Introduction
MicroRNAs (miRNAs) are short noncoding RNAs that
regulate gene expression via hybridization to complementary
sequences of mRNAs (1). miRNAs
serve critical roles in biological processes and function as either
tumor suppressors or oncogenes (2,3). An
expanding body of evidence has identified a number of miRNA
functions in cancer development, including invasion and metastasis
(4).
As one of the most common malignancies worldwide,
lung cancer is known to have high morbidity and mortality rates
(5,6). The main types of lung cancer are
small cell lung cancer and non-small-cell lung cancer (NSCLC), with
the later accounting for ~85% of lung cancer cases (7). miRNAs have been reported to serve an
important role in cancer development and are recently considered as
key factors in lung cancer (2,8,9). For
instance, miRNAs were observed to be epigenetic modulators in lung
cancer, regulating the tumor microenvironment and the immune system
(10). The members of let-7 miRNA
family are highly expressed in normal lung tissue and have been
found to negatively control multiple oncogenes (11).
A previous study has identified miR-671-3p as a
potential osteoarthritis biomarker involved in metabolic processes
(12). In breast cancer,
miR-671-3p was observed to be downregulated and potentially
function as a tumor suppressor by influencing the Wnt signaling
cascade (13). However, the
effects and mechanisms of miR-671-3p on the lung cancer cells
remain unknown.
Forkhead box protein P2 (FOXP2) is a member of the
forkhead box transcription factor family, and is expressed in
various tissues (14). FOXP2 is
involved in the pathogenesis and development of numerous cancers,
including gastric, breast and hepatocellular cancers (15–17).
In this study, the aim was to determine the role of
miR-671-3p in NSCLC progression and the potential underlying
mechanisms. It was demonstrated that the expression of miR-671-3p
was upregulated in NSCLC tissues and cell lines (A549 and H1975),
and that miR-671-3p regulated NSCLC cell proliferation, apoptosis,
migration and invasion via directly targeting FOXP2.
Materials and methods
Human tissue samples
The study was reviewed and approved by the Ethics
Committee of the First People's Hospital of Changzhou (Changzhou,
China). Clinical samples (NSCLC tissues and adjacent normal
tissues) were collected from 40 NSCLC patients [23 males and 17
females; 11 patients aged <60 years old (45–59 years old) and 29
patients aged ≥60 years old (60–75 years old)] who received surgery
at The First People's Hospital of Changzhou (Changzhou, China)
between July 2015 and November 2017; and informed consent was
obtained from all patients. All diagnoses of NSCLC were confirmed
using pathological assays, including computed tomography, nuclear
magnetic resonance imaging and immunohistochemistry; none of the
patients included in the study had previously received any cancer
treatment. All tissues were immediately frozen in liquid nitrogen
and stored at −80°C for future experiments.
Cell culture and cell
transfection
Human normal lung 16HBE and human NSCLC (A549 and
H1975) cell lines were obtained from the Chinese Academy of Science
Cell Bank (Shanghai, China). Cells were cultured in RPMI-1640
medium (HyClone; GE Healthcare Life Sciences, Logan, UT, USA)
supplied with 10% fetal bovine serum (FBS; Biowest, Barcelona,
Spain) at 37°C with 5% CO2. miR-671-3p mimics,
inhibitors, mimics NC and inhibitors NC were purchased from
Shanghai GenePharma Co., Ltd. (Shanghai, China). pcDNA.3.1-FOXP2
and pcDNA3.1 empty vector (2 µg) were inserted into a
pCDF-CMV-MCS-EF1-Peru lentiviral vector using 293T cells
(5×104; Chinese Academy of Science Cell Bank) and
Entranster™-H (Engreen Biosystem Co., Ltd., Beijing, China); all
the vectors were synthesized by Shanghai GenePharma Co., Ltd.
(Shanghai, China). The lentiviral supernatant was collected 48 h
after the transfection and used to infect cancer cells. The empty
pcDNA3.1 vector was used as negative control. The following primers
for FOXP2 were used: Sense, 5′-GGATCCATGATGCAGGAATCTGCG-3′, and
antisense, 5′-CTCGAGTCATTCCAGATCTTCAGAT-3′. Cells
(5×104) were transfected with 200 nM miR-671-3p mimic,
50 nM miR-671-3p inhibitor and 8×108 TU/ml
pcDNA3.1-FOXP2 lentivirus by using Lipofectamine® 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). After 24 or 48 h, transfected cells were collected and used
in RNA or protein assays.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
For miR-671-3p detection, total RNA was extracted
from cells and tissues with the miRcute miRNA Isolation kit
(Tiangen, Shanghai, China) and reverse-transcribed using One Step
PrimeScript miRNA cDNA Synthesis kit (Takara Biotechnology Co.,
Ltd., Dalian, China) according to the manufcaturer's protocols. For
FOXP2 mRNA expression detection, total RNA was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and then reverse-transcribed using M-MLV reverse
transcriptase (Promega Corporation, Madison, WI, USA) according to
the manufacturer's protocols. Subsequently, qPCR was performed
using the SYBR ExScript RT-qPCR kit (Takara Biotechnology Co.,
Ltd.) in an ABI 7500 Real-Time PCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc.). U6 and GAPDH served as the
endogenous control for miRNA and mRNA, respectively and each sample
was detected in triplicate. The reaction was conducted in a total
volume of 20 µl, containing 1 µl cDNA, 10 µl SYBR Premix EX Taq, 1
µl of each primer (10 µM), and 7 µl ddH2O. The PCR
program was conducted at 95°C for 3 min, followed by 40 cycles of
95°C for 10 sec, and 60°C for 30 sec. The primers used were as
follows: GAPDH, forward 5′-TGAACTGAAAGCTCTCCACC-3′, reverse,
5′-CTGATGTACCAGTTGGGGAA-3′; U6, forward 5′-CTCGCTTCGGCAGCACA-3′,
reverse, 5′-AACGCTTCACGAATTTGCGT-3′; FOXP2, forward
5′-CCACGAAGACCTCAATGGTT-3′, reverse, 5′-TCACGCTGAGGTTTCACAAG-3′;
and miR-671-3p, forward 5′-CCGGUUCUCAGGGCUCCACC-3′ and reverse,
5′-GTGCAGGGTCCGAGGT-3′ (Sangon Biotech Co., Ltd., Shanghai, China).
All fold changes were calculated using the comparative Cq method
(2−ΔΔCq), using GAPDH or U6 for normalization (18).
Western blotting
Total proteins from cells and tissues were lysed
with radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology, Haimen, China), and the concentration
of protein was measured using a bicinchoninic acid assay kit
(Beyotime Institute of Biotechnology). A total of 50 µg total
protein per lane was separated by 10% SDS-PAGE and blotted onto
polyvinylidene fluoride membranes. Next, the membrane was blocked
using 5% non-fat milk in PBS-0.5% Tween 20 at room temperature for
3 h and probed with primary antibodies against FOXP2 (1:500;
ab16046; Abcam, Cambridge, MA, USA), B-cell lymphoma 2 (Bcl-2;
1:1,000; ab32124; Abcam), Bcl-2-associated X protein (Bax; 1:1,000;
ab32503; Abcam), caspase-3 (1:1,000; ab13847; Abcam), caspase-9
(1:1,000, ab13847; Abcam), matrix metalloproteinase-2 (MMP-2;
1:1,000; ab37150; Abcam), MMP-9 (1:1,000; ab73734; Abcam) and GAPDH
(1:5,000; G8795; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at
4°C overnight. The membranes were then incubated with goat
anti-mouse secondary antibody conjugated with horseradish
peroxidase (1:5,000; ab97040; Abcam) for 1 h at room temperature.
Finally, the membranes were visualized using an enhanced
chemiluminescence kit (Beyotime Institute of Biotechnology). The
bands were scanned using a densitometer, and the gray values of the
bands were calculated automatically by the ImageJ software (version
1.45k; National Institutes of Health, Bethesda, MD, USA).
Cell proliferation and colony
formation assays
Cell proliferation was detected by Cell Counting Kit
(CCK)-8 and 5-ethynyl-2′-deoxyuridine (EdU) assays according to the
manufacturers' protocol. Briefly, cells were seeded at a density of
1×104 cells/well in 96-well plates and cultured for 24 h
before transfection. After transfection for 48 h, 10 µl of CCK-8
solution (Beyotime Institute of Biotechnology) was added and
incubated for 1 h at 37°C. Finally, the optical density value at
450 nm was determined using a microplate reader (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
For colony formation assay, cells were seeded in a
6-well plate at a density of 1,000 cells/well at 37°C for 2 weeks.
Subsequently, the cells were fixed with 95% methanol for 30 min at
room temperature and stained with 0.1% crystal violet for 15 min at
room temperature, and the colonies were examined.
Cell apoptosis assay
Cell apoptosis was detected by flow cytometry
analysis using a fluorescein isothiocyanate (FITC)-Annexin
V/propidium iodide (PI) Apoptosis Detection kit (BD Pharmingen; BD
Biosciences, San Jose, CA, USA). Briefly, the cells were collected
and adjusted to the concentration of 1×105 cells/ml. A
cell suspension of 200 µl was prepared for each sample, washed
twice with cold PBS and resuspended in 100 µl binding buffer
(Beyotime Institute of Biotechnology). Then, 5 µl Annexin V-FITC
(20 µg/ml) and 5 µl PI (50 µg/ml) were added and the cells were
incubated on ice in the dark for 15 min. Finally, 400 µl binding
buffer was added and fluorescence determination of apoptotic cells
was performed was performed using a FC500 flow cytometer equipped
with Cell Quest 3.0 software (BD Biosciences). All analyses were
performed in triplicate.
Scratch assay
Cells were seeded in 6-well plates at a density of
5×105 cells/well until 80% of the cells were attached to
the plate wall, and then rinsed in PBS twice to remove any floating
cells. Next, wounds were made in each well with sterile pipette
tips. Subsequently, the cells were washed twice with PBS, and 2 ml
RPMI-1640 culture medium containing 10% FBS was added to the
culture plate. Images of the cells were captured at 24 and 48 h
after wound generation to assess the cell migration ability.
Transwell invasion assay
Transwell assays were conducted to analyze the
migration and invasion abilities of cells. Briefly,
4×104 cells in 100 µl RPMI-1640 culture medium
supplemented with 10% FBS were placed in the upper chamber of
Transwell plates (Corning Inc., Corning, NY, USA), which were
pre-coated with Matrigel (growth factor reduced; BD Biosciences).
Dulbecco's modified Eagle's medium (Beyotime Institute of
Biotechnology) with 20% FBS was added to the lower chambers. After
24-h incubation, cells that had invaded across the membrane were
fixed in 95% methanol at room temperature and stained with 0.1%
crystal violet for 10 min at room temperature. Cells in nine
randomly selected fields were counted under an inverted microscope
(Carl Zeiss AG, Jena, Germany) at ×200 magnification, and the
number of invading cells was expressed as the mean cell number in
these fields.
Bioinformatics analysis
The TargetScan database 7.1 (http://www.targetscan.org/) was used to predict
putative targets of miR-671-3p.
Luciferase reporter assay
The wild-type 3′-untranslated region (3′UTR) of
FOXP2 gene was amplified using the following primers: Sense,
5′-GGTACCCTTTACAAACAGTTTTGACAG-3′, and antisense,
5′-AAGCTTTGGTGTGAATGATGACTGG-3′. The PCR products were cloned into
the pGL3-basic vector using KpnI/HindIII sites to
obtain the pGL3-FOXP2-3′UTR vector. A mutant version of this
construct (pGL3-FOXP2-mut-3′UTR) carrying 4-bp substitutions in the
miR-671-3p target sites was also obtained by site-directed
mutagenesis using the QuikChange Site-Directed Mutagenesis kit
(Agilent Technologies, Inc., Santa Clara, CA, USA). The following
primers were used for amplifying the mutant 3′UTR: Fragment 1,
forward 5′-CAGCGATCGCGAACTGACTTGTGAAACCTCAGCG-3′, reverse,
5′-CTCGCAGTTACTTCCAGTCCCTCAAAGCC-3′; and fragment 2, forward
5′-GTCTTTGGGTCATGATCAACGAACCGG-3′ and reverse,
5′-TATGTTTAAACTTTATAAATGGGTCAAAAAGAATTAGA-3′. A549 and H1975 cells
were seeded at a density of 2×105 cells/well onto
24-well plates 12 h prior to the transfection, and then
co-transfected with miR-671-3p mimics or negative control (NC)
mimics along with the pGL3-FOXP2-3′UTR or pGL3-FOXP2-mut-3′UTR
using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc.). After 36 h of transfection, the luciferase activity was
measured with a Dual-Glo Luciferase Assay System (Promega
Corporation, Madison, WI, USA) and normalized to Renilla
luciferase.
Statistical analysis
All results are reported as the mean ± standard
deviation, and were obtained using at least three independent
replicates. All statistical analyses were performed using GraphPad
software (version 5.0; GraphPad Software, Inc., La Jolla, CA, USA).
Differences between two groups were calculated by Student's t-test.
Differences among three or more groups were calculated by one-way
analysis of variance test, followed by Tukey's multiple comparison
test. Pearson correlation analysis was used to investigate the
association between FOXP2 and miRNA-671-3p. A P-value of <0.05
was considered to denote a difference that was statistically
significant.
Results
High expression of miR-671-3p in NSCLC
tissues and cell lines
miR-671-3p has been reported to be a tumor
suppressor in breast cancer (19).
However, the effects and underlying mechanisms of miR-671-3p in
lung cancer remain unknown. Therefore, the current study examined
the expression pattern of miR-671-3p in NSCLC. miR-671-3p
expression in 40 human NSCLC and adjacent normal lung tissues was
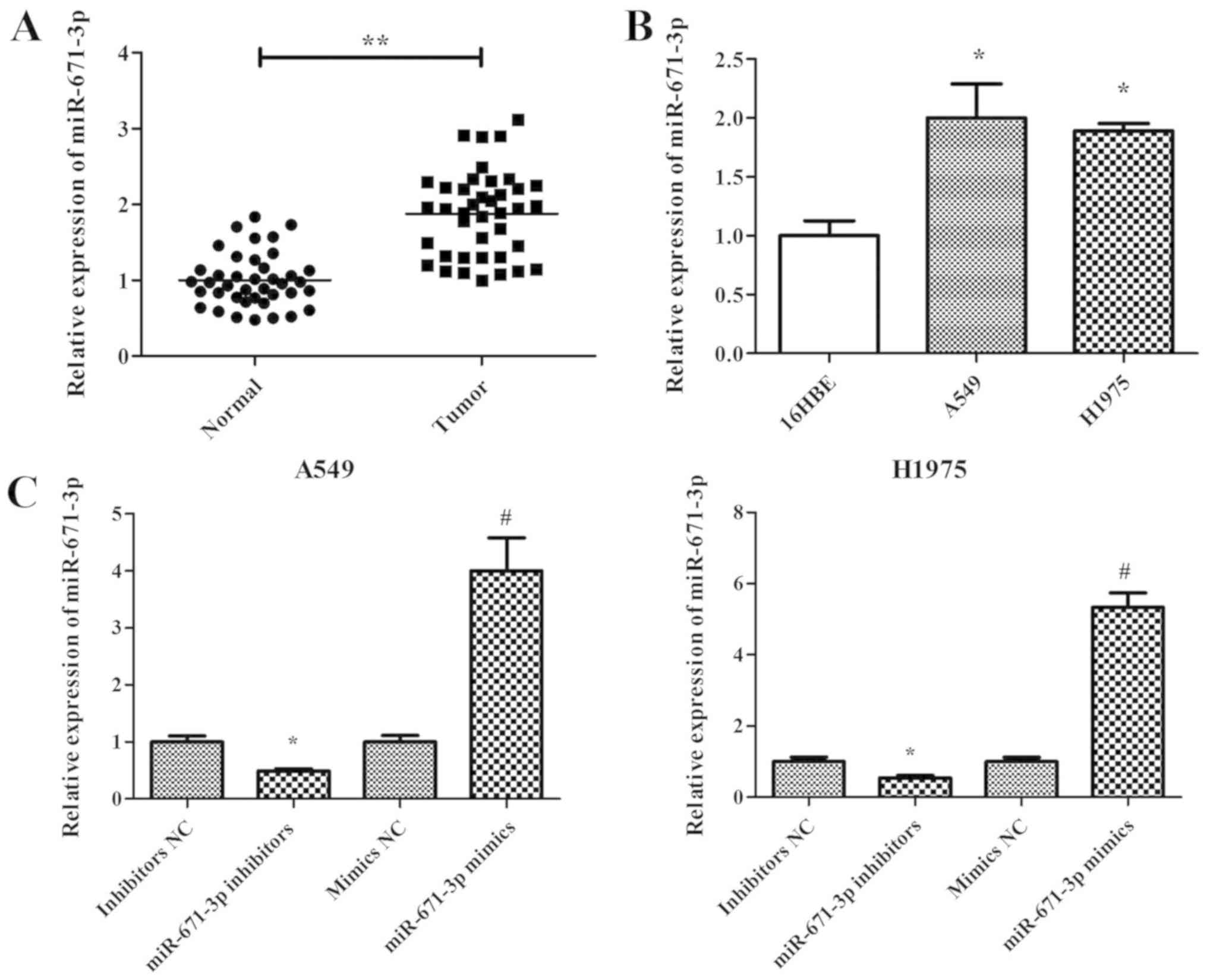
measured by RT-qPCR. The results demonstrated that miR-671-3p
expression in NSCLC tissues was significantly higher compared with
that in adjacent normal tissues (P<0.05; Fig. 1A). Furthermore, miR-671-3p
expression in two NSCLC cell lines (A549 and H1975) was measured,
and the normal lung 16HBE cell line was used as the control.
Similar to the results in NSCLC tissues, the expression level of
miR-671-3p was significantly upregulated in A549 and H1975 cells
compared with that in 16HBE cells (Fig. 1B). These findings suggested that
miR-671-3p is upregulated in NSCLC tissues and cells.
Downregulation of miR-671-3p inhibits
NSCLC cell proliferation and induces apoptosis in vitro
Sustained proliferation is one of the hallmarks of
cancer, and the fundamental basis of cancer dissemination and
metastasis (20). In order to
explore the role of miR-671-3p in NSCLC cell proliferation,
miR-671-3p was knocked down in A549 and H1975 cells by transfection
with miR-671-3p inhibitors. The RT-qPCR results confirmed that
miR-671-3p was efficiently downregulated following transfection
with miR-671-3p inhibitors, as compared with the NC group (Fig. 1C). Next, the CCK-8 assay results
indicated that the proliferation capacity was decreased in A549 and
H1975 cells transfected with miR-671-3p inhibitors (Fig. 2A). Similarly, EdU and colony
formation assays were revealed reduced cell proliferation and
colony formation in the inhibitor group compared with the NC group
(Fig. 2B and C). In addition,
fewer early and late apoptotic cells were detected in cells
transfected with NC as compared with those transfected with
miR-671-3p inhibitors, indicating that miR-671-3p downregulation
promoted the apoptosis of A549 and H1975 cells (Fig. 2D and E). Furthermore, the levels of
apoptosis-associated proteins were detected by western blotting. It
was observed that the anti-apoptotic protein Bcl-2 was
significantly downregulated in the miR-671-3p inhibitor group,
whereas pro-apoptotic proteins Bax, caspase-3 and caspase-9 were
markedly upregulated (Fig.
2F).
Downregulation of miR-671-3p
suppresses NSCLC cell migration and invasion in vitro
To explore the function of miR-671-3p in NSCLC cell
migration and invasion, scratch and transwell assays were
performed. The results of the scratch assay demonstrated that there
was significant difference in the migrated distance between the
control and miR-671-3p inhibitor groups (P<0.05; Fig. 3A). As displayed in Fig. 3B, the transwell assay revealed that
an increased number of cells had invaded to the lower chamber in
the control group as compared with that in the miR-671-3p inhibitor
group.
Matrix metalloproteinases (MMPs) have an important
role in cancer cell migration and invasion by degrading the
extracellular matrix (21,22). Therefore, western blotting was then
performed to detect the expression levels of MMP-2 and MMP-9. As
shown in Fig. 3C, MMP-2 and MMP-9
were significantly decreased in A549 and H1975 cells transfected
with miR-671-3p inhibitors. Taken together, these results indicated
that miR-671-3p downregulation inhibited NSCLC cell migration and
invasion in vitro.
miR-671-3p directly targets FOXP2 in
NSCLC
To explore the molecular mechanisms by which
miR-671-3p promoted tumor cell progression, a search for putative
miR-671-3p targets was conducted using TargetScan software. FOXP2
was predicted to be a potential candidate target of miR-671-3p
(Fig. 4A). A luciferase reporter
assay was then performed to detect the interaction between FOXP2
and miR-671-3p. As shown in Fig.
4B, overexpression of miR-671-3p decreased the luciferase
signal in FOXP2-3′UTR cells by ~60% (P<0.05). However, no
significant differences were identified between the miR-671-3p
mimics and mimics NC groups in cells transfected with
FOXP2-3′UTR-mut. These results indicated that miR-671-3p targeted
the 3′UTR of FOXP2 directly in NSCLC.
miR-671-3p expression is negatively
correlated with FOXP2 in NSCLC tissues and cell lines
RT-qPCR and western blotting were performed to
determine the mRNA and protein levels of FOXP2 in 40 human NSCLC
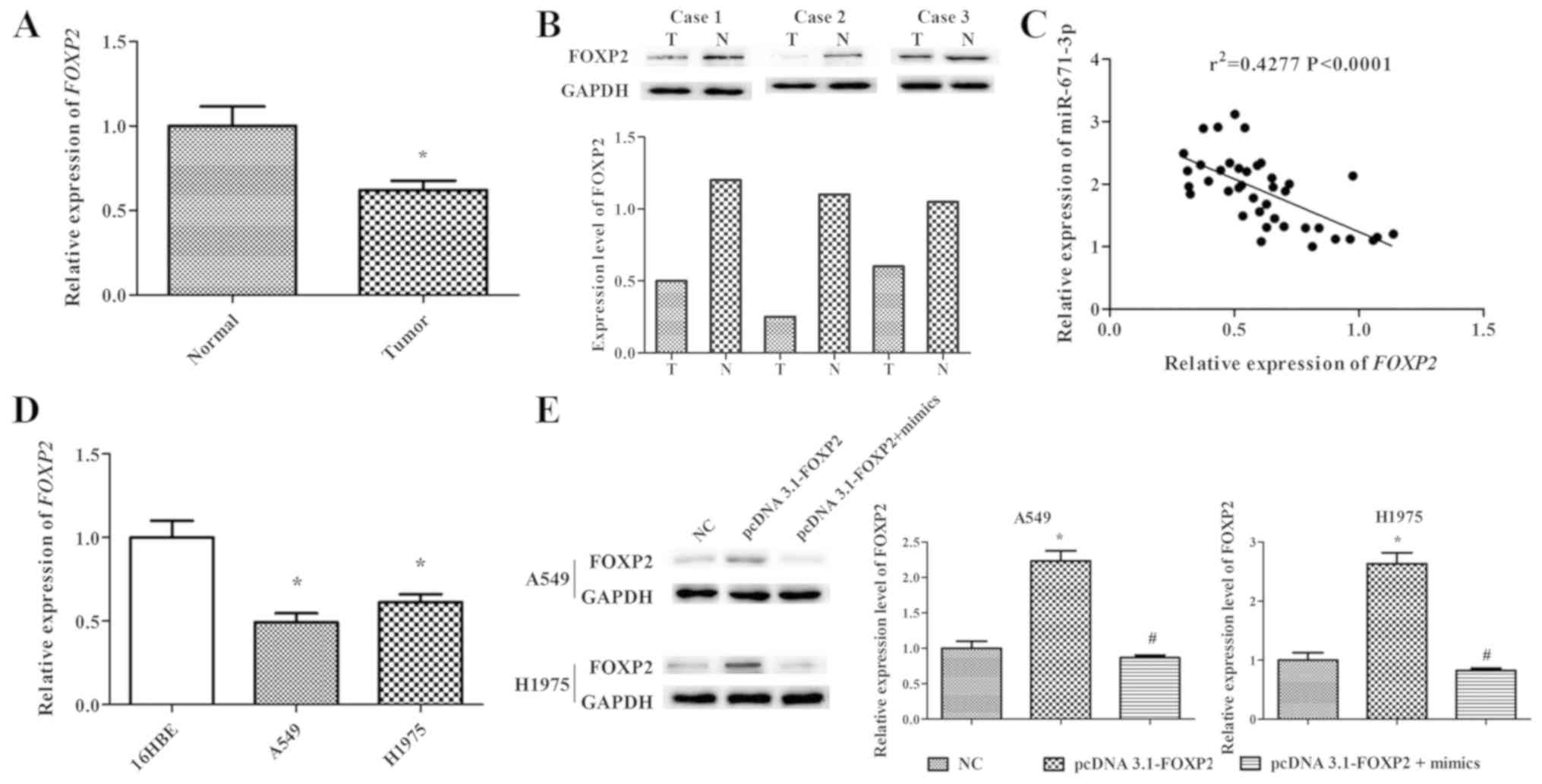
lung tissues. The results indicated that FOXP2 expression in NSCLC
lung tissues was markedly lower compared with that in adjacent
normal lung tissues (P<0.05; Fig.
5A and B). In order to further explore the association between
the FOXP2 and miR-671-3p expression levels, Pearson correlation
analysis was conducted, and the results demonstrated that
miR-671-3p expression was negatively correlated with FOXP2
expression in NSCLC tissues (r2=0.4277, P<0.0001;
Fig. 5C). Furthermore, NSCLC cell
lines (A549 and H1975) exhibited significantly lower FOXP2
expression in comparison with that in the normal lung 16HBE cell
line (Fig. 5D).
miR-671-3p promotes NSCLC cell
proliferation, migration and invasion, and suppresses apoptosis by
downregulating FOXP2 in vitro
To explore the role of FOXP2 in NSCLC cell
progression, the pcDNA3.1-FOXP2 vector was constructed. FOXP2
expression level in the pcDNA3.1-FOXP2 cell group was ~2.5 times
that in NC cells. However, transfection with miR-671-3p mimics
significantly downregulated FOXP2 expression in pcDNA3.1-FOXP2
cells (Fig. 5E).
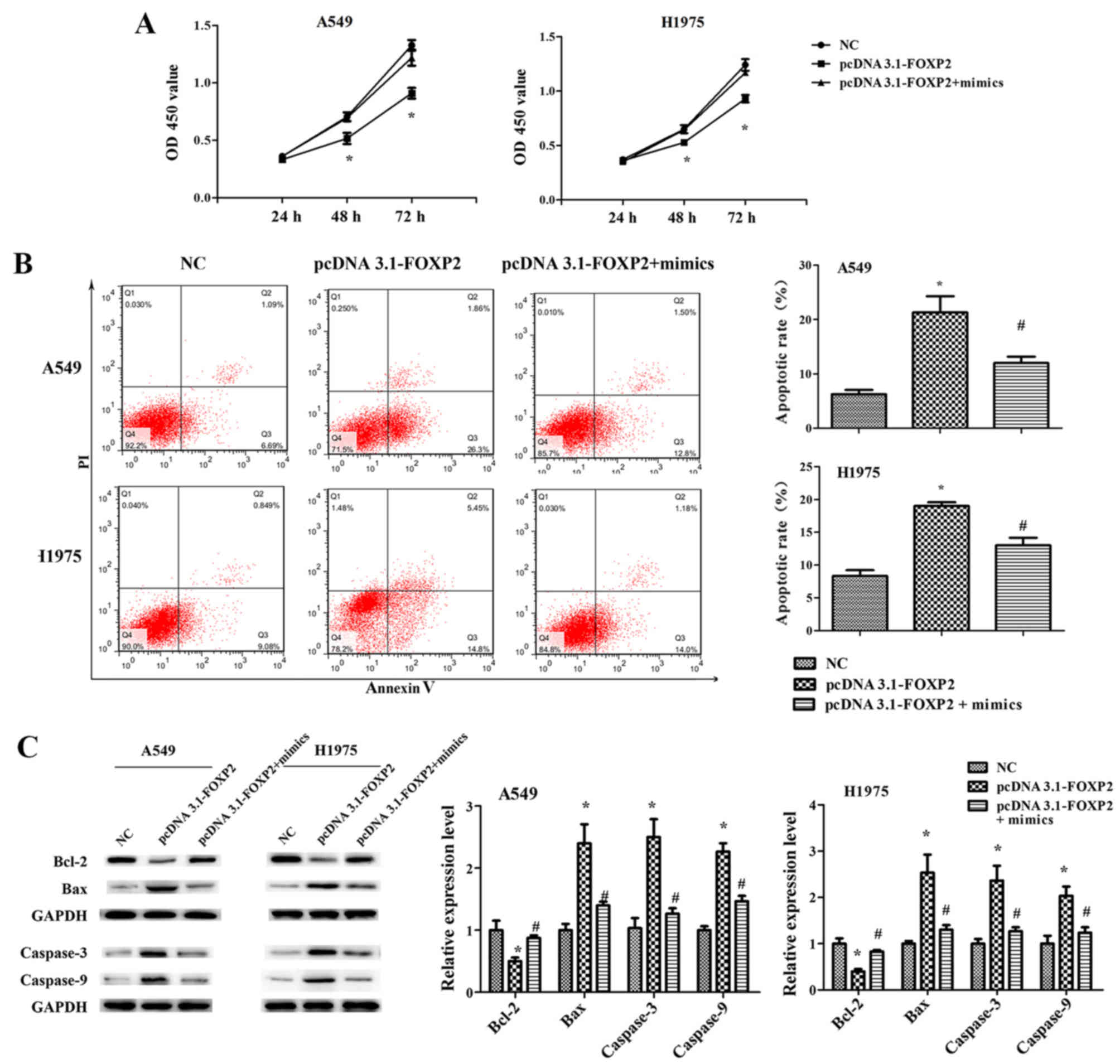
Furthermore, it was observed that FOXP2
overexpression significantly suppressed the proliferation,
migration and invasion ability of A549 and H1975 cells, while it
induced cell apoptosis (Figs. 6
and 7). However, the effects of
pcDNA3.1-FOXP2 on cell proliferation, apoptosis, migration and
invasion were reversed in the presence of miR-671-3p mimics
(Figs. 6 and 7). These findings implied that miR-671-3p
promotes NSCLC cell proliferation, migration and invasion, and
suppresses apoptosis by downregulating FOXP2 in vitro.
Discussion
It is known that dysregulation of specific miRNAs
can be involved in the initiation and progression of cancer
(23). Various miRNAs have been
reported to function as oncogenes or tumor suppressors in NSCLC by
regulating the expression of downstream target genes (24,25).
A previous study revealed that miR-671-5p served tumor suppressor
roles in breast cancer; overexpression of miR-671-5p attenuated the
proliferation and invasion of breast cancer cells by targeting
FOXM1 (26). To the best of our
knowledge, this is the first study to investigate the role of
miR-671-3p in NSCLC. In the present study, the expression of
miR-671-3p was increased in NSCLC tissues and cell lines (A549 and
H1975), as compared with normal lung tissue and cells,
respectively. To investigate the potential oncogenic roles of
miR-671-3p in NSCLC cells, cells were transfected with miR-671-3p
inhibitors. Inhibition of miR-671-3p was reported to suppress the
proliferation, migration and invasion, and promote the apoptosis of
A549 and H1975 cells.
FOXP2 is a member of the FOX family of proteins,
which are involved in a number of important biological processes
including cell metabolism, development, differentiation,
proliferation, apoptosis, migration and invasion (27,28).
FOXP2 was the first characterized FOX family member that was
relevant to the human ability to develop speech (29,30).
In recent years, FOXP2 has been reported to serve an important role
in cancer (31). FOXP2 was found
to be overexpressed in lymphoma cells (32), while low expression of FOXP2 was
correlated with poor survival in hepatocellular carcinoma (33). In addition, Cuiffo et al
(16) reported that elevated
miR-199a levels and suppressed FOXP2 expression levels are
prominent features of malignant clinical breast cancer.
Furthermore, downregulation of FOXP2 expression was observed in
gastric cancer cells and tissues, while miR-190 was observed to
regulate the pathological process of gastric cancer through the
FOXP2 pathway (15). Another study
indicated that miR-618 inhibited prostate cancer migration and
invasion by targeting FOXP2 (34).
The role of FOXP2 in cancer development has also been reported in
several other studies (35–37).
However, to the best of our knowledge, the current study is the
first to report on the expression of FOXP2 and its direct targeting
by miR-671-3p in NSCLC.
In the present study, downregulation of FOXP2 was
observed in NSCLC tissues and cells. Bioinformatics and luciferase
assays revealed that FOXP2 was direct target gene of miR-671-3p in
NSCLC cells. RTq-PCR and western blotting demonstrated that
overexpression of miR-671-3p reduced the expression of FOXP2 at the
mRNA and protein levels. Furthermore, it was observed that the
overexpression of FOXP2 inhibited the proliferation, migration and
invasion, and promoted the apoptosis of A549 and H1975 cells.
Conversely, co-transfection with miR-671-3p mimics attenuated the
effects of pcDNA3.1-FOXP2. The results indicated that FOXP2 is a
downstream target, and a functional mediator of the effects of
miR-671-3p in NSCLC.
In conclusion, the present study revealed that
miR-671-3p improved the proliferation, migration and invasion of
NSCLC cells, as well as suppressed cell apoptosis, by regulating
FOXP2 expression. These findings suggested that miR-671-3p
functions as an oncogene and may be a potential therapeutic target
in NSCLC.
Acknowledgements
Not applicable.
Funding
No funding was recieved.
Availability of data and materials
The datasets used during the current study are
available from the corresponding author on reasonable request.
Authors' contributions
ZYL and ZZZ were major contributors in sample
collection, the conduction of experiments, data interpretation and
manuscript preparation. HB, QDZ, SJZ, LZ and XQZ participated in
sample collection, conduction of experiments and data
interpretation. JZ was responsible for experimental design and
revised this manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Informed consent was obtained from each patient, and
this study was approved by the Ethics Committee of the First
People's Hospital of Changzhou, China.
Patient consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
References
|
1
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang Y, Yang Q and Wang S: MicroRNAs: A
new key in lung cancer. Cancer Chemother Pharmacol. 74:1105–1111.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yin R, Guo L, Zhang W and Zheng J: The
pleiotropic effects of mirnas on tumor angiogenesis. J Cell
Biochem. 116:1807–1815. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ma L, Teruya-Feldstein J and Weinberg RA:
Tumour invasion and metastasis initiated by microRNA-10b in breast
cancer. Nature. 449:682–688. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hess KR, Varadhachary GR, Taylor SH, Wei
W, Raber MN, Lenzi R and Abbruzzese JL: Metastatic patterns in
adenocarcinoma. Cancer. 106:1624–1633. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McIntyre A and Ganti AK: Lung cancer-A
global perspective. J Surg Oncol. 115:550–554. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cho WC: Role of miRNAs in lung cancer.
Expert Rev Mol Diagn. 9:773–776. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hashemi ZS, Khalili S, Forouzandeh
Moghadam M and Sadroddiny E: Lung cancer and miRNAs: A possible
remedy for anti-metastatic, therapeutic and diagnostic
applications. Expert Rev Respir Med. 11:147–157. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rusek AM, Abba M, Eljaszewicz A, Moniuszko
M, Niklinski J and Allgayer H: MicroRNA modulators of epigenetic
regulation, the tumor microenvironment and the immune system in
lung cancer. Mol Cancer. 14:342015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Johnson CD, Esquela-Kerscher A, Stefani G,
Byrom M, Kelnar K, Ovcharenko D, Wilson M, Wang X, Shelton J,
Shingara J, et al: The let-7 microRNA represses cell proliferation
pathways in human cells. Cancer Res. 67:7713–7722. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ntoumou E, Tzetis M, Braoudaki M, Lambrou
G, Poulou M, Malizos K, Stefanou N, Anastasopoulou L and Tsezou A:
Serum microRNA array analysis identifies miR-140-3p, miR-33b-3p and
miR-671-3p as potential osteoarthritis biomarkers involved in
metabolic processes. Clin Epigenetics. 9:1272017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xiong DD, Chen H, He RQ, Lan AH, Zhong JC,
Chen G, Feng ZB and Wei KL: MicroRNA-671-3p inhibits the
development of breast cancer: A study based on in vitro
experiments, in-house quantitative polymerase chain reaction and
bioinformatics analysis. Int J Oncol. 52:1801–1814. 2018.PubMed/NCBI
|
|
14
|
Chen MT, Sun HF, Li LD, Zhao Y, Yang LP,
Gao SP and Jin W: Downregulation of FOXP2 promotes breast cancer
migration and invasion through TGFβ/SMAD signaling pathway. Oncol
Lett. 15:8582–8588. 2018.PubMed/NCBI
|
|
15
|
Jia WZ, Yu T, An Q, Yang H, Zhang Z, Liu X
and Xiao G: MicroRNA-190 regulates FOXP2 genes in human gastric
cancer. Onco Targets Ther. 9:3643–3651. 2016.PubMed/NCBI
|
|
16
|
Cuiffo BG, Campagne A, Bell GW, Lembo A,
Orso F, Lien EC, Bhasin MK, Raimo M, Hanson SE, Marusyk A, et al:
MSC-regulated microRNAs converge on the transcription factor FOXP2
and promote breast cancer metastasis. Cell Stem Cell. 15:762–774.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu Z, Lin X, Tian M and Chang W:
microRNA-196b promotes cell migration and invasion by targeting
FOXP2 in hepatocellular carcinoma. Oncol Rep. 39:731–738.
2018.PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen X, Lu P, Wang DD, Yang SJ, Wu Y, Shen
HY, Zhong SL, Zhao JH and Tang JH: The role of miRNAs in drug
resistance and prognosis of breast cancer formalin-fixed
paraffin-embedded tissues. Gene. 595:221–226. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiménez MJ, Balbin M, Alvarez J, Komori T,
Bianco P, Holmbeck K, Birkedal-Hansen H, López JM and López-Otín C:
A regulatory cascade involving retinoic acid, Cbfa1, and matrix
metalloproteinases is coupled to the development of a process of
perichondrial invasion and osteogenic differentiation during bone
formation. J Cell Biol. 155:1333–1344. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Khokha R and Denhardt DT: Matrix
metalloproteinases and tissue inhibitor of metalloproteinases: A
review of their role in tumorigenesis and tissue invasion. Invasion
Metastasis. 9:391–405. 1989.PubMed/NCBI
|
|
23
|
Tan W, Liu B, Qu S, Liang G, Luo W and
Gong C: MicroRNAs and cancer: Key paradigms in molecular therapy.
Oncol Lett. 15:2735–2742. 2018.PubMed/NCBI
|
|
24
|
Yang Y, Ding L, Hu Q, Xia J, Sun J, Wang
X, Xiong H, Gurbani D, Li L, Liu Y and Liu A: MicroRNA-218
functions as a tumor suppressor in lung cancer by targeting
IL-6/STAT3 and negatively correlates with poor prognosis. Mol
Cancer. 16:1412017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang Y, Li H, Liu Y, Chi C, Ni J and Lin
X: miR-4319 hinders YAP expression to restrain non-small cell lung
cancer growth through regulation of LIN28-mediated RFX5 stability.
Biomed Pharmacother. 115:1089562019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tan X, Fu Y, Chen L, Lee W, Lai Y Rezaei
K, Tabbara S, Latham P, Teal CB, Man YG, et al: miR-671-5p inhibits
epithelial-to-mesenchymal transition by downregulating FOXM1
expression in breast cancer. Oncotarget. 7:293–307. 2016.PubMed/NCBI
|
|
27
|
Chiu YC, Li MY, Liu YH, Ding JY, Yu JY and
Wang TW: Foxp2 regulates neuronal differentiation and neuronal
subtype specification. Dev Neurobiol. 74:723–738. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Katoh M and Katoh M: Human FOX gene family
(Review). Int J Oncol. 25:1495–1500. 2004.PubMed/NCBI
|
|
29
|
Tsui D, Vessey JP, Tomita H, Kaplan DR and
Miller FD: FoxP2 regulates neurogenesis during embryonic cortical
development. J Neurosci. 33:244–258. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Enard W, Przeworski M, Fisher SE, Lai CS,
Wiebe V, Kitano T, Monaco AP and Pääbo S: Molecular evolution of
FOXP2, a gene involved in speech and language. Nature. 418:869–872.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Herrero MJ and Gitton Y: The untold
stories of the speech gene, the FOXP2 cancer gene. Genes Cancer.
9:11–38. 2018.PubMed/NCBI
|
|
32
|
Campbell AJ, Lyne L, Brown PJ, Launchbury
RJ, Bignone P, Chi J, Roncador G, Lawrie CH, Gatter KC, Kusec R and
Banham AH: Aberrant expression of the neuronal transcription factor
FOXP2 in neoplastic plasma cells. Br J Haematol. 149:221–230. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yan X, Zhou H and Zhang T, Xu P, Zhang S,
Huang W, Yang L, Gu X, Ni R and Zhang T: Downregulation of FOXP2
promoter human hepatocellular carcinoma cell invasion. Tumour Biol.
36:9611–9619. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Song XL, Tang Y, Lei XH, Zhao SC and Wu
ZQ: miR-618 inhibits prostate cancer migration and invasion by
targeting FOXP2. J Cancer. 8:2501–2510. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen MT, Sun HF, Li LD, Zhao Y, Yang LP,
Gao SP and Jin W: Downregulation of FOXP2 promotes breast cancer
migration and invasion through TGFβ/SMAD signaling pathway. Oncol
Lett. 15:8582–8588. 2018.PubMed/NCBI
|
|
36
|
Cuiffo BG and Karnoub AE: Silencing FOXP2
in breast cancer cells promotes cancer stem cell traits and
metastasis. Mol Cell Oncol. 3:e10190222015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu J, Liu P, Tang H, Shuang Z, Qiu Q,
Zhang L, Song C, Liu L, Xie X and Xiao X: FOXP2 Promotes tumor
proliferation and metastasis by targeting GRP78 in triple-negative
breast cancer. Curr Cancer Drug Targets. 18:382–389. 2018.
View Article : Google Scholar : PubMed/NCBI
|