Introduction
Acute myocardial infarction (AMI), resulting from
sudden interruption of blood flow in the main coronary arteries
remains a leading cause of morbidity and mortality worldwide
(1). Currently, percutaneous
coronary intervention (PCI) is the most effective minimally
invasive treatment for AMI. PCI is performed in order to reopen the
affected coronary artery and restore adequate blood flow to the
ischemic myocardium. It aims to reduce the myocardial infarct size
and to maintain heart function (2,3).
However, following PCI treatment, ~4-7% of the patients experience
recurrent chest pain, elevated troponin I/T levels and onset of
hemodynamic disorders that are attributed to myocardial
ischemia-reperfusion (I/R) injury (4). Cardiac I/R is characterized by
restricted blood flow to the myocardium followed by reoxygenation
associated with blood reperfusion when the coronary artery reopens
(5). I/R injury is caused by
mitochondrial dysfunction, calcium overload and excessive
generation of mitochondrial reactive oxygen species (ROS), which
eventually contribute to cardiomyocyte apoptosis and necrosis
(6). Therefore, the prevention of
myocardial I/R injury is essential for improving prognosis
following PCI treatment.
Ginkgolide B (GB) is extracted from the leaves of an
ancient Chinese medicinal plant Ginkgo. This compound is a
diterpene lactone, which has been documented as a strong antagonist
of platelet-activating factor receptor (7). Previous studies have shown that the
anti-inflammatory, antioxidant and anti-apoptotic properties of GB
have rendered it beneficial in ischemic and hemorrhagic stroke
(8–11). Furthermore, it has been shown to
improve cognitive function and the disease outcome of different
types of cancer. Moreover, previous studies have demonstrated that
GB plays a therapeutic role in cardiovascular diseases (12,13).
For instance, GB can ameliorate oxidized low-density
lipoprotein-induced endothelial cell dysfunction in human umbilical
vein endothelial cells and inhibit inflammatory cascades in murine
RAW264.7 macrophage-like cells (12). In addition, GB inhibits the
production of ROS in doxorubicin-induced cardiotoxicity in H9c2
cells (13). However, the effect
of GB on myocardial I/R injury remains unclear.
PI3Ks are a family of lipid kinases involved in the
regulation of cellular activation, inflammatory responses and
apoptosis (14). PI3K activation
induces the formation of the second messenger
phosphatidylinositol-3,4,5-triphosphate (PIP3) (15). PIP3 provides an Akt docking site
for Akt phosphorylation and activation (16). As the central mediator of the PI3K
signaling pathway, Akt promotes cell survival, apoptosis and
proliferation by inducing the downstream mTOR complex (17). The mTOR signaling pathway is also
involved in regulating cell growth, cell survival and protein
synthesis (18). Therefore, the
regulation of the PI3K/Akt/mTOR signaling pathway could affect cell
viability and the induction of apoptosis on different cell
types.
Hydrogen peroxide (H2O2) is a
key metabolite in oxidative stress that is widely used to simulate
myocardial I/R injury (19). The
H9c2 cell line, derived from a rat embryonic heart ventricle, is
considered a close surrogate for cardiomyocytes and has been proven
to be ideal for cardiomyocyte signal transduction studies (20). Therefore, in the present study,
H2O2 treatment in H9c2 cardiac cells was used
to simulate myocardial I/R. In addition, the present study
investigated the cardioprotective effect of GB on
H2O2-induced apoptosis in H9c2 cells and its
interaction with the PI3K/Akt/mTOR signaling pathway.
Materials and methods
Cell culture
H9c2 rat cardiomyocyte cells were purchased from The
Cell Bank of Type Culture Collection of the Chinese Academy of
Sciences and cultured in high-glucose DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and 100 U/ml
streptomycin at 37°C in a humidified atmosphere containing 5%
CO2. The cells were passaged every 4 days and the
culture medium was replaced every 2 days.
Establishment of H9c2 cell oxidative
stress model
H9c2 cells (5×103 cells per well) were
seeded in 96-well plates and incubated at 37°C in a 5%
CO2 humidified incubator overnight. Subsequently, the
cells were exposed to different H2O2
concentrations (200, 400, 600 and 800 µM) and harvested at
different time points (4, 8 and 12 h). The appropriate
concentration and exposure time of H2O2 used
for the establishment of the oxidative stress model in H9c2 cells
was determined by the Cell Counting Κit-8 (CCK-8) assay.
Drug preparation and cell
treatment
GB was purchased from Sigma-Aldrich; Merck KGaA. The
stock solution was prepared by dissolving GB in DMSO at 100 mM. The
working solution of GB was obtained by diluting the stock solution
in DMEM to the desired concentrations. To avoid the DMSO-induced
cytotoxicity, the final concentration of DMSO was retained to
<1%. H9c2 cells were pretreated with GB for 20 h prior to
co-incubation with H2O2. The total time of
GB-pretreatment group is the same as single GB-treated group. Prior
to cell pretreatment with GB at 0.01, 0.1, 1, 10 and 100 µM for 1
h, the cells were treated with PI3K inhibitor LY294002 (Selleck
Chemicals) at concentrations of 5, 10, 20 and 40 µM.
Cell viability assay
Cell viability was determined by the CCK-8 assay
(Dojindo Molecular Technologies, Inc.), according to the
manufacturer's protocol. H9c2 cells (5×103 cells per
well) were seeded in 96-well plates overnight and were pretreated
with different concentrations of LY294002, GB and
H2O2. To measure cell viability, 10 µl CCK-8
assay solution was added into each well, containing 90 µl medium,
and the cells were then incubated in the dark at 37°C for
additional 2 h. Absorbance was measured at 450 nm in a microplate
reader (BioTek Instruments, Inc.).
Hoechst 33342 staining
Typical morphological features of apoptotic cells
were observed by Hoechst 33342 staining (Nanjing KeyGen Biotech
Co., Ltd.). Briefly, the cells were washed twice with PBS following
H2O2 incubation with or without GB
pretreatment and stained with Hoechst 33342 for 15 min in a 5%
CO2 incubator at 37°C in the dark. Finally, the cells
were washed once with PBS and the images were captured by
fluorescence microscopy (magnification, ×200; Zeiss Axiovert A1).
The apoptotic cells with nuclear chromatin condensation and
fragmentation were stained bright blue. A total of three
fluorescence images per well were captured randomly, and the
percentage of apoptotic cells was calculated using the following
equation: (The number of apoptotic cells/the total number of cells)
×100.
Annexin V-FITC/propidium iodide (PI)
assay
The induction of apoptosis was examined using the
Annexin V-FITC/PI double staining assay (BD Biosciences;
Becton-Dickinson and Company). Briefly, H9c2 cells were gently
washed with ice-cold PBS, digested with 0.25% Trypsin without
ethylenediaminetetraacetic acid (Gibco; Thermo Fisher Scientific,
Inc.) and resuspended in complete culture medium. The cells were
then centrifuged at 120 × g for 5 min at 4°C and the cell pellet
was resuspended in ice-cold PBS and centrifuged at the same
conditions. Finally, the cells were resuspended in 500 µl 1X
binding buffer supplemented with 5 µl Annexin V-FITC and 5 µl PI.
The cells were incubated at room temperature for 15 min in the dark
and were analyzed by flow cytometry (BD FACSCalibur™; BD
Biosciences; Becton-Dickinson and Company). Data was analyzed using
FlowJo version 7.6.1 software (FlowJo LLC).
Western blot analysis
H9c2 cells were washed with ice-cold PBS, pelleted
by cell scraper and lysed in 100 µl lysis buffer containing 1 µl
phosphatase inhibitor, 0.1 µl protease inhibitor and 0.5 µl PMSF.
The extracted proteins were quantified with the bicinchoninic acid
Protein assay kit (Thermo Fisher Scientific, Inc.). A total of 20
µg protein extracts per lane were separated by 6–15% SDS-PAGE. The
proteins were transferred onto polyvinylidene difluoride membranes
or nitrocellulose filter membrane. The membranes were blocked with
5% BSA (Gibco; Thermo Fisher Scientific, Inc.) at room temperature
for 2 h and incubated overnight at 4°C with primary antibodies
against rabbit anti-Bax (1:1,000; cat. no. 2772), rabbit anti-Bcl-2
(1:1,000; cat. no. 2870), rabbit anti-cleaved caspase-3 (1:1,000;
cat. no. 9664), rabbit anti-caspase-3 (1:1,000; cat. no. 9662),
rabbit anti-Akt (1:1,000; cat. no. 9272), rabbit
anti-phosphorylated (p)-Akt (p-Akt; 1:1,000; cat. no. 4060), rabbit
anti-mTOR (1:1,000; cat. no. 2983), rabbit anti-p-mTOR (p-mTOR;
1:1,000; cat. no. 5536), and rabbit anti-GAPDH (1:1,000; cat. no.
5174; all from Cell Signaling Technology, Inc.). The membranes were
subsequently washed for 2 h with TBST (10 mM Tris, pH 7.5, 150 mM
NaCl, and 0.05% Tween-20), and incubated with horseradish
peroxidase-conjugated goat anti-rabbit secondary antibody (1:500;
cat. no. sc-2004; Santa Cruz Biotechnology, Inc.) for 1 h at room
temperature. The antigen-antibody complexes on the membranes were
detected using the SuperSignal™ West Femto Maximum Sensitivity
substrate (Thermo Fisher Scientific, Inc.) and quantified on the
ChemiDoc™ XRS Imaging system (Bio-Rad Laboratories, Inc.).
Statistical analysis
All experiments were repeated three times. The data
are presented as the mean ± SD. Statistical differences among
groups were analyzed using a one-way ANOVA, followed by a Tukey's
post-hoc test. All statistical analyses were performed using the
GraphPad Prism 5 software (GraphPad Software, Inc.). P<0.05 or
P<0.001 were considered to indicate a statistically significant
difference.
Results
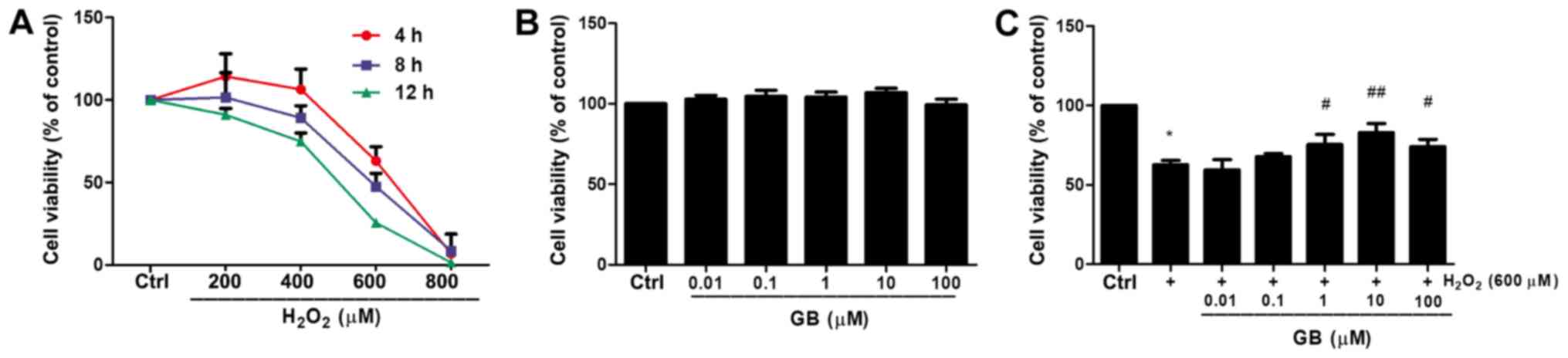
GB decreases
H2O2-induced cytotoxicity in H9c2 cells
H2O2 reduced cell viability in
a dose- and time- dependent manner (Fig. 1A). For instance, treatment of H9c2
cells with 600 µM H2O2 for 4 h reduced cell
viability to 63.11% compared with that noted in untreated cells.
The H9c2 cell state in this condition was not too poor to conduct
subsequent experiments. Therefore, 600 µΜ
H2O2 was selected as the optimal treatment
concentration for subsequent experiments. The effect of GB alone on
cell viability was also investigated. Cell viability was not
significantly affected by GB treatment for 24 h even at
concentrations up to 100 µM (Fig.
1B). Subsequently, the effects of H9c2 cell-GB pretreatment in
the protection of the cells against
H2O2-induced cytotoxicity were examined. GB
pretreatment at 10 µM concentration for 20 h prior to co-incubation
with 600 µM H2O2 for 4 h revealed the highest
inhibitory effect on the induction of cell cytotoxicity caused by
treatment of 600 µM H2O2 for 4 h (Fig. 1C). Therefore, 10 µM GB was used in
subsequent experiments.
GB reduces
H2O2-induced cell apoptosis in H9c2
cells
Chromatin condensation and fragmentation are the
typical features of apoptotic cells so Hoechst 33342 staining was
used to evaluate induction of cell apoptosis caused by
H2O2. Pretreatment of the cells with 10 µM GB
for 20 h significantly decreased the percentage of apoptotic cells,
compared with that noted following treatment of the cells with 600
µM H2O2 for 4 h (Fig. 2A and B). Furthermore, the Annexin
V-FITC/PI assay was further used to evaluate the induction of cell
apoptosis. Treatment of the cells with 600 µM
H2O2 for 4 h led to a significant increase in
cell apoptosis compared with that noted in the control group,
whereas this effect was inhibited by GB pretreatment (Fig. 2C and D). The expression levels of
the apoptotic proteins Bcl-2, Bax, caspase-3 and cleaved caspase-3,
were determined by western blot analysis. Both Bax and cleaved
caspase-3 protein levels were significantly elevated in the
H2O2-treated group, whereas Bcl-2 levels were
downregulated (Fig. 2E-H). In
addition, GB pretreatment significantly decreased the expression
levels of Bax and cleaved caspase-3 and increased the expression
levels of Bcl-2 compared with those of the
H2O2-treated group. Both the Bax/Bcl-2 and
the cleaved caspase-3/caspase-3 ratios were significantly decreased
in the GB-pretreatment group compared with those of the
H2O2-treated group.
GB enhances the phosphorylation levels
of Akt and mTOR
The expression levels of p-Akt and p-mTOR were
downregulated in the H2O2-treated group,
whereas GB pretreatment was able to reverse the effects of
H2O2 (Fig. 3A
and B).
Effect of the PI3K inhibitor LY294002
on the H2O2-induced cytotoxicity in H9c2
cells
To determine whether the activation of Akt and mTOR
was associated with the protective effects of GB, the LY294002
inhibitor was used to investigate the expression levels of p-Akt
and to assess cell viability. Pretreatment of the cells with
LY294002 and GB downregulated the expression levels of p-Akt in a
dose-dependent manner, compared with those noted in the single
GB-pretreatment group. In addition, LY294002 and GB pretreatment
reduced cell viability compared with that of the single
GB-pretreatment group. These results indicated that GB elicited its
protective effects via activation of the PI3K/Akt/mTOR signaling
pathway (Fig. 4A and B).
Discussion
The present study provides evidence regarding the
protective effects of GB on H2O2-induced
cytotoxicity in H9c2 cells. These effects were mediated by the
inhibition of cell apoptosis. Furthermore, it was shown that the
GB-induced protective effect was mediated via activation of the
PI3K/Akt/mTOR signaling pathway.
The process of apoptosis that was initially
described by Kerr et al (21) is a form of programmed cell death
with certain morphological features, such as narrowed cell volume,
chromatin condensation, nuclear fragmentation and apoptotic body
formation (22). In the present
study, GB inhibited the induction of cell apoptosis by
H2O2. Two major pathways of apoptosis, namely
the death receptor-mediated and the mitochondrial-mediated
apoptotic pathways have been identified. Both pathways result in
caspase-dependent cell death (23). The members of the Bcl-2 family of
proteins, which is composed of anti- and pro-apoptotic factors, are
involved in the mitochondrial-mediated apoptotic pathway (24). Bax is a pro-apoptotic protein of
the Bcl-2 family that is negatively regulated by Bcl-2
(anti-apoptotic protein). Consequently, the Bax/Bcl-2 ratio can act
as an indicator that determines the cell susceptibility to
apoptosis and the balance between anti- and pro-apoptotic factors
(25). Caspase-3 is one of the
most important members of the caspase family and is considered the
central effector of apoptosis activated by upstream initiator
caspases. Caspase-3 is cleaved to produce the final cleaved
caspase-3 protein form (26). In
the present study, GB pretreatment significantly decreased cleaved
caspase-3 and Bax expression levels, whereas it upregulated Bcl-2
expression levels in H2O2-treated H9c2 cells,
resulting in a declined Bax/Bcl-2 ratio and increased cell
viability. These results indicated that GB exhibited protective
effects against the H2O2-induced cytotoxicity
in H9c2 cells partly through its anti-apoptotic properties.
Previous studies have shown that the activation of
the PI3K/Akt/mTOR signaling pathway promotes cell proliferation and
inhibits cell apoptosis (27,28).
In the current study, GB pretreatment inhibited cell apoptosis by
inducing Akt and mTOR phosphorylation. To further confirm this
observation, H9c2 cells were treated with the PI3K inhibitor
LY294002 and it was shown that the GB-induced Akt phosphorylation
was partially blocked by the LY294002 inhibitor. In addition,
LY294002 treatment partially reversed the protective effect of GB
in maintaining cell viability. The aforementioned results suggested
that GB exerted protective effects against cell apoptosis via the
activation of the PI3K/Akt/mTOR signaling pathway.
All in vitro experiments were repeated three
times in this study, this is a limitation of the study so in future
experiments there should be a higher number of repeats. Additional
in vivo and clinical studies are also required to support
the in vitro results reported in the current study. Previous
studies demonstrated that PCI treatment followed by remote ischemic
preconditioning (RIPC) exhibited protective effects on myocardial
I/R injury (29) and
contrast-induced nephropathy (CIN) (30,31).
Furthermore, the activation of Akt may mediate the target organ
protection by RIPC (32). The
present study suggested that GB pretreatment could trigger the
activation of Akt during oxidative stress. In conclusion, GB
pretreatment could be used to alleviate myocardial I/R injury and
CIN following PCI treatment. However, additional clinical trials
need to be conducted in the future in order to confirm this
hypothesis.
Acknowledgements
Not applicable.
Funding
The present study was funded by the grant from
Jiangsu Province Nature Science Youth Foundation (grant no.
BK20141020).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JL and PW performed the majority of the experiments
and drafted the manuscript; ZX, JZ and JL performed some of the
experiments and collected the data; and JL and ZY designed the
study. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Anderson JL and Morrow DA: Acute
myocardial infarction. N Engl J Med. 376:2053–2064. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Canfield J and Totary-Jain H: 40 years of
percutaneous coronary intervention: History and future directions.
J Pers Med. 8:E332018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhou QL, Teng F, Zhang YS, Sun Q, Cao YX
and Meng GW: FPR1 gene silencing suppresses cardiomyocyte apoptosis
and ventricular remodeling in rats with ischemia/reperfusion injury
through the inhibition of MAPK signaling pathway. Exp Cell Res.
370:506–518. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Luo Y, Pan YZ, Zeng C, Li GL, Lei XM, Liu
Z and Zhou SF: Altered serum creatine kinase level and cardiac
function in ischemia-reperfusion injury during percutaneous
coronary intervention. Med Sci Monit. 17:Cr474–Cr479. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hausenloy DJ and Yellon DM: Myocardial
ischemia-reperfusion injury: A neglected therapeutic target. J Clin
Invest. 123:92–100. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Basheer WA, Fu Y, Shimura D, Xiao S,
Agvanian S, Hernandez DM, Hitzeman TC, Hong T and Shaw RM: Stress
response protein GJA1-20k promotes mitochondrial biogenesis,
metabolic quiescence, and cardioprotection against
ischemia/reperfusion injury. JCI Insight. 3:1219002018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nabavi SM, Habtemariam S, Daglia M, Braidy
N, Loizzo MR, Tundis R and Nabavi SF: Neuroprotective effects of
ginkgolide B against ischemic stroke: A review of current
literature. Curr Top Med Chem. 15:2222–2232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gill I, Kaur S, Kaur N, Dhiman M and
Mantha AK: Phytochemical ginkgolide B attenuates amyloid-β1-42
induced oxidative damage and altered cellular responses in human
neuroblastoma SH-SY5Y cells. J Alzheimers Dis. 60 (Suppl
1):S25–S40. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhi Y, Pan J, Shen W, He P, Zheng J, Zhou
X, Lu G, Chen Z and Zhou Z: Ginkgolide B inhibits human bladder
cancer cell migration and invasion through MicroRNA-223-3p. Cell
Physiol Biochem. 39:1787–1794. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nash KM and Shah ZA: Current perspectives
on the beneficial role of ginkgo biloba in neurological and
cerebrovascular disorders. Integr Med Insights. 10:1–9. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shu ZM, Shu XD, Li HQ, Sun Y, Shan H, Sun
XY, Du RH, Lu M, Xiao M, Ding JH and Hu G: Ginkgolide B protects
against ischemic stroke via modulating microglia polarization in
mice. CNS Neurosci Ther. 22:729–739. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Feng Z, Yang X, Zhang L, Ansari IA, Khan
MS, Han S and Feng Y: Ginkgolide B ameliorates oxidized low-density
lipoprotein-induced endothelial dysfunction via modulating
Lectin-like ox-LDL-receptor-1 and NADPH oxidase 4 expression and
inflammatory cascades. Phytother Res. 32:2417–2427. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gao J, Chen T, Zhao D, Zheng J and Liu Z:
Ginkgolide B exerts cardioprotective properties against
doxorubicin-induced cardiotoxicity by regulating reactive oxygen
species, akt and calcium signaling pathways in vitro and in vivo.
PLoS One. 11:e01682192016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu X, Xu C, Zhou X, Cui B, Lu Z and Jiang
H: PI3K/Akt signaling pathway involved in cardioprotection of
preconditioning with high mobility group box 1 protein during
myocardial ischemia and reperfusion. Int J Cardiol. 150:222–223.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Latronico MV, Costinean S, Lavitrano ML,
Peschle C and Condorelli G: Regulation of cell size and contractile
function by AKT in cardiomyocytes. Ann NY Acad Sci. 1015:250–260.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yao H and Han X and Han X: The
cardioprotection of the insulin-mediated PI3K/Akt/mTOR signaling
pathway. Am J Cardiovasc Drugs. 14:433–442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Aoyagi T and Matsui T: Phosphoinositide-3
kinase signaling in cardiac hypertrophy and heart failure. Curr
Pharm Des. 17:1818–1824. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sciarretta S, Forte M, Frati G and
Sadoshima J: New insights into the role of mTOR signaling in the
cardiovascular system. Circ Res. 122:489–505. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang X, Jiang H and Shi Y: Upregulation of
heme oxygenase-1 expression by curcumin conferring protection from
hydrogen peroxide-induced apoptosis in H9c2 cardiomyoblasts. Cell
Biosci. 7:202017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Watkins SJ, Borthwick GM and Arthur HM:
The H9C2 cell line and primary neonatal cardiomyocyte cells show
similar hypertrophic responses in vitro. In Vitro Cell Dev Biol
Anim. 47:125–131. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kerr JF, Wyllie AH and Currie AR:
Apoptosis: A basic biological phenomenon with wide-ranging
implications in tissue kinetics. Br J Cancer. 26:239–257. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Takemura G, Kanoh M, Minatoguchi S and
Fujiwara H: Cardiomyocyte apoptosis in the failing heart-a critical
review from definition and classification of cell death. Int J
Cardiol. 167:2373–2386. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mughal W, Dhingra R and Kirshenbaum LA:
Striking a balance: Autophagy, apoptosis, and necrosis in a normal
and failing heart. Curr Hypertens Rep. 14:540–547. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Watson EC, Grant ZL and Coultas L:
Endothelial cell apoptosis in angiogenesis and vessel regression.
Cell Mol Life Sci. 74:4387–4403. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lomonosova E and Chinnadurai G: BH3-only
proteins in apoptosis and beyond: An overview. Oncogene. 27 (Suppl
1):S2–S19. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Edlich F: BCL-2 proteins and apoptosis:
Recent insights and unknowns. Biochem Biophys Res Commun.
500:26–34. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chang J, Xue X, Song C, Liu B and Gao L:
Ginkgolide B promotes cell growth in endothelial progenitor cells
through miR-126 and the Akt signaling pathway. Mol Med Rep.
16:5627–5632. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fulda S: Synthetic lethality by
co-targeting mitochondrial apoptosis and PI3K/Akt/mTOR signaling.
Mitochondrion. 19:85–87. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Slagsvold KH, Moreira JB, Rognmo O, Hoydal
M, Bye A, Wisloff U and Wahba A: Remote ischemic preconditioning
preserves mitochondrial function and activates pro-survival protein
kinase Akt in the left ventricle during cardiac surgery: A
randomized trial. Int J Cardiol. 177:409–417. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Moretti C, Cerrato E, Cavallero E, Lin S,
Rossi ML, Picchi A, Sanguineti F, Ugo F, Palazzuoli A, Bertaina M,
et al: The EUROpean and Chinese cardiac and renal remote ischemic
preconditioning study (EURO-CRIPS CardioGroup I): A randomized
controlled trial. Int J Cardiol. 257:1–6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pistolesi V, Regolisti G, Morabito S,
Gandolfini I, Corrado S, Piotti G and Fiaccadori E: Contrast medium
induced acute kidney injury: A narrative review. J Nephrol.
31:797–812. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao Y, Zheng ZN, Pi YN, Liang X and Jin
SQ: Cardioprotective effects of transfusion of late-phase
preconditioned plasma may be induced by activating the reperfusion
injury salvage kinase pathway but not the survivor activating
factor enhancement pathway in rats. Oxid Med Cell Longev.
2017:85265612017. View Article : Google Scholar : PubMed/NCBI
|