Chronic kidney disease (CKD) is becoming a serious
public health problem because it is related to an increased risk of
cardiovascular disease and mortality (1–4). As
one of the main types of CKD, membranous nephropathy (MN), is an
autoimmune disease characterized by diffuse deposition of immune
complexes under glomerular epithelial cells (GECs), with diffuse
thickening of the glomerular basement membrane (GBM) (5,6).
According to primary diseases, MN can be categorized into
idiopathic membranous nephropathy (IMN) and secondary membranous
nephropathy (SMN), while IMN accounts for approximately 80% of the
total incidence of MN (7). In
recent years, the morbidity of IMN in China significantly increased
from 6.48% between 1997 and 1999 to 22.79% between 2009 and 2011
(8–11). The clinical prognosis of IMN varies
greatly. Among untreated patients, approximately one third undergo
spontaneous remission, one-third progress to end-stage renal
disease over 10 years and the remainder develop non-progressive
CKD. In short, approximately two-thirds of cases progress to CKD
(12–15). Thus far, no curative therapies have
been achieved for CKD or IMN (16,17),
which may be attributed to unclear mechanisms. Therefore, the
establishment of ideal model for IMN is important for mechanism
research and a number of studies have tried to establish new models
(18–21). This review comprehensively
discussed the available animal and cell models for IMN. The
limitations and advantages of the current models were discussed and
two improved models were provided. Overall, this review could
enable the selection of an appropriate model for studies on IMN and
further the current understanding of human IMN.
Heymann nephritis, also known as homologous immune
complex nephritis, is a widely used model of MN (22). On the basis of whether the
antibodies are autologous or homologous or not, Heymann nephritis
is divided into active and passive nephritis models (23). In 1959, Heymann et al
(24) used autogenous or
homologous rat homogenized proximal tubular brush border in immune
rats to develop a model of nephritic syndrome, known as active
Heymann nephritis (AHN). AHN is characterized by granular
glomerular capillary wall deposits of rat immunoglobulin G (IgG)
and subepithelial electron-dense deposits after 3–4 weeks. It has
been demonstrated that 30–80% of AHN rats developed proteinuria
within 8–10 weeks after immunization (25). Another study demonstrated that
injected rats with resistance to the proximal tubule brush border
antigen (Fx1A) antibodies also showed IgG, C3 and C5b-9 depositions
under the glomerular epithelium and a significant level of
proteinuria (26). This is termed
passive Heymann nephritis. It indicates that the subepithelial
deposits are formed by the circulating antibodies combining with
the intrinsic antigen in the glomerulus rather than by the
circulating immune complex. In this model, subepithelial
electron-dense deposits should be detectable after 3–5 days of
injecting anti-FxlA and rats developed persistent proteinuria after
about 7–10 days (27).
The pathogenesis of Heymann nephritis has been a
controversial issue for a long time. At present, its antigen,
mainly megalin (gp330), is believed to exist in the brush border of
the proximal convoluted tubule and epithelial cell membrane of the
glomerulus (24,28–30).
In animal models, active immunization with megalin resulted in
immune complex deposition under the epithelium of the glomerulus,
without activation of C3 or C5b-9 and proteinuria. However, when
the animals were injected with antibodies against megalin
monoclonal antibody and complement regulatory proteins such as
cluster of differentiation (CD)59 and CR1-related gene/protein Y at
the same time, pathological proteinuria occurred (31). These studies confirm that the
membrane attack complex (MAC; C5b-9) formed by the activation of
the immune complex is the main inflammatory mediator of Heymann
nephritis and is closely related to the production of pathological
proteinuria (32,33). Furthermore, although megalin is
expressed in human podocytes (34,35),
it is not detected in the glomerular subepithelial immune complex
and no circulating anti-megalin antibodies are found in patients
with IMN (36). In addition, the
complement pathway and subclass of IgG in this model are still
unknown (37–39). Therefore, it is not equivalent to
human IMN.
Dipeptidyl peptidase IV (DPP IV) is identified as a
major antigen (gp108) of FxlA (40), which is mainly expressed on the
brush borders of renal tubules, intestinal microvilli and
glomerular capillary loops. After injecting rabbit anti-DPP IV in
the rats, the rabbit IgG was deposited in the glomerular capillary
loops for 4–8 h and proteinuria occurred within 8 h. After 2 days,
proteinuria peaked and then rapidly decreased. To find the target
antigen, serum DPP IV-depleted rats were used and similar results
were obtained. The results suggest that DPP IV located along the
glomerular capillary wall plays an important role in the induction
of proteinuria (40). Compared
with the model induced by gp330, this method induced the activation
of a urinary protein that appears transiently, no deposition of C3
and faster disappearance of IgG (41). Taken together, these efforts reveal
the target antigen in MN and the pathogenesis of the kidney disease
model.
Thrombospondin type-1 domain-containing 7A (THSD7A),
which was identified in 2014, is one of the target podocyte
autoantibodies in IMN (42). These
receptors are type I transmembrane glycoproteins consisting of
three regions, namely the transmembrane domain, short intracellular
C-terminal tail and large extracellular domain (43,44).
In the human glomerulus, THSD7A is commonly expressed in foot
processes, with observation of 250 kDa proteins (45) in non-reducing states. The intrinsic
antigen respectively binds to anti-THSD7A IgG from the serum of the
patient with MN, forming immune complexes in situ along with
the glomerular filtration barrier (GFB) (46).
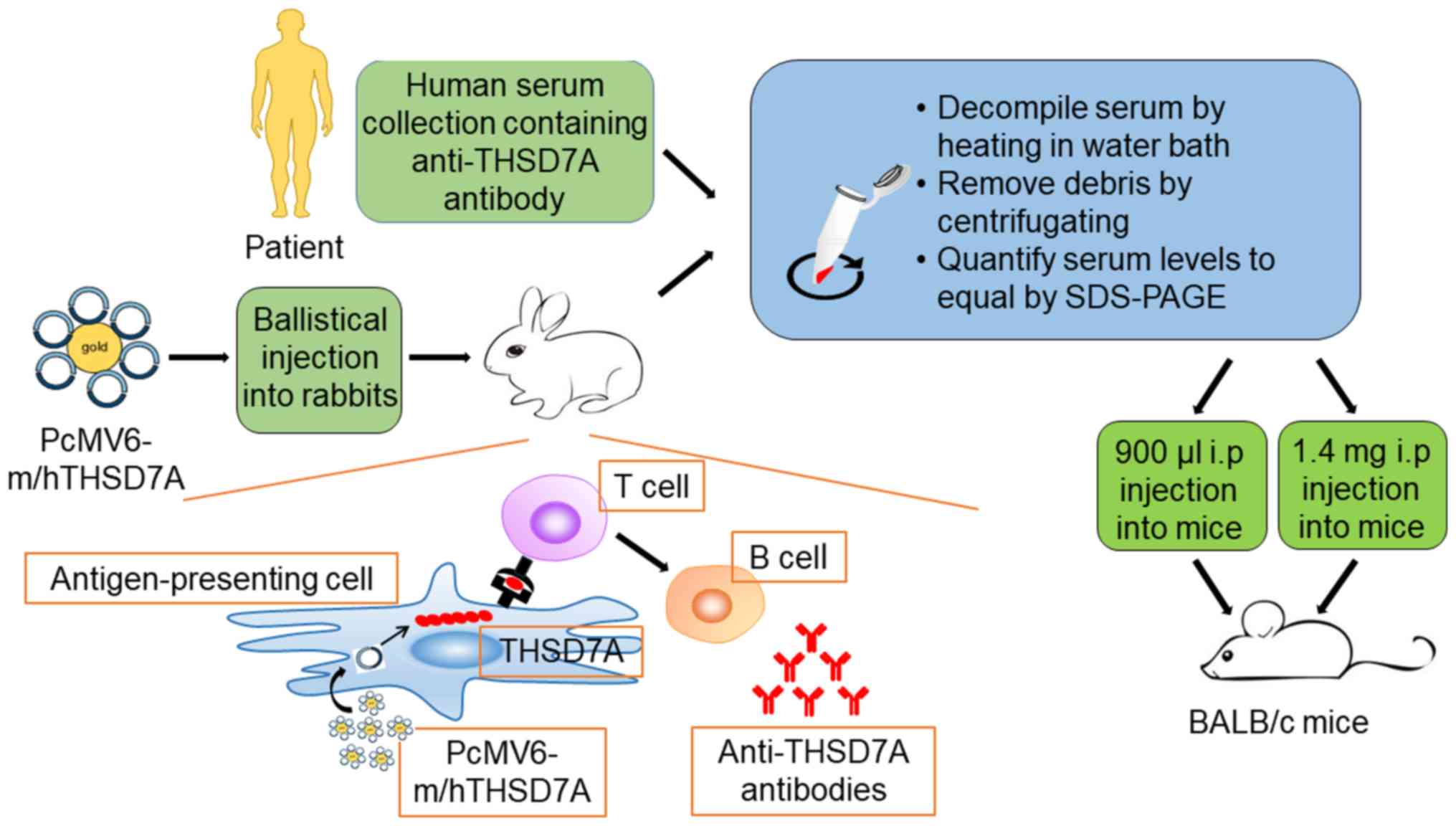
Previous findings have demonstrated that THSD7A is
strongly expressed in murine podocyte (47). This has enabled the establishment
of a model of THSD7A-related MN, which is closer to human IMN
(Fig. 1). Tomas et al
(20) injected human anti-THSD7A
antibody-containing serum in mice and demonstrated that human
anti-THSD7A autoantibodies can specifically bind to mouse
(m)THSD7A. The mice manifest significant albuminuria around day 3
and until day 70. The researchers observed a granular pattern on
the subepithelial aspect of the GFB, huIgG-mTHSD7A colocalization
and immune complex deposition. In addition, immunofluorescence
staining analyses revealed complement C3 accumulations within the
subepithelial granular pattern. Consistent with the above-mentioned
results, electron microscopic images showed electron-dense deposits
with a strictly subepithelial localization and extensive foot
process effacement around the local regions.
In the following year, a heterologous model of
THSD7A-associated MN was introduced (48). First, the researchers generated
rabbit anti-THSD7A antibodies by co-immunization with a combination
of mTHSD7A and human (h)THSD7A cDNAs. Then, the mice were injected
with anti-THSD7A IgG purified from rabbit serum. The mice that
received rabbit IgGs developed severe nephritic syndrome and the
albumin-to-creatinine ratio increased gradually during the entire
observation period of 14 days. The mice presented the same
histopathological changes as the abovementioned THSD7A-associated
model, with the only differences being the unknown IgG hypotype and
undetected C3 (Table I).
APA is a hydrolase present in the kidney of mice and
is mainly located on the podocyte membrane and the brush border of
proximal tubules. The model was established by injecting anti-APA
monoclonal antibodies in mice (49). The visible antibodies, belonging to
the IgG1 subclass, are widely distributed in the GEC membrane and
proximal tubular S1 and S2 brush borders. The dose-dependent
proteinuria lasted for 16 days and podocyte foot processes fused
but lacked activation of the complement system. Subsequently,
another study demonstrated that injection of anti-APA monoclonal
antibody can cause two slit-pore-associated proteins,
CD2-associated protein (CD2AP) and podocin structural alteration,
which may be the underlying mechanism of proteinuria (50).
According to the negative characteristic of the GBM,
injecting rabbits with cationic bovine serum albumin (C-BSA) daily
(pI>9.5) intravenously induced proteinuria and nephrotic
syndrome approximately 2 weeks later (51). Immunofluorescence showed IgG, C3
and other granule depositions in the GBM (52). Electron microscopy revealed
subepithelial electron-dense deposits and irregular GBM thickening
may occur in the later stages of the disease. To examine the
mechanism of this model, C-BSA was injected in the renal artery to
block the blood circulation after rinsing the rabbit kidney
thoroughly with saltwater and anti-BSA antibodies were subsequently
injected. Under immunofluorescence, immune complex particle sample
distribution was observed along the glomerular capillary wall,
which provided evidence that the pathogenesis of the C-BSA model
did not involve circulating immune complex deposition in the
glomerulus but immune complex deposition in situ (53). Chen et al (54) believed that this model is due to
the C-BSA deposited in the GBM, which is full of negative charge
and the corresponding antibody (principally IgG1), produced by its
own form of immune complex in situ, activates the complement
pathway. Debiec et al (55)
found bovine serum albumin in subepithelial immune deposits in
children with MN, demonstrating C-BSA binding to the anionic
glomerular capillary wall and subsequent formation of IC in
situ. They hypothesized that C-BSA may be one of the main
pathogenic targets in childhood MN, especially in children aged
<5 years.
The non-collagen 1 domain of human α3 (IV) collagen
(α3NC1) is a normal constituent of the GBM. Zhang et al
(56) hypothesized that owing to
its higher isoelectric point (pI~9), it may be conducive to the
formation of GBM deposition, similar to the mechanism of C-BSA.
Zhang et al (56) used mice
immunized with α3NC1 to develop clinical and histopathological
features of IMN. The extent and quality of autoimmunity against
α3NC1 and T-cell responses are critical to the severity of
nephropathy (57). Compared with
other models, the prominent advantage is that regulation of gene
expression is possible since mice are used. Furthermore, the model
can be developed rapidly and reliably. Luo et al (58) found that factor B-null mice lacked
glomerular deposition of C3 and C5b-9 and did not develop
albuminuria. Albuminuria was reduced but not completely abolished
in the C5-deficient mice. It indicates that the alternative pathway
is necessary for pathogenic complement activation. Only the DBA/1
mice are associated with a high success rate for mouse modeling and
other mice have a certain degree of resistance to α3NC1 and cannot
be used for developing such models (59).
Owing to different preparation methods, the
abovementioned models all have limitations or advantages. Active
Heymann nephritis is an autoimmune disease model and more closely
resembles human membranous nephropathy, thus, it is more suitable
for the study of the immune mechanism. However, it usually has
large individual variability, which makes it unsuitable for studies
of pharmacodynamics and injury (25). The passive Heymann nephritis model
is widely used as it is relatively stable and its pathological
changes are similar to human MN. The anti-DPP IV model is different
from the pathological manifestation of human IMN due to C3
deficiency (47). The
THSD7A-associated MN model is comparable to human IMN as a result
of the common THSD7A antigen in mice and humans, similar to the
complement activation pathways (lectin pathway) (64) and the common subclass of IgG
(IgG4). However, the detection rate of THSD7A is approximately 3%
in all patients with IMN (65,66).
This model may not represent the pathogenesis and all pathological
changes of IMN. Further, it is similar to the Heymann nephritis
model, which is induced by the binding of antigens and antibodies.
The C-BSA model is a new tool for IMN study that has a relatively
low cost, simple operation and wide application. In addition, the
degree of pathological changes is related to the dose; thus, models
of different stages of IMN can be developed. The limitations of the
model include the high mortality due to several tail vein
injections and dosage. A unified standard to establish a model that
reflects conditions in young children is imperative. As in the
anti-aminopeptidase A, anti-α3NC1 mouse and APN models, although
these models show different pathogenesis that of human IMN and are
currently underutilized, they can still be used to examine the
pathogenesis of IMN. For example, a study suggested that the
incidence of IMN is closely associated with the lungs (67), as in the anti-α3NC1 mouse
model.
Research on zymosan began in the 1940s. It is a
non-specific immune system stimulatory substance, existing in the
yeast cell wall (68). Some
researchers used zymosan to assemble C5b-9 complement complexes,
which were developed to interfere with mouse serum to obtain ZAS.
Subsequently, ZAS was applied to stimulate podocytes to develop the
MN model in vitro (69–71)
(Fig. 3).
Zymosan can promote rapid C3 cleavage in the
alternate pathway (AP) through assembly and protection of the
amplification convertase on its surface. It is different from the
classic complement pathway (CP), in which the antigen-antibody
complexes bind and convert C1 to its activated state on cell
membranes (72). However, the
formation of C5b-9 by zymosan induces antibody-independent
complement activation in the fluid phase via the AP, which differs
from that assembled on cell membranes via the CP (73,74).
Zymosan, one of the major activators of the AP, provides a contact
surface that initiates the complement cascade process by directly
activating C3 in serum without apparent utilization of C1, C4 or C2
(72). Subsequently, the AP
surface-bound C3 and C5 convertases assembled on zymosan particles
initiate the production of the C5b-9 complex (75,76).
To assemble sublytic C5b-9 MAC in the fluid phase,
normal human serum is treated with zymosan as previously reported
(70,77). Then, the zymosan particles are
removed from the serum sample by centrifugation. In addition, the
output rate of C5b-9 can be measured using an anti-human C5b-9
monoclonal antibody, which can specifically bind with the C5b-9
complex and then be removed from serum by immunoprecipitation, as
previously described (70). The
alteration in the membrane integrity of podocytes could be
determined by measuring the release of the intracellular enzyme
lactate dehydrogenase (76).
At present, the pathogenesis of IMN is remains to be
elucidated. A number of studies have demonstrated the pathogenesis
of immunological damage (78–80).
IMN is believed to be an autoimmune disease. As a result, research
on IMN has shifted from verifying the Heymann nephritis hypothesis
to further studying immune mechanisms. The current research on IMN
mainly focused on complement activation, the specific antigen, IgG4
and gene.
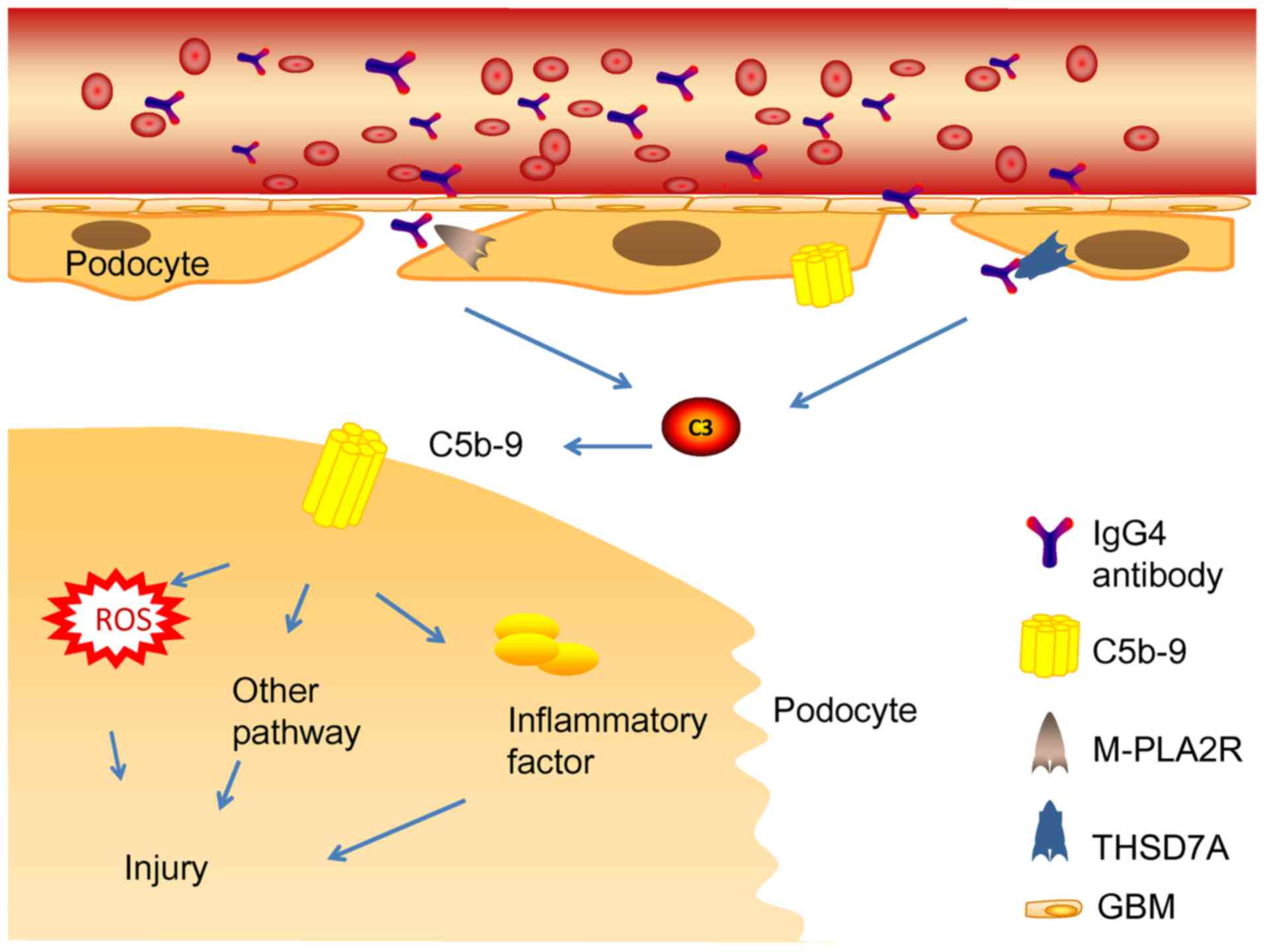
Glomerular subepithelial immune complexes play a
vital role in the pathogenetic mechanism of MN (14). The activation of the complement
system induces the production of MAC (C5b-9). Complement system
activation and MAC formation cause inflammatory mediator release
and podocyte damage (81)
(Fig. 4). Generally, immune
complexes are the major substances that activate the CP (82). Nevertheless, it remains
undiscovered as a pathway to complement system activation by IgG4
immune complexes in phospholipase A2 receptor 1 (PLA2R1) and
THSD7A-associated MN (83–85). IgG4 could hardly activate the CP
because of its low affinity to C1q or Fcγ receptors (86). Such studies support the hypothesis
that the lectin pathway may be the main initiation pathway
(87). In line with this,
mannose-binding lectin (MBL) deposition has been detected in the
subepithelial space of the glomerular capillaries (88). MBL may bind to specific
carbohydrates present in PLA2R or THSD7A, which leads to complement
system activation (89,90). Patients with C4 deficiency also
develop IMN, while the classic and lectin pathways cannot be
activated in patients with C4 deficiency. In addition, deposition
of factor B in IMN has been reported, which appears to be an
indirect evidence of the activation of the AP pathway (91).
The next focus is antigen exposure in human MN.
Several reported cases of MN might aid our understanding of the
research question. One example is the case of a 40-year-old female
patient with THSD7A-associated MN concomitant with mixed
adenoneuroendocrine carcinoma of the gallbladder. THSD7A was
detected in her gallbladder tumor, which corresponded to the
immunohistochemical test result. No anti-THSD7A antibodies were
detected in the plasma and her urinary protein level decreased
significantly after chemotherapy (92). Another example is that of two
patients with THSD7A-associated MN accompanied by angiolymphoid
hyperplasia with eosinophilia. THSD7A expression outside the kidney
may be vital to the pathogenesis of IMN (93). These cases seem to indicate that
antibodies produced outside the kidney help with the initial
recognition of antigens. However, the aforementioned cases have
limitations. Electron microscopy observation results were not
reported in the articles, so whether the diagnosis of SMN is
consistent is uncertain. In addition, a cross-sectional study
(94) found a significant
correlation between the incidence of IMN and exposure of
particulate matter of <2.5 (PM2.5). In areas with
severe air pollution, the incidence of IMN increased significantly.
PLA2R can be found in neutrophils and the lung (95,96).
In addition, PLA2R and THSD7A are disparate paths to the same
disease (97). Thus, the authors
of the present review may hypothesize that an unidentified
extrarenal cause exists in IMN or SMN.
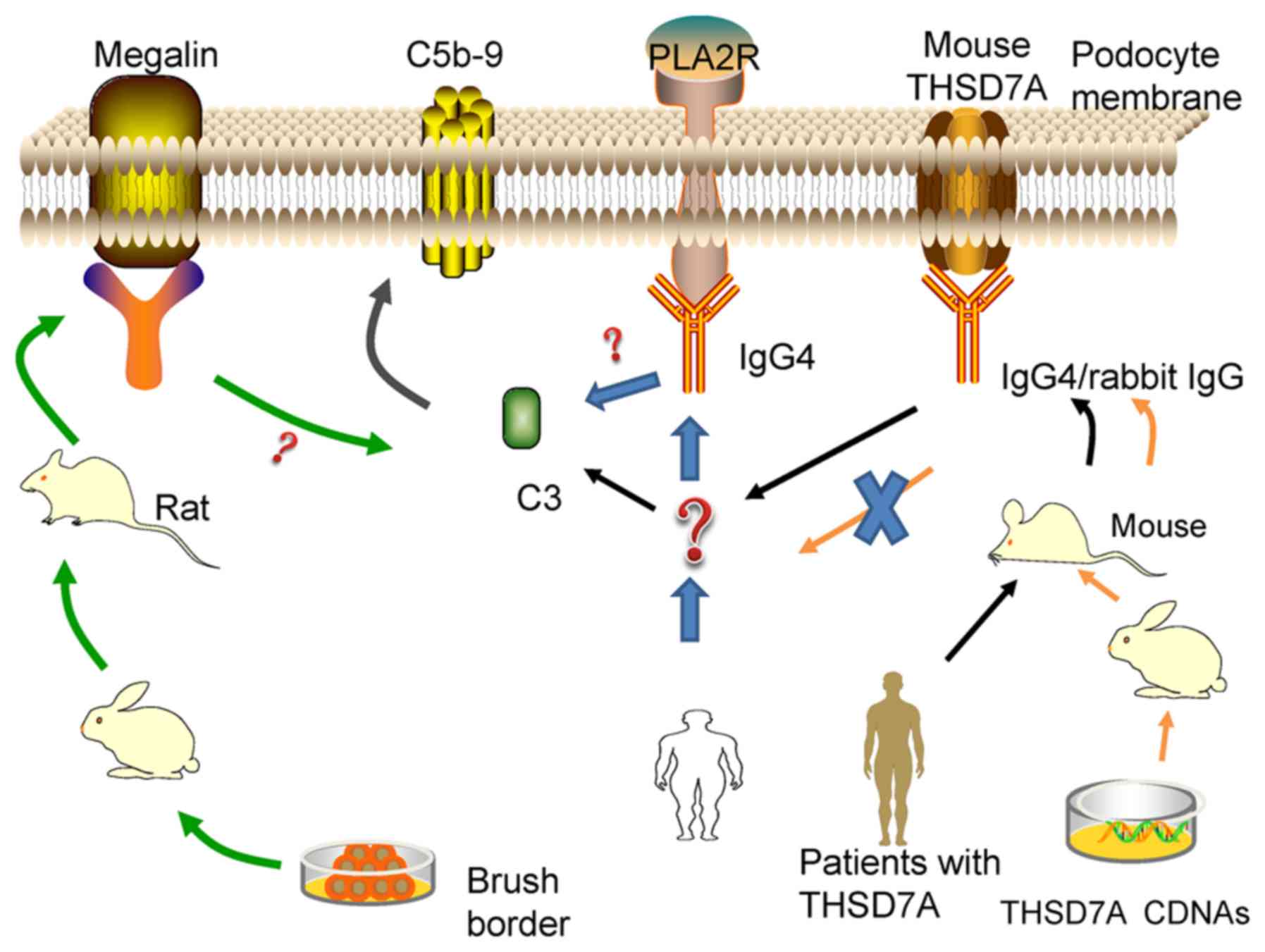
The authors of the present review hypothesized that
tumor tissues overexpress THSD7A, or PLA2R is expressed in
neutrophils and lung cells, which leads to complement system
activation and MAC formation (92,93,98).
However, the current models do not help us answer the question of
antigen production. Most current models were established by
injecting antibodies produced heterogeneously. This approach is not
helpful for examining the original antigens in human IMN. To
clarify whether an undiscovered mechanism exists, a more
appropriate model with PLA2R-expressing podocytes should be
established.
Finally, the reason that IgG4 is the most common
antibody deposited in the subepithelial immune complex in most
human IMN types remains unclear. IgG4 is related to inflammation
and autoimmune disorders (99),
but it is widely found in IMN. A previous study suggested that IgG4
could have a partially protective effect by acting on IgG1 or IgG3
(100). Another study suggested
that IgG autoantibodies transform from IgG1 to IgG4 during disease
progression (101). Probably
owing to the different antigens or ways antibodies are induced,
only few known models detect IgG4 (Table II). IgG4 is the main antibody of
the THSD7A-associated MN model. Whether purified anti-THSD7A IgG4
can form immune deposits in this model will likely advance IgG4
research
Evidence of the significant correlation between
specific genetic factors and IMN is accumulating (102,103). The human leukocyte antigen (HLA)
gene has a strong gene correlation with the PLA2R gene and
the interaction may be the basis of autoimmune injury (104). Although no relevant study has
described the polymorphism of the THSD7A gene, the combined
application of THSD7A and PLA2R gene tests is helpful
to further examine the correlation between the IMN pathogenesis and
genes, providing an important value for the establishment of the
genetic model of MN.
In human MN, three complement activation pathways
have been reported, but reports about each pathway are conflicting.
The common feature is that complement activation induces assembly
of the MAC (C5b-9) on podocytes, which is necessary for sublethal
damage to the renal intrinsic cells (22). Therefore, establishing a model with
MAC (C5b-9) podocytes may avoid the disadvantages of the difference
in pathways between the models and human MN (Fig. 6). First, sublytic C5b-9 is
assembled in vitro and then incubated with microbubbles that
carry cationic charges. The specific steps follow the protocol
provided by ultrasonic-targeted microbubble destruction (105,106). Then, the suspension of
microvesicles is injected immediately in mice through the tail
vein. Microvesicles are likely to contact the GBM and podocytes
because they carry cationic charges. Ultrasonic sensors are used to
scan the kidney repeatedly and when microbubbles are observed
traveling into the kidney, the ultrasonic pressure will be
increased to destroy the microbubbles, delivering C5b-9 to attack
podocytes.
Subsequently, the anti-PLA2R antibody can be
injected to induce IMN. In addition, as mentioned previously,
several extrarenal factors may activate the autoimmune system to
produce anti-PLA2R antibody. Previous findings have shown that
PLA2R-deficient mice had significantly expressed inflammatory cells
around the airways as compared with wild-type mice (110). Thus, using PM2.5 to
stimulate mice with podocytes expressing PLA2R, may expose PLA2R in
the lung or neutrophils and then generating antibodies to bind to
podocytes.
An ideal model is the one that will contribute to
understand the pathogenesis of IMN, so as to improve the diagnosis
and therapy. It is worth noting that increasing evidence has shown
the traditional Chinese medicine (TCM) is beneficial for both
antifibrosis therapy in CKD (111–113) and treating primary disease such
as IMN (114–116). However, their mechanisms have not
been elucidated. TCM could bring a new understanding of the
pathogenesis of IMN.
Not applicable.
The present study was supported by National Major
Scientific and Technological Special Project for ‘Significant New
Drugs Development’ (grant no. 2017ZX09304019).
All data generated or analyzed during this study are
included in this published article.
HXJ and ZF wrote the main parts of this manuscript;
ZBZ, CHX, WZ and JG wrote other parts and designed the figures; BLL
and YW improved the language of the manuscript; YNL and WJL
conceived the structure and revised the manuscript. All authors
read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Cernaro V, Coppolino G, Visconti L, Rivoli
L, Lacquaniti A, Santoro D, Buemi A, Loddo S and Buemi M:
Erythropoiesis and chronic kidney disease-related anemia: From
physiology to new therapeutic advancements. Med Res Rev.
39:427–460. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen DQ, Cao G, Chen H, Argyopoulos CP, Yu
H, Su W, Chen L, Samuels DC, Zhuang S, Bayliss GP, et al:
Identification of serum metabolites associating with chronic kidney
disease progression and anti-fibrotic effect of
5-methoxytryptophan. Nat Commun. 10:14762019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen DQ, Cao G, Zhao H, Chen L, Yang T,
Wang M, Vaziri ND, Guo Y and Zhao YY: Combined melatonin and
poricoic acid A inhibits renal fibrosis through modulating the
interaction of Smad3 and β-catenin pathway in AKI-to-CKD continuum.
Ther Adv Chronic Dis. 10:20406223198691162019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
MacKinnon HJ, Wilkinson TJ, Clarke AL,
Gould DW, O'Sullivan TF, Xenophontos S, Watson EL, Singh SJ and
Smith AC: The association of physical function and physical
activity with all-cause mortality and adverse clinical outcomes in
nondialysis chronic kidney disease: A systematic review. Ther Adv
Chronic Dis. 9:209–226. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xiaofan H, Jing X, Chenni G, Yifan W,
Xialian Y, Li L, Hong R, Wen Z, Weiming W, Xiaoxia P, et al: New
risk score for predicting progression of membranous nephropathy. J
Transl Med. 17:412019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Petrosyan A, Cravedi P, Villani V,
Angeletti A, Manrique J, Renieri A, De Filippo RE, Perin L and Da
Sacco S: A glomerulus-on-a-chip to recapitulate the human
glomerular filtration barrier. Nat Commun. 10:36562019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Glassock RJ: The pathogenesis of
idiopathic membranous nephropathy: A 50-year odyssey. Am J Kidney
Dis. 56:157–167. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu X, Wang G, Chen N, Lu T, Nie S, Xu G,
Zhang P, Luo Y, Wang Y, Wang X, et al: Long-term exposure to air
pollution and increased risk of membranous nephropathy in China. J
Am Soc Nephrol. 27:3739–3746. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu P, Zhou FD, Wang SX, Zhao MH and Wang
HY: Increasing frequency of idiopathic membranous nephropathy in
primary glomerular disease: A 10-year renal biopsy study from a
single Chinese nephrology centre. Nephrology (Carlton). 20:560–566.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pan X, Xu J, Ren H, Zhang W, Xu Y, Shen P,
Li X, Wang W, Chen X, Wu P, et al: Changing spectrum of
biopsy-proven primary glomerular diseases over the past 15 years: A
single-center study in China. Contrib Nephrol. 181:22–30. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen X, Chen Y, Shi K, Lv Y, Tong H, Zhao
G, Chen C, Chen B, Li D and Lu Z: Comparison of prognostic,
clinical, and renal histopathological characteristics of
overlapping idiopathic membranous nephropathy and IgA nephropathy
versus idiopathic membranous nephropathy. Sci Rep. 7:114682017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maisonneuve P, Agodoa L, Gellert R,
Stewart JH, Buccianti G, Lowenfels AB, Wolfe RA, Jones E, Disney
AP, Briggs D, et al: Distribution of primary renal diseases leading
to end-stage renal failure in the United States, Europe, and
Australia/New Zealand: Results from an international comparative
study. Am J Kidney Dis. 35:157–165. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Latt KZ, Honda K, Thiri M, Hitomi Y, Omae
Y, Sawai H, Kawai Y, Teraguchi S, Ueno K, Nagasaki M, et al:
Identification of a two-SNP PLA2R1 Haplotype and HLA-DRB1 Alleles
as primary risk associations in idiopathic membranous nephropathy.
Sci Rep. 8:155762018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Couser WG: Primary Membranous Nephropathy.
Clin J Am Soc Nephrol. 12:983–997. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Troyanov S, Wall CA, Scholey JW, Miller JA
and Cattran DC: Idiopathic membranous nephropathy: Definition and
relevance of a partial remission. Kidney International.
66:1199–1205. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cattran D and Brenchley P: Membranous
nephropathy: Thinking through the therapeutic options. Nephrol Dial
Transplant. 32:i22–i29. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fontecha-Barriuso M, Martin-Sanchez D,
Ruiz-Andres O, Poveda J, Sanchez-Niño MD, Valiño-Rivas L,
Ruiz-Ortega M, Ortiz A and Sanz AB: Targeting epigenetic DNA and
histone modifications to treat kidney disease. Nephrol Dial
Transplant. 33:1875–1886. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jefferson JA, Pippin JW and Shankland SJ:
Experimental models of membranous nephropathy. Drug Discov Today
Dis Models. 7:27–33. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Borza DB, Zhang JJ, Beck LH Jr,
Meyer-Schwesinger C and Luo W: Mouse models of membranous
nephropathy: The road less travelled by. Am J Clin Exp Immunol.
2:135–145. 2013.PubMed/NCBI
|
|
20
|
Tomas NM, Hoxha E, Reinicke AT, Fester L,
Helmchen U, Gerth J, Bachmann F, Budde K, Koch-Nolte F, Zahner G,
et al: Autoantibodies against thrombospondin type 1
domain-containing 7A induce membranous nephropathy. J Clin Invest.
126:2519–2532. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lim WH, Wong G, McDonald SP, Chakera A,
Luxton G, Isbel NM, Pilmore HL, Barbour T, Hughes P and Chadban SJ:
Long-term outcomes of kidney transplant recipients with end-stage
kidney disease attributed to presumed/advanced glomerulonephritis
or unknown cause. Sci Rep. 8:90212018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cybulsky AV, Quigg RJ and Salant DJ:
Experimental membranous nephropathy redux. Am J Physiol Renal
Physiol. 289:F660–F671. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ma H, Sandor DG and Beck LH Jr: The role
of complement in membranous nephropathy. Semin Nephrol. 33:531–542.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Heymann W, Hackel DB, Harwood S, Wilson SG
and Hunter JL: Production of nephrotic syndrome in rats by Freund's
adjuvants and rat kidney suspensions. Proc Soc Exp Biol Med.
100:660–664. 1959. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Salant DJ, Quigg RJ and Cybulsky AV:
Heymann nephritis: Mechanisms of renal injury. Kidney Int.
35:976–984. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Christiansen RE, Kolmannskog O, Leh S,
Iversen BM and Tenstad O: Glomerular charge barrier and development
of proteinuria in passive Heymann nephritis. Kidney Blood Press
Res. 31:203–209. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Salant DJ and Cybulsky AV: Experimental
glomerulonephritis. Methods Enzymol. 162:421–461. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kerjaschki D and Farquhar MG: The
pathogenic antigen of Heymann nephritis is a membrane glycoprotein
of the renal proximal tubule brush border. Proc Natl Acad Sci USA.
79:5557–5561. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kerjaschki D and Farquhar MG:
Immunocytochemical localization of the Heymann nephritis antigen
(GP330) in glomerular epithelial cells of normal Lewis rats. J Exp
Med. 157:667–686. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kerjaschki D, Ullrich R, Diem K,
Pietromonaco S, Orlando RA and Farquhar MG: Identification of a
pathogenic epitope involved in initiation of Heymann nephritis.
Proc Natl Acad Sci USA. 89:11179–11183. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Raychowdhury R, Zheng G, Brown D and
McCluskey RT: Induction of Heymann nephritis with a gp330/megalin
fusion protein. Am J Pathol. 148:1613–1623. 1996.PubMed/NCBI
|
|
32
|
Couser WG: Mediation of immune glomerular
injury. J Am Soc Nephrol. 1:13–29. 1990.PubMed/NCBI
|
|
33
|
Baker PJ, Ochi RF, Schulze M, Johnson RJ,
Campbell C and Couser WG: Depletion of C6 prevents development of
proteinuria in experimental membranous nephropathy in rats. Am J
Pathol. 135:185–194. 1989.PubMed/NCBI
|
|
34
|
Prabakaran T, Nielsen R, Larsen JV,
Sørensen SS, Feldt-Rasmussen U, Saleem MA, Petersen CM, Verroust PJ
and Christensen EI: Receptor-mediated endocytosis of
α-galactosidase A in human podocytes in Fabry disease. PLoS One.
6:e250652011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Larsen C, Etzerodt A, Madsen M, Skjodt K,
Moestrup SK and Andersen CBF: Structural assembly of the
megadalton-sized receptor for intestinal vitamin B12
uptake and kidney protein reabsorption. Nat Commun. 9:52042018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ronco P and Debiec H: Pathophysiological
advances in membranous nephropathy: Time for a shift in patient's
care. Lancet. 385:1983–1992. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vinaiphat A and Thongboonkerd V:
Characterizations of PMCA2-interacting complex and its role as a
calcium oxalate crystal-binding protein. Cell Mol Life Sci.
75:1461–1482. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Beausang JF, Fan HC, Sit R, Hutchins MU,
Jirage K, Curtis R, Hutchins E, Quake SR and Yabu JM: B cell
repertoires in HLA-sensitized kidney transplant candidates
undergoing desensitization therapy. J Transl Med. 15:92017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rudkin FM, Raziunaite I, Workman H, Essono
S, Belmonte R, MacCallum DM, Johnson EM, Silva LM, Palma AS, Feizi
T, et al: Single human B cell-derived monoclonal anti-Candida
antibodies enhance phagocytosis and protect against disseminated
candidiasis. Nat Commun. 9:52882018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Natori Y, Shindo N and Natori Y:
Proteinuria induced by anti-dipeptidyl peptidase IV (gp108); role
of circulating and glomerular antigen. Clin Exp Immunol.
95:327–332. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hunter JL, Hackel DB and Heymann W:
Nephrotic syndrome in rats produced by sensitization to rat kidney
proteins: Immunologic studies. J Immunol. 85:319–327.
1960.PubMed/NCBI
|
|
42
|
Tomas NM, Beck LH Jr, Meyer-Schwesinger C,
Seitz-Polski B, Ma H, Zahner G, Dolla G, Hoxha E, Helmchen U,
Dabert-Gay AS, et al: Thrombospondin type-1 domain-containing 7A in
idiopathic membranous nephropathy. N Engl J Med. 371:2277–2287.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Glassock RJ: Pathogenesis of membranous
nephropathy: A new paradigm in evolution. Contrib Nephrol.
181:131–142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tan K, Duquette M, Liu JH, Dong Y, Zhang
R, Joachimiak A, Lawler J and Wang JH: Crystal structure of the
TSP-1 type 1 repeats: A novel layered fold and its biological
implication. J Cell Biol. 159:373–382. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Allison SJ: Glomerular disease:
Thrombospondin type-1 domain-containing 7A-a new player in
membranous nephropathy. Nat Rev Nephrol. 11:632015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
De Vriese AS, Glassock RJ, Nath KA, Sethi
S and Fervenza FC: A Proposal for a serology-based approach to
membranous nephropathy. J Am Soc Nephrol. 28:421–430. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Godel M, Grahammer F and Huber TB:
Thrombospondin type-1 domain-containing 7A in idiopathic membranous
nephropathy. N Engl J Med. 372:10732015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tomas NM, Meyer-Schwesinger C, von Spiegel
H, Kotb AM, Zahner G, Hoxha E, Helmchen U, Endlich N, Koch-Nolte F
and Stahl RAK: A Heterologous model of thrombospondin type 1
domain-containing 7A-associated membranous nephropathy. J Am Soc
Nephrol. 28:3262–3277. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Assmann KJ, van Son JP, Dijkman HB and
Koene RA: A nephritogenic rat monoclonal antibody to mouse
aminopeptidase A. Induction of massive albuminuria after a single
intravenous injection. J Exp Med. 175:623–635. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Dijkman HB, Gerlofs-Nijland ME, van der
Laak JA, Wetzels JF, Groenen PJ and Assmann KJ: Podocyte changes
after induction of acute albuminuria in mice by anti-aminopeptidase
A mAb. Nephron Exp Nephrol. 94:e85–e93. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Border WA, Ward HJ, Kamil ES and Cohen AH:
Induction of membranous nephropathy in rabbits by administration of
an exogenous cationic antigen. J Clin Invest. 69:451–461. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Liu B, Lu R, Li H, Zhou Y, Zhang P, Bai L,
Chen D, Chen J, Li J, Yu P, et al: Zhen-wu-tang ameliorates
membranous nephropathy rats through inhibiting NF-κB pathway and
NLRP3 inflammasome. Phytomedicine. 59:1529132019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Adler SG, Wang H, Ward HJ, Cohen AH and
Border WA: Electrical charge. Its role in the pathogenesis and
prevention of experimental membranous nephropathy in the rabbit. J
Clin Invest. 71:487–499. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chen JS, Chen A, Chang LC, Chang WS, Lee
HS, Lin SH and Lin YF: Mouse model of membranous nephropathy
induced by cationic bovine serum albumin: Antigen dose-response
relations and strain differences. Nephrol Dial Transplant.
19:2721–2728. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Debiec H, Lefeu F, Kemper MJ, Niaudet P,
Deschênes G, Remuzzi G, Ulinski T and Ronco P: Early-childhood
membranous nephropathy due to cationic bovine serum albumin. N Engl
J Med. 364:2101–2110. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang JJ, Malekpour M, Luo W, Ge L, Olaru
F, Wang XP, Bah M, Sado Y, Heidet L, Kleinau S, et al: Murine
membranous nephropathy: Immunization with α3(IV) collagen fragment
induces subepithelial immune complexes and FcγR-independent
nephrotic syndrome. J Immunol. 188:3268–3277. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Hopfer H, Hunemorder S, Treder J, Turner
JE, Paust HJ, Meyer-Schwesinger C, Hopfer U, Sachs M, Peters A,
Bucher-Kocaoglu B, et al: Glomerulopathy induced by immunization
with a peptide derived from the goodpasture antigen α3IV-NC1. J
Immunol. 194:3646–3655. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Luo W, Olaru F, Miner JH, Beck LH Jr, van
der Vlag J, Thurman JM and Borza DB: Alternative pathway is
essential for glomerular complement activation and proteinuria in a
mouse model of membranous nephropathy. Front Immunol. 9:14332018.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Hopfer H, Maron R, Butzmann U, Helmchen U,
Weiner HL and Kalluri R: The importance of cell-mediated immunity
in the course and severity of autoimmune anti-glomerular basement
membrane disease in mice. FASEB J. 17:860–868. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Meyer TN, Schwesinger C, Wahlefeld J,
Dehde S, Kerjaschki D, Becker JU, Stahl RA and Thaiss F: A new
mouse model of immune-mediated podocyte injury. Kidney Int.
72:841–852. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Tsai SF, Wu MJ and Chen CH: Low serum C3
level, high neutrophil-lymphocyte-ratio, and high
platelet-lymphocyte-ratio all predicted poor long-term renal
survivals in biopsy-confirmed idiopathic membranous nephropathy.
Scie Rep. 9:62092019. View Article : Google Scholar
|
|
62
|
Meyer-Schwesinger C, Dehde S, Klug P,
Becker JU, Mathey S, Arefi K, Balabanov S, Venz S, Endlich KH,
Pekna M, et al: Nephrotic syndrome and subepithelial deposits in a
mouse model of immune-mediated anti-podocyte glomerulonephritis. J
Immunol. 187:3218–3229. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Beck LH Jr, Bonegio RG, Lambeau G, Beck
DM, Powell DW, Cummins TD, Klein JB and Salant DJ: M-type
phospholipase A2 receptor as target antigen in idiopathic
membranous nephropathy. N Engl J Med. 361:11–21. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Debiec H and Ronco P: Immune response
against autoantigen PLA2R is not gambling: Implications for
pathophysiology, prognosis, and therapy. J Am Soc Nephrol.
27:1275–1277. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Pandey P, Roy KK, Liu H, Ma G, Pettaway S,
Alsharif WF, Gadepalli RS, Rimoldi JM, McCurdy CR, Cutler SJ and
Doerksen RJ: Structure-based identification of potent natural
product chemotypes as cannabinoid receptor 1 inverse agonists.
Molecules. 23(pii): E26302018. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang J, Cui Z, Lu J, Probst C, Zhang YM,
Wang X, Qu Z, Wang F, Meng LQ, Cheng XY, et al: Circulating
antibodies against thrombospondin type-I domain-containing 7A in
Chinese patients with idiopathic membranous nephropathy. Clin J Am
Soc Nephrol. 12:1642–1651. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Liu W, Gao C, Dai H, Zheng Y, Dong Z, Gao
Y, Liu F, Zhang Z, Liu Z, Liu W, et al: Immunological pathogenesis
of membranous nephropathy: Focus on PLA2R1 and Its role. Front
Immunol. 10:18092019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Song JS, Kim YJ, Han KU, Yoon BD and Kim
JW: Zymosan and PMA activate the immune responses of Mutz3-derived
dendritic cells synergistically. Immunol Lett. 167:41–46. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Gawryl MS, Simon MT, Eatman JL and Lint
TF: An enzyme-linked immunoabsorbent assay for the quantitation of
the terminal complement complex from cell membranes or in activated
human sera. J Immunol Methods. 95:217–225. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Ishikawa S, Tsukada H and Bhattacharya J:
Soluble complex of complement increases hydraulic conductivity in
single microvessels of rat lung. J Clin Invest. 91:103–109. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Liu WJ, Li ZH, Chen XC, Zhao XL, Zhong Z,
Yang C, Wu HL, An N, Li WY and Liu HF: Blockage of the
lysosome-dependent autophagic pathway contributes to complement
membrane attack complex-induced podocyte injury in idiopathic
membranous nephropathy. Sci Rep. 7:86432017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Fearon DT and Austen KF: Activation of the
alternative complement pathway due to resistance of zymosan-bound
amplification convertase to endogenous regulatory mechanisms. Proc
Natl Acad Sci USA. 74:1683–1687. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Tegla CA, Cudrici C, Patel S, Trippe R
III, Rus V, Niculescu F and Rus H: Membrane attack by complement:
The assembly and biology of terminal complement complexes. Immunol
Res. 51:45–60. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Harboe M, Garred P, Lindstad JK, Pharo A,
Müller F, Stahl GL, Lambris JD and Mollnes TE: The role of
properdin in zymosan- and Escherichia coli-induced complement
activation. J Immunol. 189:2606–2613. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Rawal N and Pangburn MK: C5 convertase of
the alternative pathway of complement. Kinetic analysis of the free
and surface-bound forms of the enzyme. J Biol Chem.
273:16828–16835. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Rawal N and Pangburn M: Formation of
high-affinity C5 convertases of the alternative pathway of
complement. J Immunol. 166:2635–2642. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhang MH, Fan JM, Xie XS, Deng YY, Chen
YP, Zhen R, Li J, Cheng Y and Wen J: Ginsenoside-Rg1 protects
podocytes from complement mediated injury. J Ethnopharmacol.
137:99–107. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhang C, Leng L, Zhang X, Zhao Y and Li Z:
Comprehensive identification of immune-associated biomarkers based
on network and mRNA expression patterns in membranous
glomerulonephritis. J Transl Med. 16:2102018. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Bruschi M, Petretto A, Santucci L, Vaglio
A, Pratesi F, Migliorini P, Bertelli R, Lavarello C, Bartolucci M,
Candiano G, et al: Neutrophil Extracellular Traps protein
composition is specific for patients with Lupus nephritis and
includes methyl-oxidized alphaenolase (methionine sulfoxide 93).
Scie Rep. 9:79342019. View Article : Google Scholar
|
|
80
|
Li LZ, Hu Y, Ai SL, Cheng L, Liu J, Morris
E, Li Y, Gou SJ and Fu P: The relationship between thyroid
dysfunction and nephrotic syndrome: A clinicopathological study.
Sci Rep. 9:64212019. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Pozdzik A, Brocheriou I, David C, Touzani
F, Goujon JM and Wissing KM: Membranous nephropathy and
anti-podocytes antibodies: Implications for the diagnostic workup
and disease management. Biomed Res Int. 2018:62810542018.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Borsos T: Immune complex mediated
activation of the classic complement pathway. Behring Inst Mitt.
93–101. 1989.PubMed/NCBI
|
|
83
|
Cattran DC and Brenchley PE: Membranous
nephropathy: Integrating basic science into improved clinical
management. Kidney Int. 91:566–574. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhang Q, Huang B, Liu X, Liu B, Zhang Y,
Zhang Z, Hua J, Fan Y, Hu L, Meng M, et al: Ultrasensitive
quantitation of anti-phospholipase A2 receptor antibody as a
diagnostic and prognostic indicator of idiopathic membranous
nephropathy. Sci Rep. 7:120492017. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Fresquet M, Jowitt TA, McKenzie EA, Ball
MD, Randles MJ, Lennon R and Brenchley PE: PLA2R binds
to the annexin A2-S100A10 complex in human podocytes. Sci Rep.
7:68762017. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Vidarsson G, Dekkers G and Rispens T: IgG
subclasses and allotypes: From structure to effector functions.
Front Immunol. 5:5202014. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Borza DB: Alternative pathway
dysregulation and the conundrum of complement activation by IgG4
immune complexes in membranous nephropathy. Front Immunol.
7:1572016. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Wang Z, Wen L, Dou Y and Zhao Z: Human
anti-thrombospondin type 1 domain-containing 7A antibodies induce
membranous nephropathy through activation of lectin complement
pathway. Biosci Rep. 38(pii): BSR201801312018. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Hayashi N, Okada K, Matsui Y, Fujimoto K,
Adachi H, Yamaya H, Matsushita M and Yokoyama H: Glomerular
mannose-binding lectin deposition in intrinsic antigen-related
membranous nephropathy. Nephrol Dial Transplant. 33:832–840. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Garred P, Genster N, Pilely K,
Bayarri-Olmos R, Rosbjerg A, Ma YJ and Skjoedt MO: A journey
through the lectin pathway of complement-MBL and beyond. Immunol
Rev. 274:74–97. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Bally S, Debiec H, Ponard D, Dijoud F,
Rendu J, Fauré J, Ronco P and Dumestre-Perard C: Phospholipase A2
Receptor-related membranous nephropathy and mannan-binding lectin
deficiency. J Am Soc Nephrol. 27:3539–3544. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Hoxha E, Wiech T, Stahl PR, Zahner G,
Tomas NM, Meyer-Schwesinger C, Wenzel U, Janneck M, Steinmetz OM,
Panzer U, et al: A mechanism for Cancer-associated membranous
nephropathy. N Engl J Med. 374:1995–1996. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Matsumoto A, Matsui I, Namba T, Sakaguchi
Y, Mizuno H, Shirayama Y, Shimada K, Hashimoto N, Doi Y, Yamaguchi
S, et al: VEGF-A links angiolymphoid hyperplasia with eosinophilia
(ALHE) to THSD7A membranous nephropathy: A report of 2 cases. Am J
Kidney Dis. 73:880–885. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Xu X, Wang G, Chen N, Lu T, Nie S, Xu G,
Zhang P, Luo Y, Wang Y, Wang X, et al: Long-term exposure to air
pollution and increased risk of membranous nephropathy in China. J
Am Soc Nephrol. 27:3739–3746. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Silliman CC, Moore EE, Zallen G, Gonzalez
R, Johnson JL, Elzi DJ, Meng X, Hanasaki K, Ishizaki J, Arita H, et
al: Presence of the M-type sPLA(2) receptor on neutrophils and its
role in elastase release and adhesion. Am J Physiol Cell Physiol.
283:C1102–C1113. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Granata F, Petraroli A, Boilard E, Bezzine
S, Bollinger J, Del Vecchio L, Gelb MH, Lambeau G, Marone G and
Triggiani M: Activation of cytokine production by secreted
phospholipase A2 in human lung macrophages expressing the M-type
receptor. J Immunol. 174:464–474. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Beck LH Jr: PLA2R and THSD7A: Disparate
paths to the same disease? J Am Soc Nephrol. 28:2579–2589. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Chiorazzo MG, Tunset HM, Popov AV,
Johansen B, Moestue S and Delikatny EJ: Detection and
differentiation of breast cancer Sub-types using a cPLA2α
activatable fluorophore. Sci Rep. 9:61222019. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Pan Q, Lan Q, Peng Y, Cai J, Zheng J,
Dickerson C, Xiao H and Liu HF: Nature, functions, and clinical
implications of IgG4 autoantibodies in systemic lupus erythematosus
and rheumatoid arthritis. Discov Med. 23:169–174. 2017.PubMed/NCBI
|
|
100
|
Salant DJ: Unmet challenges in membranous
nephropathy. Curr Opin Nephrol Hypertens. 28:70–76. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Borza DB: Alternative pathway
dysregulation and the conundrum of complement activation by IgG4
immune complexes in membranous nephropathy. Front Immunol.
7:1572016. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Liu D, Zhang J, Shi Y and Liu Z: Gene
polymorphism and risk of idiopathic membranous nephropathy. Life
Sci. 229:124–131. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Canadas-Garre M, Anderson K, McGoldrick J,
Maxwell AP and McKnight AJ: Genomic approaches in the search for
molecular biomarkers in chronic kidney disease. J Transl Med.
16:2922018. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Kamyshova ES, Bobkova IN, Gorelova IA,
Каkhsurueva PA and Filatova EE: Genetic determinants of the
development and course of membranous nephropathy. Ter Arkh.
90:105–111. 2018.PubMed/NCBI
|
|
105
|
Lan HY, Mu W, Tomita N, Huang XR, Li JH,
Zhu HJ, Morishita R and Johnson RJ: Inhibition of renal fibrosis by
gene transfer of inducible Smad7 using ultrasound-microbubble
system in rat UUO model. J Am Soc Nephrol. 14:1535–1548. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Fujii H, Li SH, Wu J, Miyagi Y, Yau TM,
Rakowski H, Egashira K, Guo J, Weisel RD and Li RK: Repeated and
targeted transfer of angiogenic plasmids into the infarcted rat
heart via ultrasound targeted microbubble destruction enhances
cardiac repair. Eur Heart J. 32:2075–2084. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Huang B, Wang L, Zhang Y, Zhang J, Zhang
Q, Xiao H, Zhou B, Sun Z, Cao YN, Chen Y, et al: A novel
Time-resolved Fluoroimmunoassay for the quantitative detection of
Antibodies against the phospholipase A2 receptor. Sci Rep.
7:460962017. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Pan Y, Wan J, Liu Y, Yang Q, Liang W,
Singhal PC, Saleem MA and Ding G: sPLA2 IB induces human podocyte
apoptosis via the M-type phospholipase A2 receptor. Sci Re.
4:66602014.
|
|
109
|
Lambeau G and Lazdunski M: Receptors for a
growing family of secreted phospholipases A2. Trends Pharmacol Sci.
20:162–170. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Tamaru S, Mishina H, Watanabe Y, Watanabe
K, Fujioka D, Takahashi S, Suzuki K, Nakamura T, Obata JE, Kawabata
K, et al: Deficiency of phospholipase A2 receptor exacerbates
ovalbumin-induced lung inflammation. J Immunol. 191:1021–1028.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Chen DQ, Feng YL, Cao G and Zhao YY:
Natural products as a source for antifibrosis therapy. Trends
Pharmacol Sci. 39:937–952. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Chen DQ, Hu HH, Wang YN, Feng YL, Cao G
and Zhao YY: Natural products for the prevention and treatment of
kidney disease. Phytomedicine. 50:50–60. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Wang M, Chen DQ, Chen L, Cao G, Zhao H,
Liu D, Vaziri ND, Guo Y and Zhao YY: Novel inhibitors of the
cellular renin-angiotensin system components, poricoic acids,
target Smad3 phosphorylation and Wnt/beta-catenin pathway against
renal fibrosis. Br J Pharmacol. 175:2689–2708. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Chen Y, Deng Y, Ni Z, Chen N, Chen X, Shi
W, Zhan Y, Yuan F, Deng W and Zhong Y: Efficacy and safety of
traditional Chinese medicine (Shenqi Particle) for patients with
idiopathic membranous nephropathy: A multicenter randomized
controlled clinical trial. Am J Kidney Dis. 62:1068–1076. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Zhang L, Li P, Xing CY, Zhao JY, He YN,
Wang JQ, Wu XF, Liu ZS, Zhang AP, Lin HL, et al: Efficacy and
safety of Abelmoschus manihot for primary glomerular disease: A
prospective, multicenter randomized controlled clinical trial. Am J
Kidney Dis. 64:57–65. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Liu S, Li X, Li H, Liang Q and Chen J and
Chen J: Comparison of tripterygium wilfordii multiglycosides and
tacrolimus in the treatment of idiopathic membranous nephropathy: A
prospective cohort study. BMC Nephrol. 16:2002015. View Article : Google Scholar : PubMed/NCBI
|