Introduction
Post-operative cognitive dysfunction (POCD) is a
complication of the central nervous system after surgery that
manifests as mental disorder, anxiety, personality changes and
memory impairment (1,2). A number of risk factors are
associated with the POCD including age, preoperative cognition and
emotional state (3). A previous
study noted that older men with APOE4 allele may be more vulnerable
to POCD than older women with APOE4 allele (4). After coronary artery bypass grafting
(CABG), the incidence of early cognitive dysfunction is high, at
30–60%, and POCD severely affects the post-operative quality of
life of patients (5–7). Currently, the diagnosis of POCD is
based on the description of symptoms from the patient, assessment
of mental status and evaluation of clinical behaviors, which are
complex and take time to evaluate (8,9).
Although some biomarkers have been found to be associated with POCD
such as S100B and NSE, the mechanism and genes which associated
with these biomarkers remain to be elucidated (10).
Circular (circ) RNAs have recently emerged as a
novel class of transcripts. circRNAs are noncoding RNA (ncRNA)
molecules that regulate the stability or translational efficiency
of target mRNAs through a competitive endogenous RNA (ceRNA)
mechanism (11). The mammalian
brain is the organ in which circRNAs are more abundant and first
evidence of their functional significance started to emerge among
many tissues and species (12).
Several studies have reported that circRNAs are important
regulatory factors in normal developmental, physiological and
disease states, including cancer, mental disorders and
cardiovascular diseases (13–15).
However, to the best of our knowledge, no circRNAs have been
reported to be associated with POCD.
Unlike tumor tissue, some tissues are not easy to
obtain in clinical practice, including from the heart and brain.
Recent studies have found that diseased tissues from the heart or
brain can release exosomes, containing miRNAs, long non-coding RNAs
(lncRNAs) and circRNAs, and that these exosomes can cross the
blood-brain barrier and enter the circulation. These ncRNAs may be
new biological markers that can be obtained with less invasive
procedures from patients with cardiovascular and cerebral disease
(16,17).
This present study aimed to identify circRNAs in the
plasma exosomes of patients with POCD. These circRNAs were probed
as potential specific biomarkers for the diagnosis of POCD.
Materials and methods
Patients
The present study was approved by the Medical Ethics
Committee of The Affiliated Beijing Chaoyang Hospital of Capital
Medical University. Written informed consent was obtained from each
participant before enrollment. Patients, between 40 and 90 years of
age, who were scheduled for first-time elective off-pump CABG
surgery were recruited between June 2018 and October 2018.
Surgeries were performed at The Affiliated Chaoyang Hospital of
Capital Medical University. The exclusion criteria included
pre-existing neurological disease, including a Mini-Mental State
Examination (MMSE) score <24 points; a history of taking
antidepressants or anti-anxiety drugs; anticipated difficulty with
neuropsychological assessment, including severe visual or hearing
impairment; deep hypothermic circulatory arrest during surgery;
perioperative insulin therapy; and poor ventricular function
(ejection fraction <30%).
Anesthesia protocol
Patients were given standard perioperative care and
surgery was performed by the same team at The Affiliated Beijing
Chaoyang Hospital of Capital Medical University. Anesthesia was
administered according to a standard protocol, including induction
with midazolam (0.05–0.1 mg/kg), fentanyl (5–8 µg/kg), etomidate
(0.2–0.3 mg/kg) and vecuronium (1 mg/kg), and maintenance with
fentanyl (0.1–0.15 µg/kg/min), midazolam (0.6–1 µg/kg/min),
sevoflurane (0.5–1%) in oxygen and vecuronium (0.05 mg/kg, once
every 30 min). Standard monitoring was performed for all patients,
including electrocardiography, pulse oximetry, end-tidal carbon
dioxide, nasopharyngeal temperatures, central venous and arterial
blood pressure, arterial blood gas, cardiac output and bispectral
index.
Neuropsychological assessment
Patients were first screened with the MMSE to
exclude severe cognitive impairment. The enrolled patients
completed neuropsychological tests 1 day before surgery and again 1
week after surgery. The neuropsychological evaluation test applied
in this present study was based on a Chinese version of
neuropsychological testing (18).
The series of tests assessed several cognitive functions, including
attention, motor skills, executive function, learning and memory.
Concentration and the ability to ignore distracting stimuli was
assessed with the Stroop Colour and Word Test (SCWT) (19), including SCWT-1, SCWT-2 and SCWT-3.
The time required to complete the tests and the number of errors
was recorded. Mental processing speed and visual scanning with the
Trail Making Test (TMT) (20),
including TMT-A and TMA-B, were measured and the time required to
complete these tests was recorded. Concentration, processing speed
and visual scanning abilities were tested with the Digit Symbol
Substitution Test (21), and the
correct matching quantity was measured. Short-term and long-term
memory, and attention, were assessed by the Verbal Learning Test
(VLT) (22), including VLT-A1,
VLT-A2 and VLTA3. The total number of correct words immediately
recalled and recalled after a 30 min delay were recorded. The
Symbol Digits Modalities Test (SDMT) (23) was used to measure short-term
memory, visuospatial skills and attention, and the number of errors
recorded. These tests have been used to evaluate the POCD in a
previous study (10). POCD was
defined by a reduction in the post-operative test score compared
with the pre-operative test score ≥20% in at least two of the eight
tests, following the same criteria established by the previous
study (10).
Data collection
Demographic and intraoperative data, including age,
weight, sex, body mass index (BMI), ejection fraction (EF), New
York Heart Association (NYHA) (24) class of heart failure and surgical
time, were collected. Blood samples (10 ml) were collected 1 day
before anesthesia and 7 days after surgery (at 08:00 a.m. following
overnight fasting). Samples were collected in anticoagulant tubes,
centrifuged at 4°C at 300 × g for 5 min and then at 1,200 × g and
4°C for 20 min. The plasma was stored at −80°C until further
analysis.
Exosome isolation from plasma
The ultracentrifugation exosome isolation protocol
was used to isolate exosomes from plasma. Briefly, plasma samples
were centrifuged at 500 × g and 4°C for 5 min to remove the cells
from the samples. The supernatant was transferred to a new
polycarbonate tube and centrifuged at 2,000 × g for 10 min at 4°C.
The supernatant was collected and transferred to a new
polycarbonate tube. The samples were centrifuged at 10,000 × g for
30 min at 4°C to eliminate shed microvesicles (200–1,000 nm). The
supernatants were collected and filtered using a 0.22 µm membrane
filter (EMD Millipore). The samples were centrifuged at 100,000 × g
for 2 h and 4°C. For RNA isolation, the exosome pellet was washed
once with 1X PBS, followed by centrifugation 100,000 × g for 2 h at
4°C. The exosomes were resuspended in 1X PBS and stored at −80°C
for further use.
Electron microscopy
Exosomes isolated from plasma were washed in PBS to
further purify the sample, filtered using a 0.22-µm membrane filter
(EMD Millipore), and ultracentrifuged at 100,000 × g for 2 h at 4°C
to re-pellet the exosomes. The exosome pellet was resuspended and
fixed in PBS containing 2% glutaraldehyde (4°C for 5 min) and then
loaded onto former/carbon-coated electron microscopy grids. The
samples were contrasted with 0.5% uranyl acetate to visualize the
membranes and the samples were viewed with a Tecnai G2 Spirit 120
KV electron microscope (Thermo Fischer Scientific, Inc.).
circRNA microarray analysis
TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) and a RNeasy mini kit (74104; Qiagen,
GmbH) were used for total RNA extraction according to the
manufacturers' protocol. RNA quality and quantity were determined
using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies;
Thermo Fisher Scientific, Inc.). Samples with an absorbance ratio
>1.8 at 260 and 280 nm (A260/280), and at 260 and 230 nm
(A260/230), an RNA concentration >60 ng and a total quantity
>800 ng were defined as having sufficient quality and quantity
for further experiments. Sample preparation and microarray
hybridization was performed based on the standard protocol from
Arraystar, Inc. Briefly, total RNA was digested with RNase R
(Epicentre; Illumina, Inc.) to remove linear RNAs and enrich for
circRNAs. The enriched circular RNAs were amplified and transcribed
into fluorescent complementary RNA (cRNA) utilizing a random
priming method (Arraystar Super RNA Labeling kit; Arraystar, Inc.).
The labeled cRNAs were hybridized onto the Arraystar Rat circRNA
Array (8×15K; Arraystar, Inc.). After washing the slides, the
arrays were scanned with the Agilent G2505C Scanner (Agilent
Technologies, Inc.). Agilent Feature Extraction software (version
11.0.1.1; Agilent Technologies, Inc.) was used to analyze the
acquired array images. Quantile normalization and subsequent data
processing was performed using the R software (3.4.2) limma package
(v3.22.7, http://bioinf.wehi.edu.au/limma/). Differentially
expressed circRNAs with statistical significance between patients
with POCD and healthy individuals were identified through volcano
plot filtering (25).
Differentially expressed circRNAs between the two groups were
identified through fold change filtering. Hierarchical clustering
was performed to show the circRNA expression pattern among samples.
circRNAs with a relative up- or downregulation of at least 1.5-fold
signal density and P<0.05 were considered to be differentially
expressed.
Reverse transcription-quantitative
RT-q(PCR) validation of differentially expressed circRNAs
The expression levels of different circRNAs from the
circRNA microarray analysis were validated using RT-qPCR.
Complementary DNA templates were generated from 800 ng of total RNA
and used for RT-qPCR. β-actin was used as an internal reference for
template normalization. The primers for the different circRNAs were
designed using Primer software (5.0, Premier, http://www.premierbiosoft.com/primerdesign; Table I). The ViiA 7 Real-time PCR System
(Applied Biosystems; Thermo Fisher Scientific, Inc.) was used for
the amplification and detection of the circRNAs. The standard curve
was prepared using a gradient dilution of the DNA template.
Briefly, cDNA templates expressing the target and housekeeper genes
were selected for the RT-qPCR reaction as follows: 95°C for 10 min,
followed by 40 cycles of 95°C for 10 sec and 60°C for 60 sec. The
amplified PCR product and a 100-bp DNA ladder were electrophoresed
on a 2% agarose gel and single specific bands were detected by
ethidium bromide staining. After purification of the DNA of
interest from the gel, a gradient dilution (10 dilutions, each
decreasing by a factor of 10) was prepared and a standard curve was
generated for the RT-qPCR assay. For RT-qPCR of the experimental
groups, the target and housekeeping genes of each sample were
subjected to the aforementioned RT-qPCR reaction. The
concentrations of the target and housekeeping genes in each sample
were directly generated by the machine using the standard curve as
a reference. The relative level of the target gene=concentration of
target gene/concentration of housekeeping gene.
 | Table I.Sequences for all primers used. |
Table I.
Sequences for all primers used.
| Gene name | Bidirectional
primer sequence | Annealing
temperature (°C) | Product length
(bp) |
|---|
| β-actin (H) | F:
5′-GTGGCCGAGGACTTTGATTG-3′ | 60 | 73 |
|
| R:
5′-CCTGTAACAACGCATCTCATATT-3′ |
|
|
|
hsa_circRNA_038651 | F:
5′-TACATCCAGGCCCACATCG-3′ | 60 | 80 |
|
| R:
5′-TTCATCACGCAGCGCTTG-3′ |
|
|
|
hsa_circRNA_017502 | F:
5′-GCATCCGAACTGTCCCCTAC-3′ | 60 | 107 |
|
| R:
5′-ACCTGGCTCCTGTGTCATCAT-3′ |
|
|
|
hsa_circRNA_089763 | F:
5′-GGTGGGCCATACGGTAGTATT-3′ | 60 | 88 |
|
| R:
5′-TTACTTCCTCTCTTTCTTCTTCCC-3′ |
|
|
|
hsa_circRNA_091840 | F:
5′-TGTCTTCTTTGGTTCTGGGAG-3′ | 60 | 131 |
|
| R:
5′-CAATCTGAATGGTGGGGC-3′ |
|
|
|
hsa_circRNA_049784 | F:
5′-CATCCCTGGCCGGTTCAA-3′ | 60 | 70 |
|
| R:
5′-AGTGGGGGTTTCTCTGTTTCCT-3′ |
|
|
|
hsa_circRNA_000798 | F:
5′-ACTTGGGGTTTCTGTGGTTC-3′ | 60 | 70 |
|
| R:
5′-GAGCTCATGCCCTATGAGGA-3′ |
|
|
|
hsa_circRNA_101688-1 | F:
5′-CTTATCCCCAGAGAGCAAACAA-3′ | 60 | 69 |
|
| R:
5′-GGCAGGAACATAACACCACG-3′ |
|
|
|
hsa_circRNA_101688-2 | F:
5′-CAGATCACAGATTTTGGAACAGC-3′ | 60 | 131 |
|
| R:
5′-ACTCAGTCTGGGTCCTCACCAT-3′ |
|
|
|
hsa_circRNA_002281-1 | F:
5′-CCAGGTCATAGTAATGGGAGC-3′ | 60 | 140 |
|
| R:
5′-GCTGTAGCTTCATGGTCAGAA-3′ |
|
|
|
hsa_circRNA_002281-2 | F:
5′-GCCAGGTCATAGTAATGGGAG-3′ | 60 | 118 |
|
| R:
5′-TGAACGGTTTCGTAGAAAGGA-3′ |
|
|
|
hsa_circRNA_100075 | F:
5′-CGATAACATTTAGCCTGGAACA-3′ | 60 | 74 |
|
| R:
5′-GTAGGTCTGATTGAAGCAAGCC-3′ |
|
|
|
hsa_circRNA_003558-1 | F:
5′-CTTCTCCCCAGCCAATGTCG-3′ | 60 | 140 |
|
| R:
5′-CTGAAGGTAGGTGGTGAATAGGG-3′ |
|
|
|
hsa_circRNA_003558-2 | F:
5′-TGGAGAGACAGTAAAGGAAAAGAC-3′ | 60 | 107 |
|
| R:
5′-GGTCAAGAATCTGAAGGTAGGTG-3′ |
|
|
|
hsa_circRNA_101006-1 | F:
5′-GACTGGAGCAAGGTCGTCCT-3′ | 60 | 104 |
|
| R:
5′-AGAGCATGGGCCACTTTCTG-3′ |
|
|
|
hsa_circRNA_101006-2 | F:
5′-GTACTGGCAAGACTGCAACACC-3′ | 60 | 84 |
|
| R:
5′-AGGCGATTACTCCGAGTCCC-3′ |
|
|
|
hsa_circRNA_031757-1 | F:
5′-GGCTTTTGGCTGATGAGGATT-3′ | 60 | 62 |
|
| R:
5′-TAGCTGCACTATTACGAAGGGAC-3′ |
|
|
|
hsa_circRNA_031757-2 | F:
5′-TGGCTGATGAGGATTGATGC-3′ | 60 | 125 |
|
| R:
5′-AGTCGAGCTTCATTGCAGAAT-3′ |
|
|
|
hsa_circRNA_004954-1 | F:
5′-GGGAGCCCTGAATATACACG-3′ | 60 | 114 |
|
| R:
5′-CACCACCCGTTTCATTTTTAC-3′ |
|
|
|
hsa_circRNA_004954-2 | F:
5′-ATACACGAGAAAGCCTGGAATG-3′ | 60 | 79 |
|
| R:
5′-GGCCTCAGCCACAGAATACAG-3′ |
|
|
|
hsa_circRNA_012969-1 | F:
5′-TTTGACGGTGGATTTGGTTG-3′ | 60 | 96 |
|
| R:
5′-GTCCGGTGCATCTGACTTGA-3′ |
|
|
|
hsa_circRNA_012969-2 | F:
5′-CAGGTTCTGAATCCCATGCT-3′ | 60 | 129 |
|
| R:
5′-CATCTTTCTTTGAGCCATAGGA-3′ |
|
|
Analysis of target genes
The miRDB V5 database (http://mirdb.org/miRDB/) was used to predict miRNA
target genes in the human, mouse, and rat genomes. An additional
database, targetscan7.1, was used for miRNA target predictions in
human (http://www.targetscan.org/vert_71/) and rat
(http://www.targetscan.org/mmu_71/).
The overlapping results of two databases for each of these species
are generally accepted.
Gene Ontology (GO) analysis
The GO project provides a controlled vocabulary to
describe gene and gene product attributes in any organism
(http://www.geneontology.org). The
ontology covers three domains: Biological process, cellular
component and molecular function. Fisher's exact test was used in
Bioconductor topGO software (version 2.32.0, http://www.bioconductor.org/packages/release/bioc/html/topGO.html)
to determine whether there was more overlap between the
differentially expressed (DE) list and the GO annotation list than
would be expected by chance. The P-value produced by topGO denotes
the significance of GO term enrichment in the DE genes. The lower
the P-value, the more significant the GO term (P≤0.05 is
recommended).
Pathway analysis
Pathway analysis is a functional analysis mapping of
genes to Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways.
The P-value (Expression Analysis Systemic Explorer-score, Fisher
P-value or Hypergeometric P-value) denotes the significance of the
pathway correlated to the conditions. The lower the P-value, the
more significant the pathway (the recommended P-value cut-off is
0.05).
Constructing the circRNA-miRNA-mRNA
co-expression network
The co-expression network of the circRNA- miRNA-mRNA
was constructed using the analysis of circRNA-miRNA and the
miRNA-gene networks. The related miRNA and mRNA of the DE circRNAs
were screened and the circRNA-miRNA-mRNA network was
constructed.
Statistical analysis
All statistical analyses were performed using SPSS
13.0 software (SPSS, Inc.). Normally distributed data are presented
as the mean ± SD and were analyzed using the paired or unpaired
t-test (including baseline analysis of age and operation time) or
repeated measure analysis of covariance. Categorical data are
expressed as percentages and frequencies, and were analyzed using
the Pearson's χ2 test. Pearson's correlation analysis
was used to illustrate the relationships among age, sex and weight.
Regression analysis was used to investigate the factors
contributing to the risk of POCD. P<0.05 was considered to
indicate a statistically significant difference.
Results
Patients included and perioperative
characteristics of patients
As shown in Fig. 1,
35 patients completed the cognitive tests and 12 patients (34.3%)
developed POCD 7 days after surgery. The peri-operative
characteristics of patients are shown in Table II. Patients were divided into two
groups according to ‘cognitive tests’: POCD (n=12) and non-POCD
(n=23). There were no statistically significant differences between
the two groups in terms of sex, height, weight, BMI, EF, NYHA class
of heart failure or surgical time (P>0.05). Patients with POCD
were older than patients without POCD (P<0.05).
| Table II.Characteristics of the patients. |
Table II.
Characteristics of the patients.
|
Characteristics | POCD group
(n=12) | Non-POCD group
(n=23) | P-value |
|---|
| Age, years | 67.67±8.50 | 60.78±8.95 | 0.04 |
| Female, n | 1/9 | 4/23 | 0.48 |
| Height, cm |
1.71±0.06 |
1.67±0.08 | 0.15 |
| Weight, kg | 74.33±7.41 |
71.65±12.93 | 0.52 |
| BMI,
kg/cm2 | 25.36±2.24 | 25.50±3.86 | 0.91 |
| EF, % | 57.75±7.30 |
57.74±10.90 | 0.99 |
| NYHA |
|
| 0.78 |
| II | 5 | 10 |
|
|
III | 5 | 11 |
|
| IV | 2 | 2 |
|
| Surgical time,
min | 266.67±43.99 | 238.04±51.54 | 0.12 |
Exosome extraction results
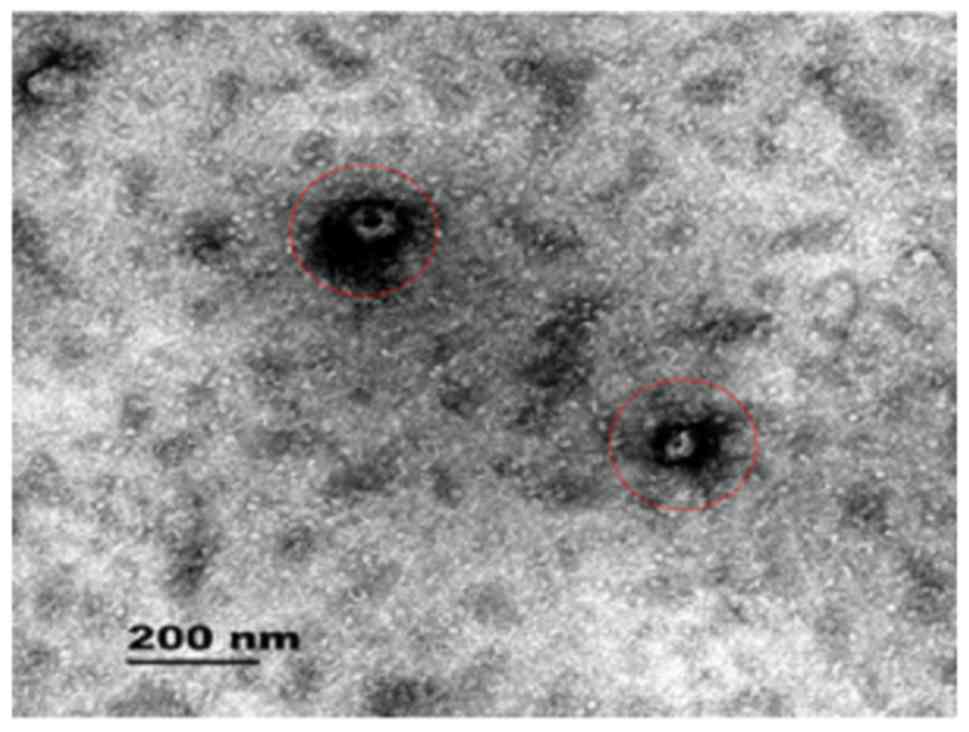
As shown in Fig. 2,
the red circle represents the plasma exosome. A transmission
electron microscope image was taken of an exosome; the image shows
the vesicular structure of the exocrine bodies and the size.
Expression profile of circRNAs in the
plasma exosomes of patients with POCD using microarray analysis and
secondary RT-qPCR validation
RNA concentration and purity was measured using a
NanoDrop ND-1000 spectrophotometer. Samples with an A260/280 and
A260/230 absorbance ratio >1.8, an RNA concentration >60 ng
and total quantity >800 ng, were defined as having sufficient
quality and quantity for further experimentation. The RNA purity,
concentration and total quantity of 8 of the 12 samples from
patients with POCD and 16 of the 23 samples from patients without
POCD met these requirements and were used for subsequent microarray
detection of the circRNA expression profile (3 samples from the
POCD and 3 from the non-POCD group were analyzed by microarray, and
5 from each group analyzed by RTqPCR). As shown in Table III, circRNA microarray analysis
of the plasma exosomes of patients with POCD compared with patients
without POCD (P<0.05) indicated that 9 circRNAs were upregulated
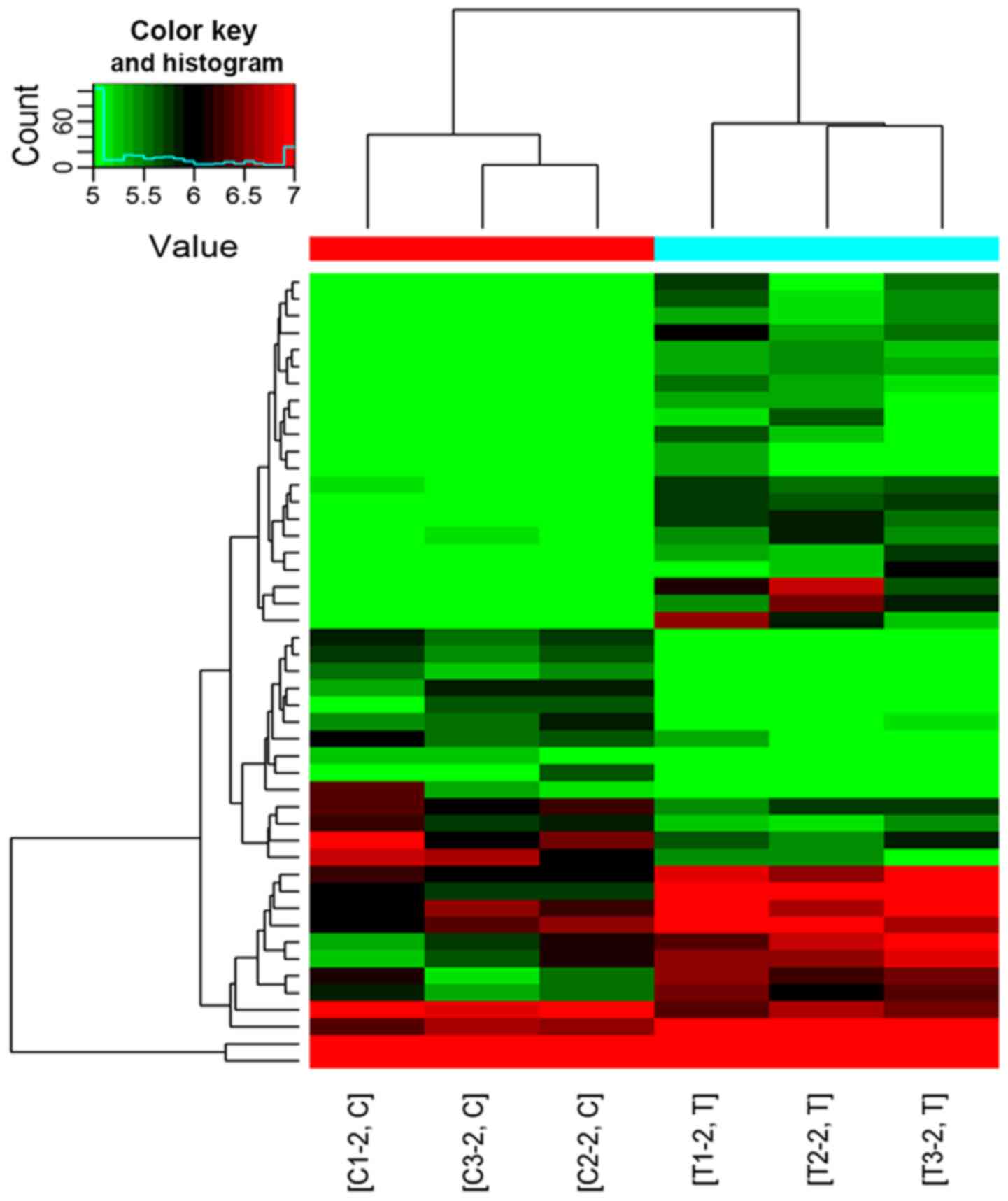
and 6 circRNAs were downregulated. As shown in Fig. 3, significantly different circRNAs
were selected for hierarchical clustering analysis. The color key
reflects the expression intensity of the circRNAs; with red
indicating high expression, green indicating low expression and the
color depth (ranging from black to color) indicating different
expression intensities. The circRNA hierarchical clustering map
revealed circRNA expression in all samples from both groups and
indicated that the expression of a single circRNA was altered in
all samples. RT-qPCR was used to validate the 15 circRNAs
identified by microarray analysis (fold change ≥1.5; P<0.05). As
shown in Table IV, of the 15
circRNAs analyzed by RT-qPCR only hsa_circRNA_089763 was confirmed
to be differentially expressed in the plasma exosomes of patients
with POCD compared to patients without POCD (P<0.05).
| Table III.Aberrantly expressed circRNAs in the
plasma exosomes of patients with post-operative cognitive
dysfunction from microarray analysis (fold change ≥1.5;
P<0.05). |
Table III.
Aberrantly expressed circRNAs in the
plasma exosomes of patients with post-operative cognitive
dysfunction from microarray analysis (fold change ≥1.5;
P<0.05).
| A, Upregulated |
|---|
|
|---|
| circRNA | Fold change | P-value | Normalized
intensity |
|---|
|
has_circRNA_038651 | 1.88 | 0.0407 | 10.04 |
|
has_circRNA_017502 | 1.71 | 0.0376 | 6.67 |
|
has_circRNA_089763 | 2.39 | 0.0415 | 7.16 |
|
has_circRNA_000798 | 3.39 | 0.0056 | 5.28 |
|
has_circRNA_102054 | 1.76 | 0.0038 | 4.90 |
|
has_circRNA_100075 | 1.60 | 0.0016 | 5.31 |
|
has_circRNA_101688 | 1.61 | 0.0059 | 4.99 |
|
has_circRNA_002281 | 2.10 | 0.0004 | 6.40 |
|
has_circRNA_003558 | 1.61 | 0.0048 | 5.35 |
|
| B,
Downregulated |
|
| circRNA | Fold
change | P-value | Normalized
intensity |
|
|
hsa_circRNA_012969 | 1.51 | 0.0042 | 5.12 |
|
hsa_circRNA_004954 | 1.89 | 0.0028 | 5.27 |
|
hsa_circRNA_101006 | 1.76 | 0.0036 | 5.21 |
|
hsa_circRNA_031757 | 1.86 | 0.0096 | 5.25 |
|
hsa_circRNA_091840 | 1.51 | 0.0193 | 6.78 |
|
hsa_circRNA_049784 | 1.61 | 0.0269 | 9.35 |
| Table IV.Expression profile of 15 circRNA in
the plasma exosomes of patients with post-operative cognitive
dysfunction using reverse transcription-quantitative PCR
validation. |
Table IV.
Expression profile of 15 circRNA in
the plasma exosomes of patients with post-operative cognitive
dysfunction using reverse transcription-quantitative PCR
validation.
| circRNA | Fold change | Regulation | P-value | Cq value |
|---|
|
hsa_circRNA_038651 | 6.1 | Up | 0.26321 | 34.0±0.8 |
|
hsa_circRNA_017502 | 1.5 | Up | 0.20196 | 35.0±0.7 |
|
hsa_circRNA_089763 | 2.1 | Up | 0.00055 | 34.7±0.7 |
|
hsa_circRNA_000798 | – | – | – | >40 |
|
hsa_circRNA_102054 | – | – | – | – |
|
hsa_circRNA_100075 | – | – | – | >40 |
|
hsa_circRNA_101688 | – | – | – | >40 |
|
hsa_circRNA_002281 | – | – | – | >40 |
|
hsa_circRNA_003558 | – | – | – | >40 |
|
hsa_circRNA_012969 | – | – | – | >40 |
|
hsa_circRNA_004954 | – | – | – | >40 |
|
hsa_circRNA_101006 | – | – | – | >40 |
|
hsa_circRNA_031757 | – | – | – | >40 |
|
hsa_circRNA_091840 | 0.8 | Down | 0.66827 | 35.7±0.6 |
|
hsa_circRNA_049784 | 0.8 | Down | 0.22230 | 32.1±0.3 |
Predicting the miRNAs regulated by
circRNA_089763 and their corresponding target genes
circRNAs act as miRNA sponges, binding and
inhibiting miRNAs from regulating their target genes, and are also
known as ceRNAs (11). As shown in
Fig. 4, circRNA_089763 may
sequester 10 miRNAs, including hsa-miR-670-3p, hsa-miR-6856-3p,
hsa-miR-6769b-3p, hsa-miR-7111-3p, hsa-miR-6803-3p, hsa-miR-614,
hsa-miR-6784-3p, hsa-miR-454-5p, hsa-miR-6798-3p and hsa-miR-3684.
Each of these miRNAs regulates target genes. To identify these
genes, prediction analysis was performed using the mirdb V5
database. As shown in Fig. 4,
circRNA-089763 can indirectly regulate a number of target genes
through endogenous competition. circRNA-089763 can bind
miR-7111-3p, miR-6769b-3p and miR-670-3p simultaneously. These
three microRNAs regulate the expression of insulin-like growth
factor binding protein 5 (IGFBP5). A total of two other miRNAs,
miR-3684 and miR-6798-3p, regulate the expression of stanniocalcin
1 (STC-1) and tyrosine 3-monooxygenase/tryptophan 5-monooxygenase
activation protein γ (YWHAG).
GO analysis results
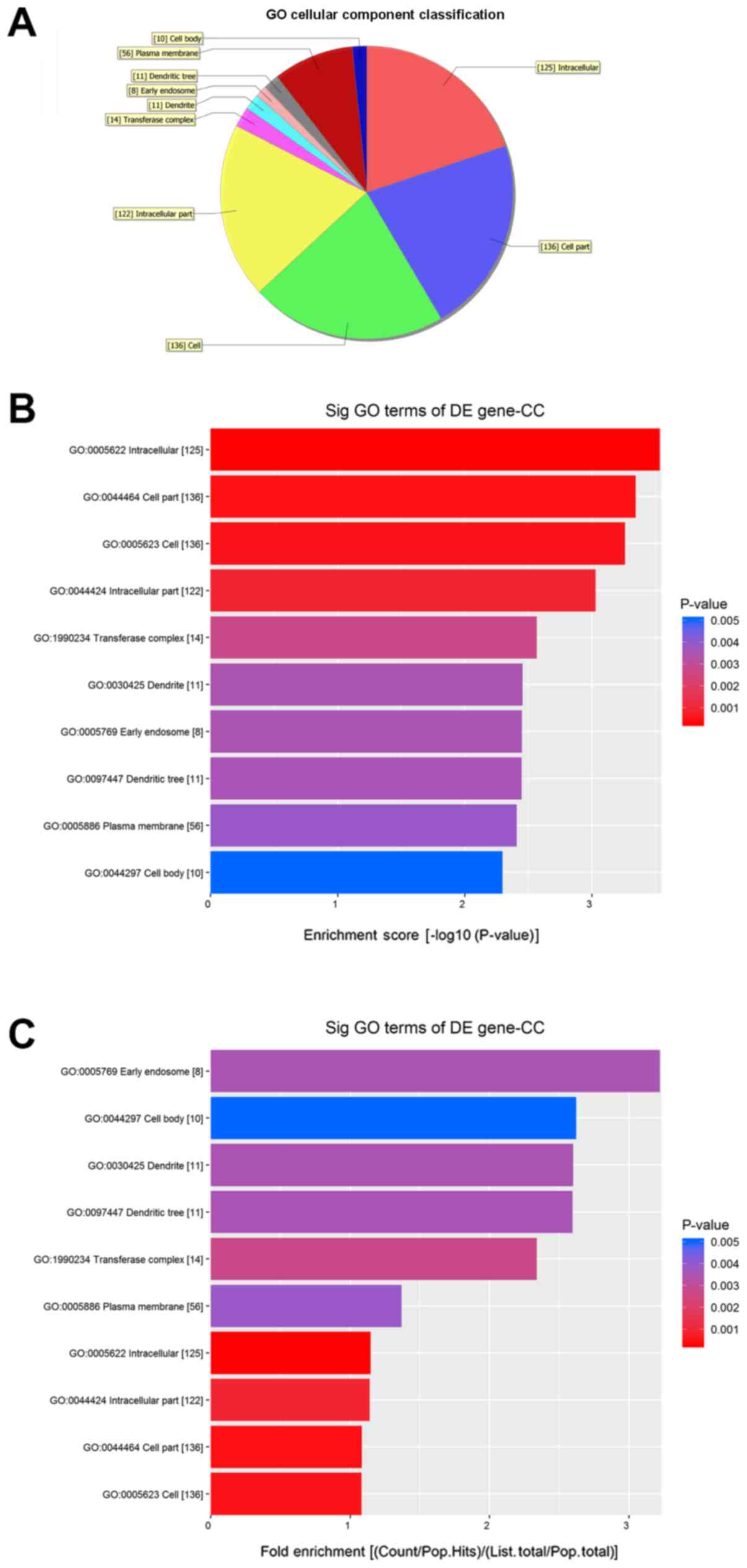
Fig. 5 shows the
molecular functions (MFs) of neurons that may be regulated by the
target genes of circRNA_089763 (P<0.05). Fig. 5A shows the classifications of these
molecular functions. Fig. 5B
orders the molecular functions based on their GO analysis
enrichment scores. Fig. 5C shows
the cellular processes, predicted using fold enrichment, regulated
by the target genes. Fig. 6 shows
the biological processes in neurons that may be regulated by the
target genes of circRNA_089763 (P<0.05). Fig. 6A shows the classifications of the
predicted biological processes and Fig. 6B orders the biological processes
based on their GO analysis enrichment score. Fig. 6C shows the biological processes,
predicted using fold enrichment, regulated by the target genes.
Fig. 7 shows the cellular
components that may be regulated by the target genes of
circRNA_089763 (P<0.05). Fig.
7A shows the classifications of these cellular components and
Fig. 7B orders the cellular
components based on their GO analysis enrichment scores. Fig. 7C shows the cellular components,
predicted using fold enrichment, regulated by the target genes.
KEGG pathway analysis results
Table V shows the
signaling pathways regulated by target genes of circRNA_089763 as
revealed by KEGG pathway analysis (P<0.05). The identified
pathways are involved in the regulation of central nervous system
functions, including the ‘sphingolipid signaling pathway’, ‘FoxO
signaling pathway’, ‘hippo signaling pathway’, ‘mRNA surveillance
pathway’, ‘endocytosis’ and ‘glucagon signaling pathway’.
| Table V.Signaling pathways regulated by
targeted genes of circular RNA_089763 (P<0.05). |
Table V.
Signaling pathways regulated by
targeted genes of circular RNA_089763 (P<0.05).
| PathwayID | Definition | Fisher P-value |
Enrichment_Score |
|---|
| hsa04666 | Fc gamma R-mediated
phagocytosis-Homo sapiens (human) | 0.004481613 | 2.348566 |
| hsa04071 | Sphingolipid
signaling pathway-Homo sapiens (human) | 0.01110983 | 1.954293 |
| hsa05230 | Central carbon
metabolism in cancer-Homo sapiens (human) | 0.01217405 | 1.914565 |
| hsa04380 | Osteoclast
differentiation-Homo sapiens (human) | 0.01462926 | 1.834777 |
| hsa05203 | Viral
carcinogenesis-Homo sapiens (human) | 0.01612574 | 1.79248 |
| hsa04068 | FoxO signaling
pathway-Homo sapiens (human) | 0.01621421 | 1.790104 |
| hsa04390 | Hippo signaling
pathway-Homo sapiens (human) | 0.02684785 | 1.57109 |
| hsa04934 | Cushing
syndrome-Homo sapiens (human) | 0.02684785 | 1.57109 |
| hsa03015 | mRNA surveillance
pathway-Homo sapiens (human) | 0.02960263 | 1.52867 |
| hsa04144 | Endocytosis-Homo
sapiens (human) | 0.03379667 | 1.471126 |
| hsa05165 | Human
papillomavirus infection-Homo sapiens (human) | 0.03837845 | 1.415913 |
| hsa04922 | Glucagon signaling
pathway-Homo sapiens (human) | 0.04051731 | 1.392359 |
| hsa04973 | Carbohydrate
digestion and absorption-Homo sapiens (human) | 0.04185221 | 1.378282 |
| hsa05152 |
Tuberculosis-Homo sapiens
(human) | 0.04307338 | 1.365791 |
| hsa04928 | Parathyroid hormone
synthesis, secretion and action-Homo sapiens (human) | 0.04352435 | 1.361268 |
| hsa05202 | Transcriptional
misregulation in cancer-Homo sapiens (human) | 0.04842783 | 1.314905 |
Discussion
Many patients develop POCD after heart surgery
(26). POCD is a significant risk
factor for a decline in quality of life after CABG (27). Thus, it is important to find
methods for the prevention and treatment of POCD. POCD is difficult
to accurately diagnose and few effective therapies are available;
this is in part due to the fact that the mechanism of POCD remains
unclear (28).
Because the pathophysiological mechanism of POCD is
complex and the clinical efficacy of drugs is poor, a more
comprehensive method to treat POCD is required (2). For many years, researchers have
focused their work on protein-coding genes and the proteins encoded
by these genes (10). Advances in
big data research have revealed that only a small fraction of the
genetic material encodes proteins. A large number of
transcriptional products result from functionally diverse ncRNAs,
including miRNA, lncRNAs and circRNAs. It is likely that the role
of a single miRNA in the regulation of a specific target gene is
relatively weak. However, a factor that could enrich multiple
miRNAs that regulate the same specific target gene could greatly
enhance the regulation of that gene.
circRNA molecules are different from traditional
linear RNA, with 5′ and 3′ ends, in that they have a closed-ring
structure that is unaffected by RNA exonucleases, has a more stable
expression and is not easily degraded (29). Research has shown that circRNA
molecules are rich in miRNA binding sites and that they act as an
miRNA sponge. By functioning as ceRNAs, circRNAs can relieve the
inhibitory effects of an miRNA on its target gene, thereby
increasing the expression of that target gene (30,31).
However, to the best of our knowledge, no circRNAs have been
reported to be associated with POCD. Therefore, in the present
study, the aim was to identify and investigate the potential role
of circRNAs in POCD.
To facilitate the clinical diagnosis of cognitive
function impairment, a number of studies have focused on biomarkers
based on molecules found in the cerebrospinal fluid (CSF) and
peripheral blood (32). However,
it is difficult to obtain CSF from patients with POCD by lumbar
puncture. Therefore, blood-based biomarkers for cognitive function
impairment for routine testing would be more suitable. One approach
could be to screen for plasma-specific circRNAs as biomarkers for
POCD. Exosomes are membrane vesicles of 40–100 nm that are released
from numerous cell types across the body. Exosomes contain various
substances, including ncRNAs, lipids and a variety of proteins
(33). It has been reported that
exosomes may serve as vesicular carriers for intercellular
communication in neurodegenerative disorders (34). A previous study described
neuronal-derived exosomal proteins in serum from healthy people and
patients with Alzheimer's disease (AD) (35). Accumulating evidence indicates that
exosomes are important in the pathogenesis of AD (33,35,36).
In the present study, candidate circRNAs were detected in the
plasma exosomes of patients with POCD. These circRNAs were further
investigated to determine whether they could provide information
about regulatory factors involved in POCD or could be utilized as
new biomarkers in the diagnosis and treatment of POCD.
The results in this present study showed that
circRNAs from plasma exosomes from patients with POCD could be
detected when there was sufficient quality and quantity of plasma
exosomes. The RNA purity, concentration and total quantity from 8
out of 12 samples from patients with POCD and 16 out of 23 samples
from patients without POCD met these requirements.
circRNA changes in the plasma exosomes of patients
with POCD were examined and it was found that the expression of
circRNA-089763 was significantly increased in patients with POCD.
Different levels of change were detected for circRNAs when
comparing the microarray and RT-qPCR method. However, the RT-qPCR
results are more credible. Data obtained using a chip, for example
in microarray experiments, provide a reference. This is due to the
many unspecific factors in the hybridization of the chip and the
number of objects measured by the chip may also cause distortion of
the results. RT-qPCR results are generally taken as the standard
(37).
As to the best of our knowledge, no previous studies
had investigated the role of circRNAs in POCD, the regulation of
target genes and signaling pathways required in-depth analysis.
Structural prediction analysis indicated that the miRNAs
sequestered by circRNA-089763 included hsa-miR-670-3p,
hsa-miR-6856-3p, hsa-miR-6769b-3p, hsa-miR-7111-3p,
hsa-miR-6803-3p, hsa-miR-614, hsa-miR-6784-3p, hsa-miR-454-5p,
hsa-miR-6798-3p and hsa-miR-3684. Therefore, circRNAs, including
circRNA-089763, could potentially induce the loss of function of
multiple miRNAs. Using circRNAs to control the expression of a
target gene would be expected to have a greater impact on gene
function than using a single miRNA targeting the same gene. This
hypothesis needs further testing in future studies.
In addition, according to the circRNA-miRNA-mRNA
network analysis conducted in this present study, it was found that
circRNA-089763 may function with miR-7111-3p, miR-6769b-3p and
miR-670-3p simultaneously, and that these three miRNAs may regulate
the IGFBP5 gene. IGFBP5 is involved in the regulation of amyloid-β
and cognitive functions (38–40).
circRNA-089763 may enrich miR-3684 and miR-6798-3p, two microRNAs
that regulate STC-1 and YWHAG. Shahim et al (41) found that the level of STC-1 in
cerebrospinal fluid is a potential biomarker in the differential
diagnosis of dementia. Ramocki et al (42) reported that the YWHAG gene is
involved in changes in cognitive and behavioral function.
Therefore, it is hypothesized that the pathogenesis of POCD is
associated with the abnormal expression levels of circRNA-089763
caused by peri-operative-related stimuli. circRNA-089763 may
indirectly regulate cognitive function by affecting the expression
of the IGFBP5, STC and YWHAG genes by sequestering the
corresponding microRNAs. As proteins are the factors that carry out
the functions of the target genes, it is important to investigate
the association between circRNAs and POCD at the protein level in
the future.
GO analysis of circRNA-089763 showed that its target
genes regulated many molecular functions, including ‘protein
binding’, ‘cell adhesion molecule binding’ and ‘DNA binding
transcription factor activity’; biological processes, including
‘regulation of biological process’, ‘regulation of signaling’ and
‘regulation of developmental process’; and cellular components,
including ‘dendritic tree’, ‘neuronal cell body’ and ‘postsynaptic
density’. KEGG pathway analysis revealed that some of the signaling
pathways regulated by the target genes of circRNA-089763 are
involved in ‘sphingolipid signaling pathway’, ‘FoxO signaling
pathway’, ‘hippo signaling pathway’, ‘mRNA surveillance pathway’,
‘endocytosis’ and ‘glucagon signaling pathway’. Many of these
signaling pathways are closely associated with the occurrence and
development of cognitive functions. Previous studies of signaling
pathways are consistent with the GO and KEGG pathway analysis
predictions in this present paper (43–51).
For example, the sphingolipid signaling pathway was reported to
play a role in AD and neurodegenerative diseases (43–45).
The FoxO signaling pathway has also been shown to be involved in
the pathogenesis of AD (46).
Neuroinflammation has emerged as an important cause of cognitive
decline during aging and in AD, and the glucagon signaling pathway
has been shown to be related to neuroinflammation and cognitive
function (47–50). Yang et al (51) reported a link between central
autophagy and cognitive dysfunction.
Due to the multifaceted mechanisms underlying POCD,
novel therapies should consider the multiple aspects of molecular
imbalance. As miRNAs are involved in several pathophysiological
aspects of POCD, they could serve as novel targets for POCD
therapies. circRNAs may be used as steady-state regulators in
locations associated with cognitive function or POCD. Although
overexpression of circRNAs acting as miRNA sponges would increase
the translation of their target genes, a decrease in circRNA
expression would result in the inappropriate silencing of an array
of downstream target genes. To prevent circRNA dysfunction in the
body, artificial circRNA mimics could be developed to restore
normal transcriptional regulation.
Due to the limited number of clinical CABG patients,
the number of samples in this present study was low. However, this
present study is the basis for further research. To increase the
sample size, other surgical diseases will be considered in future
studies to further investigate whether circRNA_089763 is elevated
in the circulating blood of patients with POCD. The genes and
pathways related to circRNA-089763 require further verification. In
addition, a unified world standard for POCD is still lacking, the
diagnostic criteria used in this present study are based on the
criteria used in a previous study (10).
In summary, the present study found that POCD
resulted in the abnormal expression of circRNA-089763 in patients
who underwent CABG. circRNA-089763 may be a novel biomarker that
could be utilized for the treatment of POCD after CABG.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Projects of
the National Natural Science Foundation of China (grant no.
81271478) and the Department of Science & Technology of Sichuan
Province (grant no. 14JC0093).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
MW, XW and SG designed and performed the
experiments, analyzed the data and wrote the manuscript. MW, PS, YL
and XZ provided the patient samples, conceived the study and
revised the manuscript. MW, JY, XW and XA performed the
experiments, analyzed the data and revised the manuscript. XW and
SG designed the experiments, provided reagents and revised the
manuscript.
Ethics approval and consent to
participate
The study was performed in accordance with the
Declaration of Helsinki, and was approved by the Ethics Committee
of Beijing Chaoyang Hospital (no. 2017-221-1). The participants
provided written informed consent to participate in this study.
Patient consent for publication
Consent for publication was obtained from all
participants.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
O'Brien H, Mohan H, Hare C, Reynolds J and
Kenn R: Mind over matter? The hidden epidemic of cognitive
dysfunction in the older surgical patient. Ann Surg. 265:677–691.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Skvarc D, Berk M, Byrne L, Dean O, Dodd S,
Lewis M, Marriott A, Moore EM, Morris G, Page RS and Gray L:
Post-operative cognitive dysfunction: An exploration of the
inflammatory hypothesis and novel therapies. Neurosci Biobehav Rev.
84:116–133. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kulason K, Nouchi R, Hoshikawa Y, Noda M,
Okada Y and Kawashima R: Indication of cognitive change and
associated risk factor after thoracic surgery in the elderly: A
pilot study. Front Aging Neurosci. 9:3962017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schenning KJ, Murchison CF, Mattek NC,
Kaye JA and Quinn JF: Sex and genetic differences in postoperative
cognitive dysfunction: A longitudinal cohort analysis. Biol Sex
Differ. 10:142019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Boodhwani M, Rubens FD, Wozny D, Rodriguez
R, Alsefaou A, Hendry PJ and Nathan HJ: Predictors of early
neurocognitive deficits in low-risk patients undergoing on-pump
coronary artery bypass surgery. Circulation. 114:I461–I466. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu YH, Wang DX, Li LH, Wu XM, Shan GJ, Su
Y, Li J, Yu QJ, Shi CX, Huang YN and Sun W: The effects of
cardiopulmonary bypass on the number of cerebral microemboli and
the incidence of cognitive dysfunction after coronary artery bypass
graft surgery. Anesth Analg. 109:1013–1022. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bishawi M, Hattler B, Almassi GH, Spertus
JA, Quin JA, Collins JF, Grover FL and Shroyer AL: Preoperative
factors associated with worsening in health-related quality of life
following coronary artery bypass grafting in the Randomized On/Off
Bypass (ROOBY) trial. Am Heart J. 198:33–38. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Needham MJ, Webb CE and Bryden DC:
Postoperative cognitive dysfunction and dementia: What we need to
know and do. Br J Anaesth. 119:i115–i125. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rundshagen I: Postoperative cognitive
dysfunction. Dtsch Arztebl Int. 111:119–125. 2014.PubMed/NCBI
|
|
10
|
Silva FP, Schmidt AP, Valentin LS, Pinto
KO, Zeferino SP, Oses JP, Wiener CD, Otsuki DA, Tort AB, Portela
LV, et al: S100B protein and neuron-specific enolase as predictors
of cognitive dysfunction after coronary artery bypass graft
surgery: A prospective observational study. Eur J Anaesthesiol.
33:681–689. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Deng L, Liu G, Zheng C, Zhang L, Kang Y
and Yang F: Circ-LAMP1 promotes T-cell lymphoblastic lymphoma
progression via acting as a ceRNA for miR-615-5p to regulate DDR2
expression. Gene. 701:146–151. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hanan M, Soreq H and Kadener S: CircRNAs
in the brain. RNA Biol. 14:1028–1034. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu C, Yao M, Li C, Shan K, Yang H, Wang
J, Liu B, Li XM, Yao J, Jiang Q and Yan B: Silencing Of circular
RNA-ZNF609 ameliorates vascular endothelial dysfunction.
Theranostics. 7:2863–2877. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Han B, Chao J and Yao H: Circular RNA and
its mechanisms in disease: From the bench to the clinic. Pharmacol
Ther. 187:31–44. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Floris G, Zhang L, Follesa P and Sun T:
Regulatory role of circular RNAs and neurological disorders. Mol
Neurobiol. 54:5156–5165. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li S, Li Y, Chen B, Zhao J, Yu S, Tang Y,
Zheng Q, Li Y, Wang P, He X and Huang S: ExoRBase: A database of
circRNA, lncRNA and mRNA in human blood exosomes. Nucleic Acids
Res. 46:D106–D112. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Otero-Ortega L, Laso-García F, Gómez-de
Frutos M, Fuentes B, Diekhorst L, Díez-Tejedor E and
Gutiérrez-Fernández M: Role of exosomes as a treatment and
potential biomarker for stroke. Transl Stroke Res. 10:241–249.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li X, Zhang S, Zhang J, Zhu J, He H, Zhang
Y, Zhang W and Tian D: Construct validity and reliability of the
test your memory chinese version in older neurology outpatient
attendees. Int J Ment Health Syst. 12:642018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Seo EH, Lee DY, Choo IH, Kim SG, Kim KW,
Youn JC, Jhoo JH and Woo JI: Normative study of the stroop color
and word test in an educationally diverse elderly population. Int J
Geriatr Psychiatry. 23:1020–1027. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Płotek W, Łyskawa W, Kluzik A, Grześkowiak
M, Podlewski R, Żaba Z and Drobnik L: Evaluation of the trail
making test and interval timing as measures of cognition in healthy
adults: Comparisons by age, education, and gender. Med Sci Monit.
20:173–181. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Forn C, Belloch V, Bustamante JC, Garbin
G, Parcet-Ibars MA, Sanjuan A, Ventura N and Avila C: A symbol
digit modalities test version suitable for functional MRI studies.
Neurosci Lett. 456:11–14. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Geffen GM, Butterworth P and Geffen LB:
Test-retest reliability of a new form of the auditory verbal
learning test (AVLT). Arch Clin Neuropsychol. 9:303–316. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Benedict RH, DeLuca J, Phillips G, LaRocca
N, Hudson LD and Rudick R; Multiple Sclerosis Outcome Assessments
Consortium, : Validity of the symbol digit modalities test as a
cognition performance outcome measure for multiple sclerosis. Mult
Scler. 23:721–733. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
The Criteria Committee of the New York
Heart Association, . Nomenclature and criteria for diagnosis of
diseases of the heart and great vessels. Dolgin M: 9th. Little,
Brown & Co; Boston: pp. 253–256. 1994
|
|
25
|
Li W, Freudenberg J, Suh YJ and Yang Y:
Using volcano plots and regularized-chi statistics in genetic
association studies. Comput Biol Chem. 48:77–83. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gilman S: Cerebral disorders after
open-heart operations. N Engl J Med. 272:489–498. 1965. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nijs K, Vandenbrande J, Vaqueriza F, Ory
JP, Yilmaz A, Starinieri P, Dubois J, Jamaer L, Arijs I and Stessel
B: Neurological outcome after minimal invasive coronary artery
surgery (NOMICS): Protocol for an observational prospective cohort
study. BMJ Open. 7:e0178232017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cascella M and Bimonte S: The role of
general anesthetics and the mechanisms of hippocampal and
extra-hippocampal dysfunctions in the genesis of postoperative
cognitive dysfunction. Neural Regen Res. 12:1780–1785. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jeck WR, Sorrentino JA, Wang K, Slevin MK,
Burd CE, Liu J, Marzluff WF and Sharpless NE: Circular RNAs are
abundant, conserved, and associated with ALU repeats. RNA.
19:141–157. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Van Rossum D, Verheijen BM and Pasterkamp
RJ: Circular RNAs: Novel regulators of neuronal development. Front
Mol Neurosci. 9:742016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ji MH, Yuan HM, Zhang GF, Li XM, Dong L,
Li WY, Zhou ZQ and Yang JJ: Changes in plasma and cerebrospinal
fluid biomarkers in aged patients with early postoperative
cognitive dysfunction following total hip-replacement surgery. J
Anesth. 27:236–242. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Iranifar E, Seresht BM, Momeni F, Fadaei
E, Mehr MH, Ebrahimi Z, Rahmati M, Kharazinejad E and Mirzaei H:
Exosomes and microRNAs: New potential therapeutic candidates in
alzheimer disease therapy. J Cell Physiol. 234:2296–2305. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Frühbeis C, Fröhlich D, Kuo WP, Amphornrat
J, Thilemann S, Saab AS, Kirchhoff F, Möbius W, Goebbels S, Nave
KA, et al: Neurotransmitter-triggered transfer of exosomes mediates
oligodendrocyte-neuron communication. PLoS Biol. 11:e10016042013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang TT, Liu CG, Gao SC, Zhang Y and Wang
PC: The serum exosome derived microRNA-135a, −193b, and −384 were
potential alzheimer's disease biomarkers. Biomed Environ Sci.
31:87–96. 2018.PubMed/NCBI
|
|
36
|
Ding M, Shen Y, Wang P, Xie Z, Xu S, Zhu
Z, Wang Y, Lyu Y, Wang D and Xu L: Exosomes isolated from human
umbilical cord mesenchymal stem cells alleviate neuroinflammation
and reduce amyloid-beta deposition by modulating microglial
activation in alzheimer's disease. Neurochem Res. 43:2165–2177.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhou M, Wang MH, Wang XB, Liu KZ, Wan YQ,
Li M, Liu L and Zhang C: Abnormal expression of microRNAs induced
by chronic unpredictable mild stress in rat hippocampal tissues.
Mol Neurobiol. 55:917–913. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Barucker C, Sommer A, Beckmann G, Eravci
M, Harmeier A, Schipke CG, Brockschnieder D, Dyrks T, Althoff V,
Fraser PE, et al: Alzheimer amyloid peptide aβ42 regulates gene
expression of transcription and growth factors. J Alzheimers Dis.
44:613–624. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fernandes J, Vieira AS, Kramer-Soares JC,
Da Silva EA, Lee KS, Lopes-Cendes I and Arida RM: Hippocampal
microRNA-mRNA regulatory network is affected by physical exercise.
Biochim Biophys Acta Gen Subj. 1862:1711–1720. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jing XP, Peng QH, Hu R, Zou HW, Wang HZ,
Yu XQ, Zhou JW, Degen A and Wang ZS: Dietary supplements during the
cold season increase rumen microbial abundance and improve rumen
epithelium development in tibetan sheep. J Anim Sci. 96:293–305.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shahim P, Blennow K, Johansson P, Svensson
J, Lista S, Hampel H, Andersson LC and Zetterberg H: Cerebrospinal
fluid stanniocalcin-1 as a biomarker for alzheimer's disease and
other neurodegenerative disorders. Neuromolecular Med. 19:154–160.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ramocki MB, Bartnik M, Szafranski P,
Kolodziejska KE, Xia Z, Bravo J, Miller GS, Rodriguez DL, Williams
CA, Bader PI, et al: Recurrent distal 7q11.23 deletion including
HIP1 and YWHAG identified in patients with intellectual
disabilities, epilepsy, and neurobehavioral problems. Am J Hum
Genet. 87:857–865. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Noel A, Ingrand S and Barrier L:
Inhibition of GSK3β by pharmacological modulation of sphingolipid
metabolism occurs independently of ganglioside disturbance in a
cellular model of Alzheimer's disease. Exp Neurol. 271:308–318.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mielke MM and Lyketsos CG: Alterations of
the sphingolipid pathway in Alzheimer's disease: New biomarkers and
treatment targets? Neuromolecular Med. 12:331–340. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ariga T, Jarvis WD and Yu RK: Role of
sphingolipid-mediated cell death in neurodegenerative diseases. J
Lipid Res. 39:1–16. 1998.PubMed/NCBI
|
|
46
|
Wang X, Wang Z, Chen Y, Huang X, Hu Y,
Zhang R, Ho MS and Xue L: FoxO mediates APP-induced AICD-dependent
cell death. Cell Death Dis. 5:e12332014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Qin L, Chong T, Rodriguez R and
Pugazhenthi S: Glucagon-like peptide-1-mediated modulation of
inflammatory pathways in the diabetic brain: Relevance to
alzheimer's disease. Current Alzheimer Res. 13:1346–1355. 2016.
View Article : Google Scholar
|
|
48
|
Tramutola A, Arena A, Cini C, Butterfield
DA and Barone E: Modulation of GLP-1 signaling as a novel
therapeutic approach in the treatment of Alzheimer's disease
pathology. Expert Rev Neurother. 17:59–75. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang Y, Chen S, Xu Z, Chen S, Yao W and
Gao X: GLP-1 receptor agonists downregulate aberrant GnT-III
expression in Alzheimer's disease models through the
Akt/GSK-3β/β-catenin signaling. Neuropharmacology. 131:190–199.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
An FM, Chen S, Xu Z, Yin L, Wang Y, Liu
AR, Yao WB and Gao XD: Glucagon-like peptide-1 regulates
mitochondrial biogenesis and tau phosphorylation against advanced
glycation end product-induced neuronal insult: Studies in vivo and
in vitro. Neuroscience. 300:75–84. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang N, Li L, Li Z, Ni C, Cao Y, Liu T,
Tian M, Chui D and Guo X: Protective effect of dapsone on cognitive
impairment induced by propofol involves hippocampal autophagy.
Neuroscience Lett. 649:85–92. 2017. View Article : Google Scholar
|