Introduction
Atrial fibrillation (AF) is one of the most common
forms of cardiac arrhythmia and is accompanied by a high risk of
stroke, heart failure and mortality (1). AF is also regarded as a significant
contributor towards morbidity and increased healthcare costs
(2). The mechanisms underlying AF
can be typically classified into three types: Autonomic neural
remodeling, electrical remodeling and structural remodeling
(3), which can act as triggers or
favor the creation of an AF-prone substrate. However, the
mechanisms of AF are intricate, which poses challenges for the
effective medical intervention of AF.
Recently, multiple molecular factors have been
reported to be involved in the pathophysiological progression of
AF, including fibrosis, abnormal Ca2+ handling and
inflammation (4–6). Previous studies have demonstrated
the significant role of microRNAs (miRNAs/miRs) in mediating these
molecular factors in AF (7).
miRNAs are a type of small non-coding RNA of 19–25 nucleotides in
length, and which regulate the expression of target genes at the
post-transcriptional level by promoting the degradation of target
mRNA or repressing target mRNA translation (8). Zhao et al (9) reported that miR-29a-3p serves roles
in the development of AF by downregulating the L-type
Ca2+ current (9).
Furthermore, Cañón et al (10) demonstrated that the aberrant
expression of miR-208b could reduce the expression and function of
L-type Ca2+ channel subunits, as well as the
sarcoplasmic reticulum Ca2+ pump, ATPase
sarcoplasmic/endoplasmic reticulum Ca2+ transporting 2,
in myocytes isolated from patients with chronic AF, suggesting that
miR-208b may be an important mediator in impaired Ca2+
handling during atrial remodeling. Our previous study demonstrated
that miR-27b-3p could regulate the Wnt/β-catenin signaling pathway
and attenuate atrial fibrosis in rats with AF by targeting Wnt3a
(11). miRNAs have therefore been
considered as promising targets for AF intervention.
Accumulating evidence has indicated that long
non-coding RNAs (lncRNAs), a class of non-coding RNAs of >200
nucleotides in length, may also serve an important role in cardiac
diseases by acting as competing endogenous RNAs (ceRNAs) (12). Salmena et al (13) first proposed the ceRNA theory in
2011, and hypothesized that mRNAs and lncRNAs could communicate
with each other by binding to shared miRNAs using miRNA response
elements (14). For example, the
lncRNA-long intergenic non-protein coding RNA 472 was discovered to
promote AF by downregulating the expression of ryanodine receptor 2
via miR-24 (15). However, the
underlying regulatory ceRNA mechanism in AF remains to be fully
elucidated.
The present study firstly aimed to identify
significantly differentially expressed miRNAs in AF by screening
microarray data from the Gene Expression Omnibus (GEO) database.
Then, right atrial appendage (RAA) tissues from patients with AF
were collected to verify the expression of the identified miRNAs.
Combined with the validation results, Gene Ontology (GO) functional
term and Kyoto Encyclopedia of Genes and Genomes (KEGG) signaling
pathway enrichment analyses were performed for the target genes of
the differentially expressed miRNAs of interest to fully understand
the role of these miRNAs. The potential upstream lncRNAs targeting
the identified miRNAs were predicted using Bioinformatics method.
Finally, a dual luciferase assay was used to verify whether a
targeted relationship existed between significantly differentially
expressed miRNAs and lncRNAs of interest. These findings may
provide a solid theoretical basis for future research to help
identifying potential targets for the treatment of AF.
Materials and methods
Microarray data collection
The present study used the GEO database (https://www.ncbi.nlm.nih.gov/geo), which is a
public gene expression database from the National Center for
Biotechnology Information. For microarray retrieval, ‘atrial
fibrillation’ was used as the key word to search for microarrays
that had studied the expression levels of miRNAs in AF. The
inclusion criteria were as follows: i) Human atrial appendage
tissue as the research sample type; ii) the Sinus rhythm (SR) group
was set as the control group and the persistent AF group was set as
the experimental group, and each group contained >3 research
samples; and iii) miRNA expression in atrial appendage tissues was
the main research focus. The exclusion criteria were as follows: i)
Incomplete microarray matrix data; ii) the source of the research
sample was unclear; iii) the main research focus of the microarray
matrix was not miRNAs; iv) the microarray annotation file was
missing or unknown; and v) the microarray data were unable to meet
the subsequent data analysis standards after standardized
processing. After retrieval, the platform files and microarray data
that met the criteria were downloaded for subsequent analysis.
Data preprocessing
To reduce the errors in data analysis, data
preprocessing of the downloaded microarray data was performed.
Firstly, the corresponding microarray platform files were used to
annotate the matrix data of the microarray and convert the miRNA
IDs. After annotation, the annotated matrix data were supplemented
using the impute toolkit of R software (version 3.5.1; RStudio,
Inc.). All matrix data that did not match the miRNAs were deleted.
For the matrix data with the same miRNA ID, the average value was
obtained after merging. Finally, Bioconductor's limma toolkit
(version 3.10.3; http://www.bioconductor.org/packages/2.9/bioc/html/limma.html)
in R software was used for background correction, provision of
missing values and data standardization.
Identification of differentially
expressed miRNAs
After data preprocessing, the limma toolkit in R
software was used to identify the significantly differentially
expressed miRNAs between AF and SR samples. The screening threshold
of significantly differentially expressed miRNAs was set as
|log2 fold change (FC)| ≥0.5 and P<0.05. The heatmap
and volcano map of significantly differentially expressed miRNAs
were drawn using R software. The top four most significantly
upregulated and downregulated miRNAs, according to the
|log2FC| values, were selected for verification using
reverse transcription-quantitative PCR (RT-qPCR).
Patient studies
We continuously included patients with SR and
persistent AF who were hospitalized at the First Affiliated
Hospital of Guangxi Medical University (Nanning, China) and were
undergoing open-heart surgery for coronary bypass grafting or valve
replacement. The patient's electrocardiogram was independently
judged by two cardiovascular physicians and combined with medical
history. The diagnostic criteria of AF was in line with the
diagnostic criteria of AF recommended in the 2016 European Society
of Cardiology AF management guidelines (16). Patients were excluded if they had
the additional following conditions: left ventricular ejection
fraction <50%, infectious diseases, active myocarditis,
endocarditis, active rheumatism, pulmonary disease,
hyperthyroidism, hematological diseases, tumors and autoimmune
diseases. Since September 2018, RAA tissues were collected at the
beginning of surgical intervention under extracorporeal circulation
and rapidly cryopreserved in liquid nitrogen, then stored at −80°C
until the RAA tissue of the last patient was collected in June
2019. All samples would then be used for RT-qPCR analysis. The
experimental protocol was approved by the Ethics Committee of The
First Affiliated Hospital of Guangxi Medical University (Nanning,
China). Each patient provided written informed consent prior to
surgery for RAA tissue collection.
RT-qPCR
Total RNA was extracted from the RAA tissues from
patients with AF or SR (n=8 per group) using TRIzol®
reagent (cat. no. 9108; Takara Bio, Inc.). A NanoDrop ND 1000
spectrophotometer was used to assess the absorbance of the
extracted total RNA at wavelengths of 260 and 280 nm. The 260/280
ratio was calculated to confirm the quality of the RNA, and only
samples with a 260/280 ratio between 1.8 and 2.1 were used. Total
RNA was reverse transcribed into cDNA using a tailing reaction kit
(cat. no. B532451; Sangon Biotech Co., Ltd.) for miRNA or a reverse
transcription kit (cat. no. RR047A; Takara Bio, Inc.) for mRNA
according to the manufacturers' instructions. When the universal
reverse primers provided by the tailing reaction kit could not
complete the cDNA synthesis of miRNA experiment, we replaced the
universal reverse primers or redesigned the reverse primers to
complete this part of the study. qPCR was subsequently performed on
a StepOne Real-Time PCR system (Thermo Fisher Scientific, Inc.).
The PCR procedure was as follows: One cycle at 95°C for 30 sec to
complete the initial denaturation, followed by 40 cycles of 95°C
for 5 sec and 60°C for 30 sec for denaturation, annealing and
extension. Melting curve analysis was performed at ~65–95°C.
Expression levels were quantified using the 2−∆∆Cq
method (17) and normalized to
GAPDH (for mRNA) or U6 (for miRNA). The sequences of the primers
used are listed in Table I.
 | Table I.Sequences of the primers used for
reverse transcription quantitative PCR. |
Table I.
Sequences of the primers used for
reverse transcription quantitative PCR.
| Target | Sequences
(5′-3′) |
|---|
| hsa-KCNQ1OT1 |
|
|
Forward |
GAACTCTGTTTTGTTTTCTGCTGC |
|
Reverse |
CATAAATCCTGAAACAGACCCACTT |
| hsa-GAS5 |
|
|
Forward |
AAGCCATTGGCACACAGGCATTAG |
|
Reverse |
AGAACCATTAAGCTGGTCCAGGCA |
| hsa-MINCR |
|
|
Forward |
CAGAAGAGCTTCATCGGCCC |
|
Reverse |
TCACAGACGCACTCTTCCCA |
| U6 |
|
|
Forward |
CTCGCTTCGGCAGCACA |
|
Reverse |
AACGCTTCACGAATTTGCGT |
| GAPDH |
|
|
Forward |
GGAAGCTTGTCATCAATGGAAATC |
|
Reverse |
TGATGACCCTTTTGGCTCCC |
| hsa-miR-196b |
|
|
Forward |
CGGGCTAGGTAGTTTCCTGT |
|
Reverse |
CAGCCACAAAAGAGCACAAT |
| hsa-miR-642b |
|
|
Forward |
AGACACAUUUGGAGAGGGACCC |
|
Reverse |
GAGUGUGCCCAAGAGAAAGUUU |
| hsa-miR-3164 |
|
|
Forward |
TGTGACTTTAAGGGAAATGGCG |
|
Reverse |
CTCTACAGCTATATTGCCAGCCAC |
| hsa-miR-223-3p |
|
|
Forward |
TGTCAGTTTGTCAAATACC |
|
Reverse |
AACTGGTGTCGTGAG |
| hsa-miR-425 |
|
|
Forward |
ATGACACGATCACTCCCGTTG |
|
Reverse |
GTGCAGGGTCCGAGGTATTC |
| hsa-miR-122 |
|
|
Forward |
GCGTGATGGAGTGTGACAAT |
|
Reverse |
GTGCAGGGTCCGAGGTATTC |
| hsa-miR-483-3p |
|
|
Forward |
GGTGTCACTCCTCTCCTCC |
|
Reverse |
CAGTGCGTGTCGTGGA |
| hsa-miR-33b |
|
|
Forward |
GTGCATTGCTGTTGCATTGC |
|
Reverse |
GTGCAGGGTCCGAGGT |
Bioinformatics analysis
By combining the data from the microarray analysis
and patient tissues, the differentially expressed miRNAs of
interest were selected for bioinformatics analysis to further
explore their functions. GO functional term and KEGG signaling
pathway enrichment analyses were conducted for the target genes of
the differentially expressed miRNAs of interest (18,19). The GO functional term enrichment
analysis consisted of biological process (BP), cellular component
(CC) and molecular function (MF) aspects. P<0.05 and a count
value ≥2 were used as the significant cutoff values. The results
from the GO functional term and KEGG signaling pathway enrichment
analyses were sorted using the Revigo toolkit in R software and
visualized in the form of bubble charts.
Prediction of lncRNAs targeting the
differentially expressed miRNAs of interest
The miRbase (http://www.mirbase.org) and StarBase (http://starbase.sysu.edu.cn/index.php)
databases were used to predict the upstream lncRNAs targeting the
differentially expressed miRNAs of interest. The identified lncRNAs
were inputted into Cytoscape software (version 3.7.2) to construct
the ceRNA network.
Dual luciferase reporter assay
The recombinant luciferase construct,
pcDNA3.1-KCNQ1OT1 3′-untranslated region (UTR)-wild-type (WT;
Hanbio Biotechnology Co., Ltd.) harboring the WT binding site for
miR-223-3p and another recombinant construct, pcDNA3.1-KCNQ1OT1
3′-UTR-mutant (MUT), containing the mutated binding site for
miR-223-3p, were constructed. Then, 293T cells (The Cell Bank of
Type Culture Collection of The Chinese Academy of Sciences) were
co-transfected with the pcDNA3.1-KCNQ1OT1 3′-UTR-WT (KCNQ1OT1 WT)
or pcDNA3.1-KCNQ1OT1 3′-UTR-MUT (KCNQ1OT1 MUT) recombinant plasmid
and miR-223-3p mimic (sense 5′-UGUCAGUUUGUCAAAUACCCC-3′, antisense
5′-GGGGUAUUUGACAAACUGACA-3′) or miR-223-3p mimic-negative control
(sense 5′-UUUGUACUACACAAAAGUACUG-3′, antisense
5′-CAGUACUUUUGUGUAGUACAAA-3′) by Liposomal Transfection Reagent
(Hanbio Biotechnology Co., Ltd.). The miR-223-3p mimic and
miR-223-3p mimic-negative control were constructed by GenePharma
Co., Ltd. After 48 h of transfection, the Dual-Luciferase Reporter
Assay system (cat. no. E1910; Promega Corporation) was used to
measure the relative luciferase activity in each group and the data
normalization was performed by comparing with the Renilla
luciferase activity of KCNQ1OT1 WT + miR-223-3p mimic group.
Statistical analysis
Statistical analysis was performed using SPSS
version 23 software (IBM Corp.). The categorical data are presented
as the frequency and the continuous data are presented as the means
± standard deviation. A χ2 test was used to determine
the statistical differences between categorical data. For
continuous and normally distributed data, statistical differences
between two groups were compared using a unpaired two-tailed
Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Microarray data
After screening, the miRNA microarray dataset
GSE68475, which contained 10 patients with AF and 11 patients with
SR, was selected for further analysis. The GSE68475 dataset, which
was uploaded by Morishima et al (20), aimed to detect the miRNAs
expression in human atrial appendages, and reported that miR-30d
was essential for the electrical remodeling of AF. After
preprocessing the miRNA microarray data according to the degree of
samples dispersion, two samples from both the AF and SR groups were
removed. Finally, eight samples from the AF group and nine samples
from the SR group were obtained.
Identification of differentially
expressed miRNAs in AF
miRNAs were screened for according to the threshold
cutoff values used in the present study. The results identified 43
differentially expressed miRNAs in the AF and SR groups, including
23 upregulated miRNAs, such as miR-33b, miR-483-3p, miR-122 and
miR-425 (Table II), and 20
downregulated miRNAs, such as miR-196b, miR-642b, miR-3164 and
miR-223-3p (Table III). The
heatmap and volcano map displaying the differentially expressed
miRNAs were drawn using R software and are presented in Figs. 1 and 2.
| Table II.miRNAs with increased expression in
patients with atrial fibrillation. |
Table II.
miRNAs with increased expression in
patients with atrial fibrillation.
| Target | |Log2FC| | t | P-value |
|---|
| hsa-miR-33b | 0.869586221 | 2.365376187 | 0.02954409 |
| hsa-miR-483-3p | 0.817867704 | 2.597063925 | 0.018293799 |
| hsa-miR-122 | 0.772442509 | 2.541789208 | 0.020534659 |
| hsa-miR-425 | 0.75856261 | 3.360950977 | 0.00351163 |
| hsa-miR-196a | 0.708341176 | 2.607334223 | 0.017903851 |
| hsa-miR-2276 | 0.660417901 | 2.909299897 | 0.009413676 |
| hsa-miR-493 | 0.649498855 | 3.344417162 | 0.00364197 |
| hsa-miR-4271 | 0.63060241 | 3.202575415 | 0.004974358 |
| hsa-miR-1539 | 0.622510073 | 2.536416632 | 0.020765806 |
| hsa-miR-4296 | 0.607087624 | 3.262255063 | 0.004363723 |
| hsa-miR-587 | 0.606329463 | 4.071871233 | 0.000726406 |
| hsa-miR-4298 | 0.602845228 | 2.412892937 | 0.026808013 |
| hsa-miR-548p | 0.574869493 | 4.30723264 | 0.000431555 |
| hsa-miR-605 | 0.550447818 | 2.738410535 | 0.013572845 |
| hsa-let-7i | 0.549892958 | 3.102281544 | 0.006194148 |
| hsa-miR-621 | 0.541751345 | 2.533866076 | 0.0208764 |
| hsa-miR-3125 | 0.538749883 | 3.107985712 | 0.006117547 |
| hsa-miR-3915 | 0.534540407 | 2.802455828 | 0.011840557 |
| hsa-miR-3180 | 0.528461253 | 2.632995349 | 0.016963765 |
| hsa-miR-492 | 0.52778329 | 3.813405954 | 0.001289044 |
| hsa-miR-193b | 0.506441031 | 2.255811103 | 0.036874622 |
| hsa-miR-641 | 0.505252296 | 2.308449626 | 0.033164494 |
| hsa-miR-127-5p | 0.500711212 | 4.461170028 | 0.000307439 |
| Table III.miRNAs with decreased expression in
patients with atrial fibrillation. |
Table III.
miRNAs with decreased expression in
patients with atrial fibrillation.
| Target | |Log2FC| | t | P-value |
|---|
| hsa-miR-196b | −0.953061347 | −2.387936306 | 0.028214045 |
| hsa-miR-642b | −0.684669912 | −5.232846332 | 5.78E-05 |
| hsa-miR-3164 | −0.667260096 | −3.14912112 | 0.005591877 |
| hsa-miR-223-3p | −0.667052957 | −3.778345029 | 0.001393388 |
| hsa-let-7e | −0.652894577 | −3.358265208 | 0.003532485 |
| hsa-miR-4264 | −0.643875178 | −3.129959531 | 0.005831009 |
| hsa-miR-301b | −0.613918625 | −3.965039772 | 0.000920629 |
| hsa-miR-4254 | −0.601866722 | −3.137886674 | 0.005730892 |
| hsa-miR-548o | −0.591841785 | −4.056385355 | 0.000751777 |
| hsa-miR-556-3p | −0.585672054 | −2.622710761 | 0.017334758 |
| hsa-miR-518c | −0.577279497 | −4.166962591 | 0.000588435 |
| hsa-miR-628-5p | −0.575507912 | −5.543327212 | 3.00E-05 |
| hsa-miR-518b | −0.57442567 | −2.137014059 | 0.046696381 |
| hsa-miR-130a | −0.570996268 | −2.417688503 | 0.026545462 |
|
hsa-miR-3678-3p | −0.566643877 | −2.618079507 | 0.017504327 |
| hsa-miR-578 | −0.556014824 | −2.409518058 | 0.026994239 |
| hsa-miR-889 | −0.544245666 | −6.228489463 | 7.35E-06 |
| hsa-miR-433 | −0.524268417 | −2.193339731 | 0.041773836 |
| hsa-miR-19a | −0.52202772 | −2.739368993 | 0.013545215 |
| hsa-miR-553 | −0.520224796 | −5.015733997 | 9.19E-05 |
Identification of differentially
expressed miRNAs in RAA tissues from patients with AF
To verify the expression of the aforementioned
differentially expressed miRNAs identified by screening the
microarray data, RT-qPCR was used to determine the expression
levels of the identified miRNAs in the RAA tissues from patients
with AF or SR. In total, 16 patients (age range, 42–69 years) were
used, including eight patients with persistent AF. The
clinicopathological characteristics of the included patients are
listed in Table IV, and no
significant differences were observed in the sex, age, medical
history or therapeutic regimen between the two groups.
| Table IV.Clinicopathological characteristics
of patients with atrial fibrillation. |
Table IV.
Clinicopathological characteristics
of patients with atrial fibrillation.
|
Characteristics | SR (n=8) | AF (n=8) | P-value |
|---|
| Sex ratio,
male/female | 5/3 | 7/1 | 0.119 |
| Age, years | 53.125±8.839 | 54.125±7.338 | 0.907 |
| Medical
history |
|
|
|
|
Coronary heart disease
(n) | 3 | 1 | 0.569 |
|
Diabetes mellitus (n) | 0 | 3 | 0.200 |
|
Hypertension (n) | 5 | 4 | 0.334 |
| Therapy |
|
|
|
|
CABG | 3 | 1 | 0.569 |
|
AVR | 0 | 2 | 0.467 |
|
MVR | 5 | 3 | 0.619 |
|
Combined valve
replacement | 0 | 2 | 0.467 |
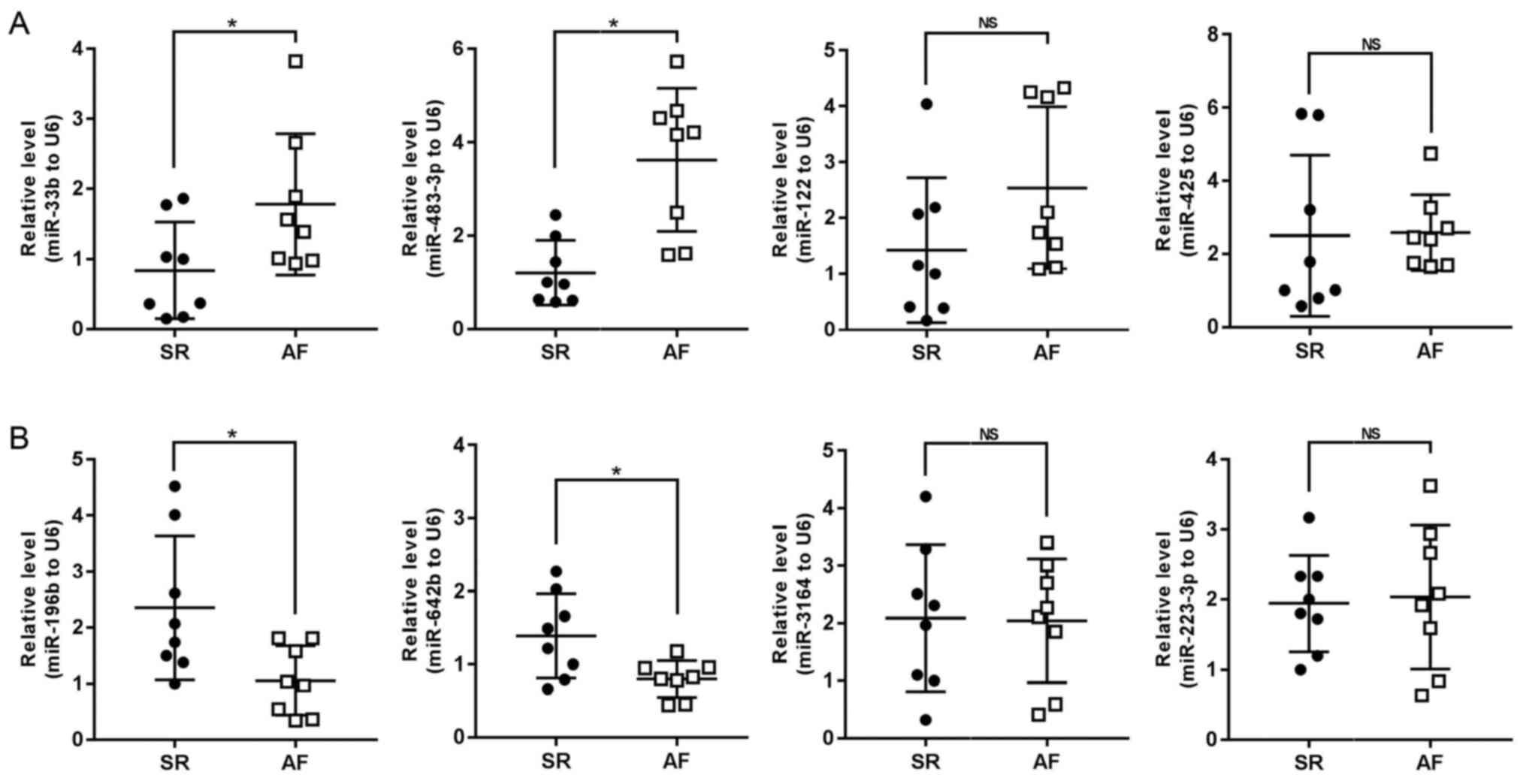
Compared with patients with SR, the expression
levels of miR-33b and miR-483-3p were found to be upregulated in
the AF group, while the expression levels of miR-196b and miR-642b
were downregulated. These findings were consistent with the
microarray data. However, no significant differences were
identified in the expression levels of miR-122, miR-425, miR-3164
and miR-223-3p between the two groups (Fig. 3). Our previous study reported that
miR-223-3p could mediate the generation of angiotensin
(Ang)II-induced reactive oxygen species and thereby regulate the
levels of oxidative stress (Weiran Dai; unpublished data).
Oxidative stress is one of the important mechanisms required for
the initiation and maintenance of AF. Zhang et al (21) reported that miR-223-3p regulates
the activation of paired box 6, resulting in apoptosis, a mechanism
that plays a significant role in the progression of the
prothrombotic state in AF. Novel evidence has suggested that the
inhibition of miR-223-3p expression could upregulate the expression
of FOXO3 and activate the autophagy pathway, thereby significantly
inhibiting myocardial fibrosis and improving myocardial remodeling
in AF (22). miR-223-3p may
therefore have multiple regulatory mechanisms in AF and
understanding the biological function and specific regulatory
mechanisms of miR-223-3p in AF may be of great significance. For
these reasons, miR-223-3p was selected for further analysis in the
present study.
Functional enrichment analyses for
target genes of miR-223-3p
According to the predictions of the bioinformatics
database, 415 target genes were identified as having a potential
regulatory relationship with miR-223-3p. To analyze the potential
biological functions of target genes of miR-223-3p, GO functional
terms and KEGG signaling pathways enriched by the target genes of
miR-223-3p were identified using the Database for Annotation,
Visualization and Integrated Discovery (version 6.8; http://david.ncifcrf.gov/). A total of 57 GO
functional terms and 21 KEGG signaling pathways were obtained. For
GO enrichment analysis, target genes of miR-223-3p were enriched in
20 BP, 17 CC and 20 MF, including ‘nuclear function’,
‘transcription factor activity regulation’, ‘RNA polymerase II
promoter’, ‘DNA template translation’ and ‘cytoplasmic composition’
(Fig. 4). The results from the
KEGG signaling pathway enrichment analysis revealed that target
genes of miR-223-3p were enriched in 21 KEGG signaling pathways,
including ‘AMPK signaling pathway’, ‘stem cell pluripotency
regulation’, ‘cGMP PKG signaling pathway’ and ‘insulin signaling
pathway’ (Fig. 5).
Identification of lncRNAs targeting
miR-223-3p in the RAA tissues from patients with AF
According to the bioinformatics database prediction,
69 lncRNAs targeting miR-223-3p were identified, including lncRNA
growth arrest specific transcript 5 (GAS5), lncRNA KCNQ1 opposite
strand/antisense transcript 1 (KCNQ1OT1) and lncRNA MYC-induced
long non-coding RNA (MINCR; Fig.
6A). As presented in Fig. 6B,
the results from RT-qPCR analysis revealed that, compared with SR
group, the expression level of KCNQ1OT1 was upregulated in RAA
tissues from patients with AF. Conversely, the expression level of
GAS5 was lower in patients with AF compared with that in patients
with SR (Fig. 6C). However, no
significant differences in the expression level of MINCR were
identified between the two groups (P>0.05; Fig. 6D). Furthermore, KCNQ1OT1 was
identified as an AF-related lncRNA using GeneCards (https://www.genecards.org). KCNQ1OT1 was therefore
selected for further analysis in the present study.
 | Figure 6.miR-223-3p is one direct target of
KCNQ1OT1. (A) Bioinformatics database predicted the lncRNAs
targeting miR-223-3p. (B-D) Expression of KCNQ1OT1, GAS5 and MINCR
in RAA of patients with SR or AF (n=8 per group); *P<0.05 vs. SR
group. (E) Binding site of KCNQ1OT1 and miR-223-3p. (F) Dual
luciferase assay confirmed that miR-223-3p was one direct target of
KCNQ1OT1 (n=3 per group). *P<0.05 vs. KCNQ1OT1 WT + miR-223-3p
NC group. SR, sinus rhythm; AF, atrial fibrillation; RAA, right
atrial appendage; NS, non-significant; WT, wild type; MUT, mutant;
miR, microRNA; NC, negative control; KCNQ1OT1, KCNQ1 opposite
strand/antisense transcript 1. |
miR-223-3p is a direct target of
KCNQ1OT1
To further investigate whether KCNQ1OT1 may directly
mediate miR-223-3p, a dual luciferase reporter assay was performed.
The results from the bioinformatics analysis suggested that
miR-223-3p may be considered as a potential target of KCNQ1OT1,
with two conserved putative binding sites identified between
miR-223-3p and KCNQ1OT1 (Fig.
6E). The results from the dual luciferase reporter assay
demonstrated that, compared with the KCNQ1OT1 WT + miR-223-3p
mimic-NC group, the relative luciferase activity of the KCNQ1OT1 WT
+ miR-223-3p mimic group was significantly decreased (1.0102±0.0234
vs. 0.4903±0.0316), while the decrease in relative luciferase
activity was reversed in the KCNQ1OT1 MUT + miR-223-3p mimic and
KCNQ1OT1 MUT + miR-223-3p mimic-NC groups (Fig. 6F). Taken together, these results
suggested that miR-223-3p may be a direct target of KCNQ1OT1, and
that a ceRNA regulatory relationship may exist between KCNQ1OT1 and
miR-223-3p in AF. These findings provided a novel potential target
for the treatment of AF and might initiate further
investigations.
Discussion
Despite the significant progress that has been made
in the treatment of AF, AF continues to negatively impact
individuals' lives, and the incidence increases with age. A recent
study suggested that lncRNAs function as ceRNAs and could regulate
miRNA expression and thereby affect the expression of target genes
of miRNAs to contribute to the pathogenesis of AF (23). Therefore, screening for
differentially expressed miRNAs using bioinformatics analysis
remains an effective method to identify potential targets for
future research into AF intervention. To the best of our knowledge,
the present study was the first to identify and verify the specific
regulatory relationship between KCNQ1OT1 and miR-223-3p using
microarray data and may provide a promising target for future AF
intervention experiments.
In cardiology, miR-223-3p has been shown to exert
multiple regulatory effects. Previous studies have reported that
miR-223-3p can regulate cell proliferation, apoptosis and necrosis
at the post-transcriptional level via targeted inhibition
mechanisms, and can also exert anti-inflammatory effects (24–26). A previous study reported that
miR-223-3p could directly inhibit IκB kinase α to regulate the
inflammatory response and necrosis induced by myocardial
ischemia/reperfusion injury (27). Furthermore, another study
suggested that circulating miR-223-3p levels might be used as a
circulating marker of unstable plaques in coronary atherosclerotic
heart disease (28). To fully
understand the biological function of miR-223-3p, the present study
performed GO functional term and KEGG signaling pathway enrichment
analyses for potential miR-223-3p target genes. The results from
the GO functional term enrichment analysis revealed that target
genes were mainly enriched in ‘nuclear function’, ‘transcription
factor activity regulation’, ‘RNA polymerase II promoter’, ‘DNA
template translation’ and ‘cytoplasmic composition’. The results
from the KEGG signaling pathway enrichment analysis demonstrated
that target genes were mainly enriched in the ‘AMPK signaling
pathway’, ‘stem cell pluripotency regulation’, ‘cGMP PKG signaling
pathway’ and ‘insulin signaling pathway’.
The AMPK signaling pathway is an important energy
metabolism pathway, which plays an essential role in the regulation
of cardiomyocyte energy homeostasis. AMPK regulates the metabolism
of fatty acids and glycogen to maintain energy balance within the
cell. Previous evidence also revealed that AMPK could regulate
protein synthesis and cell proliferation via the eukaryotic
translation elongation factor 2 and TSC complex subunit 2/mTOR
signaling pathways and regulate blood flow via endothelial or
neuronal nitric oxide synthase (29). In addition, the AMPK signaling
pathway is also considered as a cardiac metabolite regulator. AMPK
is a sensitive sensor of cell stress, which can be activated by
phosphorylation of the upstream signal at position 172 threonine,
and then exerts a protective effect by preventing metabolic stress,
cell proliferation and hypertrophic remodeling, and regulating
membrane electrophysiology. In addition, a previous study
demonstrated that AMPK can also regulates the expression levels of
Ca2+ channels and ion transporters to mediate
intracellular Ca2+ signaling processing and transmission
(30). In addition, abnormal
Ca2+ handling may affect the action potential duration,
which is an important mechanism of AF induction. Harada et
al (31) report that
downregulation of AMPK inhibits the abnormal Ca2+
transmission mediated by the integrity of atrial L-type
Ca2+ channels, suggesting that the AMPK signaling
pathway might play an important role in maintaining cardiac ion
homeostasis and arrhythmias, including AF. Amongst the
susceptibility models of AF, collagen-induced arthritis (CIA) is
associated with the increased expression of inflammatory cytokines
(interleukin-6 and tumor necrosis factor-α), and it has been shown
to lead to atrial remodeling and AF. Zhang et al (32) used a CIA rat model and
demonstrated that CIA could induce atrial energy metabolism
disorder by inhibiting the AMPK/PPARG coactivator 1 α signaling
pathway, while resveratrol could effectively reverse this
inhibitory effect and reduce the susceptibility of AF in CIA model
rats, suggesting that the increased risk of AF may be associated
with the downregulation of the AMPK signaling pathway (33,34). Another study demonstrated that
fisetin can promote AMPK phosphorylation in the atrium, reducing AF
susceptibility following myocardial infarction (35). The AMPK signaling pathway may
therefore represent a potential research direction for future
studies into AF intervention.
lncRNAs are important upstream regulators of the
miRNA-associated ceRNA network by sponging miRNAs with related
binding sites. In the present study, bioinformatics analysis
predicted that 69 lncRNAs could target miR-223-3p, including GAS5,
KCNQ1OT1 and MINCR. The expression of these lncRNAs was
subsequently verified in RAA tissues from patients with AF, and the
results demonstrated that compared with patients with SR, KCNQ1OT1
was upregulated and GAS5 was downregulated.
KCNQ1OT1 is located in the 15.5 region of the KCNQ1
locus on chromosome 11 (36).
KCNQ1OT1 has been reported to be associated with numerous types of
disease, including cardiovascular diseases (37). Li et al (38) demonstrated that KCNQ1OT1 could
prevent myocardial ischemia/reperfusion injury following acute
myocardial infarction by regulating adiponectin receptor 1.
Furthermore, KCNQ1OT1 was identified as an AF-related lncRNA using
the GeneCard database. Consistent with the results from the present
study, Shen et al (39)
reported that KCNQ1OT1 expression is significantly upregulated in a
time-dependent manner in an angiotensin II (AngII) infusion mouse
model. AngII treatment has been demonstrated to effectively
increase AF susceptibility. AngII is therefore widely used in
scientific research to establish an AF susceptibility model. In
addition, KCNQ1OT1 was shown to be an essential lncRNA involved in
the regulation of a cardiovascular disease ceRNA network, and it
could function as a molecular sponge by binding to miRNAs to
regulate the expression of their downstream target genes. Yang
et al (40) report that
KCNQ1OT1 is highly expressed in patients with diabetic
cardiomyopathy and that silencing KCNQ1OT1 successfully inhibits
cell apoptosis by increasing the expression of miR-214-3p and then
downregulating its target gene by acting as a ceRNA in vivo.
Furthermore, a previous study indicated that KCNQ1OT1 might be
involved in the occurrence of AF as a ceRNA by mediating the
miR-384b/Ca2+ voltage-gated channel subunit α 1C axis
(39). In the present study, the
results from the dual luciferase reporter assay revealed that
miR-223-3p could directly bind to KCNQ1OT1, suggesting that
KCNQ1OT1 may regulate the expression of downstream genes targeted
by miR-223-3p to exert its biological molecular effects. The
findings from the present study may therefore provide a new
promising target for AF intervention.
The lncRNA GAS5 plays an important role in a variety
of cardiovascular diseases. For example, in hypertension, GAS5 has
been considered as a new vascular remodeling regulator (41). In AF, GAS5 can reverse the
proliferation of cardiac fibroblasts by inhibiting transforming
growth factor β receptor 1, thus decreasing the synthesis of
collagen fibers in the extracellular matrix and inhibiting the
structural remodeling of the myocardium and the progression of AF
(42). In addition, GAS5 has also
been reported to serve as a ceRNA in cardiovascular diseases and
inhibit the negative regulation of miRNAs on downstream gene
expression. Zhou et al (43) demonstrated that GAS5, by acting as
a ceRNA, can inhibit miR-21 via its molecular sponge action, thus
regulating the expression of programmed cell death 4, a direct
target of miR-21, and mediating cell apoptosis in myocardial
infarction. Increasing evidence has suggested that GAS5 might also
act as a ceRNA and inhibit miR-223-3p, thereby regulating the
expression of its downstream target genes (44). Similarly, Yao et al
(45) reported that miR-223-3p
could directly bind to the 3′-UTR of nicotinamide
phosphoribosyltransferase (NAMPT) and that GAS5 could sponge
miR-223-3p and relieve the inhibitory effect of miR-223-3p on NAMPT
via regulating PI3K/AKT signaling. As there have been numerous
studies demonstrating that GAS5 can directly target miR-223-3p
(44,45), the present study did not perform
dual luciferase reporter assays to verify whether miR-223-3p was a
target of GAS5. However, the underlying mechanism of GAS5 in AF
based on the ceRNA theory has not been studied in detail, to the
best of our knowledge, and will require further experimental
verification.
The results from the present study provided novel
promising targets for further experimentation to develop strategies
for AF intervention. However, there were some limitations to the
present study. Firstly, human samples were used to verify the
differentially expressed miRNAs of interest found within the
microarray dataset, in addition to the trends in the expression
levels of upstream lncRNAs in AF. Unfortunately, the study did not
perform in vitro transfection experiments to further verify
the direct targeted regulatory relationship between the identified
miRNAs and lncRNAs. Hence, the lack of functional in vitro
experiments was a limitation to this study. The author's research
group is currently performing in vivo and in vitro
experiments to validate the results from the present study.
Secondly, the sample size used in this study was relatively small,
which may produce selection offset. Some evidence has suggested
that miR-223-3p expression in AF is controversial. For example,
Wang et al (46) studied
the miRNA transcriptome in AF using a tissue microarray and
demonstrated that miR-223-3p expression level is upregulated in AF
tissues. Conversely, a recent study using exosomes isolated from
patients with SR and AF reported that compared patient with SR,
miR-223-3p expression levels were downregulated in circulation
expsomes of patients with AF (47). Thus, when the sample size used is
relatively small and the reported expression of the gene is
inconsistent with the literature, the selection offset should be
expanded. Subsequently, further verification using larger sample
sizes and multi-center research investigations is required.
In summary, the present study used microarray data
on AF to identify significantly differently expressed miRNAs, and
the results were subsequently verified in RAA tissues from patients
with AF. Amongst the 43 significantly differently expressed miRNAs
identified, miR-223-3p was discovered to be significantly
upregulated in AF. Through GO functional term and KEGG signaling
pathway enrichment analyses of the target genes of miR-223-3p, the
‘AMPK signaling pathway’ was identified as an important pathway. In
addition, lncRNAs, including KCNQ1OT1, GAS5 and MINCR, were
identified as targeting miR-223-3p. Furthermore, the results from
dual luciferase reporter assay confirmed the existence of a
targeted relationship between KCNQ1OT1 and miR-223-3p. These
findings suggested that KCNQ1OT1 and GAS5 may serve crucial roles
as ceRNAs in AF. These findings provided a solid theoretical basis
for future intervention experiments and offered some potential
novel targets for AF intervention. However, further transfection
experiments both in vivo and in vitro should be
performed to verify the findings from this study.
Acknowledgements
Not applicable.
Funding
This study was supported by the Natural Science
Foundation of China (grant nos. 82060068 and 81760060).
Availability of data and materials
The GSE68475 dataset is available from the Gene
Expression Omnibus website (http://www.ncbi.nlm.nih.gov/geo/).
Authors' contributions
WD and XC performed the experiments. WD wrote the
manuscript. GZ and ZJ designed the experiments. ZJ carefully
revised the important intellectual content of the manuscript. WD
and GZ confirm the authenticity of all the raw data. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental protocol was approved by the Ethics
Committee of The First Affiliated Hospital of Guangxi Medical
University (Nanning, China). Each patient provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chugh SS, Havmoeller R, Narayanan K, Singh
D, Rienstra M, Benjamin EJ, Gillum RF, Kim YH, McAnulty JH Jr,
Zheng ZJ, et al: Worldwide epidemiology of atrial fibrillation: A
Global Burden of Disease 2010 Study. Circulation. 129:837–847.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zimetbaum P: Atrial Fibrillation. Ann
Intern Med. 166:ITC33–ITC48. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nattel S and Harada M: Atrial remodeling
and atrial fibrillation: Recent advances and translational
perspectives. J Am Coll Cardiol. 63:2335–2345. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ferrari R, Bertini M, Blomstrom-Lundqvist
C, Dobrev D, Kirchhof P, Pappone C, Ravens U, Tamargo J, Tavazzi L
and Vicedomini GG: An update on atrial fibrillation in 2014: From
pathophysiology to treatment. Int J Cardiol. 203:22–29. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu Y, Shi Q, Ma Y and Liu Q: The role of
immune cells in atrial fibrillation. J Mol Cell Cardiol.
123:198–208. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vyas V, Hunter RJ, Longhi MP and Finlay
MC: Inflammation and adiposity: New frontiers in atrial
fibrillation. Europace. 22:1609–1618. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Arroyo AB, de Los Reyes-García AM,
Rivera-Caravaca JM, Valledor P, García-Barberá N, Roldán V, Vicente
V, Martínez C and González-Conejero R: miR-146a regulates
neutrophil extracellular trap formation that predicts adverse
cardiovascular events in patients with atrial fibrillation.
Arterioscler Thromb Vasc Biol. 38:892–902. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Farazi TA, Hoell JI, Morozov P and Tuschl
T: MicroRNAs in human cancer. Adv Exp Med Biol. 774:1–20. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao Y, Yuan Y and Qiu C: Underexpression
of CACNA1C caused by overexpression of microRNA-29a underlies the
pathogenesis of atrial fibrillation. Med Sci Monit. 22:2175–2181.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cañón S, Caballero R, Herraiz-Martínez A,
Pérez-Hernández M, López B, Atienza F, Jalife J, Hove-Madsen L,
Delpón E and Bernad A: miR-208b upregulation interferes with
calcium handling in HL-1 atrial myocytes: Implications in human
chronic atrial fibrillation. J Mol Cell Cardiol. 99:162–173. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lv X, Li J, Hu Y, Wang S, Yang C, Li C and
Zhong G: Overexpression of miR-27b-3p targeting Wnt3a regulates the
signaling pathway of Wnt/β-catenin and attenuates atrial fibrosis
in rats with atrial fibrillation. Oxid Med Cell Longev.
2019:57037642019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kong F, Jin J, Lv X, Han Y, Liang X, Gao Y
and Duan X: Long noncoding RNA RMRP upregulation aggravates
myocardial ischemia-reperfusion injury by sponging miR-206 to
target ATG3 expression. Biomed Pharmacother. 109:716–725. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: the Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lou W, Ding B and Fu P: Pseudogene-derived
lncRNAs and their miRNA sponging mechanism in human cancer. Front
Cell Dev Biol. 8:852020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang LY, Shen H, Yang Q, Min J, Wang Q, Xi
W, Yin L, Le SG, Zhang YF, Xiao J, et al: LncRNA-LINC00472
contributes to the pathogenesis of atrial fibrillation (Af) by
reducing expression of JP2 and RyR2 via miR-24. Biomed
Pharmacother. 120:1093642019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kirchhof P, Benussi S, Kotecha D, Ahlsson
A, Atar D, Casadei B, Castella M, Diener HC, Heidbuchel H, Hendriks
J, et al ESC Scientific Document Group, : 2016 ESC Guidelines for
the management of atrial fibrillation developed in collaboration
with EACTS. Eur Heart J. 37:2893–2962. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gene Ontology Consortium, . The Gene
Ontology (GO) project in 2006. Nucleic Acids Res. 34:D322–D326.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kanehisa M, Sato Y, Kawashima M, Furumichi
M and Tanabe M: KEGG as a reference resource for gene and protein
annotation. Nucleic Acids Res. 44(D1): D457–D462. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Morishima M, Iwata E, Nakada C, Tsukamoto
Y, Takanari H, Miyamoto S, Moriyama M and Ono K: Atrial
Fibrillation-Mediated Upregulation of miR-30d Regulates Myocardial
Electrical Remodeling of the G-Protein-Gated K(+) Channel, IK.ACh.
Circ J. 80:1346–1355. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang H, Yang G, Zhong N, Shan J, Li X, Wu
Y, Xu Y and Yuan Y: Possible key microRNAs and corresponding
molecular mechanisms for atrial fibrillation. Anatol J Cardiol.
23:324–333. 2020.PubMed/NCBI
|
|
22
|
Hu J, Wang X, Cui X, Kuang W, Li D and
Wang J: Quercetin prevents isoprenaline-induced myocardial fibrosis
by promoting autophagy via regulating miR-223-3p/FOXO3. Cell Cycle.
20:1253–1269. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Z, Wang X, Wang W, Du J, Wei J, Zhang
Y, Wang J and Hou Y: Altered long non-coding RNA expression profile
in rabbit atria with atrial fibrillation: TCONS_00075467 modulates
atrial electrical remodeling by sponging miR-328 to regulate
CACNA1C. J Mol Cell Cardiol. 108:73–85. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ji Q, Xu X, Song Q, Xu Y, Tai Y, Goodman
SB, Bi W, Xu M, Jiao S, Maloney WJ, et al: miR-223-3p inhibits
human osteosarcoma metastasis and progression by directly targeting
CDH6. Mol Ther. 26:1299–1312. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Matsuzaki J and Ochiya T: Extracellular
microRNAs and oxidative stress in liver injury: A systematic mini
review. J Clin Biochem Nutr. 63:6–11. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Boxberger N, Hecker M and Zettl UK:
Dysregulation of Inflammasome Priming and Activation by MicroRNAs
in Human Immune-Mediated Diseases. J Immunol. 202:2177–2187. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qin D, Wang X, Li Y, Yang L, Wang R, Peng
J, Essandoh K, Mu X, Peng T, Han Q, et al: MicroRNA-223-5p and −3p
Cooperatively Suppress Necroptosis in Ischemic/Reperfused Hearts. J
Biol Chem. 291:20247–20259. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Singh S, de Ronde MWJ, Kok MGM, Beijk MA,
De Winter RJ, van der Wal AC, Sondermeijer BM, Meijers JCM,
Creemers EE and Pinto-Sietsma SJ: MiR-223-3p and miR-122-5p as
circulating biomarkers for plaque instability. Open Heart.
7:e0012232020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hardie DG: The AMP-activated protein
kinase pathway--new players upstream and downstream. J Cell Sci.
117:5479–5487. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chakraborty P, Nattel S and Nanthakumar K:
Linking cellular energy state to atrial fibrillation pathogenesis:
Potential role of adenosine monophosphate-activated protein kinase.
Heart Rhythm. 17:1398–1404. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Harada M, Tadevosyan A, Qi X, Xiao J, Liu
T, Voigt N, Karck M, Kamler M, Kodama I, Murohara T, et al: Atrial
Fibrillation Activates AMP-Dependent Protein Kinase and its
Regulation of Cellular Calcium Handling: Potential Role in
Metabolic Adaptation and Prevention of Progression. J Am Coll
Cardiol. 66:47–58. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Y, Zhang S, Liu Z, Zhao X, Yuan Y,
Sheng L and Li Y: Correction to: Resveratrol prevents atrial
fibrillation by inhibiting atrial structural and metabolic
remodeling in collagen-induced arthritis rats. Naunyn Schmiedebergs
Arch Pharmacol. 393:927–928. 2020.Erratum for: Naunyn Schmiedebergs
Arch Pharmacol 391: 1179-1190, 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Abe I, Teshima Y, Kondo H, Kaku H, Kira S,
Ikebe Y, Saito S, Fukui A, Shinohara T, Yufu K, et al: Association
of fibrotic remodeling and cytokines/chemokines content in
epicardial adipose tissue with atrial myocardial fibrosis in
patients with atrial fibrillation. Heart Rhythm. 15:1717–1727.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Maesen B, Nijs J, Maessen J, Allessie M
and Schotten U: Post-operative atrial fibrillation: A maze of
mechanisms. Europace. 14:159–174. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu L, Gan S, Li B, Ge X, Yu H and Zhou H:
Fisetin Alleviates Atrial Inflammation, Remodeling, and
Vulnerability to Atrial Fibrillation after Myocardial Infarction.
Int Heart J. 60:1398–1406. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kanduri C: Kcnq1ot1: A chromatin
regulatory RNA. Semin Cell Dev Biol. 22:343–350. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang Y, Du W, Chu Q, Qin Y, Tuguzbaeva G,
Wang H, Li A, Li G, Li Y, Chai L, et al: Downregulation of Long
Non-Coding RNA Kcnq1ot1: An Important Mechanism of Arsenic
Trioxide-Induced Long QT Syndrome. Cell Physiol Biochem.
45:192–202. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li X, Dai Y, Yan S, Shi Y, Han B, Li J,
Cha L and Mu J: Down-regulation of lncRNA KCNQ1OT1 protects against
myocardial ischemia/reperfusion injury following acute myocardial
infarction. Biochem Biophys Res Commun. 491:1026–1033. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shen C, Kong B, Liu Y, Xiong L, Shuai W,
Wang G, Quan D and Huang H: YY1-induced upregulation of lncRNA
KCNQ1OT1 regulates angiotensin II-induced atrial fibrillation by
modulating miR-384b/CACNA1C axis. Biochem Biophys Res Commun.
505:134–140. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang F, Qin Y, Wang Y, Li A, Lv J, Sun X,
Che H, Han T, Meng S, Bai Y, et al: LncRNA KCNQ1OT1 Mediates
Pyroptosis in Diabetic Cardiomyopathy. Cell Physiol Biochem.
50:1230–1244. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang YN, Shan K, Yao MD, Yao J, Wang JJ,
Li X, Liu B, Zhang YY, Ji Y, Jiang Q, et al: Long Noncoding
RNA-GAS5: A Novel Regulator of Hypertension-Induced Vascular
Remodeling. Hypertension. 68:736–748. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lu J, Xu FQ, Guo JJ, Lin PL, Meng Z, Hu
LG, Li J, Li D, Lu XH and An Y: Long noncoding RNA GAS5 attenuates
cardiac fibroblast proliferation in atrial fibrillation via
repressing ALK5. Eur Rev Med Pharmacol Sci. 23:7605–7610.
2019.PubMed/NCBI
|
|
43
|
Zhou XH, Chai HX, Bai M and Zhang Z:
LncRNA-GAS5 regulates PDCD4 expression and mediates myocardial
infarction-induced cardiomyocytes apoptosis via targeting miR-21.
Cell Cycle. 19:1363–1377. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dong X, Kong C, Liu X, Bi J, Li Z, Li Z,
Zhu Y and Zhang Z: GAS5 functions as a ceRNA to regulate hZIP1
expression by sponging miR-223 in clear cell renal cell carcinoma.
Am J Cancer Res. 8:1414–1426. 2018.PubMed/NCBI
|
|
45
|
Yao J, Shi Z, Ma X, Xu D and Ming G:
lncRNA GAS5/miR-223/NAMPT axis modulates the cell proliferation and
senescence of endothelial progenitor cells through PI3K/AKT
signaling. J Cell Biochem. 120:14518–14530. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang JG, Meng X, Han J, Li Y, Luo TG, Wang
J, Xin M and Xi JZ: Differential expressions of miRNAs in patients
with nonvalvular atrial fibrillation. Zhonghua Yi Xue Za Zhi.
92:1816–1819. 2012.(In Chinese). PubMed/NCBI
|
|
47
|
Wang S, Min J, Yu Y, Yin L, Wang Q, Shen
H, Yang J, Zhang P, Xiao J and Wang Z: Differentially expressed
miRNAs in circulating exosomes between atrial fibrillation and
sinus rhythm. J Thorac Dis. 11:4337–4348. 2019. View Article : Google Scholar : PubMed/NCBI
|