Introduction
Gout, a type of inflammation caused by the
disturbance of purine metabolism and deposition of monosodium urate
(MSU) crystals in synovial fluid and other tissues, is one of the
most common forms of inflammatory arthritis, usually in the
presence of prolonged hyperuricemia. A longitudinal study reported
that the prevalence of gout and hyperuricemia in China were 2.8 and
18.1%, respectively (1). The
incidence of gout and hyperuricemia are increasing due to obesity,
insulin resistance, metabolic syndrome, renal impairment,
cardiovascular disease and hypertension (2).
Dioscorea collettii is a traditional Chinese
medicine that been used for the treatment of inflammatory
conditions, such as gouty arthritis and hyperuricemia, in China for
several years (3). Total saponin
from Dioscorea collettii (TSD) extracted from
Dioscorea has been reported to have significant
anti-inflammatory, analgesic and anti-hyperuricemia effects. A
previous study in rats with chronic hyperuricemia revealed that TSD
exhibited its effects through the downregulation of solute carrier
family 22 member 12 and solute carrier family 2 member 6 and the
upregulation of solute carrier family 22 members 6 and 8 (4). A previous study (5) suggested that the monosodium urate
(MSU)-induced inflammatory response is dependent on the
inflammatory cytokine interleukin (IL)-1β. The IL-1-dependent
innate inflammatory phenotype relies on the formation of the
macromolecular NLR family pyrin domain containing 3 (NALP3)
inflammasome complex in response to the MSU ‘danger signal’
(6). Therefore, the NALP3
inflammasome may be a potential target of TSD in gouty arthritis.
The present demonstrated that TSD inhibited the secretion of
inflammatory cytokines, including IL-1β, IL-18 and tumor necrosis
factor (TNF)-α, in THP-1 macrophages treated with MSU crystals.
Furthermore, the present study revealed that TSD inhibited the
assembly of the NALP3 inflammasome and the activation of
caspase-1.
Materials and methods
Drug and reagents
Dioscorea rhizomes were purchased from The
First Affiliated Hospital of Anhui University of Chinese Medicine.
According to the literature, the saponins were extracted from
Dioscorea (4). The total
content of TSD in the extract of Dioscorea was 53.1%
(4). Urate sodium was purchased
from Sigma-Aldrich (Merck KGaA). Colchicine and rotenone were
purchased from Shanghai Aladdin Biochem Technology Co., Ltd. ELISA
kits for IL-1β (cat. no. F0179A), IL-18 (cat. no. F0138A) and TNF-α
(cat. no. F0121A) were purchased from Shanghai Fankewei Technology
Industry Co., Ltd (www.shfksc.com).
Preparation of MSU crystals
MSU was prepared according to the method of Huang
el al's study (7). Briefly,
1 g uric acid was dissolved in 200 ml boiling water and the
solution pH was adjusted to 7.2 with 1N NaOH. The solution was
cooled gradually by stirring at room temperature. The crystals were
collected by centrifugation at 3,000 × g at 4°C for 2 min and
settled at 4°C for 6 h. The crystals were evaporated and sterilized
by heating at 180°C for 2 h and stored in a sterile environment
until use. The crystals were suspended in PBS at a concentration of
50 mg/ml and sonicated 10 min in 40 kHz at room temperature. 10 min
to obtain rod-shaped crystals with uniform sizes (5–25 µm in
length). A Limulus amebocyte cell lysate assay (cat. no. L00350;
GenScript) was used to verify the absence of endotoxin in the
preparation. The assay was performed according to the
manufacturer's protocol.
Cell culture and drug treatments
The human THP-1 cell line was purchased from the
Type Culture Collection of the Chinese Academy of Sciences. THP-1
cells were cultured in RPMI-1640 medium (Hyclone; GE Healthcare),
containing 10% FBS (Zhejiang Tianhang Biotechnology Co., Ltd.). The
air in the cell incubator was humidified and contained 5%
CO2 and 95% air at 37°C. The medium was changed every 2
days. In order to certify the effect of macrophages on MSU
crystals, THP-1 cells were induced into macrophage-like cells.
THP-1 cells (2×106 cells/well) were seeded in six-well
culture plates and incubated with phorbol 12-myristate acetate
(PMA) from 25–200 ng/ml for 24 h, then cells were washed by PBS and
observed the morphology under an inverted light microscope at ×200
magnification. Images were captured of each well in at least 5
random fields, the result was calculated by the ratio of adhered or
pseudopodia-formed THP-1 cells to the total cells. The cells were
identified by morphology and cluster of differentiation (CD)11b
protein level was characteristic of macrophages.
Viability assays
To evaluate the effects of MSU crystals or TSD on
the viability of THP-1 macrophages, THP-1 macrophages were treated
with MSU (0, 25, 50, 100, 200, 300 and 400 µg/ml) or TSD (0, 0.1,
0.3, 1, 3, 10 and 30 µg/ml) for 24 h. The viability of THP-1
macrophages was examined by MTT assay and the formazan was
dissolved by DMSO (≥99.7%; Sigma-Aldrich; Merck KGaA). Every well
was measured at a wavelength of 490 nm (optical density at 490)
with the Thermo Varioskan Flash (Thermo Fisher Scientific, Inc.).
Cell viability was expressed as a percentage of control cells,
which were defined as 100% viable. All the assays were performed in
triplicate.
Inflammatory cytokine ELISAs
In order to investigate the most appropriate MSU
crystals concentration in THP-1 macrophages, cells were treated
with MSU crystals at different concentrations (0, 50, 100, 200, 300
and 400 µg/ml). In follow-up experiments, THP-1 macrophages were
treated with TSD (0.3, 1.0 and 3.0 µg/ml) or colchicines (0.5 and 5
µM) for 24 h prior to the stimulation with MSU crystals (400 µg/ml)
or rotenone (80 µM). The level of the inflammatory cytokines, such
as IL-1β, IL-18 and TNF-α in the supernatants of media were
quantitatively measured with ELISA kits as listed above. The ELISA
plates were measured using a Thermo Varioskan Flash (Thermo Fisher
Scientific, Inc.).
Western blot analysis
THP-1 macrophages grouping was the same as above.
Cells were lysed in RIPA (Beyotime Institute of Biotechnology)
buffer containing 1 mM PMSF (Beyotime Institute of Biotechnology).
Protein samples (30 µg/lane) extracted from cell lysate was
separated by 10% SDS-PAGE. The separated proteins in the gel were
transferred onto a polyvinylidene difluoride membrane (EMD
Millipore), blocked with 2.5% TBST milk [10 mM Tris HCl (pH 8.0),
150 mM NaCl, 0.5% Tween-20, 2.5% skim milk] for 2 h at room
temperature and probed with anti-CD11b antibody (cat. no. ab133357;
Abcam), anti-NALP3 antibody (D2P5E; cat. no. 13158; Cell Signaling
Technology, Inc.), anti-ASC antibody (AW5459; Abgent, Inc.)
anti-caspase-1 p20 antibody (D57A2; cat. no. 4199; Cell Signaling
Technology, Inc.), anti-caspase-1 antibody (cat. no. sc-2225; Santa
Cruz Biotechnology, Inc.), or anti-actin antibody (8H10D10; cat.
no. 3700; Cell Signaling Technology, Inc.), respectively. All
antibodies were used at 1:1,000 dilution at 4°C overnight. All
antibodies were diluted with TBST milk. The secondary antibodies,
1:10,000, (Alexa Fluor® 790 goat anti-rabbit IgG, cat.
no. 111-655-144 and Alex Fluor® 680 goat anti-mouse IgG;
cat. no. 115-625-146) were incubated with the membranes for 2 h at
room temperature and detected with an Odyssey® CLx
Infrared Imaging System (LI-COR Biosciences), and the grey value
were analyzed with ImageJ 1.52a (National Institutes of Health).
All the assays were performed in triplicate.
Assay of mRNA expression level of
NALP3 with reverse transcription (RT)-PCR
The THP-1 macrophage grouping was the same as above.
The mRNA expression levels of NALP3 gene were determined by RT-PCR.
Total RNA was extracted from THP-1 macrophages with different
treatments using the total RNA extraction kit. RT-PCR was performed
using SuperScript™ IV One-Step RT-PCR system (Thermo Fisher
Scientific, Inc.). The β-actin gene was used as an internal
reference for normalization of expression levels. The following
primer sequences were used: NALP3 (NALP3 forward,
5′-TTCTCTGATGAGGCCCAAG-3′ and NALP3 reverse,
5′-GGATCTTCATGAGGTAGTCAG-3′); and β-actin (β-actin forward,
5′-GAGACCTTCAACACCCCAGCC-3′ and β-actin reverse,
5′-GGATCTTCATGAGGTAGTCAG-3′). RT-PCR amplification (Thermo Fisher
Scientific, Inc.) was performed using a protocol of 50°C for 2 min
and 95°C for 10 min, then 95°C for 15 sec followed by 53°C for 1
min for 40 cycles. The analysis method of RT-PCR was carried out
using the 2−ΔΔCq method (8).
Statistical analysis
Data were expressed as the mean ± standard
deviation. The results were analyzed for statistical significance
using one-way ANOVA followed by Tukey's post hoc test. P<0.05
was considered to indicate a statistically significant difference.
All data analysis was performed with SPSS 17.0 software (IBM
Corp.).
Results
THP-1 cells are successfully
differentiated into macrophages by PMA
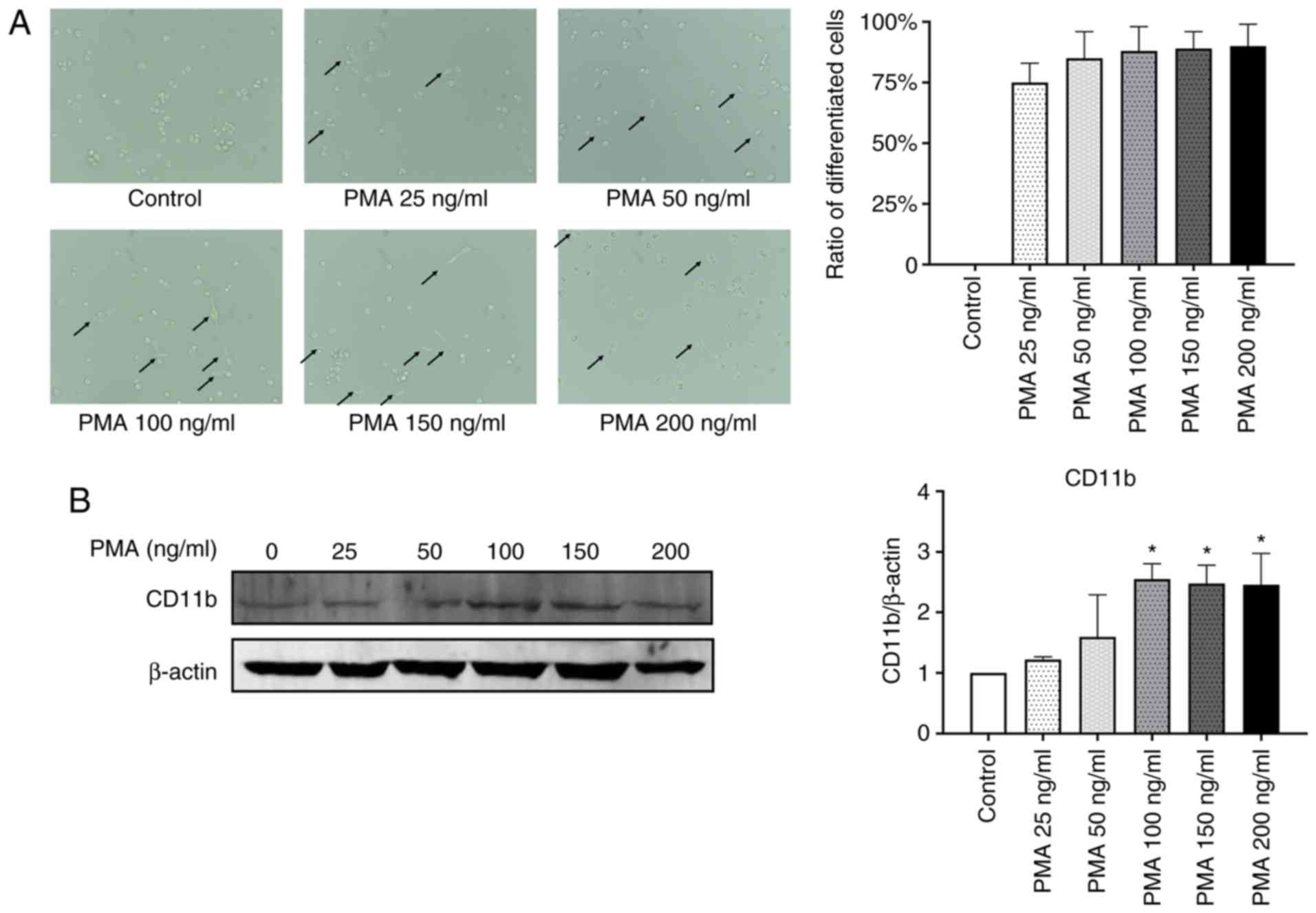
THP-1 cells were exposed to 25–200 ng/ml PMA for 24
h and examined under an inverted microscope (9,10).
As shown in Fig. 1A, THP-1 cells
treated with PMA at concentrations >25 ng/ml exhibited cell
adhesion, spreading and formed pseudopodia, suggesting that the
cells successfully differentiated into macrophages (11,12).
Furthermore, the level of CD11b, a macrophage surface marker
(13), was analyzed in the THP-1
macrophages. There was a significant increase in the CD11b protein
level in cells treated with PMA at concentrations exceeding 100
ng/ml (P<0.05; Fig. 1B).
Therefore, 100 ng/ml PMA was used in subsequent experiments.
Cytotoxic effect of MSU and TSD on
THP-1 macrophages
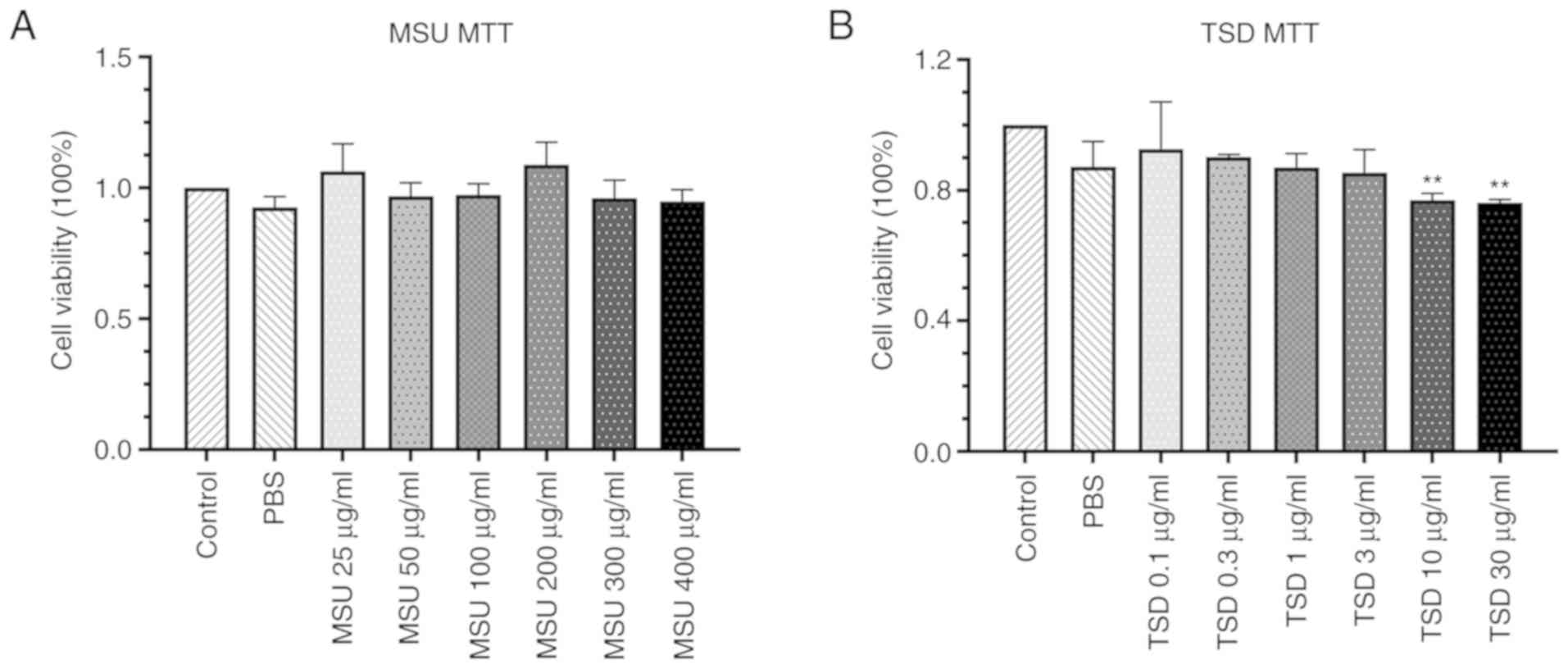
The cytotoxic effect of MSU and TSD on
differentiated THP-1 cells was measured using the MTT assay. As
shown in Fig. 2, no significant
changes in cell viability were observed in THP-1 macrophages
treated with MSU at concentrations up to 400 µg/ml or TSD at
concentrations up to 3 µg/ml for 24 h (48 or 72 h-treatment showed
the similar results with 24 h, Fig.
S1). Based on these results, MSU at a concentration of 400
µg/ml and TSD at a concentration range of 0.1–3 µg/ml were used for
further experimentation.
MSU induces THP-1 macrophages to
secrete IL-1β
Previous studies reported that macrophages respond
to MSU crystals during the progression of the gout inflammatory
response (14–16). MSU crystals subsequently lead to
the production of inflammatory cytokines such as IL-1β, which is a
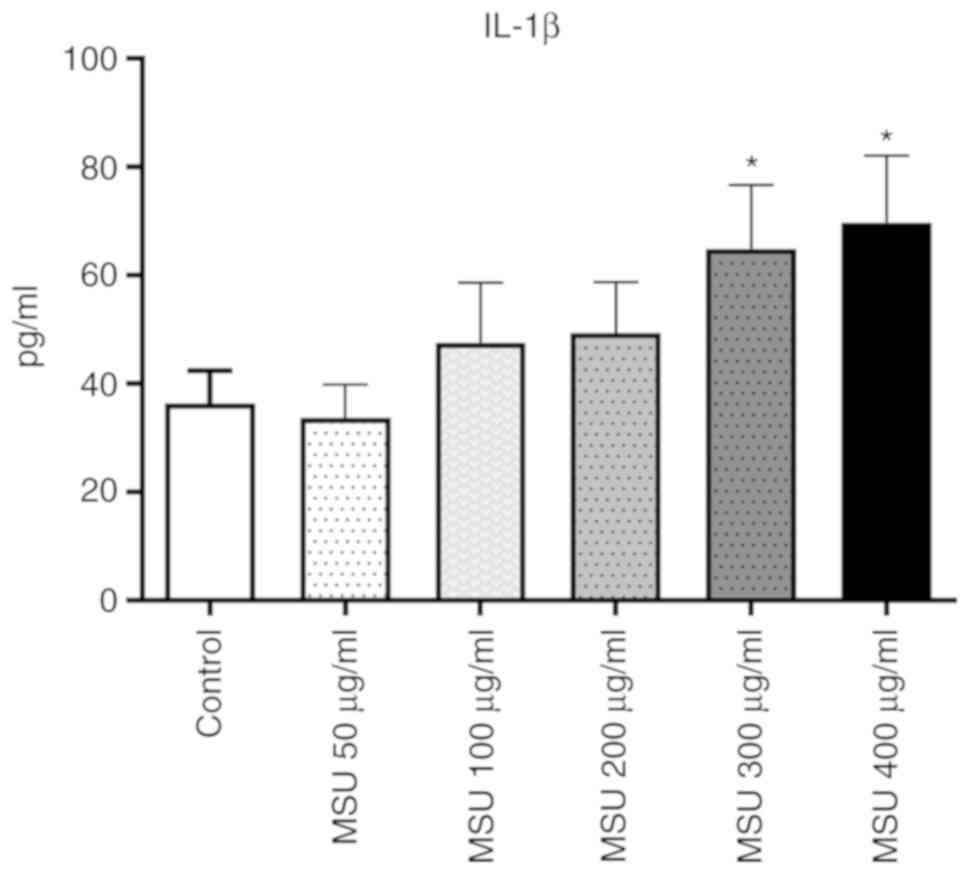
key regulatory cytokine in gout (6). The present study investigated whether
MSU induces the secretion of IL-1β in THP-1 macrophages. As shown
in Fig. 3, the level of IL-1β in
the cell culture medium increased ~1.79- and 1.92-fold following
treatment with MSU at concentrations of 300 and 400 µg/ml,
respectively. Based on these results, MSU at a concentration of 400
µg/ml was selected for subsequent experimentation.
TSD significantly inhibits MSU-induced
secretion of inflammatory cytokines in THP-1 macrophages
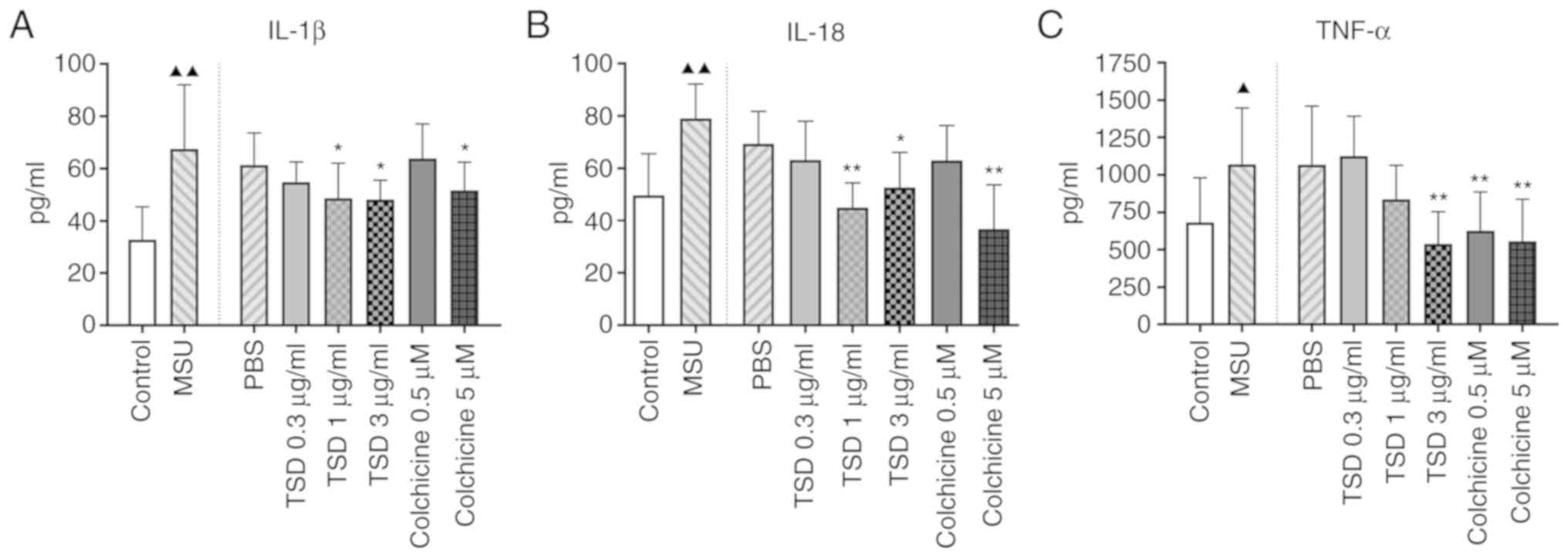
In order to investigate whether TSD inhibits the
production of inflammatory cytokines in THP-1 macrophages treated
with MSU crystals, cells were treated with or without TSD for 24 h
prior to stimulation with 400 µg/ml MSU. Cells treated with
colchicine (0.5 or 5 µM) served as the positive controls. As shown
in Fig. 4, treatment with MSU
crystals significantly increased the protein levels of IL-1β, IL-18
and TNF-α in THP-1 macrophages (P<0.05). However, the levels of
IL-1β decreased to ~72.2 and 71.2% in cells treated with 1 and 3
µg/ml TSD, respectively (P<0.01; Fig. 4A). The levels of IL-18
significantly decreased to ~57.0 and 66.6% in cells treated with 1
and 3 µg/ml TSD, respectively (P<0.01; Fig. 4B). The levels of TNF-α decreased to
~50.1% in cells treated with 3 µg/ml TSD (P<0.01; Fig. 4C). Cells treated with colchicine
exhibited similar results. These data indicated that TSD decreased
the secretion of IL-1β, IL-18 and TNF-α from THP-1 macrophages.
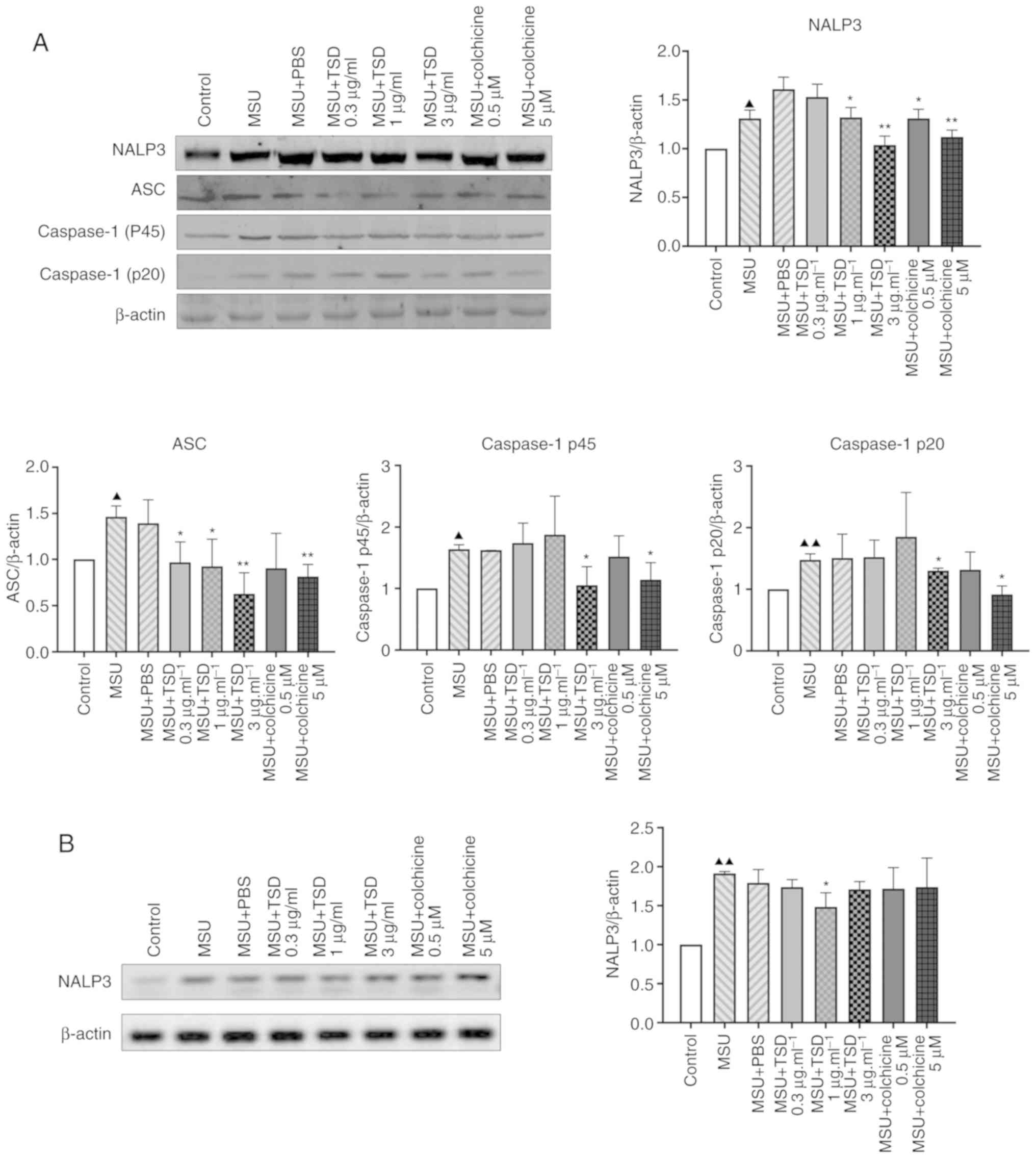
TSD inhibits the activation of the
NALP3 inflammasome
The release of IL-1β and IL-18 is dependent on the
NALP3 inflammasome and caspase-1 (17–19).
The present study therefore investigated whether TSD inhibited the
secretion of IL-1β via the NALP3 inflammasome. As NALP3 and
apoptosis-associated speck-like (ASC) serve an important role in
NALP3 inflammasome assembly (20,21),
the protein levels of NALP3 and ASC in TSD-treated THP-1
macrophages were measured in the current study. As shown in
Fig. 5A, the protein level of
NALP3 decreased to ~64.5 and 38.8% in cells treated with 1 and 3
µg/ml TSD compared with the MSU + PBS (control) group, respectively
(P<0.05). The protein level of ASC decreased to ~66.3, 63.2 and
42.9% in cells treated with 0.3, 1 and 3 µg/ml TSD, respectively
(P<0.05; Fig. 5A).
Additionally, the protein levels of caspase-1 p45 and p20, which
are activated by the NALP3 inflammasome, were investigated. As
presented in Fig. 5A, the protein
levels of caspase-1 p45 and p20 significantly decreased to ~64.4
and 88.1% in cells treated with 3 µg/ml TSD compared with the MSU +
PBS (control) group, respectively (P<0.05). The same trend was
observed in cells treated with colchicine (Fig. 5A). However, there were no
significant differences in the protein levels of NALP3, ASC,
caspase-1 p45 and p20 in cells treated with MSU + PBS compared with
the MSU group (Fig. 5A).
NALP3 is a critical inflammasome component which
recognize numerous exogenous and host ligands (14). Once activated, NALP3 recruits the
adapter ASC, which in turn recruits pro-caspase-1 (5,17).
It was reported that MSU crystals directly increased intercellular
NALP3 mRNA expression in human FLS cells (22). Additionally a previous study showed
that NALP3 transcript levels are increased after treatment of
macrophages for 6 h with MSU (23). Therefore it was investigated
whether TSD could inhibit the transcriptional level of NALP3. As
shown in Fig. 5B, the level of
NALP3 decreased in cells treated with 3 µg/ml TSD. However, no
significant difference in the level of NALP3 was observed in cells
treated with colchicine.
Mitochondrial reactive oxygen species (ROS) may be a
major trigger for the activation of the NLRP3 inflammasome and
production of inflammatory cytokines (6). The current study investigated the
effects of TSD in rotenone-induced THP-1 macrophages. Rotenone is a
respiration chain complex I inhibitor and induces mitochondrial ROS
production. As shown in Fig. 6,
THP-1 macrophages NALP3 inflammasome-associated protein levels were
decreased in THP-1 macrophages which were incubated with TSD before
and after incubation with rotenone for 6 h. The protein levels of
NALP3, caspase-1 p45 and caspase-1 p20 significantly decreased
~78.7, 38.7 and 47.4% in cells treated with 3 µg/ml TSD,
respectively, compared with rotenone + PBS-treated cells
(P<0.05). The protein level of ASC decreased ~45.0 and 61% in
cells treated with 1 and 3 µg/ml TSD, respectively.
Discussion
MSU deposition is the major source of inflammation
in gouty arthritis (24–26). Current treatment strategies for
gout remain suboptimal. Clinical studies have revealed that a
number of novel drugs, including IL-1R antagonists, may reduce the
pain and inflammation associated with gout (27–30).
Furthermore, several trials demonstrated that IL-1 inhibitors
prevent flares during the initial stages of allopurinol therapy
(31).
Dioscorea collettii has been widely used for
the treatment of gouty arthritis in traditional Chinese medicine.
TSD, the extract prepared from Dioscorea collettii, is
primarily used for the treatment of gout and hyperuricemia in rats
(32). However, mechanisms by
which TSD exerts its effects in gout have not been fully
elucidated. The present study therefore investigated the mechanism
of TSD in gouty arthritis.
Previous studies demonstrated that MSU crystals
recruit monocytes and promote their differentiation into
proinflammatory M1-like macrophages (33–35).
The resulting inflammation is likely to serve a role in the
deposition of MSU crystals in gout (36). A growing body of evidence suggests
that the MSU crystal-induced differentiation of monocytes into
macrophages is associated with the initiation, progression and
resolution of acute gouty inflammation (5,37).
In addition, it was reported that MSU crystals increase the
secretion of cytokines and chemokines, including TNF-α, IL-1β,
IL-18, IL-6, and IL-8, by macrophages and monocytes (38–40).
IL-1β and TNF-α have been implicated in the development of gout
flare-ups (41). A previous study
demonstrated increased levels of NALP3 mRNA and protein in
MSU-treated human fibroblast-like synoviocytes 6 to 48 h after
treatment (42). Additionally, MSU
crystals induced a significant increase in IL-1β in the culture
medium with a peak concentration at 6 h following MSU treatment
(21). The authors' previous study
revealed that 400 µg/ml MSU significantly increased the level of
IL-1β and NALP3 inflammasome activation in THP-1 macrophages 6 h
following treatment (43). Similar
results were obtained in the current study. Additionally, the
present study demonstrated that TSD inhibited the secretion of
inflammatory cytokines, including IL-1β, IL-18 and TNF-α, from
THP-1 macrophages treated with MSU crystals, suggesting that TSD
may exert a therapeutic effect in gout by decreasing the
inflammation induced by MSU.
A previous study reported that innate immunity is
involved in the pathogenesis of gout (44), suggesting that MSU crystals may be
recognized by pattern-recognition receptors (PRRs), such as
nucleotide-binding oligomerization domain-like receptors (NLRs) and
toll-like receptors (TLRs). NALP3 is a member of the NLR family,
intracellular PRRs that recognize pathogen-associated molecular
patterns and danger-associated molecular patterns (14).
MSU crystal-induced inflammation relies on the
activation of the NALP3 inflammasome, which consists of NALP3, ASC
and caspase-1 (17,45,46).
Following activation by MSU crystals, NALP3 assembles with the
adaptor protein ASC to form a protein-complex termed the NALP3
inflammasome. The NALP3 inflammasome subsequently cleaves caspase-1
to the active enzyme form. The activated caspase-1 cleaves
pro-IL-1β to IL-1β (47–49). Several studies have indicated that
the aberrant activation of the NALP3 inflammasome is associated
with the pathogenesis of autoimmune and chronic inflammatory and
metabolic diseases, including atherosclerosis, type 2 diabetes and
gout (50–53). A previous study revealed that
macrophages deficient of the NALP3 inflammasome components
caspase-1 and ASC exhibited significantly reduced MSU
crystals-induced inflammatory responses, and did not produce IL-1β.
This suggests that the NALP3 inflammasome and activated caspase-1
play an important role in MSU crystal-induced inflammation
(14). The current study revealed
that TSD decreased the protein levels of NALP3 and ASC in
MSU-treated THP-1 macrophages. Additionally, the present study
demonstrated that TSD inhibited the activation of caspase-1 in
THP-1 macrophages treated with MSU crystals. The aforementioned
results suggested that TSD may attenuate MSU-induced inflammation
by inhibiting the activation of the NALP3 inflammasome and
caspase-1. Zhou et al (54)
reported that the Chinese herbal medicine Dioscorea
nipponica decreased the activities of β-galactosidase, β-N
acetyl glucosamine, β-glucuronidase, acid phosphatase and
malonaldehyde and decreased the levels of TNF-α, IL-1β and IL-8 in
rats with gouty arthritis, suggesting that Dioscorea
nipponica decreased the extent of the self-limiting responses
by the NALP3 inflammasome.
ROS are major mediators of the NALP3/IL-1β signaling
pathway. Production of pro-inflammatory cytokines is associated
with the generation of ROS (55).
Mitochondria-and NADPH oxidase-derived ROS have been implicated in
pro-inflammatory microglia activation (56). As the abnormal activation of the
NALP3 inflammasome is linked to the pathogenesis of autoimmune and
chronic inflammatory and metabolic diseases, including
atherosclerosis, and type 2 diabetes and gout (50–52),
regulating the NALP3 inflammasome may prevent unwanted host damage
and excessive inflammation. Dysfunction of the mitochondrial
respiratory chain (MRC) is associated with the activation of the
NALP3 inflammasome partially due to the release of mitochondrial
ROS and DNA. Impairment of the MRC by rotenone confers a selective
priming signal for the activation of the NALP3 inflammasome
(57).
Ma et al (24) demonstrated that intracerebral
hemorrhage-induced inflammatory activation was associated with the
activation of the NALP3 inflammasome and revealed that rotenone
induced ROS production as well as NALP3 inflammasome activation.
Similarly, the present study demonstrated that rotenone induced the
activation of the NALP3 inflammasome, which was subsequently
suppressed by TSD. The aforementioned results suggested that ROS
may trigger the activation of the NALP3 inflammasome and that the
NALP3 inflammasome is a potential therapeutic target of TSD in
gouty arthritis.
While the present study suggested that TSD may exert
a therapeutic effect in gouty arthritis, future in vivo
experimental models are required to substantiate the results
obtained. Additionally, the effects of TSD on the expression of
TLRs and the pro-IL-1β or NALP3 signaling pathway warrant further
investigation, and these are the main areas of interest for the
next study.
In summary, the current study demonstrated that TSD
attenuated MSU-induced production of inflammatory cytokines, such
as TNF-α, IL-1β and IL-18, via inhibition of the NALP3 inflammasome
and caspase-1 in THP-1 macrophages. The NALP3 inflammasome and
caspase-1 may therefore serve as therapeutic targets for TSD in
gouty arthritis.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Ms. Guoying Li
(College of Integrative Medicine, Anhui University of Chinese
Medicine) for her help with extracting Dioscorea
collettii.
Funding
The current study was supported by the Fund of the
National Natural Science Foundation of China (grant no.
81573670).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article
Authors' contributions
GC and LL conceived and designed the study. LW, LZ
and CD performed the experiments and data analysis. LW, LZ and LL
wrote the manuscript. All authors reviewed, edited and approved the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
TSD
|
total saponins of Dioscorea
collettii
|
|
MSU
|
monosodium urate
|
|
MRC
|
mitochondrial respiratory chain
|
References
|
1
|
Lu X, Li X, Zhao Y, Zheng Z, Guan S and
Chan P: Contemporary epidemiology of gout and hyperuricemia in
community elderly in Beijing. Int J Rheum Dis. 17:400–407. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stamp LK and Chapman PT: Gout and its
comorbidities: Implications for therapy. Rheumatology (Oxford).
52:34–44. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pan BQ, Pan JK, Liu J and Yang JY:
Analysis on medication rule in herbal prescriptions for gout based
on apriori and clustering algorithm. China J Trad Chin Med
Pharmacy. 29:2040–2043. 2014.(In Chinese).
|
|
4
|
Zhu L, Dong Y, Na S, Han R, Wei C and Chen
G: Saponins extracted from Dioscorea collettii rhizomes
regulate the expression of urate transporters in chronic
hyperuricemia rats. Biomed Pharmacother. 93:88–94. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kingsbury SR, Conaghan PG and McDermott
MF: The role of the NLRP3 inflammasome in gout. J Inflamm Res.
4:39–49. 2011.PubMed/NCBI
|
|
6
|
Martinon F and Tschopp J: Inflammatory
caspases and inflammasomes: Master switches of inflammation. Cell
Death Differ. 14:10–22. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang HG, Sun YF, Hu M, Yuan GS, Wang Q
and Liu YX: Characteristics of monosodium urate monohydrate
crystal-induced acute arthritis in rats that mimicked human gouty
arthritis. Bulletin of the Academy of Military (Medicinal
Sciences). 29:538–542. 2005.
|
|
8
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kawakami A, Tani M, Chiba T, Yui K,
Shinozaki S, Nakajima K, Tanaka A, Shimokado K and Yoshida M:
Pitavastatin inhibits remnant lipoprotein-induced macrophage foam
cell formation through ApoB48 receptor-dependent mechanism.
Arterioscler Thromb Vasc Biol. 25:424–429. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Park EK, Jung HS, Yang HI, Yoo MC, Kim C
and Kim KS: Optimized THP-1 differentiation is required for the
detection of responses to weak stimuli. Inflamm Res. 56:45–50.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xian H, Wang P, Jing H, Chen GQ, Cheng DF,
Ji F, Song S and Zhang L: Comparative study of components and
anti-oxidative effects between sulfated polysaccharide and its iron
complex. Int J Biol Macromol. 118:1303–1309. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao Y, Zheng J, Yu Y and Wang L: Panax
notoginseng saponins regulate macrophage polarization under
hyperglycemic condition via NF-κB signaling pathway. Biomed Res
Int. 2018:92393542018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schwende H, Fitzke E, Ambs P and Dieter P:
Differences in the state of differentiation of THP-1 cells induced
by phorbol ester and 1,25-dihydroxyvitamin D3. J Leukoc Biol.
59:555–561. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Martinon F, Petrilli V, Mayor A, Tardivel
A and Tschopp J: Gout-associated uric acid crystals activate the
NALP3 inflammasome. Nature. 440:237–241. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chhana A, Pool B, Callon KE, Tay ML,
Musson D, Naot D, McCarthy G, McGlashan S, Cornish J and Dalbeth N:
Monosodium urate crystals reduce osteocyte viability and indirectly
promote a shift in osteocyte function towards a proinflammatory and
proresorptive state. Arthritis Res Ther. 20:2082018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sil P, Wicklum H, Surell C and Rada B:
Macrophage-derived IL-1 beta enhances monosodium urate
crystal-triggered NET formation. Inflamm Res. 66:227–237. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Eisenbarth SC, Colegio OR, O'Connor W,
Sutterwala FS and Flavell RA: Crucial role for the Nalp3
inflammasome in the immunostimulatory properties of aluminium
adjuvants. Nature. 453:1122–1126. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
van de Veerdonk FL, Netea MG, Dinarello CA
and Joosten LA: Inflammasome activation and IL-1β and IL-18
processing during infection. Trends Immunol. 32:110–116. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mankan AK, Dau T, Jenne D and Hornung V:
The NLRP3/ASC/Caspase-1 axis regulates IL-1β processing in
neutrophils. Eur J Immunol. 42:710–715. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wen H, Gris D, Lei Y, Jha S, Zhang L,
Huang MT, Brickey WJ and Ting JP: Fatty acid-induced NLRP3-ASC
inflammasome activation interferes with insulin signaling. Nat
Immunol. 12:408–415. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
He X, Mekasha S, Mavrogiorgos N,
Fitzgerald KA, Lien E and Ingalls RR: Inflammation and fibrosis
during Chlamydia pneumoniae infection is regulated by IL-1 and the
NLRP3/ASC inflammasome. J Immunol. 184:5743–5754. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zheng SC, Zhu XX, Xue Y, Zhang LH, Zou HJ,
Qiu JH and Liu Q: Role of the NLRP3 inflammasome in the transient
release of IL-1β induced by monosodium urate crystals in human
fibroblast-like synoviocytes. J Inflamm (Lond). 12:302015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gicquel T, Robert S, Loyer P, Victoni T,
Bodin A, Ribault C, Gleonnec F, Couillin I, Boichot E and Lagente
V: IL-1β production is dependent on the activation of purinergic
receptors and NLRP3 pathway in human macrophages. FASEB J.
29:4162–4173. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ma Q, Chen S, Hu Q, Feng H, Zhang JH and
Tang J: NLRP3 inflammasome contributes to inflammation after
intracerebral hemorrhage. Ann Neurol. 75:209–219. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kawai T and Akira S: The roles of TLRs,
RLRs and NLRs in pathogen recognition. Int Immunol. 21:317–337.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Landis RC and Haskard DO: Pathogenesis of
crystal-induced inflammation. Curr Rheumatol Rep. 3:36–41. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lamprecht P, Till A and Kabelitz D: New
aspects of the pathogenesis of gout. Danger signals,
autoinflammation and beyond. Z Rheumatol. 67:151–156. 2008.(In
German). View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Janssen CA, Oude Voshaar MAH, Vonkeman HE,
Jansen TLTA, Janssen M, Kok MR, Radovits B, van Durme C, Baan H and
van de Laar MAFJ: Anakinra for the treatment of acute gout flares:
A randomized, double-blind, placebo-controlled, active-comparator,
non-inferiority trial. Rheumatology (Oxford). 2019.(Epub ahead of
print). View Article : Google Scholar :
|
|
29
|
Sundy JS, Schumacher HR, Kivitz A,
Weinstein SP, Wu R, King-Davis S and Evans RR: Rilonacept for gout
flare prevention in patients receiving uric acid-lowering therapy:
results of RESURGE, a phase III, international safety study. J
Rheumatol. 41:1703–1711. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schlesinger N, Alten RE, Bardin T, et al:
Canakinumab for acute gouty arthritis in patients with limited
treatment options: Results from two randomised, multicentre,
active-controlled, double-blind trials and their initial
extensions. Ann Rheum Dis. 71:1839–1848. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Richette P, Doherty M, Pascual E, Barskova
V, Becce F, Castañeda-Sanabria J, Coyfish M, Guillo S, Jansen TL,
Janssens H, et al: 2016 updated EULAR evidence-based
recommendations for the management of gout. Ann Rheum Dis.
76:29–42. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen GL, Wei W and Xu SY: Effect and
mechanism of total saponin of dioscorea on animal experimental
hyperuricemia. Am J Chin Med. 34:77–85. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Martin WJ, Walton M and Harper J: Resident
macrophages initiating and driving inflammation in a monosodium
urate monohydrate crystal-induced murine peritoneal model of acute
gout. Arthritis Rheum. 60:281–289. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pope RM and Tschopp J: The role of
interleukin-1 and the inflammasome in gout: Implications for
therapy. Arthritis Rheum. 56:3183–3188. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Scott P, Ma H, Viriyakosol S, Terkeltaub R
and Liu-Bryan R: Engagement of CD14 mediates the inflammatory
potential of monosodium urate crystals. J Immunol. 177:6370–6378.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Martin WJ, Shaw O, Liu X, Steiger S and
Harper JL: Monosodium urate monohydrate crystal-recruited
noninflammatory monocytes differentiate into M1-like
proinflammatory macrophages in a peritoneal murine model of gout.
Arthritis Rheum. 63:1322–1332. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Schroder K, Zhou R and Tschopp J: The
NLRP3 inflammasome: A sensor for metabolic danger? Science.
327:296–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Choe JY, Jung HY, Park KY and Kim SK:
Enhanced p62 expression through impaired proteasomal degradation is
involved in caspase-1 activation in monosodium urate
crystal-induced interleukin-1β expression. Rheumatology (Oxford).
53:1043–1053. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Terkeltaub R, Zachariae C, Santoro D,
Martin J, Peveri P and Matsushima K: Monocyte-derived neutrophil
chemotactic factor/interleukin-8 is a potential mediator of
crystal-induced inflammation. Arthritis Rheum. 34:894–903. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu Q, Zhang D, Hu D, Zhou X and Zhou Y:
The role of mitochondria in NLRP3 inflammasome activation. Mol
Immunol. 103:115–124. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Punzi L, Scanu A, Ramonda R and Oliviero
F: Gout as autoinflammatory disease: New mechanisms for more
appropriated treatment targets. Autoimmun Rev. 12:66–71. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zheng SC, Zhu XX, Xue Y, Zhang LH, Zou HJ,
Qiu JH and Liu Q: Role of the NLRP3 inflammasome in the transient
release of IL-1 β induced by monosodium urate crystals in human
fibroblast-like synoviocytes. J Inflamm (Lond). 12:302015.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lu W, Sha N and Guang-liang C: Effect of
total saponin of Dioscorea on NALP3 inflammasome signaling pathway
with acute gouty in rats. Chinese Pharmacological Bulletin.
33:354–360. 2017.
|
|
44
|
Blomgran R, Patcha Brodin V, Verma D,
Bergström I, Söderkvist P, Sjöwall C, Eriksson P, Lerm M, Stendahl
O and Särndahl E: Common genetic variations in the NALP3
inflammasome are associated with delayed apoptosis of human
neutrophils. PLoS One. 7:e313262012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kool M, Petrilli V, De Smedt T, Rolaz A,
Hammad H, van Nimwegen M, Bergen IM, Castillo R, Lambrecht BN and
Tschopp J: Cutting edge: Alum adjuvant stimulates inflammatory
dendritic cells through activation of the NALP3 inflammasome. J
Immunol. 181:3755–3759. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hornung V, Bauernfeind F, Halle A, Samstad
EO, Kono H, Rock KL, Fitzgerald KA and Latz E: Silica crystals and
aluminum salts activate the NALP3 inflammasome through phagosomal
destabilization. Nat Immunol. 9:847–856. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Dostert C, Petrilli V, Van Bruggen R,
Steele C, Mossman BT and Tschopp J: Innate immune activation
through Nalp3 inflammasome sensing of asbestos and silica. Science.
320:674–677. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Cassel SL, Eisenbarth SC, Iyer SS, Sadler
JJ, Colegio OR, Tephly LA, Carter AB, Rothman PB, Flavell RA and
Sutterwala FS: The Nalp3 inflammasome is essential for the
development of silicosis. Proc Natl Acad Sci USA. 105:9035–9040.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Halle A, Hornung V, Petzold GC, Stewart
CR, Monks BG, Reinheckel T, Fitzgerald KA, Latz E, Moore KJ and
Golenbock DT: The NALP3 inflammasome is involved in the innate
immune response to amyloid-beta. Nat Immunol. 9:857–865. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ozaki E, Campbell M and Doyle SL:
Targeting the NLRP3 inflammasome in chronic inflammatory diseases:
Current perspectives. J Inflamm Res. 8:15–27. 2015.PubMed/NCBI
|
|
51
|
Menu P and Vince JE: The NLRP3
inflammasome in health and disease: The good, the bad and the ugly.
Clin Exp Immunol. 166:1–15. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mason DR, Beck PL and Muruve DA:
Nucleotide-binding oligomerization domain-like receptors and
inflammasomes in the pathogenesis of non-microbial inflammation and
diseases. J Innate Immun. 4:16–30. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ogura Y, Sutterwala FS and Flavell RA: The
inflammasome: First line of the immune response to cell stress.
Cell. 126:659–662. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhou Q, Yu DH, Zhang N and Liu SM:
Anti-inflammatory effect of total saponin fraction from
Dioscorea nipponica Makino on gouty arthritis and its
influence on NALP3 inflammasome. Chin J Integr Med. 25:663–670.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yang D, Elner SG, Bian ZM, Till GO, Petty
HR and Elner VM: Pro-inflammatory cytokines increase reactive
oxygen species through mitochondria and NADPH oxidase in cultured
RPE cells. Exp Eye Res. 85:462–472. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bordt EA and Polster BM: NADPH oxidase-
and mitochondria-derived reactive oxygen species in proinflammatory
microglial activation: A bipartisan affair? Free Radic Biol Med.
76:34–46. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Won JH, Park S, Hong S, Son S and Yu JW:
Rotenone-induced impairment of mitochondrial electron transport
chain confers a selective priming signal for NLRP3 inflammasome
activation. J Biol Chem. 290:27425–27437. 2015. View Article : Google Scholar : PubMed/NCBI
|