaling pathway and
reactive oxygen species production in vitro (18–20).
However, these previous studies did not yield any conclusive
evidence, and the precise mechanisms of hyperglycemia in the
remodeling and fibrosis of the diabetic heart still remain
elusive.
To further elucidate the role of CaSR in myocardial
fibrosis in DCM, a rat model of T1D was generated. Polydipsia,
polyuria, evident emaciation, increased blood glucose, TC and TG,
and decreased insulin activity were observed in rats treated with
STZ, and optionally with R568 or Calhex231, thus indicating that
the T1D rat model had been successfully generated.
At 12 weeks after modeling, the HW/BW was
significantly increased in the T1D group and the T1D+R568 group,
which may have been associated with the weight loss and an increase
in the myocardial ECM. This speculation is supported by the results
of the analysis of cardiac morphology and determination of
associated proteins. H&E staining indicated that the cardiac
myocytes of T1D rats were disordered and hypertrophic. Masson
staining and Sirius red staining revealed large amounts of Col
deposition in the interstitial and perivascular areas, particularly
in denatured and necrotic areas, while the CaSR agonist and the
CaSR inhibitor respectively promoted and inhibited these changes.
The expression of Col-I and Col-III proteins in the myocardial
tissue was significantly increased in the T1D group and the
T1D+R568 group, but was significantly decreased in the
T1D+Calhex321 group. These results demonstrated that myocardial
remodeling and myocardial fibrosis had clearly occurred in the T1D
rats and CaSR may be associated with the increased ECM and
deposition of Col.
It is well known that the proliferation and
activation of CFs represent the major pathways for Col secretion
and the increased ECM (21). A
previous study by our group indicated that CaSR is expressed in CFs
(22). However, the association of
changes of CaSR expression in CFs in DCM has remained to be fully
elucidated. To address this question, a series of experiments was
performed.
In the present study, CCK-8 assays indicated that HG
treatment increased the proliferation of CFs. It was also observed
that R568 further promoted the proliferation of CFs; however,
Calhex231 significantly inhibited these changes. This indicated
that CaSR is closely associated with the proliferation of CFs.
The proliferation and activation of CFs, as well as
the increased ECM, are important mechanisms of myocardial fibrosis
(23). Intracellular calcium is an
important second messenger and the driving force of CF activation
(24,25). Studies by our group (26) and other research groups (12,27)
have demonstrated that the increase or activation of CaSR
expression increases intracellular calcium through the G
protein/phospholipase C/inositol triphosphate pathway. To
investigate the role of CaSR in the activation of CFs, the effect
of HG treatment on intracellular calcium and a cell scratch assay
were determined. The results of the Fluo-3/AM fluorescence probe
analyses and cell scratch assay indicated that HG increased
intracellular calcium release and the migration of CFs.
Furthermore, R568 or Calhex231 promoted or inhibited these changes,
respectively. It is therefore evident that CaSR activation in CFs
promotes the proliferation and migration of CFs.
MMPs participate in the degradation of various
protein components of the ECM. Different types of MMPs degrade
different types of protein; MMP2 mainly degrades Col-IV, while MMP9
breaks down laminin and fibronectin (28). The present study indicated that HG
conditions and exogenous TGF-β1 activated CFs and
upregulated the expression of MMP2 and MMP9. The upregulated MMP2
and MMP9 provided additional ECM space for cell migration and the
secretion of Col-I and Col-III through the degradation of laminin
and fibronectin.
TGF-β1 is a potent cytokine with a
driving role in development, fibrosis and cancer (29). It promotes differentiation of CFs
and activation of the renin angiotensin aldosterone system, and
causes an increased abundance of NADPH (30). Furthermore, accumulation of
intracellular Ca2+, which promotes mitosis, induces cell
proliferation (25,29). More importantly, it promotes the
secretion of TGF-β1 modulated by mitogen-activated
protein kinase family members in CFs (30,31).
In the present study, HG and HG + R568 increased the levels of
p-PKC, p-p38 and the content of TGF-β1 in CFs and their
culture medium, while Calhex231 caused a significant reduction. To
further verify the role of TGF-β1, CFs were treated with
TβRI/II kinase inhibitor, and it was revealed that the expression
of TβRI/II and Smad2 was downregulated, while TGF-β1 was
not affected; this indicated the regulatory role of CaSR through
alteration of the intracellular calcium concentration. The effect
of increased TGF-β1 caused by CaSR was further assessed,
and the results demonstrated increased mRNA levels of Col-I,
Col-III, MMP-2 and MMP-9 and enhanced protein expression as well as
release of the relevant proteins into the culture medium.
It has previously been reported that the occurrence
and development of myocardial fibrosis is closely associated with
the activation of the TGF-β1/Smad pathway (32). TGF-β1 is associated with
its receptor TβRII, which activates TβRI kinase, causing the
phosphorylation of Smad2/3, which then combines with Smad4 and
forms a complex. This complex then translocates to the nucleus and
regulates the transcription of target genes, including Smad7. Smad7
is an inhibitory Smad, which degrades Smad2, Smad3 and
TGF-β1 via the ubiquitin protease degradation system
(33). The present study indicated
that HG levels and CaSR agonists significantly increased
TGF-β1 and p-Smad2, while CaSR inhibitors exerted the
opposite effects.
Collectively, based on the aforementioned
experimental results and previous studies, it may be hypothesized
that during diabetes (hyperglycemia), the upregulated expression of
CaSR in the CFs may lead to increases in intracellular
Ca2+ (a second messenger) (34), and further activate the
TGF-β1/Smads pathway. This results in the proliferation
and activation of fibroblasts, and eventually leads to myocardial
fibrosis (Fig. 7). Future studies
by our group will further clarify the role and specific mechanisms
of CaSR in myocardial fibrosis, and provide novel targets and an
experimental basis for the prevention and treatment of DCM.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (no. 81800260) and the Heilongjiang
Postdoctoral Fund (no. LBH-Z17103).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CX and HL conceived and supervised the study. HY and
YW designed experiments. HY, YF, BZ and YS performed experiments.
TG, CW and HY analyzed the data. HY drafted the manuscript. All
authors reviewed the results and approved the final version of the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
The animal raising and handling procedures were
performed in accordance with the Guide for the Care and Use of
Laboratory The study was approved by the Harbin Medical University
Medical Science Ethics Committee (Harbin, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MF
|
myocardial fibrosis
|
|
CaSR
|
calcium-sensing receptor
|
|
DCM
|
diabetic cardiomyopathy
|
|
HG
|
high glucose
|
|
MMP2/9
|
matrix metalloproteinase2/9
|
|
α-SMA
|
α-smooth muscle actin
|
|
TGF-β1
|
transforming growth factor
β1
|
|
STZ
|
streptozotocin
|
|
CFs
|
cardiac fibroblasts
|
|
TC
|
total cholesterol
|
|
H&E
|
hematoxylin and eosin
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
CCK-8
|
Cell Counting Kit-8
|
|
EdU
|
5-ethynyl-2′-deoxyuridine
|
References
|
1
|
Echouffo-Tcheugui JB and Dagogo-Jack S:
Preventing diabetes mellitus in developing countries. Nat Rev
Endocrinol. 8:557–562. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang X, McLennan SV, Allen TJ, Tsoutsman
T, Semsarian C and Twigg SM: Adverse effects of high glucose and
free fatty acid on cardiomyocytes are mediated by connective tissue
growth factor. Am J Physiol Cell Physiol. 297:C1490–C1500. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Westermeier F, Riquelme JA, Pavez M,
Garrido V, Díaz A, Verdejo HE, Castro PF, García L and Lavandero S:
New Molecular Insights of Insulin in Diabetic Cardiomyopathy. Front
Physiol. 7:1252016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fowlkes V, Clark J, Fix C, Law BA, Morales
MO, Qiao X, Ako-Asare K, Goldsmith JG, Carver W, Murray DB, et al:
Type II diabetes promotes a myofibroblast phenotype in cardiac
fibroblasts. Life Sci. 92:669–676. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cavalera M, Wang J and Frangogiannis NG:
Obesity, metabolic dysfunction, and cardiac fibrosis:
Pathophysiological pathways, molecular mechanisms, and therapeutic
opportunities. Transl Res. 164:323–335. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hutchinson KR, Lord CK, West TA and
Stewart JA Jr: Cardiac fibroblast-dependent extracellular matrix
accumulation is associated with diastolic stiffness in type 2
diabetes. PLoS One. 8:e720802013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tharmalingam S and Hampson DR: The
Calcium-Sensing Receptor and Integrins in Cellular Differentiation
and Migration. Front Physiol. 7:1902016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hendy GN and Canaff L: Calcium-Sensing
Receptor Gene: Regulation of Expression. Front Physiol. 7:3942016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Peng X, Li HX, Shao HJ, Li GW, Sun J, Xi
YH, Li HZ, Wang XY, Wang LN, Bai SZ, et al: Involvement of
calcium-sensing receptors in hypoxia-induced vascular remodeling
and pulmonary hypertension by promoting phenotypic modulation of
small pulmonary arteries. Mol Cell Biochem. 396:87–98. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu C, Zhang W, Jiang C, Sun Y and Wang R:
Involvement of calcium sensing receptor in myocardial
ischemia/reperfusion injury and apoptosis. J Mol Cell Cardiol.
42:S80–S81. 2007. View Article : Google Scholar
|
|
11
|
Wang Y, Gao P, Wei C, Li H, Zhang L, Zhao
Y, Wu B, Tian Y, Zhang W, Wu L, et al: Calcium sensing receptor
protects high glucose-induced energy metabolism disorder via
blocking gp78-ubiquitin proteasome pathway. Cell Death Dis.
8:e27992017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dong S, Li G, Zheng D, Wu J, Sun D, Yang
F, Yu X, Li T, Sun A, Liu J, et al: A novel role for the calcium
sensing receptor in rat diabetic encephalopathy. Cell Physiol
Biochem. 35:38–50. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liang CC, Park AY and Guan JL: In vitro
scratch assay: A convenient and inexpensive method for analysis of
cell migration in vitro. Nat Protoc. 2:329–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Adeghate E: Molecular and cellular basis
of the aetiology and management of diabetic cardiomyopathy: A short
review. Mol Cell Biochem. 261:187–191. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lam S, Verhagen NAM, Strutz F, van der
Pijl JW, Daha MR and van Kooten C: Glucose-induced fibronectin and
collagen type III expression in renal fibroblasts can occur
independent of TGF-β1. Kidney Int. 63:878–888. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Spector KS: Diabetic cardiomyopathy. Clin
Cardiol. 21:885–887. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kehlet SN, Willumsen N, Armbrecht G,
Dietzel R, Brix S, Henriksen K and Karsdal MA: Age-related collagen
turnover of the interstitial matrix and basement membrane:
Implications of age- and sex-dependent remodeling of the
extracellular matrix. PLoS One. 13:e01944582018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Russo I and Frangogiannis NG:
Diabetes-associated cardiac fibrosis: Cellular effectors, molecular
mechanisms and therapeutic opportunities. J Mol Cell Cardiol.
90:84–93. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Loboda A, Sobczak M, Jozkowicz A and Dulak
J: TGF-β1/Smads and miR-21 in Renal Fibrosis and Inflammation.
Mediators Inflamm. 2016:83192832016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yao M, Wang X, Wang X, Zhang T, Chi Y and
Gao F: The Notch pathway mediates the angiotensin II-induced
synthesis of extracellular matrix components in podocytes. Int J
Mol Med. 36:294–300. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Olson ER: Signaling mechanisms controlling
the proliferation and differentiation of cardiac fibroblasts
(unpublished PhD thesis). Kent State University, College of
Biomedical Sciences. 2006.
|
|
22
|
Zhang X, Zhang T, Wu J, Yu X, Zheng D,
Yang F, Li T, Wang L, Zhao Y, Dong S, et al: Calcium sensing
receptor promotes cardiac fibroblast proliferation and
extracellular matrix secretion. Cell Physiol Biochem. 33:557–568.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Park S, Ranjbarvaziri S, Lay FD, Zhao P,
Miller MJ, Dhaliwal JS, Huertas-Vazquez A, Wu X, Qiao R, Soffer JM,
et al: Genetic Regulation of Fibroblast Activation and
Proliferation in Cardiac Fibrosis. Circulation. 138:1224–1235.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Du G, Fischer BE, Voss KO, Becker G,
Taucher-Scholz G, Kraft G and Thiel G: The absence of an early
calcium response to heavy-ion radiation in Mammalian cells. Radiat
Res. 170:316–326. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu W, Wang X, Mei Z, Gong J, Huang L, Gao
X, Zhao Y, Ma J and Qian L: BNIP3L promotes cardiac fibrosis in
cardiac fibroblasts through [Ca2+]i-TGF-β-Smad2/3 pathway. Sci Rep.
7:19062017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang WH, Fu SB, Lu FH, Wu B, Gong DM, Pan
ZW, Lv YJ, Zhao YJ, Li QF, Wang R, et al: Involvement of
calcium-sensing receptor in ischemia/reperfusion-induced apoptosis
in rat cardiomyocytes. Biochem Biophys Res Commun. 347:872–881.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li GW, Miao HZ, Bo L, Wang GZ, Jin L, Lin
Y, Deng ZH and Xiao W: Calcium-sensing receptor modulates pulmonary
artery tension through G-protein-PLC-IP_3 pathways. Chin J
Pathophysiol. 31:2015.(In Chinese).
|
|
28
|
Jabłońska-Trypuć A, Matejczyk M and
Rosochacki S: Matrix metalloproteinases (MMPs), the main
extracellular matrix (ECM) enzymes in collagen degradation, as a
target for anticancer drugs. J Enzyme Inhib Med Chem. 31 (Suppl
1):177–183. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nüchel J, Ghatak S, Zuk AV, Illerhaus A,
Mörgelin M, Schönborn K, Blumbach K, Wickström SA, Krieg T, Sengle
G, et al: TGFB1 is secreted through an unconventional pathway
dependent on the autophagic machinery and cytoskeletal regulators.
Autophagy. 14:465–486. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wei Y, Meng T and Sun C: Protective effect
of diltiazem on myocardial ischemic rats induced by isoproterenol.
Mol Med Rep. 17:495–501. 2018.PubMed/NCBI
|
|
31
|
Ryu JM, Lee MY, Yun SP and Han HJ: High
glucose regulates cyclin D1/E of human mesenchymal stem cells
through TGF-beta1 expression via Ca2+/PKC/MAPKs and PI3K/Akt/mTOR
signal pathways. J Cell Physiol. 224:59–70. 2010.PubMed/NCBI
|
|
32
|
Shi BH, Zong-Pei XU and Fan GW: The
research progress on treatment of myocardial fibrosis by regulating
TGF-β1/Smads pathway. Zhongguo Yaolixue Tongbao. (In Press).
|
|
33
|
Lan HY: Diverse roles of TGF-β/Smads in
renal fibrosis and inflammation. Int J Biol Sci. 7:1056–1067. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Duran J, Troncoso MF, Lagos D, Ramos S,
Marin G and Estrada M: GDF11 Modulates Ca2+-Dependent Smad2/3
Signaling to Prevent Cardiomyocyte Hypertrophy. Int J Mol Sci.
19:1333–1342. 2018. View Article : Google Scholar :
|
















