Introduction
Colorectal cancer (CRC) is the third most common
cause of cancer-related deaths (1), and the incidence rates are rapidly
increasing, with >1 million new cases and 694,000 mortalities
each year worldwide (2). The
current first-line treatment for CRC is a combination of
radiotherapy and chemotherapy (3),
where drugs such as cisplatin are frequently employed (4). However, such treatments have serious
side effects and are often associated with drug resistance,
resulting in the constant need to identify alternative treatment
options with fewer side effects. This has led to increased interest
in the use of natural products to treat CRC (5).
Epithelial-mesenchymal transition (EMT) is a key
process in cancer metastasis, which is characterized by the
decreased expression of cell-cell adhesion molecules such as
E-cadherin, and the increased expression of mesenchymal proteins
such as vimentin. During EMT, epithelial cells adopt a mesenchymal
phenotype and exhibit increased migratory potential; this allows
for increased invasiveness and resistance to apoptosis (6,7).
Furthermore, EMT promotes cancer cell resistance to chemo- and
radiotherapy (8). Snail is a
member of the zinc-finger transcription factor family, and is
reportedly one of the most important transcriptional regulators of
EMT (9). Zheng et al
(10), revealed that in CRC, Snail
expression was significantly enhanced, which affected cancer
progression. Additionally, Kwon et al (11) suggested that Snail may be a novel
prognostic biomarker and therapeutic target in CRC. In fact; to an
extent, the activation of Snail is considered to be the initiating
factor for EMT in various malignant tumors.
Flavonoids are a diverse family of polyphenolic
compounds derived from plant-based foods, including fruit, seeds,
vegetables, herbs, tea and wine. Flavonoids have been extensively
studied and their anticancer effects are well documented (12). Epidemiological studies have
identified that the increased intake of dietary flavonoids is
associated with a decreased risk of developing CRC (13). Previous studies also revealed that
flavonoids suppressed the migration and invasion abilities
(14), influenced cell cycle
progression (15) and induced
apoptosis in CRC cells (16–18).
In Asia, S. baicalensis, whose constituents include numerous
flavonoid compounds, is widely used for the treatment of
hypertension (19), inflammation
(20) and cancer (21), as well as bacterial and viral
infectious diseases (22).
Baicalein, which is currently one of the most representative
flavonoid aglycones in S. baicalensis, has received
considerable attention for its reported ability to suppress
cellular proliferation and induce apoptosis (17,23–26).
However, the specific mechanisms of these anti-metastatic
properties remain unclear.
In the present study, baicalein was extracted and
analyzed using ultra-high performance liquid chromatography-tandem
mass spectrometry (UPLC-MS/MS), and its effect on cell
proliferation, migration and invasion, and the expression of EMT
markers was subsequently evaluated. Notably, Snail-induced EMT was
partially blocked by baicalein treatment, which provides
theoretical evidence for its use as a potential antitumor agent,
and indicates a novel mechanism for its antitumor effects.
Materials and methods
Chemicals
High performance liquid chromatography (HPLC)-grade
methanol and formic acid were obtained from Merck KGaA. Deionized
water was prepared via Milli-Q water purification (EMD Millipore).
Baicalein reference standards (cat. no. 18031608) were purchased
from the Beijing Aoke Biological Technology Corporation, and stock
solutions were prepared in dimethyl sulfoxide (DMSO) and stored at
4°C. Methyl p-hydroxybenzoate (internal standard; purity >98% by
HPLC-UV) was obtained from Sigma-Aldrich (Merck KGaA).
UPLC/MS/MS instrument and
conditions
In order to detect and analyze baicalein in rat
plasma after oral administration of S. baicalensis extract,
UPLC-MS/MS was conducted as previously described (27). An Acquity UPLC system (Waters
Corporation) and an API 4000 Triple Quadrupole mass spectrometer
(Shanghai AB SCIEX Analytical Instrument Trading Co.) equipped with
an electrospray ionization (ESI) source were used as a part of this
system, using Analyst 1.5 software (Applied Biosystems; Thermo
Fisher Scientific, Inc.) for assay control. An Acquity UPLC HSS BEH
C18 column (100 × 2.1 mm, 1.7 µm; Waters Corporation) maintained at
40°C was used for chromatographic separation, with a mobile phase
of (A) 0.1% formic acid, and (B) methanol, with a gradient elution
of 60–90% (A) for 0–6 min, 90% (A) for 6–7 min, 90–35% (A) for 7–10
min, and 35% (A) for 10–12 min. The flow rate of the mobile phase
was 0.20 ml/min, with a 1-µl injection volume. The electrospray
ionization source was performed in negative ionization mode with
the following parameters: −4.5 kV ion spray voltage, 500°C turbo
spray temperature. For gas pressures, nebulizer, heater and curtain
gas were set to 55, 50 and 25 psi, respectively, with a dwell time
of 50 msec. Detection analysis was conducted in multiple reaction
monitoring mode at the transition m/z [M-H]− 269.2à195.0
for baicalein and 150.9à136.0 for IS, with collision energies of
−35 eV and −19 eV, and cone voltages of −70 V and −56 V,
respectively.
Preparation of S. baicalensis
extract
To obtain an aqueous extraction of S.
baicalensis, 100 g S. baicalensis roots were immersed in
distilled water for 30 min with occasional stirring, and then
boiled three times for 30 min each; plant:water ratios were
maintained at 1:10. The three separate decoctions were combined and
concentrated into a final volume of 100 ml to yield S.
baicalensis extract, which was used for UPLC-MS/MS analysis.
The raw materials were identified by Professor Minghua Qiu of
Kunming Institute of Botany, Chinese Academy of Sciences (Kunming,
China), where voucher specimens are retained.
Animal study
A total of six male Sprague-Dawley rats (weight,
250±20 g; age, 8 weeks) were provided by Kunming Medical University
(Yunnan, China). The rats were housed under standard conditions
(20±2°C with 60±5% humidity and 12-h light/dark cycles) with free
access to food and water, and acclimated for 1 week. Animals were
observed daily throughout the study. All rats were fasted, with
free access to water, for 12 h prior to the experiment. The rats
were then randomized into 2 groups (n=3 per group); the
experimental group received S. baicalensis extract (4.5
g/kg) by intragastric administration once (28), and the control rats received
distilled water only (10 ml/kg of body weight). At 45 min after
oral administration, rats were then anesthetized using
pentobarbital sodium at a dose of 50 mg/kg (i.p.), and blood
samples (0.5 ml) were collected via retro-orbital bleeding. The
anesthetic agent doses selected were based on existing literature
(29). The sampling time-points
were selected based on previous pharmacokinetic studies (27). Subsequently, the rats were
sacrificed by cervical dislocation. Plasma was then separated from
blood after centrifugation at 5,000 × g for 10 min at 4°C. The
humane endpoint of this experiment was as follows: A marked
reduction in food or water intake, labored breathing, inability to
stand, and no response to external stimuli. No abnormal signs that
signified the humane endpoints of the experiment were observed from
any of the rats during the experiment. All animal procedures were
approved and performed in compliance with the guidelines set by the
Animal Care Committee of the First People's Hospital of Yunnan
Province (30).
Sample preparation
A total of 10 µl IS (methyl p-hydroxybenzoate, 1
µg/ml in methanol) and 50 µl 0.2 M HCl were spiked into a 100-µl
sample of rat plasma, mixed and allowed to rest for 10 min. Next,
800 µl ethyl acetate was added and mixed for 3 min, followed by
centrifugation for 5 min at 5,000 × g (4°C). The supernatants were
transferred to fresh tubes and evaporated using a nitrogen gas
stream at room temperature. Any remaining residue was dissolved in
100 µl mobile phase, mixed for 1 min by vortexing and centrifuged
at 15,000 × g for 5 min at 4°C . Finally, 1 µl supernatant was
injected into the UPLC-MS/MS system for baicalein detection.
Cell culture
HT29 and DLD1 human colorectal cancer cell lines
were obtained from Shanghai Cell Biological Institute of the
Chinese Academy of Sciences, and cultured in RPMI-1640 media
(Hyclone; GE Healthcare Life Sciences) containing 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin (Thermo Fisher Scientific, Inc.) at 37°C
with 5% CO2.
Cell Counting Kit-8 (CCK-8) assay
The CCK-8 assay (Beyotime Institute of
Biotechnology) was used to assess the effects of baicalein on
cancer cell viability. HT29 and DLD1 cells were seeded into a
96-well plate at a density of 2×103 cells/well, and
cultured until complete adherence. The cells were treated with
baicalein at concentrations of 0, 20, 40, 60, 80, 100 and 120
µmol/l for 24, 48 and 72 h, using DMSO as a negative control. The
media was then replaced with fresh media containing 10% CCK-8
solution. After a further 3 h of incubation, the optical density
(OD) of each well was assessed using a spectrophotometer at a
wavelength of 450 nm. The inhibition rates were calculated as
follows: Inhibition rate = [1- (OD drug treated - OD blank)/(OD
control-OD blank)] ×100%. The half inhibitory concentration
(IC50) for baicalein was determined using the Logit
method (31), indicating the
concentration of baicalein necessary to inhibit 50% cell
proliferation at a given time-point.
Wound-healing assay
HT29 cells were plated in a 6-well plate at
1×106 cells/well, and cultured to 90% confluency. A
10-µl sterile micropipette tip was used to create a wound across
the monolayer, and the cells were washed twice with sterile
phosphate-buffered saline (PBS) to remove debris. The cells were
treated with 10, 20, and 30 µmol/l baicalein in RPMI-1640 media
without FBS, and the control group was treated with 0.05% DMSO
only. Cell migration was assessed using an inverted microscope
(Zeiss Axio Vert.A1), original magnification, ×10.
Transwell invasion assay
To assess the effects of baicalein on cancer cell
invasiveness, a Transwell invasion assay was performed using
24-well Transwell chambers (pore size, 8 µm), pre-coated with
Matrigel® for 1 h at 37°C. (BD Biosciences). Following
treatment with 10, 20, and 30 µmol/l baicalein at 37°C for 24 h,
HT29 cells were digested with 0.25% trypsin and resuspended in
serum-free RPMI-1640 medium at a density of 1×106/ml,
and 100 µl cell suspension was added into the upper chambers. The
lower chambers were filled with 500 µl medium supplemented with 10%
FBS. Following incubation at 37°C for 24 h, the inserts were
detached and non-invasive cells were gently removed with a cotton
wool swab. Invaded cells were fixed with 4% paraformaldehyde for 20
min at room temperature and stained with 0.1% crystal violet for 15
min at room temperature. Stained cells were visualized using an
inverted microscope (Zeiss Axio Scope.A1) and counted in 5 randomly
selected fields (magnification, ×10).
Plasmid transfection
pcDNA3.1-vector and pcDNA3.1-Snail plasmids were
obtained from Shanghai GeneChem Co., Ltd., and verified by DNA
sequencing. The pcDNA3.1-vector plasmids were used as the controls.
HT29 and DLD1 cells (4×105 cells/well) were seeded into
6-well plates with complete medium and incubated at 37°C for 24 h
prior to transfection. For transient transfections, cells were
transfected with 2.5 µg plasmid using Lipofectamine®
3000 transfection reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. The cells were
harvested 1–2 days after transfection for further
investigation.
Western blot analysis
For western blotting, HT29 and DLD1 cells were
plated at 3×105 cells/well in 6-well plates, and treated
with 10, 20, and 30 µmol/l baicalein or DMSO only for 24 or 48 h at
37°C. Cells were washed with PBS and lysed using ice-cold RIPA
buffer (Beyotime Institute of Biotechnology) for 30 min to extract
the total protein, which was quantified using the bicinchoninic
acid assay method. The lysates were then denatured in loading
buffer containing 4% SDS, and incubated at 95°C for 10 min. In
total, 50 µg total protein per sample was separated by 10% SDS-PAGE
gel and transferred to PVDF membranes, which were subsequently
blocked for 2 h using 5% non-fat dry milk in TBS and 0.05% Tween
20. The membranes were incubated overnight at 4°C with primary
antibodies targeted against: E-cadherin (cat. no. 14472; 1:1,000;
Cell Signaling Technology, Inc.), vimentin (cat. no. 5741; 1:1,000;
Cell Signaling Technology, Inc.), Snail (cat. no. 3879; 1:1,000;
Cell Signaling Technology, Inc.), Twist1 (cat. no. 46702; 1:1,000;
Cell Signaling Technology, Inc.), p53 (cat. no. 60283-2-Ig; 1:800;
ProteinTech Group, Inc.), p21 (cat. no. 10355-1-AP; 1:800;
ProteinTech Group, Inc.) and β-actin (cat. no. 4970; 1:10,000; Cell
Signaling Technology, Inc.). Following primary incubation, the
membranes were incubated with horseradish peroxidase-conjugated
goat anti-mouse (cat. no. 6946; 1:8,000; Abcam) and anti-rabbit
(cat. no. 6721; 1:6,000; Abcam) IgG (H+L) secondary antibodies for
2 h at room temperature. Protein bands were visualized in a
darkroom using enhanced chemiluminescence reagents (New Cell &
Molecular Biotech Co., Ltd.). Protein expression was quantified
using ImageJ software (v.1.48; National Institutes of Health) with
β-actin as the loading control.
Statistical analysis
All experiments were performed in triplicate. Data
are presented as the mean ± standard deviation. Statistical
analyses were performed using SPSS software (v.22.0; IBM Corp.).
One-way ANOVA followed by Dunnett's post hoc test was used to
compare the treatment and control groups. P<0.05 was considered
to indicate a statistically significant difference. All graphs were
generated using GraphPad Prism software (v.5.0; GraphPad Software,
Inc.).
Results
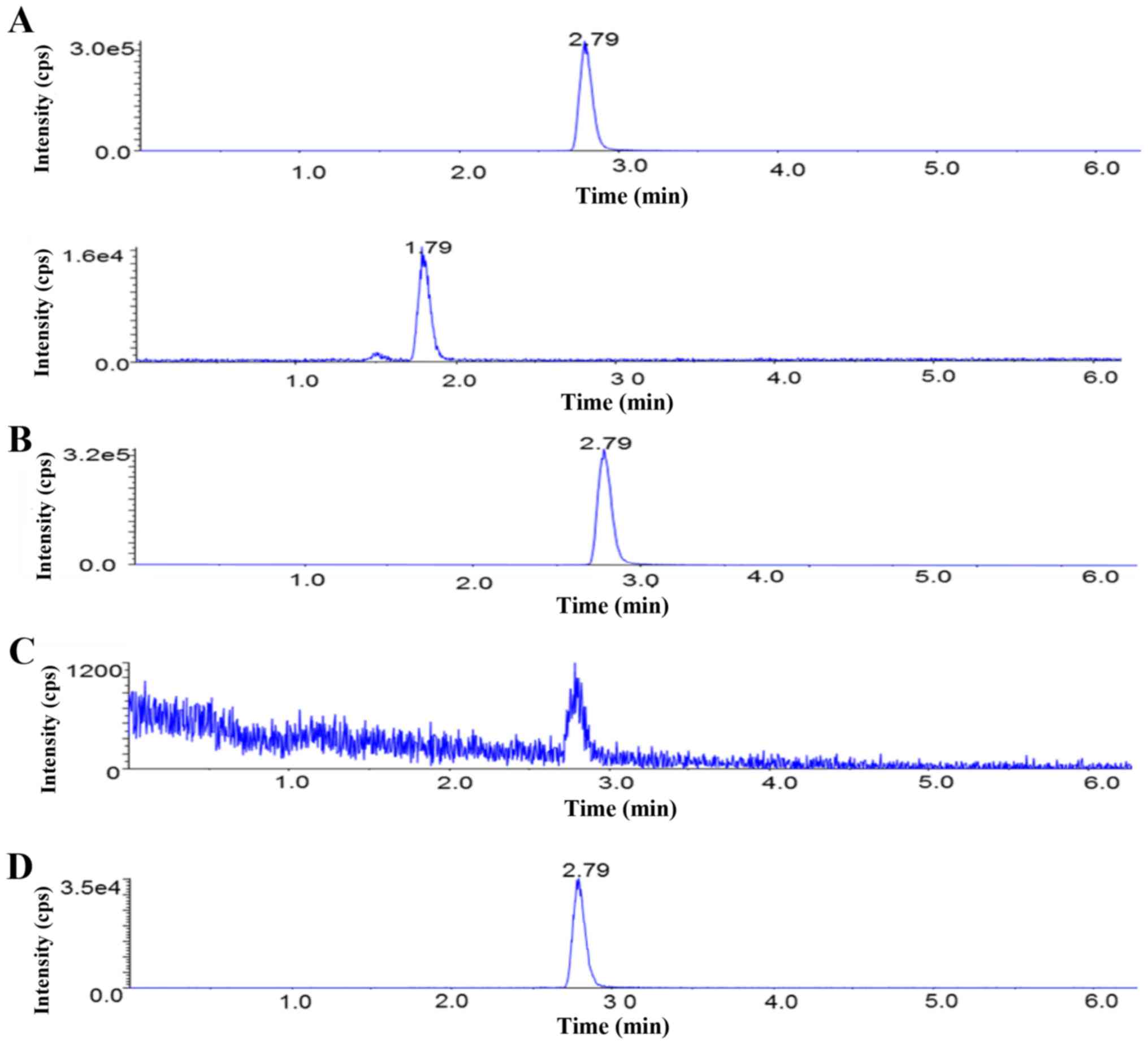
Analysis of the absorption of
baicalein from S. baicalensis extract
The S. baicalensis herb comprises a complex
mixture of different phytochemicals, and contains >60 chemical
components. Thus, the purpose of using the UPLC-MS/MS technique was
to detect and identify baicalein in rat plasma following the oral
administration of S. baicalensis extract. A representative
chromatogram of baicalein is presented in Fig. 1.
Baicalein suppresses DLD1 and HT29
cell proliferation in vitro
The anti-proliferative activity of baicalein was
assessed with a CCK-8 assay, using HT29 and DLD1 cells treated with
0–120 µmol/l baicalein to identify the minimal non-lethal dose.
Baicalein was revealed to inhibit the viability of HT29 and DLD1
cells in a dose- and time-dependent manner; in HT29 cells, the
IC50 values at 24, 48 and 72 h were 49.77, 34.35 and
16.91 µmol/l, respectively; and in DLD1 cells, the IC50
values were 60.49, 34.70, and 18.75 µmol/l at the same time-points,
respectively (Fig. 2A). To avoid
growth suppression, all subsequent experiments were conducted using
baicalein concentrations <34 µmol/l.
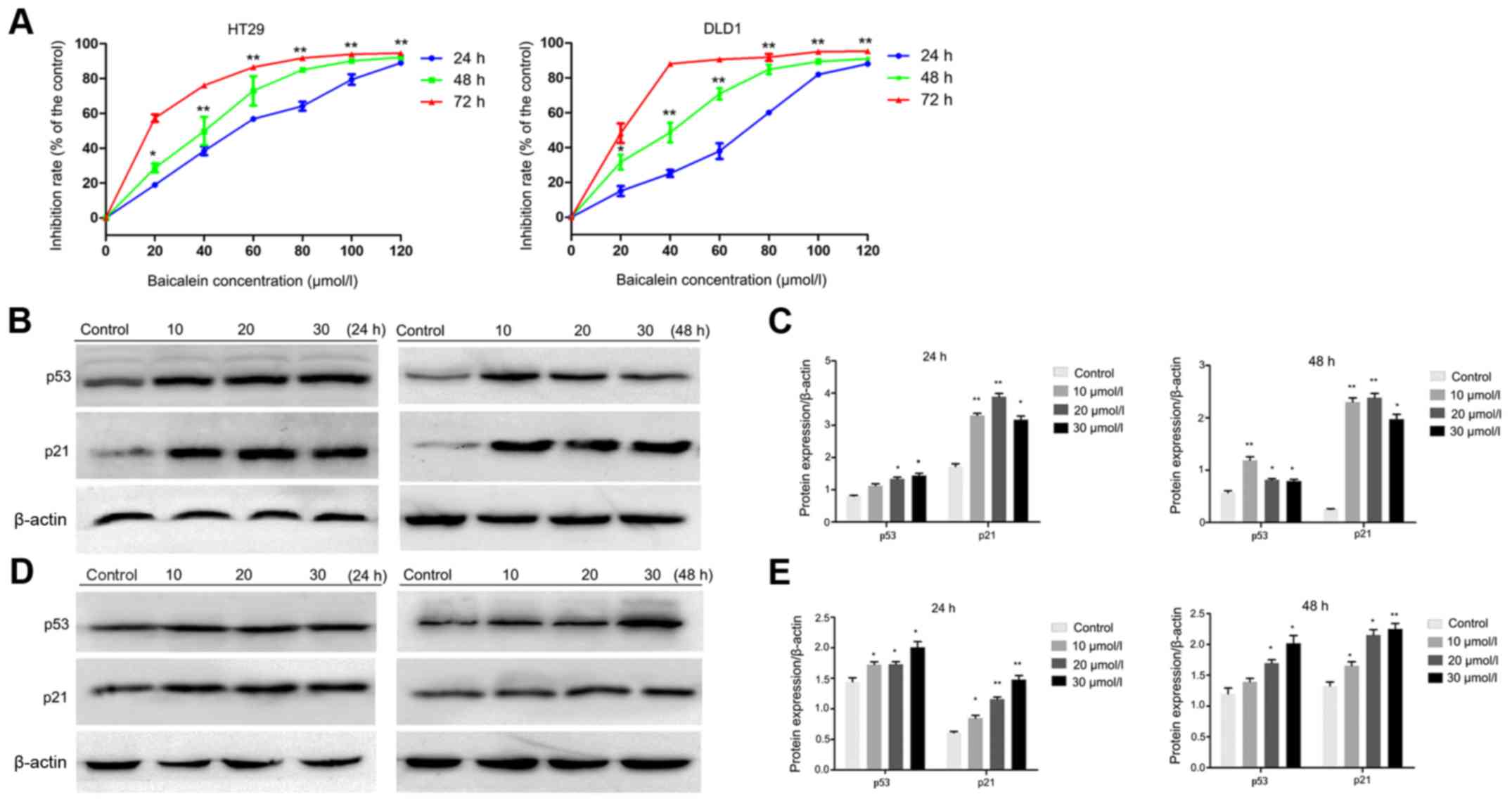 | Figure 2.Baicalein suppresses cellular
proliferation in CRC. (A) HT29 and DLD1 cells were treated with 0,
20, 40, 60, 80, 100 and 120 µmol/l baicalein for 24, 48 and 72 h,
and the CCK-8 assay was used to assess cellular proliferation. (B)
Effects of baicalein on p53 and p21 expression in HT29 cells. Cells
were treated with or without baicalein at the indicated
concentrations for 24 or 48 h, and the protein expression levels of
p53 and p21 were determined by western blotting. β-actin was used
as the internal control. (C) p53 and p21 expression in HT29 cells
was quantified using ImageJ software. (D) Effects of baicalein on
p53 and p21 expression in DLD1 cells. Cells were treated with or
without baicalein at the indicated concentrations for 24 or 48 h,
and the protein expression levels of p53 and p21 were determined by
western blotting. (E) p53 and p21 expression levels in DLD1 cells
were calculated using ImageJ software. Results are expressed as the
means ± standard deviation of three separate experiments.
*P<0.05 and **P<0.01 vs. the control. CRC, colorectal cancer;
CCK-8, Cell Counting Kit-8. |
p53 and p21 are reportedly involved in the
baicalein-associated inhibition of CRC HCT116 cell proliferation
(24). Therefore, the expression
levels of p53 and p21 in HT29 and DLD1 cells were investigated,
with or without baicalein treatment. As revealed in Fig. 2B and D, both p53 and p21 expression
were significantly increased in baicalein-treated cells compared
with the control cells, which was consistent with the
aforementioned study (24).
Baicalein affects the mobility of HT29
cells in vitro
p53 is also important for the regulation of
metastasis and E-cadherin expression (32). Chang et al (33), revealed that p53 inhibits the
invasiveness of CRC cells by regulating EMT. To further explore the
effects of baicalein on CRC cells, the migration and invasion
abilities of baicalein-treated HT29 cells were investigated. The
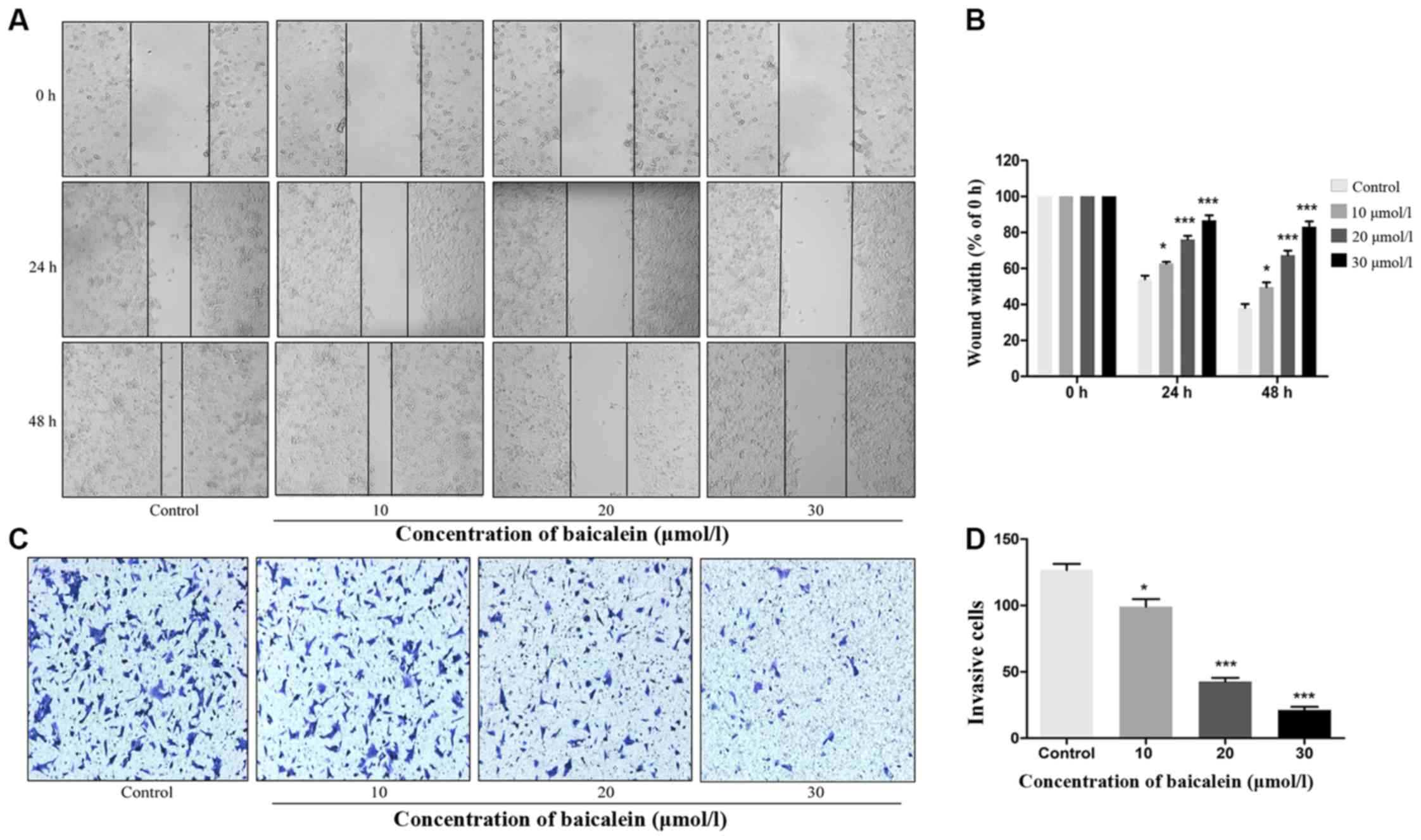
wound-healing assay results demonstrated that baicalein inhibited
the migratory ability of CRC cells in a dose-dependent manner
(Fig. 3A). Following treatment
with 10, 20 or 30 µmol/l baicalein for 48 h, HT29 cell motility was
inhibited by 49.65, 67.41 and 83.17%, respectively (Fig. 3B). In the Transwell assays, cells
from the control group exhibited a higher invasive capacity than
those that had been treated with baicalein, indicating that
baicalein significantly inhibited the invasiveness of CRC cells in
a dose-dependent manner (Fig. 3C and
D). These findings indicated that baicalein may act as a
suppressor of CRC cell migration and invasion.
Baicalein regulates the expression of
EMT markers in HT29 and DLD1 cells
EMT is an important process which is characterized
by the decreased expression of epithelial markers such as
E-cadherin, and the concomitant increased expression of mesenchymal
markers such as vimentin, matrix metalloproteinase 9 and various
transcription factors (34). In
the present study, the effects of baicalein on the expression of
EMT markers was assessed in HT29 and DLD1 cells, following a 24- or
48-h treatment with 0–30 µmol/l baicalein. Compared with the
control group, Snail, Twist1 and vimentin expression was decreased
in baicalein-treated HT29 cells, while E-cadherin expression was
increased at 24 and 48 h, respectively (Fig. 4A and B). Similar effects were
observed in DLD1 cells (Fig. 4C and
D). These results indicated that baicalein was able to impede
EMT in CRC cells.
 | Figure 4.Baicalein regulates the expression of
epithelial-mesenchymal transition-associated markers. (A) HT29
cells were treated with or without baicalein at the indicated
concentrations for 24 or 48 h, and protein expression levels of
E-cadherin, vimentin, Snail and Twist1 were determined by western
blotting. (B) E-cadherin, vimentin, Snail and Twist1 expression
levels in HT29 cells were quantified using ImageJ software. (C)
DLD1 cells were treated with or without baicalein at the indicated
concentrations for 24 or 48 h, and the protein levels of
E-cadherin, vimentin, Snail and Twist1 were determined by western
blotting. (D) E-cadherin, vimentin, Snail and Twist1 expression in
DLD1 cells were calculated using ImageJ software. Results are
presented as the means ± standard deviation of three separate
experiments. *P<0.05, **P<0.01 and ***P<0.001 vs. the
control. β-actin was used as the internal control. |
Baicalein inhibits Snail-induced EMT
in CRC cells
Since Snail plays a key role in metastasis and
baicalein suppresses the expression of Snail and Snail-associated
target genes (35), it was
hypothesized that the antitumor activity of baicalein was
influenced by Snail. In order to confirm this hypothesis, CRC cells
were transfected with a pcDNA3.1-Snail plasmid, and Snail
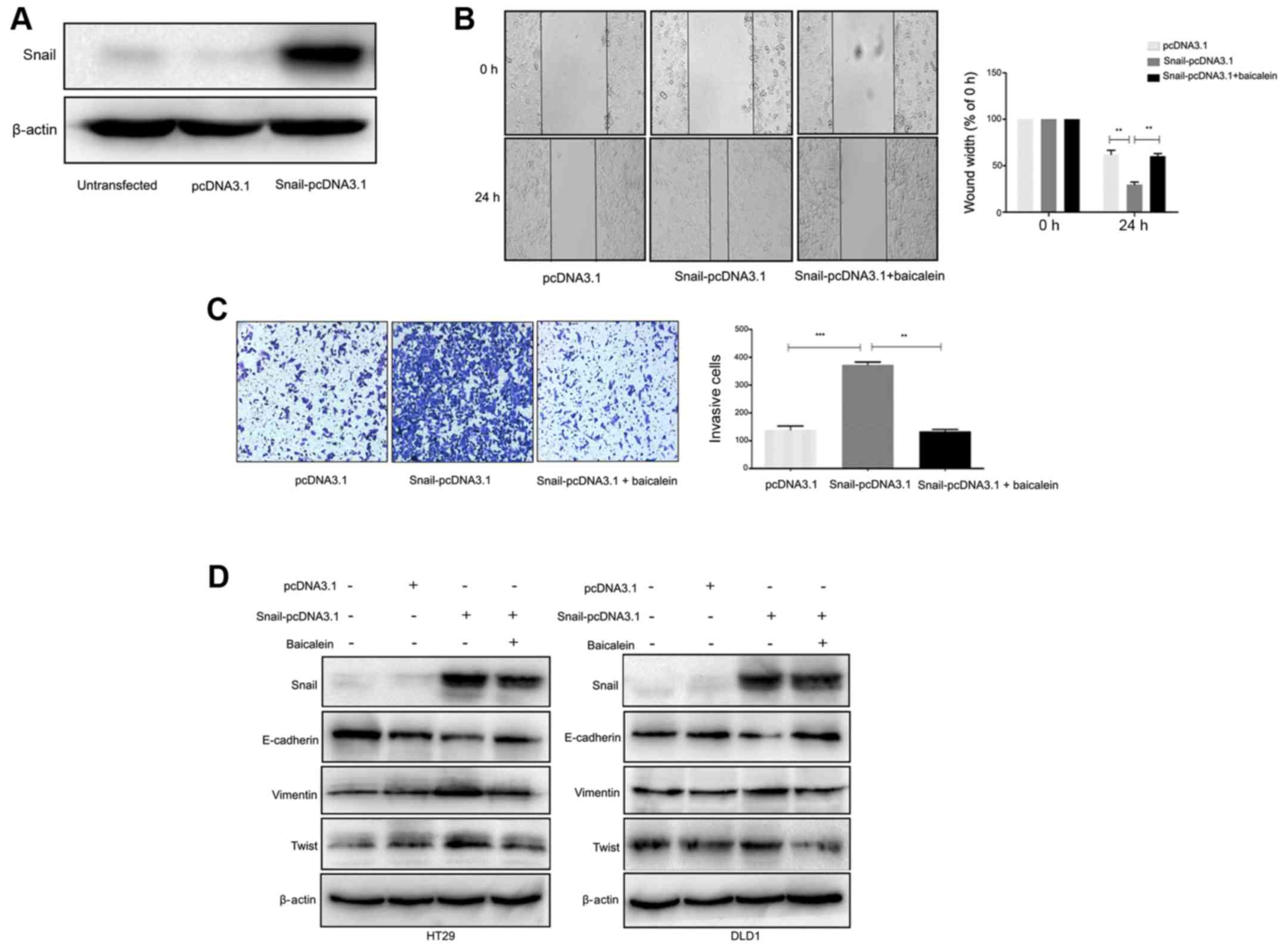
overexpression was confirmed by western blotting (Fig. 5A). The overexpression of Snail was
revealed to increase the migratory and invasive capacities of CRC
cells, which were reversed by baicalein treatment (Fig. 5B and C). Snail is reported to be
one of the most important EMT-associated transcription factors;
therefore, the effects of baicalein on Snail-induced EMT were also
evaluated. As revealed in Fig. 5D,
transfection with Snail-pcDNA3.1 markedly increased the expression
levels of Snail, vimentin and Twist1, while the level of E-cadherin
expression was significantly decreased, compared with the
control-transfected cells; these results were consistent with the
EMT expression profile. Notably, following baicalein treatment,
Snail-induced vimentin and Twist1 upregulation, as well as
E-cadherin downregulation were decreased both in HT29 and DLD1
cells, indicating that baicalein exhibits its suppressive effect
partly through the inhibition of Snail-induced EMT.
Discussion
Plants possess a complex mixture of different
phytochemicals; S. baicalensis contains >60 chemical
components, of which baicalein is the primary contributor to its
antitumor effects. In the present study, a UPLC-MS/MS technique was
employed to detect and identify baicalein in rat plasma after the
oral administration of S. baicalensis extract. The results
demonstrate that baicalein is a major bioactive ingredient of S.
baicalensis, which is absorbed into the blood via enterocytes.
However, it should be noted that S. baicalensis is most
often administered orally, and that flavonoids are usually unstable
at a neutral pH (36). On the
other hand, the flavonoid metabolite 2, 4, 6-trihydroxybenzoic acid
was reported to mediate its effects through a CDK- and
sodium-coupled monocarboxylate transporter 1-dependent pathway
contributing to the prevention of CRC (37). Therefore, it is possible that
flavonoids may be subjected to degradation by the intestinal
microflora or its metabolites (38).
S. baicalensis is commonly used to treat
cancer (39,40); baicalein is the primary active
ingredient present within extracts of S. baicalensis
(41,42). Baicalein treatment can inhibit
cellular proliferation by blocking the cell cycle and inducing
apoptosis and senescence, via the modulation of the
mitogen-activated protein kinase 1 ERK-1/-2, and p53-p21 pathways
(17,24). In the present study, baicalein
exhibited its antitumor effects even at a low concentration (10
µM), at which p53 and p21 expression were significantly increased.
Similar results were observed for E-cadherin, vimentin, Snail and
Twist1 expression levels. It is well known that p53 induces
apoptosis in response to a variety of cellular stimuli (43). Since the induction of apoptosis is
a major mechanism of most chemotherapeutic agents, it was
hypothesized that p53 may be involved in baicalein-mediated
cellular proliferation. Notably, both p53 and p21 expression were
increased in baicalein-treated cells in the present study.
Emerging evidence has revealed that EMT is
responsible for the development of metastatic dissemination, a
characteristic of the advanced clinical stages of CRC (44). Baicalein has been reported to
inhibit EMT by regulating the Wnt/β-catenin signaling pathway
(45). However, the effects of
baicalein on Snail, a primary promoter of EMT, have not been
previously reported, to the best of the authors' knowledge. In the
present study, Snail-overexpression was revealed to significantly
promote CRC cell invasiveness, which was partially reversed by
baicalein treatment, resulting in the downregulation of E-cadherin
and the upregulation of Snail and Twist1. Hence, to the best of the
authors' knowledge, the present study is the first to report that
baicalein suppresses EMT, partly through a decrease in Snail
activity.
To conclude, the present study confirmed that
baicalein is absorbed into the blood and can inhibit cellular
proliferation, migration and invasiveness in CRC, potentially by
regulating p53 and p21 expression, and disrupting EMT. These data
suggest that baicalein, a primary component of S.
baicalensis, exerts potent anticancer effects against human CRC
cells, and is potentially an effective target drug for cancer
therapy.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81302159
and 81502128) and the Joint Foundation of Kunming Medical
University and Yunnan Provincial Science and Technology Department
(grant no. 2017FE467-159), and a grant from the Internal Division
of Yunnan Provincial Health Commission (grant no. 2016NS226).
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
QG, WZ and QZ conceived and designed the
experiments. WZ and QZ performed the experiments. YZ analyzed the
data. QZ wrote the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
All animal procedures were approved and performed in
compliance with the guidelines set by the Animal Care Committee of
the First People's Hospital of Yunnan Province.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
UPLC-MS/MS
|
ultra-high performance liquid
chromatography-tandem mass spectrometric
|
|
S. baicalensis
|
Scutellaria baicalensis
|
|
CCK-8
|
Cell Counting Kit-8
|
|
EMT
|
epithelial-mesenchymal transition
|
|
CRC
|
colorectal cancer
|
|
DMSO
|
dimethyl sulfoxide
|
|
OD
|
optical density
|
|
PBS
|
phosphate-buffered saline
|
|
FBS
|
fetal bovine serum
|
References
|
1
|
Friedman S, Rubin PH, Bodian C, Goldstein
E, Harpaz N and Present DH: Screening and surveillance colonoscopy
in chronic Crohn's colitis. Gastroenterology. 120:820–826. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pan J, Xu Y, Song H, Zhou X, Yao Z and Ji
G: Extracts of Zuo Jin Wan, a traditional Chinese medicine,
phenocopies 5-HTR1D antagonist in attenuating Wnt/β-catenin
signaling in colorectal cancer cells. BMC Complement Altern Med.
17:506–517. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li K, Guo J, Wu Y, Jin D, Jiang H, Liu C
and Qin C: Suppression of YAP by DDP disrupts colon tumor
progression. Oncol Rep. 39:2114–2126. 2018.PubMed/NCBI
|
|
5
|
Ling CQ, Yue XQ and Ling C: Three
advantages of using traditional Chinese medicine to prevent and
treat tumor. J Integr Med. 12:331–335. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yeung KT and Yang J:
Epithelial-mesenchymal transition in tumor metastasis. Mol Oncol.
11:28–39. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tsai JH and Yang J: Epithelial-mesenchymal
plasticity in carcinoma metastasis. Genes Dev. 27:2192–2206. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Singh A and Settleman J: EMT, cancer stem
cells and drug resistance: An emerging axis of evil in the war on
cancer. Oncogene. 29:4741–4751. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cai W, Ye Q and She QB: Loss of 4E-BP1
function induces EMT and promotes cancer cell migration and
invasion via cap-dependent translational activation of snail.
Oncotarget. 5:6015–6027. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zheng H, Shen M, Zha YL, Li W, Wei Y,
Blanco MA, Ren G, Zhou T, Storz P, Wang HY, et al: PKD1
phosphorylation-dependent degradation of SNAIL by SCF-FBXO11
regulates epithelial-mesenchymal transition and metastasis. Cancer
Cell. 26:358–373. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kwon CH, Park HJ, Choi JH, Lee JR, Kim HK,
Jo HJ, Kim HS, Oh N, Song GA and Park DY: Snail and serpinA1
promote tumor progression and predict prognosis in colorectal
cancer. Oncotarget. 6:20312–20326. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bugel SM and Tanguay RL: Multidimensional
chemobehavior analysis of flavonoids and neuroactive compounds in
zebrafish. Toxicol Appl Pharmacol. 344:23–34. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bobe G, Sansbury LB, Albert PS, Cross AJ,
Kahle L, Ashby J, Slattery ML, Caan B, Paskett E, Iber F, et al:
Dietary flavonoids and colorectal adenoma recurrence in the Polyp
Prevention Trial. Cancer Epidemiol Biomarkers Prev. 17:1344–1353.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu K, Gao H, Wang Q, Wang L, Zhang B, Han
Z, Chen X, Han M and Gao M: Hispidulin suppresses cell growth and
metastasis by targeting PIM1 through JAK2/STAT3 signaling in
colorectal cancer. Cancer Sci. 109:1369–1381. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Razak S, Afsar T, Ullah A, Almajwal A,
Alkholief M, Alshamsan A and Jahan S: Taxifolin, a natural
flavonoid interacts with cell cycle regulators causes cell cycle
arrest and causes tumor regression by activating Wnt/β -catenin
signaling pathway. BMC Cancer. 18:1043–1061. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wenzel U, Kuntz S, Brendel MD and Daniel
H: Dietary flavone is a potent apoptosis inducer in human colon
carcinoma cells. Cancer Res. 60:3823–3831. 2000.PubMed/NCBI
|
|
17
|
Dou J, Wang Z, Ma L, Peng B, Mao K, Li C,
Su M, Zhou C and Peng G: Baicalein and baicalin inhibit colon
cancer using two distinct fashions of apoptosis and senescence.
Oncotarget. 9:20089–20102. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang CZ, Calway TD, Wen XD, Smith J, Yu C,
Wang Y, Mehendale SR and Yuan CS: Hydrophobic flavonoids from
Scutellaria baicalensis induce colorectal cancer cell apoptosis
through a mitochondrial-mediated pathway. Int J Oncol.
42:1018–1026. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang X, Wu P, Huang F, Xu M, Chen M,
Huang K, Li GP, Xu M, Yao D and Wang L: Baicalin attenuates chronic
hypoxia-induced pulmonary hypertension via adenosine A2A
receptor-induced SDF-1/CXCR4/PI3K/AKT signaling. J Biomed Sci.
24:52–66. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hong GE, Kim JA, Nagappan A, Yumnam S, Lee
HJ, Kim EH, Lee WS, Shin SC, Park HS and Kim GS: Flavonoids
identified from Korean Scutellaria baicalensis Georgi inhibit
inflammatory signaling by suppressing activation of NF- κB and MAPK
in RAW 264.7 cells. Evid Based Complement Alternat Med.
2013:912031–912042. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hussain I, Waheed S, Ahmad KA, Pirog JE
and Syed V: Scutellaria baicalensis targets the hypoxia-inducible
factor-1α and enhances cisplatin efficacy in ovarian cancer. J Cell
Biochem. 119:7515–7524. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Duan C, Matsumura S, Kariya N, Nishimura M
and Shimono T: In vitro antibacterial activities of Scutellaria
baicalensis Georgi against cariogenic bacterial. Pediatr Dent J.
17:58–64. 2017. View Article : Google Scholar
|
|
23
|
Kim SJ, Kim HJ, Kim HR, Lee SH, Cho SD,
Choi CS, Nam JS and Jung JY: Antitumor actions of baicalein and
wogonin in HT-29 human colorectal cancer cells. Mol Med Rep.
6:1443–1449. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen Z, Hou R, Gao S, Song D and Feng Y:
Baicalein inhibits proliferation activity of human colorectal
cancer cells HCT116 through downregulation of Ezrin. Cell Physiol
Biochem. 49:2035–2046. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rui X, Yan XI and Zhang K: Baicalein
inhibits the migration and invasion of colorectal cancer cells via
suppression of the AKT signaling pathway. Oncol Lett. 11:685–688.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chai Y, Xu J and Yan B: The
anti-metastatic effect of baicalein on colorectal cancer. Oncol
Rep. 37:2317–2323. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cui XB, Qian XC, Huang P, Zhang YX, Li JS,
Yang GM and Cai BC: Simultaneous determination of ten flavonoids of
crude and wine-processed Radix Scutellariae aqueous extracts in rat
plasma by UPLC-ESI-MS/MS and its application to a comparative
pharmacokinetic study. Biomed Chromatogr. 29:1112–1123. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang Y, Zhang Z and Song R: The influence
of compatibility of rhubarb and radix scutellariae on the
pharmacokinetics of anthraquinones and flavonoids in rat plasma.
Eur J Drug Metab Pharmacokinet. 43:291–300. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Aoki T, Nishimura M, Kataoka H, Ishibashi
R, Nozaki K and Miyamoto S: Complementary inhibition of cerebral
aneurysm formation by eNOS and nNOS. Lab Invest. 91:619–626. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Duan j, Song Z, Qi M, Bai X, Wang J, Zhang
Y, Zou X, Guo Q and Wan P: Increased autophagy levels mediate
cisplatin resistance in cisplatin-resistant cells while also
rendering them vulnerable to autophagy induction. BioMed Res Int.
10:1736738–1736748. 2018.
|
|
31
|
Qiu Y, Li C, Wang Q, Zeng X and Ji P:
Tanshinone IIA induces cell death via Beclin-1-dependent autophagy
in oral squamous cell carcinoma SCC-9 cell line. Cancer Med.
7:397–407. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Y, Yan W and Chen X: Mutant p53
disrupts MCF-10A cell polarity in three-dimensional culture via
epithelial-to-mesenchymal transitions. J Biol Chem.
286:16218–16228. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chang CJ, Chao CH, Xia W, Yang JY, Xiong
Y, Li CW, Yu WH, Rehman SK, Hsu JL, Lee HH, et al: p53 regulates
epithelial-mesenchymal transition and stem cell properties through
modulating miRNAs. Nat Cell Biol. 13:317–323. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fan F, Samuel S, Evans KW, Lu J, Xia L,
Zhou Y, Sceusi E, Tozzi F, Ye XC, Mani SA, et al: Overexpression of
snail induces epithelial-mesenchymal transition and a cancer stem
cell-like phenotype in human colorectal cancer cells. Cancer Med.
1:5–16. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sang Y, Cheng C, Zeng YX and Kang T: Snail
promotes metastasis of nasopharyngeal carcinoma partly by
down-regulating TEL2. Cancer Commun Lond. 38:58–68. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bermudezsoto MJ, Tomasbarberan FA and
Garciaconesa M: Stability of polyphenols in chokeberry (Aronia
melanocarpa) subjected to in vitro gastric and pancreatic
digestion. Food Chem. 102:865–874. 2007. View Article : Google Scholar
|
|
37
|
Sankaranarayanan R, Valiveti CK, Kumar DR,
Van slambrouck S, Kesharwani S, Seefeldt T, Scaria J, Tummala H and
Bhat G: Van slambrouck S, Kesharwani SS, Seefeldt T, Scaria J,
Tummala H and Bhat GJ: The flavonoid metabolite 2, 4,
6-Trihydroxybenzoic acid is a CDK inhibitor and an
anti-proliferative agent: A potential role in cancer prevention.
Cancers (Basel). 11:2–18. 2019. View Article : Google Scholar
|
|
38
|
Gao K, Xu A, Krul C, Venema K, Liu Y, Niu
Y, Lu J, Bensoussan L, Seeram NP, Heber D, et al: Of the major
phenolic acids formed during human microbial fermentation of tea,
citrus, and soy flavonoid supplements, only
3,4-dihydroxyphenylacetic acid has antiproliferative activity. J
Nutr. 136:52–57. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Du LY, Qian DW, Shang EX, Liu P, Jiang S,
Guo JM, Su SL, Duan JA, Xu J and Zhao M: UPLC-Q-TOF/MS-based
screening and identification of the main flavonoids and their
metabolites in rat bile, urine and feces after oral administration
of Scutellaria baicalensis extract. J Ethnopharmacol. 169:156–162.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Park KI, Park HS, Kang SR, Nagappan A, Lee
DH, Kim JA, Han DY and Kim GS: Korean Scutellaria baicalensis water
extract inhibits cell cycle G1/S transition by suppressing cyclin
D1 expression and matrix-metalloproteinase-2 activity in human lung
cancer cells. J Ethnopharmacol. 133:634–641. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang Y, Song L, Cai L, Wei R, Hu H and
Jin W: Effects of baicalein on apoptosis, cell cycle arrest,
migration and invasion of osteosarcoma cells. Food Chem Toxicol.
53:325–333. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kowalczyk E, Krzesiński P, Kura M,
Niedworok J, Kowalski J and Błaszczyk J: Pharmacological effects of
flavonoids from Scutellaria baicalensis. Przegl Lek. 63:95–96.
2006.(In Polish). PubMed/NCBI
|
|
43
|
Chao C, Saito S, Kang J, Anderson CW,
Appella E and Xu Y: p53 transcriptional activity is essential for
p53-dependent apoptosis following DNA damage. EMBO J. 19:4967–4975.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yilmaz M and Christofori G: EMT, the
cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev.
28:15–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu H, Dong Y, Gao Y, Du Z, Wang Y, Cheng
P, Chen A and Huang H: The fascinating effects of baicalein on
cancer: A review. Int J Mol Sci. 17:1681–1699. 2016. View Article : Google Scholar :
|