Introduction
Colorectal cancer (CRC) is the third most diagnosed
cancer worldwide and the fourth leading cause of cancer-related
mortality, accounting for >1.4 million new cases and 700,000
deaths annually (1). Thanks to
improved screening programs and standardized treatments, the
mortality rate of CRC has declined in the past 30 years; however,
the recurrence and metastasis still contribute to a poor response
to CRC treatment, with only 50% of patients surviving 5 years
(2–4). Better understanding of CRC
pathogenesis is therefore urgently required in order to develop
novel effective therapeutic strategies.
MicroRNAs (miRNAs/miRs) are a group of small,
non-coding RNAs of 18–25 nucleotides in length, which can
negatively regulate targeted genes at the post-transcriptional
level by directly binding to their 3′-untranslated region (3′-UTR)
(5). It has been reported that
miRNAs are highly involved in various biological processes,
including cell differentiation, apoptosis and metabolism (6). Previous studies demonstrated that
aberrantly expressed miRNAs may be closely associated with various
types of human cancer, in which miRNAs were reported to be involved
in tumor initiation and development (7,8). For
example, miR-324-3p is aberrantly high-expressed in gastric cancer,
and overexpression of miR-324-3p promotes cell growth, migration
and decreased apoptosis of gastric cancer (8); miR-125a is downregulated in cervical
cancer, and serves an important role in its cell proliferation and
progression (9). In addition,
miRNAs have been reported to serve as effective biomarkers for the
early diagnosis and prognosis of patients with cancer (10). A previous study demonstrated that
miR-576-5p might be a potential tumor-promoting oncomiR in several
types of human tumor (11). Kordass
et al (11) reported that
miR-576-5p enhances the invasion of melanoma cells in vitro.
In addition, miR-576-5p was also found to be upregulated in
esophageal cancer, where it was found to be involved in cancer cell
migration and invasion, suggesting that it might serve as a
predictor of cancer prognosis (12,13).
However, the role of miR-576-5p in CRC remains unknown. In a
previous study using a miRNA microassay from 50 patients with colon
cancer and 44 healthy controls, the expression level of miR-576-5p
in patients was significantly higher compared with controls
(14). Furthermore, miR-576-5p
expression levels are more highly expressed at TNM stage III/IV
compared with TNM stage I/II, suggesting that miR-576-5p might be
considered as an effective diagnostic marker for CRC (14). However, the exact role of miR-576-5p
in CRC remains unclear and the biological functions of miR-576-5p
are unknown. Further investigation is therefore required to
determine whether miR-576-5p could regulate the biological
functions of CRC cells, in addition to understanding the underlying
mechanism.
The present study performed a series of in
vitro experiments to determine the role and underlying
mechanism of miR-576-5p in CRC and its effect on CRC development.
The findings from this study may provide a novel therapeutic target
and serve the development of novel strategy for the treatment of
CRC.
Materials and methods
Data analyses and bioinformatics
miRNA-576-5p expression from 459 CRC samples and 8
normal samples (adjacent cancer tissues) were retrieved from the
TCGA data portal (https://tcga-data.nci.nih.gov.).
Cell culture and transfection
The human normal colonic epithelial cell line NCM460
and the human CRC cell lines HCT116, SW620 and SW480, were obtained
from The Cell Bank of Type Culture Collection of the Chinese
Academy of Sciences. Cells were cultured in DMEM, supplemented with
10% FBS, and 1% penicillin-streptomycin (all from Gibco; Thermo
Fisher Scientific, Inc.), and maintained in a humidified chamber
with 5% CO2 at 37°C.
miR-576-5p mimic (miR-576-5p), miR-576-5p inhibitor
(anti-miR-576-5p) and corresponding negative control (NC), as well
as pcDNA3.1-Wnt5a (Wnt5a) and its negative control (vector), were
synthesized by Shanghai GenePharma Co., Ltd. SW480 cells were
seeded in 6-well plates at a density of 1×106 cells/well
and cultured at 37°C for 24 h. SW480 cells were subsequently
transfected with 100 nM miR-576-5p mimic, 100 nM miR-576-5p
inhibitor, 100 nM negative control, as well as 10 nM pcDNA3.1-Wnt5a
and its vector using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. After 48 h, the expression of miR-576-5p and Wnt5a could
be observed, and the transfected cells were collected for
subsequent experiments.
MTT assay
SW480 Cells were seeded into 96-well plates at a
density of 1×103 cells/well and were cultured in a
humidified incubator with 5% CO2 at 37°C for 48 h.
Subsequently, 10 µl MTT solution (5 mg/ml; Beyotime Institute of
Biotechnology) was added into each well and incubated for 4 h.
Subsequently, 100 µl DMSO was then added to each well to dissolve
the formazan crystals. The absorbance was measured at 490 nm using
a microplate reader.
Wound healing assay
Cells were plated into 6-well plates in DMEM
supplemented with 10% FBS. Once cells reached 85% confluence, a
20-µl pipette tip was used to make a wound in the cell monolayer.
Cells were washed twice with PBS and incubated with complete medium
without FBS. After 24 h, the wound healing process was monitored
under a phase contrast microscope (magnification, ×100). The
migratory distance was analyzed to quantify the cell migration
ability.
Transwell assay
Transwell chambers (8-µm pore; Corning Inc.)
precoated with 50 µl Matrigel (BD Biosciences) were placed into
24-well plates. SW480 cells (1×105) were plated into the
upper chamber without serum, whereas complete medium supplemented
with 10% FBS was placed into the lower chamber. Following
incubation for 24 h, cells remaining in the upper chamber were
removed carefully with a cotton swab. Cells that have invaded the
lower chamber were stained with crystal violet and the invasive
cells were counted under a light microscope (magnification,
×100).
Reverse transcription-quantitative
(RT-q) PCR
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), according to the manufacturer's instructions. Total RNA was
reverse transcribed into cDNA using the PrimeScript™ 1st Strand
cDNA Synthesis kit (Takara Bio, Inc.). The expression levels were
determined by qPCR using the 7500 Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The thermal cycling
conditions were: Initial denaturation at 95°C for 10 min, followed
by 32 cycles of denaturation at 95°C for 15 sec, annealing at 60°C
for 30 sec and extension at 75°C for 40 sec. The primers were
obtained from Sangon Biotech (Shanghai, China) and the primer
sequences were: miR-576-5p, forward, 5′-TTGGGTCAAGAGTCAGAAGTTT-3′
and reverse, 5′-TGGCTTCTACTTGTCCTTTCC-3′; Wnt5a, forward,
5′-ATGCAGTACATTGGAGAAGGTG-3′, reverse, 5′-CGTCTCTCGGCTGCCTATTT-3′;
U6, forward, 5′-AAAGCAAATCATCGGACGACC-3′, reverse,
5′-GTACAACACATTGTTTCCTCGGA-3′; GAPDH, forward,
5′-ATCATCCCTGCCTCTACTGG-3′; reverse, 5′-GTCAGGTCCACCACTGACAC-3′.
Relative mRNA level was quantified using 2−ΔΔCq method
(15). U6 and GAPDH were used as
internal control.
Western blotting
Total protein was extracted from cells using RIPA
lysis buffer (Cell Signaling Technology, Inc.). Total protein was
measured using a bicinchoninic acid assay kit (Pierce; Thermo
Fisher Scientific, Inc.). Equal amounts of proteins (30 µg/lane)
were separated by 12% SDS-PAGE and subsequently transferred onto
nitrocellulose membranes. Membranes were blocked with 5% nonfat
dried milk at room temperature for 1 h and incubated with the
primary antibodies against Wnt/β-catenin signaling-related
proteins: Wnt5a (1:1,000; cat. no. ab179824), β-catenin (1:5,000;
cat. no. ab32572), cyclin D1 (1:200; cat. no. ab16663), c-myc
(1:1,000; cat. no. ab32072), p-c-Jun (1:1,000; cat. no. ab32385)
and c-Jun (1:1,000; cat. no. ab40766); epithelial-to-mesenchymal
transition (EMT)-related proteins: E-cadherin (1:1,000; cat. no.
ab40772), vimentin (1:1,000; cat. no. ab92547), N-cadherin
(1:5,000; cat. no. ab76011) and Snail (1:1,000; cat. no. ab216347);
GAPDH (1:1,000; cat. no. ab8245 all from Abcam) at 4°C overnight.
Membranes were then incubated with the corresponding horseradish
peroxidase-conjugated anti-rabbit (1:2,000; cat. no. sc-2004) or
anti-mouse IgG secondary antibodies (1:2,000; cat. no. sc-2005;
both Santa Cruz Biotechnology, Inc.) at room temperature for 2 h.
Bands were visualized using enhanced chemiluminescence system
(Pierce; Thermo Fisher Scientific, Inc.) and analyzed using a
LAS-4000 image document instrument (FUJIFILM Wako Pure Chemical
Corporation). ImageJ software version 1.46 (National Institutes of
Health) was used to analyze the gray value of protein bands and the
relative expression levels were normalized to endogenous control
GAPDH.
Dual-luciferase reporter assay
TargetScan (http://www.targetscan.org/) was used to predict the
putative binding sites between miR-576-5p and Wnt5a, which was then
verified by dual-luciferase reporter assay. Wild-type Wnt5a
(Wnt5a-WT) and mutated Wnt5a (Wnt5a-MUT) were cloned into the
pMIR-REPORT luciferase vectors (Ambion; Thermo Fisher Scientific,
Inc.). After the SW480 cells reached a density of 80%, they were
co-transfected with 100 ng Wnt5a-WT/Wnt5a-Mut and 150 nM miR-576-5p
mimic/miR-NC using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. After transfection for 48 h, luciferase activity was
analyzed using Dual-Luciferase Assay Kit (Promega Corporation). The
relative luciferase activity was normalized to Renilla
luciferase activity.
Statistical analysis
Statistical analysis was performed using SPSS 19.0
software (IBM Corp.) and data were presented as the means ±
standard deviation. Statistical differences were determined using
Student's t test for comparison between two groups and a one-way
ANOVA followed by a Tukey's post hoc test for multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-576-5p expression level is
upregulated in CRC tissues
TCGA database was used to determine the expression
level of miR-576-5p in clinical samples. As presented in Fig. 1A, the expression level of miR-576-5p
in CRC tissues was significantly increased compared with that of
normal tissues. Furthermore, miR-576-5p expression level was
expressed at higher levels in CRC cell lines HCT116, SW620 and
SW480 compared with the human normal colonic epithelial cell line
NCM460 (Fig. 1B). Since SW480 cells
expressed the highest level of miR-576-5p, this cell line was
selected in subsequent experiments.
miR-576-5p inhibition inhibits cell
viability and migratory and invasive abilities
To investigate the biological role of miR-576-5p in
SW480 cells, miR-576-5p was inhibited (Fig. 1C). The cell viability was
significantly decreased following inhibition of miR-576-5p
(Fig. 1D). In addition, miR-576-5p
inhibition significantly decreased the migratory and invasive
abilities of SW480 cells in the wound healing and Transwell assays,
respectively (Fig. 1E-H).
miR-576-5p inhibition may therefore alter CRC cell biology.
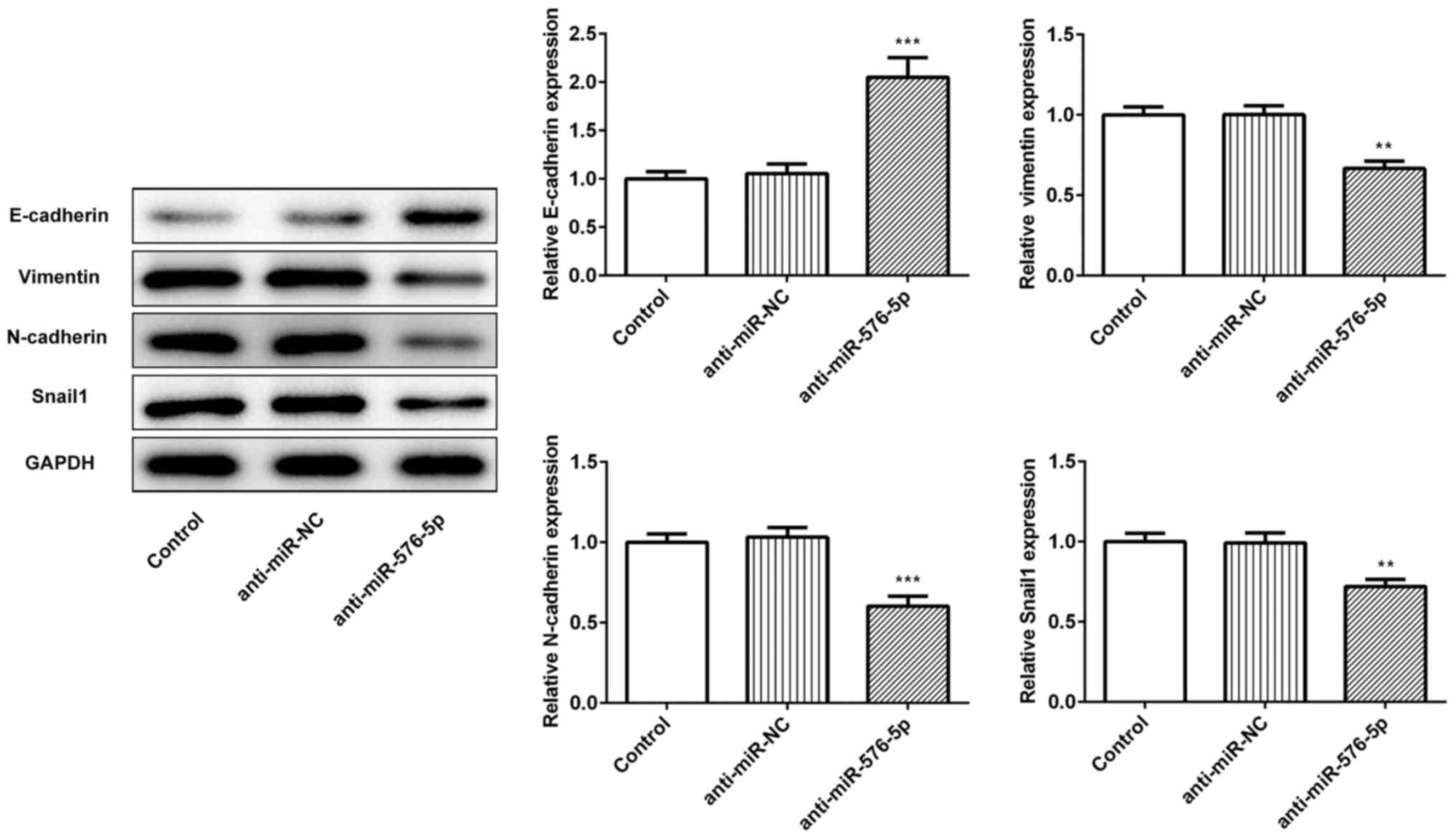
Inhibition of miR-576-5p reduces
EMT
EMT serves a pivotal role in the development of
metastatic tumors and it is responsible for most cancer-associated
mortality cases, which are usually associated with cell invasion
(16). In the present study, the
protein expression of E-cadherin, vimentin, N-cadherin and Snail
was assessed using western blotting to determine whether miR-576-5p
could induce EMT in SW480 cells. The results demonstrated that the
expression of the epithelial cell adhesion molecule E-cadherin was
significantly upregulated following miR-576-5p was inhibited,
whereas the expression of its transcriptional repressor Snail was
downregulated (Fig. 2).
Furthermore, upon inhibition of miR-576-5p, the expression of
mesenchymal-associated proteins, including vimentin and N-cadherin,
was significantly decreased, suggesting that miR-576-5p inhibition
may successfully suppress EMT.
miR-576-5p targets Wnt5a and regulates
the β-catenin signaling pathway
To further investigate the potential mechanism of
miR-576-5p function in SW480 cells, TargetScan (http://www.targetscan.org/) was used to identify the
putative genes which had a possible binding site on miR-576-5p.
Wnt5a was predicted to have binding site on miR-576-5p (Fig. 3A). To verify the interaction between
miR-576-5p and Wnt5a, 3′UTR reporter vectors (WT; Wnt5a 3′UTR)
containing the predicted sequences were constructed. The dual
luciferase reporter assay demonstrated that miR-576-5p mimic
significantly decreased the relative Renilla luciferase activity,
which was not the case when the predicted sites were mutated (MUT;
Wnt5a 3′UTR; Fig. 3B). The mRNA
level of miR-576-5p was significantly upregulated when cells were
transfected with miR-576-5p mimic (Fig.
3C). The results from RT-qPCR and western blotting confirmed
that the mRNA and protein expression levels of Wnt5a were regulated
by miR-576-5p, as its expression levels were downregulated in the
presence of the miR-576-5p mimic and upregulated following
miR-576-5p inhibition (Fig. 3D and
E). Wnt5a, one of the Wnt ligands, is known to act through the
β-catenin-independent non-canonical signaling pathway (17). The present study investigated
whether Wnt5a could activate or repress β-catenin signaling
following the differential expression of miR-576-5p in cells. The
results from western blotting demonstrated that miR-576-5p
overexpression significantly increased the expression of β-catenin,
c-Myc, cyclin D1 and phosphorylated (p)-c-Jun, whereas the
inhibition of miR-576-5p had the opposite effect (Fig. 3F). Therefore, these results
suggested that miR-576-5p may target Wnt5a and regulate the
β-catenin signaling pathway.
 | Figure 3.miR-576-5p targeted Wnt5a and
regulated β-catenin signaling pathway. (A) Putative binding site of
miR-576-5p and Wnt5a was predicted by TargetScan. (B)
Dual-luciferase report assay was conducted to confirm the predicted
binding site. (C) SW480 cells were transfected with miR-576-5p
mimic and miR-576-5p level was determined using RT-qPCR. (D and E)
miR-576-5p was upregulated or downregulated and mRNA level and
protein expression of Wnt5a were measured using RTq-PCR and western
blotting, respectively. (F) Protein expression of β-catenin, c-myc,
cyclin D1, p-c-Jun and c-Jun were determined using western
blotting. *P<0.05, **P<0.01 and ***P<0.001. miR, microRNA;
NC, negative control; CRC, colorectal cancer; RT-qPCR, reverse
transcription quantitative PCR; MUT, mutant; WT, wild-type; p,
phosphorylated. |
Wnt5a overexpression reverses the
effects of miR-576-5p on SW480 cells
The contribution of Wnt5a and β-catenin signaling in
miR-576-5p-mediated functions in SW480 cells was explored.
pcDNA-Wnt5a was transfected into SW480 cells to overexpress Wnt5a
(Fig. 4A) or cells were treated
with XAV-939 to inhibit the activation of Wnt/β-catenin signaling.
The cell viability and migratory and invasive abilities were
subsequently evaluated. The results demonstrated that upregulated
expression of miR-576-5p significantly improved SW480 cell
viability, which was partially reversed by Wnt5a overexpression or
treatment with XAV-939 (Fig. 4B).
The results also showed that the effects of upregulated miR-576-5p
on the cell migratory or invasive abilities were suppressed
following Wnt5a overexpression or XAV-939 treatment (Fig. 4C-F). These findings suggested that
miR-576-5p may exert its role on SW480 cell viability and migratory
and invasive abilities by regulating Wnt5a and β-catenin
signaling.
miR-576-5p regulates EMT by targeting
Wnt5a and activating Wnt/β-catenin signaling
The effects of Wnt5a and XAV-939 on
miR-576-5p-induced EMT were investigated. The results demonstrated
that miR-576-5p mimic significantly decreased the expression of
E-cadherin, and increased the expression of vimentin, N-cadherin
and Snail1 (Fig. 5), suggesting
that miR-576-5p may promote EMT in SW480 cells. miR-576-5p-mediated
EMT could then be reversed by either Wnt5a overexpression or
XAV-939 treatment, indicating that miR-576-5p mediated EMT in SW480
cells through Wnt5a and β-catenin signaling.
To confirm the association between miR-576-5p and
Wnt5a, the effect of miR-576-5p mimic in the presence or absence of
Wnt5a overexpression or XAV-939 treatment on Wnt/β-catenin
signaling was evaluated. The results demonstrated that miR-576-5p
significantly increased the expression of β-catenin, c-Myc, cyclin
D1 and p-c-Jun (Fig. 6). These
miR-576-5p-mediated changes were then reversed by either Wnt5a
overexpression or XAV-939 treatment, indicating that miR-576-5p
could mediate biological functions in SW480 cells via Wnt/β-catenin
signaling.
Discussion
The aberrant expression or dysfunction of miRNAs has
been reported to be closely associated with the progression of
numerous types of human cancers, by regulating the molecular
functions of cancer cells, including cell proliferation, metastatic
ability and drug resistance (18,19).
Increasing evidence has revealed that numerous miRNAs have a
prominent role in CRC, serving as either diagnostic markers or
therapeutic targets. For example, miR-92a can promote tumorigenesis
of CRC and has become a useful biomarker for early detection of CRC
in both serum and stool (20).
Reduced miR-4319 is correlated with poor prognosis in CRC patients,
and miR-4319 might become a therapeutic target for CRC treatment
(21). However, the precise role of
miR-576-5p in CRC remains unclear. The present study aimed
therefore to understand the potential role of miR-576-5p in CRC.
The results from the present study demonstrated that the expression
level of miR-576-5p was upregulated in CRC cancer cells compared
with normal colonic epithelial cells, which was consistent with a
previous study (14). A miR-576-5p
inhibitor was subsequently transfected into cancer cells to inhibit
miR-576-5p expression and changes in SW480 cells were thus
observed. The results demonstrated that miR-576-5p inhibition
significantly decreased SW480 cell viability and migratory and
invasive abilities. These results suggested that the inhibition of
miR-576-5p may exert an anti-cancer effect by weakening the
metastatic ability of CRC cells.
Distant metastases are major contributors to
cancer-associated mortality, which often lead to therapeutic
failure in patients (22). Cancer
cell metastasis involves a series of reactions regulated by various
factors, with EMT regarded as one of the most critical factors
responsible for the infiltration and metastasis of ~90% of
epithelial malignant tumors in humans (23). EMT consists in the differentiation
of epithelial cells into mesenchymal cells, which is characterized
by the loss of E-cadherin, a marker of epithelial characteristics,
and the increase in N-cadherin, a marker of mesenchymal
characteristics (24,25). In the present study, results from
western blotting demonstrated that miR-576-5p inhibition
significantly increased the expression of E-cadherin and decreased
the expression of N-cadherin, vimentin and Snail1, suggesting that
the inhibition of miR-576-5p may alleviate EMT in SW480 cells and
effectively inhibit cancer cell metastasis, providing a potential
therapeutic effect of miR-576-5p inhibition in CRC.
Subsequently, the present study investigated the
potential mechanism responsible for the suppressive effect of
miR-576-5p inhibition on cell viability, migratory and invasive
abilities, and EMT. Wnt5a was identified as the direct target of
miR-576-5p, which was regulated by miR-576-5p. Wnt5a, a member of
the Wnt signaling protein family, was found to regulate various
molecular functions through binding to different receptors on the
cellular surface (26).
Accumulating evidence has suggested that Wnt5a serves a crucial
role in the pathogenesis of numerous types of human cancers, where
Wnt5a exerts both oncogenic and tumor suppressive effects in
various types of cancer (27). For
instance, Wnt5a is highly expressed in patients with nasopharyngeal
cancer, especially those with poor survival (28). Furthermore, Wnt5a expression level
is positively correlated with the severity of melanoma as its
expression increases following the progression of melanoma,
suggesting that Wnt5a expression might be considered as a risk
factor for the outcome of the diseases (29). However, Wnt5a was identified to
serve as a tumor suppressor in hepatocellular carcinoma (HCC) and
breast cancer. Several downstream signaling pathways involved in
cancer cell metastasis were reported to be tightly controlled by
Wnt5a in HCC and breast cancer (30–32).
In CRC, Wnt5a is known to exert anti-cancer effects. For example,
Li and Chen (33) reported that
both mRNA and protein expression of Wnt5a was decreased in the
highly metastatic human colon cancer cells, and Cheng et al
(34) demonstrated that the
expression level of Wnt5a was significantly downregulated in most
patients with primary colon cancer. Restoring Wnt5a may therefore
be considered as an innovative therapeutic target. In the present
study, Wnt5a overexpression reversed miR-576-5p-induced EMT, cell
viability and migratory and invasive abilities, suggesting that
Wnt5a may be used to inhibit cell metastasis in SW480 cells.
Wnt5a, a member of the Wnt family of proteins,
activates the canonical or non-canonical Wnt signaling pathway
depending on the receptor context (35). As a key component of Wnt signaling,
β-catenin serves as the marker of the pathway activation. In
previous studies, Wnt5a was discovered to be an antagonist of the
β-catenin-dependent Wnt signaling pathway (26,33).
Furthermore, it was reported that Wnt/β-catenin signaling
participates in the regulation of tumor signaling transduction and
promotes the proliferation, differentiation and metastasis of tumor
cells (36). In addition,
Wnt/β-catenin signaling has been demonstrated to have a major
impact on EMT during cancer progression. Upon the absence of Wnt,
β-catenin and E-cadherin attach to the cell membrane (37,38).
The loss of E-cadherin activates the Wnt pathway, which facilitates
the translocation of β-catenin into the nucleus to initiate the
transcription of various downstream target genes, including cyclin
D1, c-Myc and c-Jun, which eventually promote the malignant
progression of cells (37,38). Furthermore, c-myc is an important
proto-oncogene associated with tumor occurrence and development.
The cancer gene c-myc possesses the function of molecular switch in
gene transcription and cell regulation, and is activated during the
transition from colorectal adenoma to adenocarcinoma, leading to
c-myc with abnormal activation, increased protein expression and
promotion of cell proliferation (39). Activation of β-catenin is the key
nuclear effector of Wnt pathway, and increased cytoplasmic and
nuclear translocation of β-catenin promotes the combination between
β-catenin and the T-cell factor (TCF)/lymphoid enhancer-binding
factor (LEF) transcription factor family (40). c-myc is a well-established target
gene of β-catenin/TCF transcription factor complex, and aberrant
nuclear accumulation of β-catenin and constitutive upregulation of
c-myc are believed to be the basis of colorectal tumorigenesis
(41,42). In the present study, Wnt/β-catenin
was activated following miR-576-5p overexpression. In addition, the
results from the present study revealed that the upregulation of
β-catenin, as well as of its downstream target genes c-Myc, cyclin
D1 and c-Jun, may account for the suppression of Wnt5a by
miR-576-5p, suggesting that miR-576-5p may activate Wnt/β-catenin
signaling by silencing Wnt5a. To further validate the regulatory
role of Wnt5a in CRC, Wnt5a was overexpressed in SW480 cells, and
cells were treated with XAV-939, an inhibitor of the Wnt/β-catenin
signaling pathway. The results demonstrated that both Wnt5a
overexpression and XAV-939 treatment exhibited similar inhibitory
effects on Wnt/β-catenin signaling. In addition, miR-576-5p
exhibited promotional effects on cell viability, and migratory and
invasive ability, in addition to EMT; however, the promotive
effects were partly inhibited by overexpression of Wnt5a or XAV-939
treatment, suggesting that Wnt5a may serve as an antagonist of
Wnt/β-catenin signaling pathway. Furthermore, the results suggested
that miR-576-5p may affect cell metastasis by targeting Wnt5a and
regulating Wnt5a-mediated Wnt/β-catenin signaling.
In conclusion, the present study revealed the
biological role of miR-576-5p in CRC, and the inhibition of
miR-576-5p was discovered to exert an anti-tumor effect by
inhibiting cell viability, migration, invasion and EMT. In
addition, miR-576-5p was demonstrated to directly target Wnt5a and
regulate Wnt5a-mediated Wnt/β-catenin signaling. Furthermore, the
effect of miR-576-5p in CRC cells may be reversed by Wnt5a
overexpression or inhibition of the Wnt/β-catenin signaling using
XAV-939. Taken together, these findings suggested that inhibition
of miR-576-5p may suppress CRC cell viability, migration, invasion
and EMT by targeting Wnt5a and regulating Wnt5a-mediated
Wnt/β-catenin signaling, suggesting that miR-576-5p may be
considered as a potential therapeutic target for CRC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the General
Program of Zhejiang Natural Science Foundation (grant no.
Y19H160049) and the Project of Medical and Health Science and
Technology Plan in Zhejing Province (grant no. 2020KY460).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
YZ and JL were responsible for conceiving and
designing the study. JL, LL, JS and NZho participated in the
literature collection and conducting the experiments. YF, NZha and
QS analyzed the data and created the figures. JL and LL drafted the
initial manuscript and YZ revised the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that there have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lu CW, ZhouDDXieTHaoJ L, Pant OP, Lu CB
and Liu XF: HOXAll antisense long noncoding RNA (HOXAll-AS): A
promising lncRNA in human cancers. Cancer Med. 7:3792–3799. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
McArdle CS and Hole DJ: Outcome following
surgery for colorectal cancer: Analysis by hospital after
adjustment for case-mix and deprivation. Br J Cancer. 86:331–335.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Holm M, Saraswat M, Joenväärä S, Ristimäki
A, Haglund C and Renkonen R: Colorectal cancer patients with
different C-reactive protein levels and 5-year survival times can
be differentiated with quantitative serum proteomics. PLoS One.
13:e01953542018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tsuchiya S, Okuno Y and Tsujimoto G:
MicroRNA: Biogenetic and functional mechanisms and involvements in
cell differentiation and cancer. J Pharmacol Sci. 101:267–270.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartels CL and Tsongalis GJ: MicroRNAs:
Novel biomarkers for human cancer. Clin Chem. 55:623–631. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun GL, Li Z, Wang WZ, Chen Z, Zhang L, Li
Q, Wei S, Li BW, Xu JH, Chen L, et al: miR-324-3p promotes gastric
cancer development by activating Smad4-mediated Wnt/beta-catenin
signaling pathway. J Gastroenterol. 53:725–739. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fan Z, Cui H, Xu X, Lin Z, Zhang X, Kang
L, Han B, Meng J, Yan Z, Yan X and Jiao S: MiR-125a suppresses
tumor growth, invasion and metastasis in cervical cancer by
targeting STAT3. Oncotarget. 6:25266–25280. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jiang C, Hu X, Alattar M and Zhao H: miRNA
expression profiles associated with diagnosis and prognosis in lung
cancer. Expert Rev Anticancer Ther. 14:453–461. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kordass T, Weber CEM, Eisel D, Pane AA,
Osen W and Eichmüller SB: miR-193b and miR-30c-1* inhibit, whereas
miR-576-5p enhances melanoma cell invasion in vitro. Oncotarget.
9:32507–32522. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ni XF, Zhao LH, Li G, Hou M, Su M, Zou CL
and Deng X: MicroRNA-548-3p and MicroRNA-576-5p enhance the
migration and invasion of esophageal squamous cell carcinoma cells
via NRIP1 down-regulation. Neoplasma. 65:881–887. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang L, Chen J, Wang L, Chen L, Du Z, Zhu
L, Cui M, Zhang M and Song L: Linc-PINT acted as a tumor suppressor
by sponging miR-543 and miR-576-5p in esophageal cancer. J Cell
Biochem. 120:19345–19357. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang YN, Chen ZH and Chen WC: Novel
circulating microRNAs expression profile in colon cancer: A pilot
study. Eur J Med Res. 22:512017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ghahhari NM and Babashah S: Interplay
between microRNAs and WNT/β-catenin signalling pathway regulates
epithelial-mesenchymal transition in cancer. Eur J Cancer.
51:1638–1649. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kumawat K and Gosens R: WNT-5A: Signaling
and functions in health and disease. Cell Mol Life Sci. 73:567–587.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Calin GA and Croce CM: MicroRNA-cancer
connection: The beginning of a new tale. Cancer Res. 66:7390–7394.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu W, Luo X, Fu H, Liu L, Sun P and Wang
Z: MiR-3653 inhibits the metastasis and epithelial-mesenchymal
transition of colon cancer by targeting Zeb2. Pathol Res Pract.
215:1525772019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen E, Li Q, Wang H, Yang F, Min L and
Yang J: MiR-92a promotes tumorigenesis of colorectal cancer, a
transcriptomic and functional based study. Biomed Pharmacother.
106:1370–1377. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang L, Zhang Y, Li Z, Zhao X, Xi Z, Chen
H, Shi H, Xin T, Shen R and Wang T: MiR-4319 suppresses colorectal
cancer progression by targeting ABTB1. United European
Gastroenterol J. 7:517–528. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rodrigo JP, Martínez P, Allonca E,
Alonso-Durán L, Suárez C, Astudillo A and García-Pedrero J:
Immunohistochemical markers of distant metastasis in laryngeal and
hypopharyngeal squamous cell carcinomas. Clin Exp Metastasis.
31:317–325. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hugo H, Ackland ML, Blick T, Lawrence MG,
Clements JA, Williams ED and Thompson EW: Epithelial-mesenchymal
and mesenchymal-epithelial transitions in carcinoma progression. J
Cell Physiol. 213:374–383. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li D, Tian B and Jin X: miR-630 inhibits
epithelial-to-mesenchymal transition (EMT) by regulating the
Wnt/β-catenin pathway in gastric cancer cells. Oncol Res. 27:9–17.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Montorsi L, Guizzetti F, Alecci C,
Caporali A, Martello A, Atene CG, Parenti S, Pizzini S, Zanovello
P, Bortoluzzi S, et al: Loss of ZFP36 expression in colorectal
cancer correlates to wnt/β-catenin activity and enhances
epithelial-to-mesenchymal transition through upregulation of ZEB1,
SOX9 and MACC1. Oncotarget. 7:59144–59157. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kikuchi A, Yamamoto H, Sato A and
Matsumoto S: Wnt5a: Its signalling, functions and implication in
diseases. Acta Physiol (Oxf). 204:17–33. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Asem MS, Buechler S, Wates RB, Miller DL
and Stack MS: Wnt5a Signaling in Cancer. Cancers (Basel). 8:792016.
View Article : Google Scholar
|
|
28
|
Qin L, Yin YT, Zheng FJ, Peng LX, Yang CF,
Bao YN, Liang YY, Li XJ, Xiang YQ, Sun R, et al: WNT5A promotes
stemness characteristics in nasopharyngeal carcinoma cells leading
to metastasis and tumorigenesis. Oncotarget. 6:10239–10252. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Da Forno PD, Pringle JH, Hutchinson P,
Osborn J, Huang Q, Potter L, Hancox RA, Fletcher A and Saldanha GS:
WNT5A expression increases during melanoma progression and
correlates with outcome. Clin Cancer Res. 14:5825–5832. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang T, Liu X and Wang J: Up-regulation of
Wnt5a inhibits proliferation and migration of hepatocellular
carcinoma cells. J Cancer Res Ther. 15:904–908. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bi L, Liu X, Wang C, Cao Y, Mao R, Li P
and Geng M: Wnt5a involved in regulation of the biological behavior
of hepatocellular carcinoma. Int J Clin Exp Pathol. 7:987–995.
2014.PubMed/NCBI
|
|
32
|
Prasad CP, Manchanda M, Mohapatra P and
Andersson T: WNT5A as a therapeutic target in breast cancer. Cancer
Metastasis Rev. 37:767–778. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li Q and Chen H: Silencing of Wnt5a during
colon cancer metastasis involves histone modifications.
Epigenetics. 7:551–558. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cheng R, Sun B, Liu Z, Zhao X, Qi L, Li Y
and Gu Q: Wnt5a suppresses colon cancer by inhibiting cell
proliferation and epithelial-mesenchymal transition. J Cell
Physiol. 229:1908–1917. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
van Amerongen R, Mikels A and Nusse R:
Alternative wnt signaling is initiated by distinct receptors. Sci
Signal. 1:re92008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Howard S, Deroo T, Fujita Y and Itasaki N:
A positive role of cadherin in Wnt/β-catenin signalling during
epithelial-mesenchymal transition. PLoS One. 6:e238992011.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wijnhoven BP, Dinjens WN and Pignatelli M:
E-cadherin-catenin cell-cell adhesion complex and human cancer. Br
J Surg. 87:992–1005. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
MacDonald BT, Tamai K and He X:
Wnt/beta-catenin signaling: Components, mechanisms, and diseases.
Dev Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shi L, Wu YX, Yu JH, Chen X, Luo XJ and
Yin YR: Research of the relationship between β-catenin and
c-myc-mediated Wnt pathway and laterally spreading tumors
occurrence. Eur Rev Med Pharmacol Sci. 21:252–257. 2017.PubMed/NCBI
|
|
40
|
Valenta T, Hausmann G and Basler K: The
many faces and functions of β-catenin. EMBO J. 31:2714–2736. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fang Y, Shen ZY, Zhan YZ, Feng XC, Chen
KL, Li YS, Deng HJ, Pan SM, Wu DH and Ding Y: CD36 inhibits
β-catenin/c-myc-mediated glycolysis through ubiquitination of GPC4
to repress colorectal tumorigenesis. Nat Commun. 10:39812019.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xie C, Pan Y, Hao F, Gao Y, Liu Z, Zhang
X, Xie L, Jiang G, Li Q and Wang E: C-Myc participates in
β-catenin-mediated drug resistance in A549/DDP lung adenocarcinoma
cells. APMIS. 122:1251–1258. 2014. View Article : Google Scholar : PubMed/NCBI
|