Introduction
Particulate matter (PM), also called particle
pollution, is the suspension of liquid or solid particles in the
air and includes inorganic and organic particles such as smoke,
dust, pollen, biological contaminants, heavy metals and dirt
(1–3). PM can be classified by the diameter
of the particles, including PM10, PM2.5 and
PM1.0, and size is an important factor affecting the
cellular response (3–5). Exposure to PM is linked to heart
disease (cardiopulmonary disease, cardiac arrest and
cardiovascular/hypertensive disease), lung disease (pneumonia, lung
cancer, asthma and chronic obstructive lung disease) and
neurodegeneration (1,6). The treatment of cells with PM is
associated with both an inflammatory response and a pro-oxidative
response (7). For example, PM
(PM10 and PM2.5) treatments induce Akt
activation via phosphorylation at Ser473 and Thr308 in human lung
epithelial cells (BEAS-2B) (8).
Additionally, coal and silica dust treatments induce alterations in
DNA methylation patterns via changes in DNA methyltransferase
(DNMT) 1, DNMT3α, DNMT3β, methyl CpG binding protein 2 and
methyl-CpG binding domain protein 2 expression (9,10).
In addition, the treatment of A549 cells with PM2.5
results in cytotoxicity and genotoxicity (3).
PM stimulates the immune system in the lungs and
respiratory tract. Following inhalation, PM accumulates in air
exchange regions of the lung, including the alveoli and stimulates
defense mechanisms involving alveolar epithelial cells or
macrophages (11). Lung tissue
consists of various cell types, such as epithelium and fibroblasts,
that maintain homeostasis and defense mechanisms in the lungs
(12). Fibroblasts are
responsible for maintaining the alveolar structure during repair
and proliferation in injured regions (13). The lung epithelium provides an
inert barrier for gas exchange and bulk airflow (14). Although PM exposure occurs in lung
epithelial cells, the cell-specific responses of lung epithelial
cells and fibroblasts to PM remain to be elucidated.
Thus, the present study performed an RNA-sequencing
(RNA-seq) analysis on lung epithelial cells (WI-38 VA-13) and
fibroblast cells (WI-38) following treatment with PM10
and used Gene Ontology (GO) analysis programs [Clustering GO
(ClueGO) and Database for Annotation, Visualization and Integrated
Discovery (DAVID)] to identify the cell-specific responses to
PM.
Materials and methods
PM10 sampling and
components
The fine dust ERM®-CZ100
(PM10) was purchased from Sigma-Aldrich (Merck KGaA).
PM10 packaged in amber glass vials was used to prepare
stock solutions in phosphate-buffered saline (PBS; 100 mg/ml) and
experiments using the suspended PM10 composition were
performed immediately. The PM10 used contained the
following polyaromatic hydrocarbons (PAHs): i) Benzo(a)anthracene;
ii) benzo(a)pyrene; iii) benzo(b)fluoranthene; iv)
benzo(l)fluoranthene; v) benzo(k)fluoranthene; vi)
dibenz(a,h)anthracene; vii) indeno(1,2,3-cd)pyrene; and viii) a
combination of benzo(b)fluoranthene, benzo(k)fluoranthene and
benzo(l)fluoranthene.
Cell culture and treatment
The normal human lung cell lines WI-38 and WI-38
VA-13 subclone 2RA (WI-38 VA-13) were purchased from the Korean
Cell Line Bank (Korean Cell Line Research Foundation) and cultured
in MEM (HyClone; Cytiva) and RPMI-1640 medium (Welgene, Inc.),
respectively, supplemented with 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.) and 1% penicillin/streptomycin
(Gibco; Thermo Fisher Scientific, Inc.) in a humidified atmosphere
under 5% CO2 at 37°C. The WI-38 and WI-38 VA-13 cell
lines were treated with PM10 (9 µg/cm2) for
48 h.
Cell viability assay
A Cell Counting Kit-8 (CCK-8; Dojindo Laboratories,
Inc.) was used to conduct the cell viability assays (15). WI38 and WI38 VA-13 cells were
seeded in 6-well plates at 4×105 cells/well and
incubated for 24 h. Following PM10 treatment for 48 h,
CCK-8 solution and RPMI-1640 medium with 10% FBS were added to each
well and incubated under 5% CO2 at 37°C for 2 min or 5
min. The absorbance was measured using a microplate reader at 450
nm.
RNA isolation
Total RNA was isolated from WI38 and WI38 VA-13 cell
lines using a Qiagen RNeasy Mini kit (74106; Qiagen, Inc.)
according to the manufacturer's instructions. The concentration of
the isolated RNA was confirmed through a SpectraMax QuickDrop
Micro-Volume Spectrophotometer (Molecular Devices, LLC). The
260/280 ratio of the RNA used was >2.1.
RNA-seq and analysis
Using an Illumina TruSeq RNA Sample Preparation kit
V2, purification and library construction were conducted with total
RNA and Illumina HiSeq 2500 machines (Illumina, Inc.) were used for
sequencing with a read length of 2×100 bases. FastQC v.0.11.4
(https://www.bioinformatics.babraham.ac.uk/projects/fastqc/)
was used to assess the quality of the paired-end reads. Cutadapt
v.1.15 (https://cutadapt.readthedocs.io/en/v1.15/) and Sickle
v.1.33 (https://github.com/najoshi/sickle/releases) were used
to filter low-quality reads and adaptors. Cufflinks v.2.2.1
(http://cole-trapnell-lab.github.io/cufflinks/) was
used to calculate fragments per kilobase of transcripts per million
mapped reads values. Cuffdiff (http://cole-trapnell-lab.github.io/cufflinks/) was
used to select differentially expressed genes (DEGs; fold change
>2). All GO analyses were performed using DAVID v.6.8
(https://david.ncifcrf.gov/) and ClueGO
v.2.5.5 in Cytoscape v.3.7.1 (https://cytoscape.org/) (16).
Wound healing assay
WI38 and WI38 VA-13 cells (80% confluence) were
seeded in 6-well plates at 4×105 cells/well using MEM
and RPMI-1640 medium, respectively, supplemented with 10% FBS and
wounded by scratching with sterile plastic 10 µl micropipette tips
after 24 h of PM10 treatment (17). Images were taken of the cells at 0
and 24 h after wounding using a CELENATM S Digital Imaging System
(Logos Biosystems). Cell migration distance was measured in the
images by using ImageJ software version 1.8 (National Institutes of
Health).
Flow cytometric cell sorting
analysis
After PM10 treatment for 48 h, WI38 and
WI38 VA-13 cells were collected and incubated with the Muse Annexin
V and Dead Cell Assay kit (cat. no. MCH100105; Luminex Corporation)
for 20 min at room temperature in the dark. Following incubation,
5×104 cells were analyzed using a Muse cell analyzer
(Merck KGaA). The results were analyzed using Muse 1.5 analysis
software (Merck KGaA).
Statistical analysis
The results were expressed as the means ± SDs (error
bars) of three independent experiments. An unpaired Student's
t-test was performed using GraphPad Prism 5.0 (GraphPad Software,
Inc.). P<0.05 was considered to indicate a statistically
significant difference.
Results
Research strategy for the biological
effects of PM10 based on RNA-seq analysis in lung cell
lines
To evaluate the biological effects of
PM10 treatment in lung cells, two types of normal lung
cell lines (WI-38, fibroblast cells and WI-38 VA-13, epithelial
cells) were selected. Following PM10 treatment for 48 h,
the total RNA was extracted and performed RNA-seq for a
bioinformatic analysis (GO analysis). Cell-specific PM10
effects were evaluated and the biological relationship between the
lung and PM predicted (Fig. 1).
To assess the effect of PM10 on cell viability, CCK-8
and flow cytometric cell sorting analysis was performed following
treatment with PM10 for 48 h. In Fig. 2A, although no significant effects
on cell viability were observed as a result of PM10
treatment, slightly decreased cell growth was observed using bright
field microscopy (Fig. 2B). Flow
cytometric cell sorting analysis with Annexin V showed that the
proportion of apoptotic cells in the PM10 treatment
group was increased compared with that in the PBS group in both
cell lines (Fig. 2C). This
suggested that PM10 treatment may affect cell growth via
induction of apoptosis. To verify whether PM10 treatment
affected cell migration, wound healing analysis was performed
following the treatment of two cell lines with PM10. The
rate of migration in VA-13 cells was decreased compared with that
in the PBS treatment group. However, no difference in the migration
rate in the WI-38 cell line was observed (Fig. 2D). Therefore, it was suggested
that acute treatment with PM10 could induce cell
apoptosis in the two cell lines, but inhibited the migration rate
only in epithelial cells (VA-13), not in fibroblasts (WI-38).
PM10 treatment affects the
adhesion and extracellular matrix (ECM) of fibroblasts
RNA-seq analysis was used to observe the
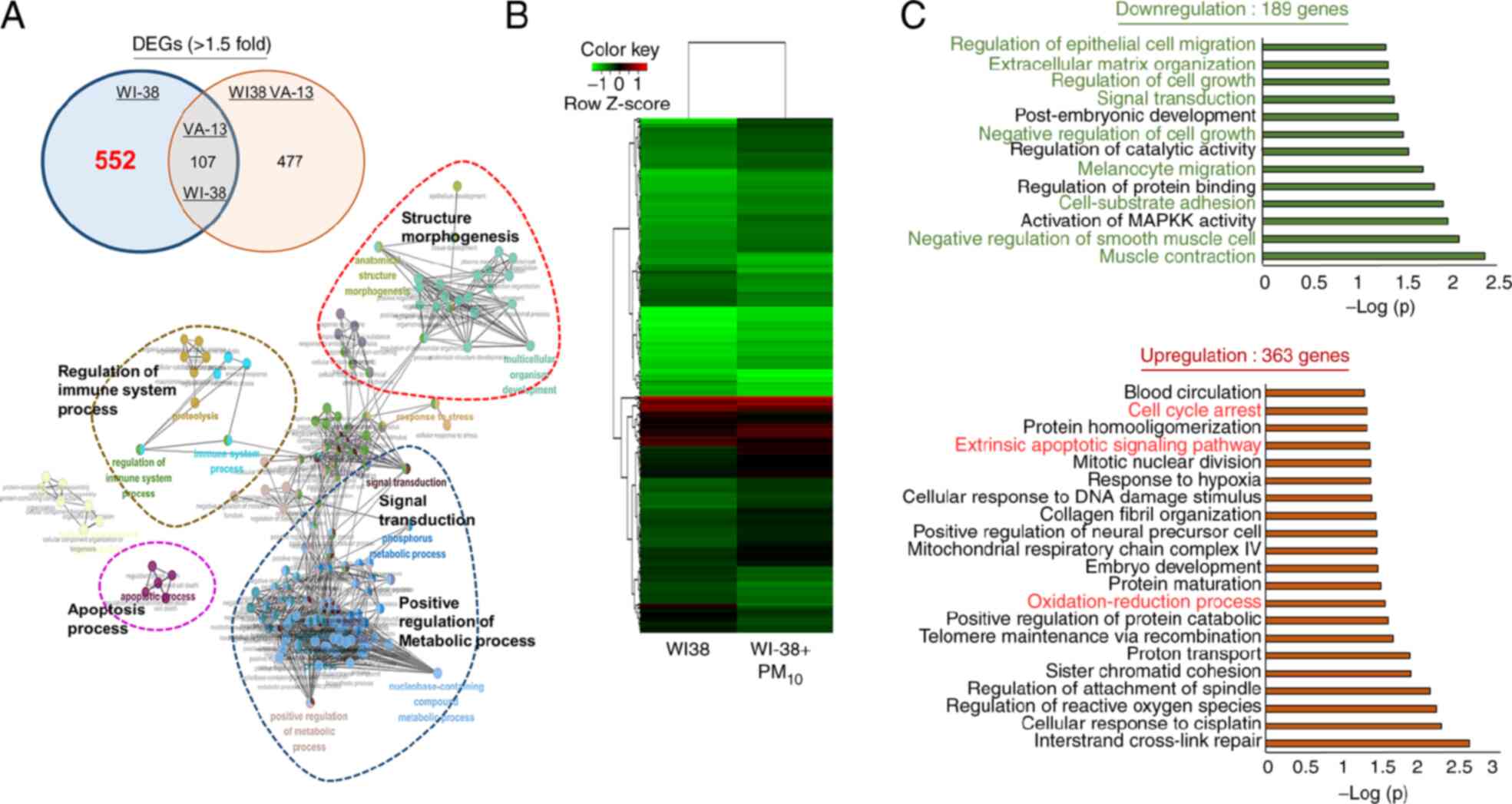
cell-specific DEGs in WI-38 and WI-38 VA-13 normal lung cell lines.
Using 552 WI-38-specific DEGs, GO analysis was performed using the
ClueGO plugin in Cytoscape (Fig.
3A). Fig. 3A shows that
PM10 treatment was associated with ‘apoptotic process’,
‘signal transduction’, ‘regulation of immune system process’,
‘structure morphogenesis’ and ‘positive regulation of metabolic
process’. Additionally, heatmap analysis identified 189
downregulated genes and 363 upregulated genes following treatment
with PM10 in WI-38 cell lines (Fig. 3B). The 189 downregulated genes in
the GO term analysis were associated with migration-related terms
(‘melanocyte migration’ and ‘regulation of epithelial cell
migration’), cell growth-related terms (‘regulation of cell growth’
and ‘negative regulation of cell growth’) and adhesion-related
terms (‘muscle contraction’, ‘cell-substrate adhesion’ and
‘extracellular matrix organization’). In addition, the 363
upregulated genes were associated with ‘cell cycle arrest’, the
‘extrinsic apoptotic signaling pathway’ and ‘oxidation-reduction
processes’ (Fig. 3C). In
conclusion, it was suggested that PM10 treatment
affected cell growth (‘extrinsic apoptotic signaling pathway’,
‘cell cycle arrest’ and ‘negative regulation of cell growth’), cell
adhesion and ECM organization in fibroblasts.
PM10 induces ER stress in
WI-38 VA-13 epithelial cells
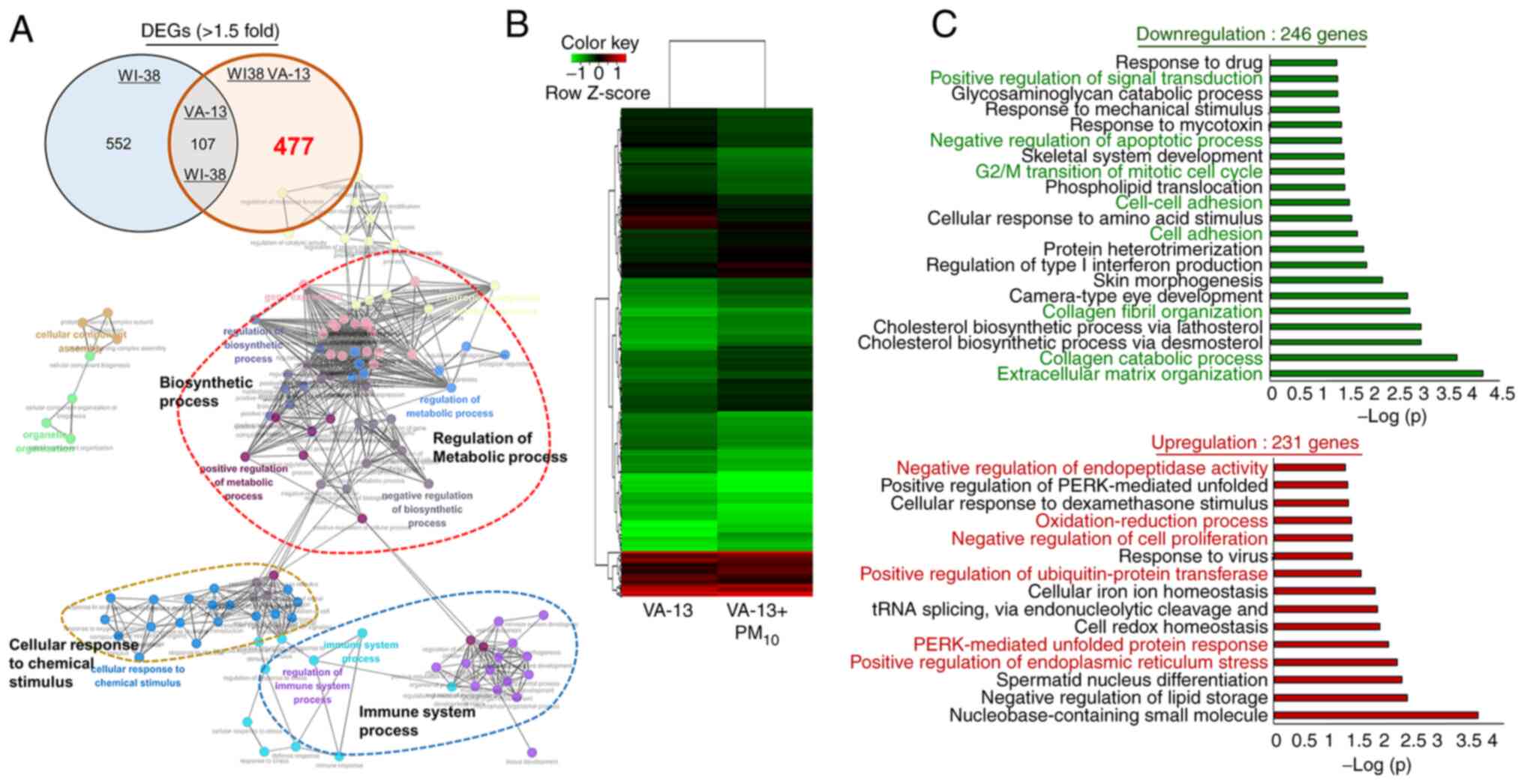
Following PM10 treatment, 477 WI-38
VA-13-specific DEGs were identified using RNA-seq. For the WI-38
VA-13-specific DEGs, a GO analysis was performed using the ClueGO
plugin in Cytoscape and associations with terms related to ‘immune
system process’, ‘cellular response to chemical stimulus’,
‘regulation of metabolic process’ and ‘biosynthetic process’ were
found (Fig. 4A). To analyze the
477 DEGs in greater detail, a heatmap analysis (246 downregulated
genes and 231 upregulated genes) was performed (Fig. 4B). The 246 downregulated genes
were clearly associated with ‘negative regulation of apoptotic
process’, ‘G2/M transition of mitotic cell cycle’, ‘cell-cell
adhesion’, ‘cholesterol biosynthetic process via lathosterol’ and
‘cholesterol biosynthetic process via desmosterol’. In addition,
the 231 upregulated genes were related to ER stress terms,
including ‘positive regulation of endoplasmic reticulum stress’,
‘PERK-mediated unfolded protein response’ and ‘negative regulation
of endopeptidase activity’ (Fig.
4C).
PM10 treatment is related to apoptotic
processes in fibroblast and epithelial cell lines. In the analysis
of DEGs, 107 genes were expressed in both the WI38 and WI-38 VA-13
cell lines following PM10 treatment. The GO analysis
showed that the terms related to ‘regulation of primary metabolic
process’ and ‘cellular response’ were enriched in genes affected by
PM10 treatment (Fig.
5A). Additionally, the heatmap and GO analysis using DAVID
indicated that DEGS in the two cell lines were associated with
apoptosis processes, protein folding and signaling pathways (‘p38
MAPK cascade’ and ‘negative regulation of protein kinase B
signaling’) following PM10 treatment (Fig. 5B and C). Thus, at the
transcriptome level, PM10 treatment was associated with
negative cell growth-related GO terms.
Discussion
GO analysis (DAVID and ClueGO) is a useful method
for evaluating the total biological response of normal lung cell
lines following PM10 treatment using cell-specific DEGs
(18). The present study
presented the cell-specific responses to PM10 observed
in the cell lines WI-38 (lung fibroblast cells) and WI-38 VA-13
(lung epithelial cells) via transcriptome analysis using RNA-seq
results. To select the PM10 concentration for the
assessment of cellular response in normal lung cell lines, a high
concentration of PM10 was selected in the present study.
Chen et al (19)
demonstrate that a concentration of 10 µg/m3
PM10 is associated with lung cancer mortality (3.4~6%
increase) in a cohort of 39,054 participants. From a cohort study
conducted for 12 years, Quezada-Maldonado et al (20) report that a high concentration (10
µg/cm3) of PM10 elicits an acute cellular
response in lung tumors, changing the expression of 45 miRNAs for
72 h. At the in vitro level, cell culture is difficult to
maintain in the long term following PM10 treatment.
Thus, the present study decided on short-term cell culture
following treatment with a high concentration of PM10 (9
µg/cm3) for 48 h to determine the cellular effects of
PM10 treatment. Hence, it demonstrated that continuous
acute stimulation by PM10 had negative effects on cells
and the loss of homeostasis in the cells may induce several types
of disease in humans. However, future animal studies are needed to
examine the long-term effects of PM10.
Lung epithelial cells serve an important role in
maintaining lung homeostasis and host defense mechanisms. In wound
sites caused by toxins, PM and pathogens, several epithelial cell
types spread and migrate as part of repair processes in the lung
(21,22). In wound healing analysis following
PM10 treatment, it was clearly observed that
PM10 treatment suppressed wound closure in epithelial
cells (VA-13), implying that PM10 exposure may inhibit
the wound healing process in the lung defense mechanisms.
Additionally, in flow cytometric cell sorting analysis, the
induction of apoptosis by PM10 treatment was associated
with GO term results (‘negative regulation of cell proliferation’
and ‘apoptosis’). Thus, PM10 treatment in epithelial
cells may be a potential risk factor for several types of lung
disease.
Lung fibroblasts produce ECM components, including
elastin, type III collagen and proteoglycan, for lung structure and
cell adhesion. The ECM is an important factor for physical support
and normal organ function (23,24). Although the present study could
not detect significant differences in cell growth and wound healing
analysis following PM10 treatment, transcriptome
analysis clearly revealed the enrichment of GO terms (‘cell
adhesion’, ‘cell growth’ and ‘extracellular matrix organization’)
suggesting that the primary functions of fibroblasts in the lung
were slightly affected by exposure to PM10 and it was
considered that continuous PM10 exposure may cause
functional defects under normal conditions in lung fibroblasts.
Finally, in the DEGs of both cell types,
PM10 induced changes related to apoptotic processes and
metabolic signaling processes. Thus, the present study suggested
that although brief exposure to PM10 was slightly
reflected in phenotypic changes in cell processes including cell
growth, various processes represented by functional GO terms
(apoptotic GO terms and cell proliferation GO terms) were fully
induced by PM10 treatment. In brief, longer exposure to
PM10 may affect cell conditions and smaller particles
(PM2.5 or PM1.0) could have stronger effects
on lung epithelial or fibroblast cells.
In summary, RNA-seq analysis was performed on lung
epithelial cells and fibroblast cells to identify cell-specific
responses to PM10 treatment. GO analysis of DEGs showed
that PM10 treatment was associated with terms related to
cell apoptosis, immune system processes, cell cycle arrest and ER
stress. Thus, the present study suggested that exposure to PM may
affect cell conditions and long-term exposure to PM is likely to
result in various disease processes (Fig. 5D).
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the National
Research Foundation of Korea (NRF) funded by the Ministry of
Science, ICT and Future Planning (grant nos.
2018M3A9H3023077/2021M3A9H3016046, 2019R1I1A2A01060140 and
2020R1A2B5 B01002028) and by the KRIBB and KIST Research Initiative
Program.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
Conception and design was conducted by DSK, MSL, YJP
and HSC. TYR, JL, KK and SJL developed the methodology. Analysis
and interpretation of data was conducted by KP, MYS and HSC.
Manuscript writing and reviewing was performed by SJL, DSK, MSL and
YJP. Study supervision was by DSK and HSC. All authors have read
and approved the final manuscript. HSC and DSK confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
PM10
|
particulate matter10
|
|
DAVID
|
Database for Annotation, Visualization
and Integrated Discovery
|
|
ClueGO
|
Clustering Gene Ontology
|
|
DEGs
|
differentially expressed genes
|
|
ECM
|
extracellular matrix
|
|
DNMT
|
DNA methyltransferase
|
|
FBS
|
fetal bovine serum
|
|
PBS
|
phosphate-buffered saline
|
|
CCK-8
|
Cell Counting Kit-8
|
|
ER stress
|
endoplasmic reticulum stress
|
References
|
1
|
Kyung SY and Jeong SH: Particulate-matter
related respiratory diseases. Tuberc Respir Dis (Seoul).
83:116–121. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li T, Hu R, Chen Z, Li Q, Huang S, Zhu Z
and Zhou LF: Fine particulate matter (PM2.5): The
culprit for chronic lung diseases in China. Chronic Dis Transl Med.
4:176–186. 2018.PubMed/NCBI
|
|
3
|
Zou Y, Wu Y, Wang Y, Li Y and Jin C:
Physicochemical properties, in vitro cytotoxic and genotoxic
effects of PM1.0 and PM2.5 from Shanghai,
China. Environ Sci Pollut Res Int. 24:19508–19516. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Huang SL, Hsu MK and Chan CC: Effects of
submicrometer particle compositions on cytokine production and
lipid peroxidation of human bronchial epithelial cells. Environ
Health Perspect. 111:478–482. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mu Q, Jiang G, Chen L, Zhou H, Fourches D,
Tropsha A and Yan B: Chemical basis of interactions between
engineered nanoparticles and biological systems. Chem Rev.
114:7740–7781. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang F, Wang P, Pan X, Wang Y and Ren S:
Effects of short-term exposure to particulate matters on heart rate
variability: A systematic review and meta-analysis based on
controlled animal studies. Environ Pollut. 256:1133062020.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pfeffer PE, Lu H, Mann EH, Chen YH, Ho TR,
Cousins DJ, Corrigan C, Kelly FJ, Mudway IS and Hawrylowicz CM:
Effects of vitamin D on inflammatory and oxidative stress responses
of human bronchial epithelial cells exposed to particulate matter.
PLoS One. 13:e02000402018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Watterson TL, Hamilton B, Martin RS and
Coulombe RA Jr: Urban particulate matter activates Akt in human
lung cells. Arch Toxicol. 86:121–135. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hesselbach K, Kim GJ, Flemming S, Häupl T,
Bonin M, Dornhof R, Günther S, Merfort I and Humar M: Disease
relevant modifications of the methylome and transcriptome by
particulate matter (PM2.5) from biomass combustion.
Epigenetics. 12:779–792. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang N, Liu K, Wang K, Zhou C, Wang H,
Che S, Liu Z and Yang H: Dust induces lung fibrosis through
dysregulated DNA methylation. Environ Toxicol. 34:728–741. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wei T and Tang M: Biological effects of
airborne fine particulate matter (PM2.5) exposure on
pulmonary immune system. Environ Toxicol Pharmacol. 60:195–201.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Travaglini KJ, Nabhan AN, Penland L, Sinha
R, Gillich A, Sit RV, Chang S, Conley SD, Mori Y, Seita J, et al: A
molecular cell atlas of the human lung from single-cell RNA
sequencing. Nature. 587:619–625. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ito Y, Correll K, Schiel JA, Finigan JH,
Prekeris R and Mason RJ: Lung fibroblasts accelerate wound closure
in human alveolar epithelial cells through hepatocyte growth
factor/c-Met signaling. Am J Physiol Lung Cell Mol Physiol.
307:L94–L105. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hsia CC, Hyde DM and Weibel ER: Lung
structure and the intrinsic challenges of gas exchange. Compr
Physiol. 6:827–895. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee J, Kim K, Ryu TY, Jung CR, Lee MS, Lim
JH, Park K, Kim DS, Son MY, et al: EHMT1 knockdown induces
apoptosis and cell cycle arrest in lung cancer cells by increasing
CDKN1A expression. Mol Oncol. 15:2989–3002. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee H, Son YS, Lee MO, Ryu JW, Park K,
Kwon O, Jung KB, Kim K, Ryu TY, Baek A, et al: Low-dose
interleukin-2 alleviates dextran sodium sulfate-induced colitis in
mice by recovering intestinal integrity and inhibiting
AKT-dependent pathways. Theranostics. 10:5048–5063. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cappiello F, Casciaro B and Mangoni ML: A
novel in vitro wound healing assay to evaluate cell migration. J
Vis Exp. 568252018.PubMed/NCBI
|
|
18
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen X, Zhang LW, Huang JJ, Song FJ, Zhang
LP, Qian ZM, Trevathan E, Mao HJ, Han B, Vaughn M, et al: Long-term
exposure to urban air pollution and lung cancer mortality: A
12-year cohort study in Northern China. Sci Total Environ.
571:855–861. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Quezada-Maldonado EM, Sanchez-Perez Y,
Chirino YI, Vaca-Paniagua F and Garcia-Cuellar CM: miRNAs
deregulation in lung cells exposed to airborne particulate matter
(PM10) is associated with pathways deregulated in lung
tumors. Environ Pollut. 241:351–358. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Croasdell Lucchini A, Gachanja NN, Rossi
AG, Dorward DA and Lucas CD: Epithelial cells and inflammation in
pulmonary wound repair. Cells. 10:3392021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Farooqui R and Fenteany G: Multiple rows
of cells behind an epithelial wound edge extend cryptic
lamellipodia to collectively drive cell-sheet movement. J Cell Sci.
118:51–63. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pardo A and Selman M: Lung fibroblasts,
aging, and idiopathic pulmonary fibrosis. Ann Am Thorac Soc. 13
(Suppl 5):S417–S421. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
White ES: Lung extracellular matrix and
fibroblast function. Ann Am Thorac Soc. 12 (Suppl 1):S30–S33. 2015.
View Article : Google Scholar : PubMed/NCBI
|