Introduction
Cancer is one of the most lethal diseases.
Conventional treatment consists of surgery, chemotherapy and
radiotherapy. This therapeutic scheme, based on 5FU and oxaliplatin
is useful only when cancer is detected early, but it lacks
specificity and is not practical in the presence of metastases
(1). Moreover, this therapy has
remained mostly unchanged for decades. Besides, all the
aforementioned treatments, and particularly radiotherapy and
chemotherapy, produce severe harmful effects (2). Recently, new approaches that focus
on cancer mechanisms to increase specificity in cancer cell
elimination efficacy and safety are being intensively analyzed.
Some of these new therapies introduce the use of nanoparticles,
monoclonal antibodies, miRNAs, gene and cell therapy with the use
of cells as delivery vectors for antitumoral proteins (3–10).
All promise to be more specific, efficacious and safer than
conventional treatments.
In the present study, engineered mesenchymal stem
cells (MSCs) were used as antitumoral therapy, considering that
MSCs gather several desirable features (11) as anticancer weapons, such as their
abundance in tissues, particularly in bone marrow (BM) (12) and adipose tissue (13). Besides, these cells have the
outstanding ability for ‘homing’ to the tumor microenvironment.
Homing allows MSCs to exert their anticancer effects directly
towards tumors (14).
Nevertheless, a serious limitation for using MSCs as anticancer
therapy is that it is hard to predict the naïve MSCs behavior in
the tumor, as these cells, under the influence of the tumor
microenvironment, can improve tumor growth (15–17). Thus, an alternative is needed that
increases the anticancer potency of MSCs and reduces the risk of
favoring cancer progression instead of fighting it. Achieving this
goal is possible since MSCs offer the practicability of being
genetically engineered to overexpress coded proteins and find and
attack malignant cells. Therefore, engineering MSCs can ensure that
these cells specifically attack malignant cells and overcome the
naïve MSCs efficacy for fighting tumors. To transduce murine
BM-MSCs, the murine soluble fraction of the TNFα-related
apoptosis-inducing ligand (sTRAIL) and interferon β (INFβ) was
chosen, considering the following features of each transgene:
sTRAIL is a powerful anticancer protein extensively used due to the
selective apoptotic effect that this protein induces in malignant
cells. TRAIL targets the death receptors (DR)4 and DR5, which
activate apoptosis by a caspase-8 derived process; the
aforementioned receptors are mainly overexpressed by malignant
cells (18,19). sTRAIL exerts its function once it
has been trimerized and bound with cell receptors. However, Wong
et al (19) pointed out
that full-length (fl)TRAIL can induce apoptosis more efficiently
than soluble (s)TRAIL. Interferon beta (IFNβ) is a type I member of
the interferon family with pleiotropic roles, including
immunomodulator, antiproliferative and cancer-inhibitory activity
(20). IFNβ has already been
examined and reported as an effective gene therapy agent (21); however, it was hypothesized in the
present study that the combination of both proteins could increase
the reach of monotherapy efficiency. To achieve this goal, a murine
syngeneic model of lymphoma was used, with an immunocompetent
strain (BALB/c) and BM-MSCs freshly obtained from mice belonging to
the same BALB/c strain. The purpose of the present study was to
determine the percentage of surviving mice and lymphoma reduction
in mice implanted with MSCs carrying sTRAIL or IFNβ transgenes and
their combinations.
Materials and methods
Mice
BALB/c mice were born, grown and maintained in the
Laboratory of Animal Experimentation of the Autonomous University
of Nuevo Leon. Male mice aged 4 to 8 weeks were used to isolate
BM-MSCs. A total of 72 female BALB/c mice (12 weeks old) weighing
19–22 g were used to induce solid lymphomas and evaluate the in
vivo anti-lymphoma efficacy. From mice birth and throughout the
experiments until the last mouse was euthanized, all animals were
kept in individually ventilated cages at 25°C with sterilized air
with EPA filters (UREAtac; LAB&Bio), with 50% humidity, ≤500
ppm of CO2, ≤15 ppm of NH3 and a light/dark
cycle of 12/12 h. Access to food (Labdiet 5001; LabSupply) and
purified sterile water was provided ad libitum.
Lymphoma cells
The thymic lymphoma cell line L5178Y was used to
produce lymphomas; the clone that was used (3.7.2C) was derived
from the L5178YS cell line. In vitro, these cells grow in
suspension; in vivo, when inoculated intramuscularly, they
produce solid lymphomas, and when injected intraperitoneally, they
grow abundantly and produce ascites. The L5178Y cell line was
purchased from the American Type Culture Collection (cat. no.
CRL-9518™; ATCC®) and cultured as instructed by the
manufacturer. The culture medium was Dulbecco's modified Eagles
medium (DMEM) high glucose medium (4.5 g/l), 0.1% pluronic, 100 µg
gentamycin/ml, 2.5 µg amphotericin B/ml, and 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.).
Lentiviral vectors
Lentiviral vectors were produced and provided by
Cyagen Biosciences, Inc. as a gene delivery tool, named as follows:
pLV[Exp]-EGFP/Neo-EF1A>{sMurTRAIL},
pLV[Exp]-EGFP/Neo-EF1A>mIfnb1[NM_010510.1],
pLV[Exp]-EGFP/Neo-EF1A>{MurIL12-70p}. The lentiviral vectors
were of 3rd generation and also included an integration cassette
for G-418 (geneticin; Gibco; Thermo Fisher Scientific, Inc.)
resistance for cell selection and a green fluorescent protein (GFP)
gene to evaluate transgene integration.
MSC isolation and cell culture
BM-MSCs were obtained and characterized as
previously described (22).
Briefly, six mice were placed inside a CO2 chamber to
sacrifice them (70% vol/min); immediately after, their femurs and
tibias were dissected under aseptic conditions. Their epiphysis was
removed, and using a hypodermic needle (27 gauge), their BM was
obtained by perfusing the shafts inner barrel with 500-1,000 µl
DMEM/Nutrient Mixture F-12 (DMEM F-12) medium supplemented (s) with
10% FBS, 100 µg gentamicin/ml, and 2.5 µg amphotericin B/ml (Gibco;
Thermo Fisher Scientific, Inc.) and poured into T-25 cell culture
flasks (Corning, Inc.). The first cell passage was accomplished as
follows: The aforementioned preparations were incubated for 24 h at
37°C in a 5% CO2 humid atmosphere. The expended medium
and the non-adherent cells were discarded. The culture flasks with
the adherent cells were replenished with 4 ml fresh medium and
incubated as before until the cell monolayer reached 80% confluence
(7–10 days). The second passage was performed by discarding the
spent medium and incubating the cell monolayer with a solution of
0.25% trypsin and 0.1% EDTA (Gibco; Thermo Fisher Scientific, Inc.)
in sterile phosphate-buffered saline (PBS), washing trice with PBS
and inoculating 5×105 cells in a new culture flask with
fresh 5 ml of supplemented DMEM F-12 and incubating it as before.
The procedure described for the second passage was repeated in each
of the following passages.
MSC characterization
The characterization of MSCs and the assays
performed with them were accomplished with cell cultures from
passage 3 or 4. MSCs were characterized by immunocytochemistry and
their multipotency was evaluated (Fig. 1). Cells were fixed with
methanol-acetone (1:1) at 4°C per 10 min. Regarding
immunocytochemistry, CD90 and CD105 surface markers were
investigated using specific monoclonal antibodies included in the
mouse and rabbit specific HRP/DAB (ABC) detection IHC kit (cat. no.
ab64264; Abcam). Antibodies were diluted in PBS to 1:200 (CD90) and
1:25 (CD105). MSC multipotency was examined by inducing the cells
to differentiate to osteoblasts or chondroblasts, using the
appropriate reagents in the Mouse Mesenchymal Stem Cell Functional
Identification kit (cat. no. SC010; R&D Systems, Inc.) and
following the manufacturer's instructions. The osteoblasts were
identified by staining with the Von Kossa technique and
chondroblasts with Alcian blue. Cells were fixed with 4%
paraformaldehyde for 20 min at ambient temperature. In Von Kossa
staining, slides were submerged in saturated lithium carbonate
solution for 20 min, and then washed with water and submerged in a
5% silver nitrate solution. Slides were exposed to bright light for
60 min and then were submerged in 5% sodium thiosulfate solution
for 5 min. Finally, the slides were counterstained with neutral
red. In Alcian blue staining, the colorant was added at ambient
temperature for 10 min, and then the slides were washed with water
and counterstained with neutral red at ambient temperature for 2
min.
Construct design
All transgene constructs contained murine sequences
of the genes of interest, a promotor cassette for Elongation Factor
1-Alpha (EF1-α) to allow a constitutive, high protein expression,
and the Kozak consensus sequence (GCC GCC ACC) (23). The constructs used in the present
study were designed with Vector Builder software (Cyagen
Biosciences, Inc.). The design of the sTRAIL transgene is presented
in Fig. 2A. This design contained
the sTRAIL nucleotide sequence codifying the amino acid fragment
118 to 291; an isoleucine zipper sequence to facilitate in
situ TRAIL's trimerization (24,25), a furin specific cleavage site
(CGCACCAAACGC), a signal sequence (SS) derived from Gaussia
sp. (a copepod) luciferase (TGG GAG
TCAAAGTTCTGTTTGCCCTGATCTGCATCGCTG-TGGCCGAGGCC) to conduct protein
secretion (26) and Kozak's
sequence. As revealed in Fig. 2B,
besides EF1-α and Kozak's sequence, the construct used to perform
the transduction of BM-MSCs with the flTRAIL transgene contained a
nucleotide sequence coding its 1–291 amino acids. The entire
flTRAIL was retracted from GeneBank (accession number U37522, URL:
http://www.ncbi.nlm.nih.gov/nuccore/U37522). Regarding
IFNβ, the murine IFNβ gene sequence was retracted from
GeneBank (accession number NM_010510, URL: http://www.ncbi.nlm.nih.gov/nuccore/NM_010510).
As demonstrated in Fig. 2C, the
scheme of the IFNβ transgene design was as follows: The complete
IFNβ nucleotide sequence was included and added with Kozak's
sequence.
Transduction of murine BM-MSC
BM-MSCs (5×105) were seeded into T-25
culture flasks (Corning, Inc.) containing 5 ml DMEM F-12
supplemented with 10% FBS and incubated overnight under standard
conditions, 50% confluency. Then, protamine sulfate
(Sigma-Aldrich), 5 µg/ml were added to the BM-MSC cultures. Each
lentivirus preparation carrying sTRAIL, flTRAIL, or INFβ transgenes
was added to separate BM-MSC cultures. The multiplicity of
infection (MOI) employed was two viral particles/one BM-MSC
(27). The BM-MSCs infected with
the lentivirus vectors were incubated overnight, to allow viral
infection. Then, spent media was discarded and replenished with
fresh DMEM-F12 and incubated 48 h before fluorescence analysis.
Lentivirus integration was validated with fluorescence emitted by
GFP, which was visualized with a fluorescence microscope (Eclipse
10i; Nikon Corporation) and a representative field photographed
with a Sight DS-2MV (Nikon Corporation) digital camera. The
selection of transduced BM-MSCs was accomplished by adding 200 µg
of geneticin/ml to the culture medium. Transduction efficiency was
calculated by applying the following equation: % Transduction
efficiency=BM-MSC-GFP/total BM-MSC ×100. Where BM-MSC-GFP are
transduced cells that exhibit a green fluorescent signal under the
epifluorescence microscope. Total BM-MSC is the total number of
cells detected by counting nuclei by fluorescence with DAPI (blue
signal). Mounting medium Vectashield with DAPI (7 µl) per slide was
applied (Vecta Mount; Vector Laboratories, Inc.).
Protein extraction
Gene expression was validated by western blot
analysis. BM-MSCs (5×105) transduced with sTRAIL,
flTRAIL or INFβ were placed, separately, into T-75 flasks (Corning,
Inc.), containing 10 ml of DMEM-F12 and incubated at 37°C with 5%
CO2 for 48 h and allowing 70% confluency. The spent
medium was discarded, and then the cell culture flasks were
replenished with 10 ml fresh supplemented media and incubated at
37°C with 5% CO2 for 48 h. For soluble proteins (sTRAIL
and IFNβ), the spent medium was centrifuged at 4,000 × g for 5 min,
concentrated 40 times with an Amicon filtration unit of 10 kDa MW
cut-off (Merck KGaA). FlTRAIL is a transmembrane protein that was
extracted and purified. FlTRAIL-BM-MSCs were cultured as
aforementioned with the sTRAIL-BM-MSCs or IFNβ-BM-MSCs until
flTRAIL-BM-MSCs culture reached 80% confluency. Then,
flTRAIL-BM-MSCs were harvested and washed twice with PBS.
Subsequently, 1×106 cells were resuspended in 50 µl of
lysis buffer [Triton X-100 (50 ml), Tris-HCl 1 M pH 7.5 (5 ml), KCl
2 M (12.5 ml), MgCl2 1 M (1 ml), DTT (1 µl, Invitrogen;
Thermo Fisher Scientific, Inc.) and a protease inhibitor cocktail
(5 µl; Sigma-Aldrich; Merck KGaA)]. The mixture was adjusted at 500
µl with ultrapure sterile water, chilled in ice for 20 min,
centrifuged at 16,000 × g at 4°C for 5 min, and the supernatant was
collected.
Western blot analysis
The aforementioned protein concentrates were
quantified with Bradford reagent (Bio-Rad Laboratories, Inc.).
After, 40 µg protein concentrate (5 µl) was submitted to 2% sodium
dodecyl sulfate (SDS), 4% stacking and 12% resolving polyacrylamide
gel electrophoresis (PAGE) at 100 V for 120–150 min and the protein
bands were electro-transferred to a 0.45-µm mesh polyvinylidene
fluoride membrane (Bio-Rad Laboratories, Inc.) at 100 V for 60 min.
Protein was Electro-transfer to a PVDF membrane. The membrane was
blocked for 1 h with 5% skimmed milk in Tris-buffered saline with
Tween-20 (TBST) at 4°C. The protein bands of sTRAIL, flTRAIL, or
IFNβ were identified with rabbit monoclonal antibodies anti-murine
TRAIL (1:200; cat. no. ab10516; Abcam) or 1:500 rabbit polyclonal
anti-murine IFNβ (cat. no. ab85803; Abcam), as primary antibodies,
incubated overnight at 4°C. The secondary antibody was an
HRP-conjugated anti-rabbit polyclonal antibody (1:10,000; cat. no.
W4011; Promega Corporation), incubated for 2.5 h at 4°C. Dilutions
of all antibodies were performed with Tris-buffered saline (TBS
buffer) in 5% skimmed milk.
The marked protein bands were revealed with the
luminol kit Clarity Western ECL blotting substrate (Bio-Rad
Laboratories, Inc.) following the manufacturer's protocol and
analyzed with the Molecular Imager Chemidoc XRS+ Imaging System
(Bio-Rad Laboratories, Inc.) equipped with the Image Lab
software.
Commercial recombinant proteins were used as
positive controls in all western blot assays; soluble TRAIL
recombinant protein (cat. no. 315-19; PeproTech, Inc.) which
consists in the extracellular fraction of murine TRAIL, weighing 20
kDa; ~200 ng of recombinant protein was loaded into the gel well.
IFNβ standard (cat. no. IF011; Merck KGaA) consisted of recombinant
murine IFNβ weighing 22 kDa; ~2 kU was loaded into the gel
well.
Induction of lymphomas
L5178Y cells (5×105) were inoculated in
the right gastrocnemius with a 27-gauge syringe. The cells were
suspended in 100 µl of saline solution for a total of 72 female
BALB/c mice. The mice exhibited mild discomfort immediately after
inoculation; however, no sign of discomfort was observed overnight.
The implanted leg of each mouse was measured with a Vernier
calibrator to calculate the gastrocnemius volume (mm3),
using the following equation: Vol=π/6 (LxWxh); where Vol is the
volume of the leg, expressed in mm3, π is a radius
constant (3.1416), L is the leg's length, measured from the
ankle to the knee. W is the width of the thickest portion of
the leg, and h is the height, measuring from the rear leg
section (gastrocnemius) to the front (tibialis anterior). These
measurements were performed every third day throughout the
experiment. When legs carrying lymphomas showed an increment in the
leg volume of 1–2 mm3, mice were implanted with the
transduced BM-MSCs.
Implantation of transduced
BM-MSCs
Mice with lymphomas were divided into nine groups of
eight mice each. Transduced BM-MSCs were implanted to one of these
groups (G) as follows: G1, BM-MSCs-sTRAIL; G2, BM-MSCs-IFNβ
plus-flTRAIL; G3, BM-MSCs-IFNβ; G4, flTRAIL; G5, sTRAIL plus IFNβ,
G6, not transduced or naïve BM-MSC plus-IFNβ plus-flTRAIL; G7,
naïve BM-MSC; G8, mice not implanted with BM-MSCs but injected with
isotonic saline; G9, mice not implanted with BM-MSCs. In addition,
the experiment was repeated with groups G5, sTRAIL plus IFNβ, and
G6, not transduced BM-MSC to confirm results. However, mice were
reduced to five animals per group with the same characteristics. In
all implanted mice, 1×106 viable BM-MSCs, suspended in
100 µl isotonic sterile saline, were distributed into the entire
volume of each tumor. The number of BM-MSCs transduced separately
with two transgenes or two transgenes plus not transduced MSCs were
inoculated with an appropriate proportion of BM-MSCs to sum a total
of 1×106 cells. For example, G5 mice were implanted with
5×105 BM-MSCs-sTRAIL and 5×105-INFβ and mice
from G6 were implanted with 3.33×105 not transduced
BM-MSC, plus 3.33×105 BM-MSCs IFNβ plus
3.33×105 flTRAIL. All inoculations were performed in
aseptic conditions with a new, sealed sterile 27-gauge needle for
each mouse. Before and throughout the experiments, mice behavior,
their ability to eat and drink, and pain indicators were monitored.
When a mouse showed intense suffering as previously described
(28), these signs included bent
down low ears, orbital tightening, bristly hair, sluggishness, or a
lack of response to external stimuli. When all signs were detected,
the mouse was euthanized. These animals were considered deceased
due to their induced lymphoma. Euthanasia was practiced by cervical
dislocation. Tumor volume and mouse weight measuring were performed
thrice a week, as at the beginning of the experiment.
Tumor biopsies of ~5 mm3 were dissected
from recently deceased animals. The samples from each lymphoma were
preserved with Tissue-Tek optimum cutting temperature (O.C.T.)
(Sakura Finetek Europe B.V.), freezing the blocks with liquid
nitrogen and stored at −80°C.
Histopathological analysis
Frozen samples were cut with a cryostat (model 2230;
Shenyang Roundfin Trade Co., Ltd.), stained with hematoxylin and
eosin, and observed with a clear field using a light microscope
(model DM IL LED; Leica Microsystems GmbH) at 100–400 amplification
diameters. Images were captured with a camera (DFC295; Leica
Microsystems GmbH) attached to the microscope.
Analysis of results
The number of deceased mice from all groups was
monitored daily. When the last not-implanted mouse with BM-MSCs
carrying lymphomas succumbed, the percentage of surviving mice from
each group and the number of days elapsed since the lymphoma cells
were inoculated were registered.
Statistical analysis
The survival rate was analyzed with a Kaplan-Meier
estimator with a Mantel-Cox test to assess statistical significance
with SPSS software (SPSS Statistics for Windows; version 22.0; IBM
Corp.). Statistical analysis was conducted using GraphPad Prism
version 7.00 for Windows (GraphPad Software, Inc.). Differences
between the analyzed groups were determined with the t-test and
one-way ANOVA (followed by Tukey's post hoc test). P<0.05 was
considered to indicate a statistically significant difference.
Ethical considerations
The present study was approved (approval no.
BI15-005) by the Scientific Research and Bioethics Committee and
the Institutional Animal Care Committees of the School of Medicine
of the Autonomous University of Nuevo Leon (Monterrey, Mexico). The
mice were handled according to the Mexican Standard
NOM-062-ZOO-1999. The mice were kept in the animal facility of the
Department of Immunology in individually ventilated cages (Lab
& Bio) that maintain filtered air flow and a barrier that
protects them from contact with external agents. The system records
pressure, oxygen, CO2, humidity, temperature, and
ammonia concentration. Furthermore, this system maintains all these
parameters in uniform conditions. The cages were cleaned every 2–3
days by the technical personnel of the vivarium. The model was
euthanized if the tumor exceeded 10% of the normal mouse body
weight according to the international manual Institutional Animal
Care and Use Committee (2002) and would be euthanized by inhalation
of CO2 with subsequent cervical dislocation and
monitoring of vital signs. They were also euthanized if they showed
signs of severe pain according to the Rodent Management Laboratory
Manual accepted by the American Association for the Accreditation
of Animal Care Laboratories (29): decreased intake of water and food,
weight loss (>20%), bristly coat, dry skin, lacerations,
posture, abnormal gait, head tilt, lethargy, bloating, diarrhea,
seizures, discharge from body orifices and dyspnea. The
experimental groups were kept under close care from the day of
inoculation until the last mouse in the control group succumbed.
The mice succumbed due to tumor growth. However, the mice in the
experimental groups that survived until the last mouse in the
untreated group were sacrificed on the same day that the last
untreated mouse lived.
Results
BM-MSCs express TRAIL and IFNβ
BM-MSC isolated from mice presented fibroblast
morphology in culture and were characterized by expression of CD90
and CD105 superficial markers that presented >95% positive cell
staining (Fig. 1A-C). In
addition, BM-MSC differentiated to chondroblasts and osteoblasts,
showing positivity to Alcian blue and Von Kossa stains,
respectively (Fig. 1D and E). A
representative field of transduced BM-MSCs is shown in Fig. 2D. All cells fluoresced intensely
in green due to expression of the GFP gene and geneticin selection,
denoting an efficient integration of each gene construct. The
percentage of transduction efficiency of BM-MSC-sTRAIL and -flTRAIL
was 90% and for BM-MSC-IFNβ was 95%. For protein expression,
western blot results are shown in Fig. 2E and F. The protein bands, whose
images are demonstrated in Fig.
2E, correspond to sTRAIL and an sTRAIL standard, respectively,
with both weighing ~20 kDa (control). The image of flTRAIL,
revealing a ladder of five bands; the smallest one weighs ~20 kDa,
which matches the 2 soluble TRAIL standard control; however, 4
heavier bands of ~30, ~39, ~45, and ~60 kDa were also revealed. In
Fig. 2F, two IFNβ bands are
demonstrated, one weighing ~22 kDa and a slightly heavier ~24 kDa
band. Alongside as control, the recombinant IFNβ standard shows a
single band weighing ~22 kDa.
BM-MSC expressing sTRAIL and IFNβ
reduce solid lymphoma engraftment
On day 7 post-inoculation (PI), a slight
inflammation was observed in all inoculated subjects. PI leg
volumes on day 9, comparing the non-inoculated and inoculated
groups are revealed in Fig. 3;
considering one standard deviation as a reference, it displays the
totality of the study subjects with tumor engraftment.
First, deceased mice from the untreated group were
registered by day 26 PI; therefore, the last tumor measure with
full subjects' groups was on day 23 PI. Fig. 4A highlights the most relevant
results: the untreated group shows the largest tumor (average 2.5
cm3) and first deceased mouse at day 26 PI; on the other
hand, the group with combined treatment of IFNβ and sTRAIL showed
the smallest tumor (1.2 cm3) by day 23, and the first
mouse succumbed by day 29 PI. Notably, separated treatments of both
sTRAIL and IFNβ also showed considerably smaller tumors (compared
with the untreated group) but slightly larger than the combined one
(1.3 and 1.5 cm3, respectively).
BM-MSC expressing sTRAIL, flTRAIL and
IFNβ treatments do not affect mice weight
During the study, mice weights were documented. Any
mouse that exhibited a loss of ≥20% of the baseline weight was
withdrawn from the study and euthanized. The weight of all groups
during the study is revealed in Fig.
4B; mice showed stable weight by day 19 PI; however, important
weight variability was registered thereafter. All mice were
carefully observed to detect a disability to eat or drink; however,
despite massive tumors, mice were eating and drinking; thus, weight
variability was associated with tumor growth. All subjects gained
weight according to the developed tumor mass.
In addition, tumor tissues were analyzed with a
light microscope. A section of the tumor of untreated mice was
compared with tissue of the inoculated leg of surviving mice.
Images of histological sections are revealed in Fig. 5. It was demonstrated how the tumor
tissue almost completely displaced muscle tissue (Fig. 5B and D), exhibiting basophilic
predominance of lymphocyte (tumoral cell) nuclei, which in contrast
to a surviving mouse, most of the tissue was acidophilic with
skeletal muscle tissue (Fig. 5A and
C). It was also observed that a few nuclei dispersed over the
muscle tissue, indicating an active immune reaction. The
statistical analysis of the infiltration rate and morphology
observed in treated tissues is revealed in Fig. 5E and F. Chi-square showed
differences between the control and the treatment group
(P<0.001); however, in morphology, no statistical differences
were identified.
BM-MSC expressing sTRAIL and IFNβ
increase survival of mice with solid lymphoma engraftment
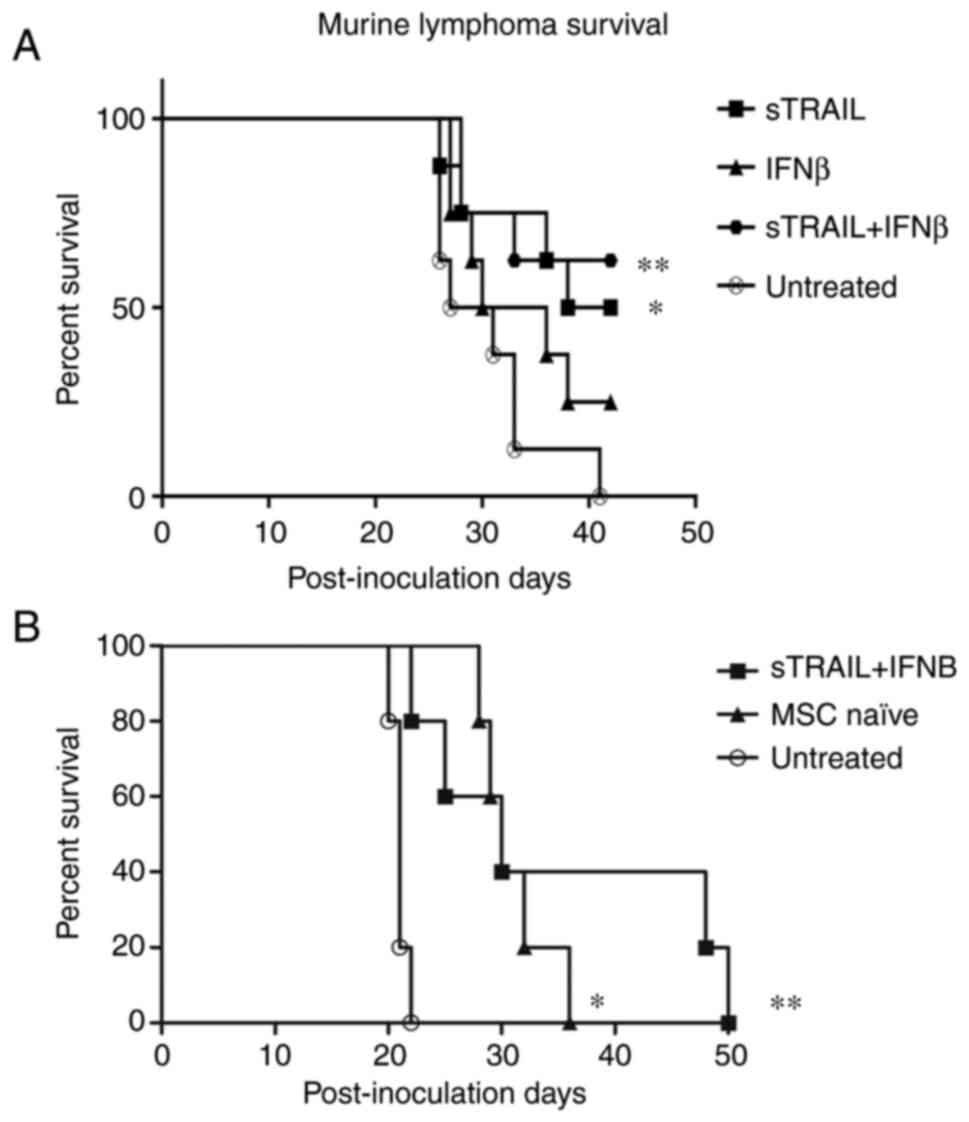
The last untreated mouse died at day 41 PI; thus, a
cut-off point was established. All groups comparing each treated
group against the untreated group were analyzed. In Fig. 6A, a Kaplan-Meier estimator with
the most relevant results is demonstrated: 100% of untreated mice
succumbed due to the tumor by day 41 PI. On the other hand, the
combined treatment of IFNβ and sTRAIL showed a 62.5% survival rate.
Additionally, the separated groups of IFNβ and sTRAIL showed 25 and
50% survival, respectively, in addition to initial tumor mass,
which showed a similar result, suggesting an additive effect. The
log-rank test showed significant differences between the groups
(P<0.05). All P-values are included in Table I.
 | Table I.Survival significant differences
between treated and non-treated groups. |
Table I.
Survival significant differences
between treated and non-treated groups.
| Treatment | P-value |
|---|
| sTRAIL | 0.017 |
| flTRAIL + IFNβ | 0.068 |
| IFNβ | 0.170 |
| flTRAIL | 0.072 |
| sTRAIL + IFNβ | 0.008 |
| MSC naïve + IFNβ +
flTRAIL | 0.042 |
| MSC naïve | 0.038 |
| Saline
solution | 0.309 |
To confirm these results, the combination treatment
of BM-MSC expressing sTRAIL and IFNβ was repeated. The results are
presented in Fig. 6B. The control
treatment of naïve BM-MSCs was added. At first, both treatments
reduced tumor growth in the right gastrocnemius; however, near day
20, mice of the groups treated with naïve BM-MSCs presented an
increase in tumor volume. On day 36 PI, the last mouse of the naïve
BM-MSC group was euthanized, and 40% of survivors remained in the
group treated with sTRAIL plus IFNβ. The log-rank test showed
significant differences between the groups (P<0.001). Mice were
kept monitored until day 50; nevertheless, mice did not present
visually solid tumors in the right gastrocnemius.
All treatments showed anti-tumorigenic activity to
some extent. Independent treatments showed poorer antitumor
activity than combined ones. IFNβ exhibited the lowest therapeutic
activity with a 25% survival rate, sTRAIL showed 37.5%. Combined
treatment showed 37.5% survival rate, indicating a non-additive
effect since it remained nearly the same as when treatment was
performed with sTRAIL alone. Notably, saline solution had a 25%
survival rate. A combined treatment of MSC naïve, IFNβ and flTRAIL
showed a 50% survival rate, indicating a slight improvement over
flTRAIL and IFNβ alone, suggesting MSC antitumoral effect.
Discussion
Cancer xenograft in vivo murine models have
been a frequent approach in numerous research groups; indeed, the
chimeric models of human tissue in immunodeficient mice have
allowed achievement of tremendous advances and testing several
cancer models and treatments (30). The current landscape deems murine
models as the cornerstone in biomedical research (31).
MSC approaches in murine cancer models involve the
following criteria: first, a cancer model; that is, a
tumor-inducing cell line in vivo of certain tissue; second,
determination of the source of the MSCs, as there are significant
phenotypic and genotypic differences among the cells in terms of
regenerative capacity, multipotency, isolation yield, and overall
behavior within the tumor; third, one or more anticancer transgenes
to be expressed and delivered; and finally, fourth, a mouse strain
according to the aims of the study (32). CD105, CD90 and CD73 are main
markers highly expressed by BM-MSCs (33). In the present study, CD105 and
CD90 were only evaluated. CD73 expression on MSCs was reported
heterogeneity from different sources as adipose tissue, BM,
amniotic membrane (34). However,
previous research has shown that CD73 has the power to promote cell
proliferation, tumor angiogenesis, and has the ability to make
tumor cells escape immunological recognition (35), so its evaluation would be very
interesting for future studies.
MSCs have been reported to possess homing ability
led by chemokines and growth factors from the tumor site. Moreover,
this therapeutic effect depends on the capacity of reaching the
tumor site, by migration, expression of adhesion molecules, and
engraftment in the tumor microenvironment. It has been previously
reported that MSCs decrease the incidence of lymphoma tumors and
improve survival of C57BL/6 mice when intravenously administered
(36). In addition,
intraperitoneal administration of MSCs in a disseminated
non-Hodgkin's lymphoma murine model increases the overall survival
of treated mice (37). Both
studies demonstrated MSC homing to tumoral cells. Nevertheless,
there is a lack of enough lymphoma studies that determine the
complete mechanisms of antitumoral activity of MSC, which opens an
important field to research. In the present study, a murine,
syngeneic, immunocompetent, lymphoma model treated with BM-MSCs
overexpressing TRAIL (soluble fraction or full-length protein) and
IFNβ by intratumoral inoculation was employed to elucidate if
unmodified MSC would exert a pro-tumoral or anti-tumoral effect and
if such an effect would be reverted or enhanced, respectively by
the transgenes alone or combined.
The tumor microenvironment is determinant in tumor
progression as T and B lymphocytes are strongly influenced by it
and vice versa, affecting other cells. T-cells are the most
important line of defense regarding cancer development;
nonetheless, tumors can produce TGF-β, which favors T cell
conversion to regulatory T-cell, which exhibits protumor effects by
immunosuppression (38).
B-cell subsets have demonstrated cooperative or
opposite roles within the tumors. Immunosuppressive regulatory B
cells or tumor-infiltrating lymphocytes have protumor effects as
they tend to be up to 25% of cells within the tumor mass, can
produce lymphotoxin, an angiogenesis-inducing protein (39), and secrete antibodies against
tumor-specific antigens (such as p53) (40) with variable outcomes. On the other
hand, there are also effector B-cells with antitumor effects by
antigen presentation, CD4 T-cell activation, and macrophage
conversion to antitumoral M1 phenotype (41). In aims for an improved
understanding of cancer development and treatments, in our opinion,
a homogenous, syngeneic model provides substantial advantages.
To the best of our knowledge, there are no reports
of lymphoma immunocompetent models with combined TRAIL (soluble
fraction and complete) and IFNβ treatments. Soluble TRAIL required
several accessories to be secreted extracellularly to exert its
intended function: an isoleucine zipper to facilitate ligand
trimerization (25), a signal
sequence-derived Gaussia princeps luciferase (26) to conduct protein extraction, and a
furin cleavage site to eliminate all upstream accessory sequences
(24); all transgenes proved to
be recognizable as they were easily detected by western blotting.
FlTRAIL showed 5 bands (weighing ~20, ~30, ~39, ~45 and ~60 kDa),
which differs from a 24–25 kDa expected weight; however, similar
reports suggested a glycosylated version of the protein (27) or possible oligomerization of the
ligand. Additionally, a ~20 kDa band suggests the expression of
soluble TRAIL as well. IFNβ showed 2 bands (~22 and ~24 kDa),
similar to flTRAIL; the smaller band matched the recombinant
standard, and the observed ~24 kDa band is probably a glycosylated
version of the protein.
TRAIL has been examined in numerous studies and
clinical trials due to its unique property of targeting transformed
cells, but its short half-life and toxicity make it impractical;
thus, the need for a dose-based therapy by cell delivery is
plausible. Soluble TRAIL fraction is normally secreted by the cell
to procure DR4+ or DR5+ cells more easily
than the complete protein; thus, flTRAIL is anchored to the cell
membrane. Therefore, it requires a markedly closer cell-cell
approach to trigger apoptosis. Notably, Yuan et al (27) demonstrated that flTRAIL exhibited
a more efficient cell-killing effect in TRAIL sensitive and
resistant cell lines in vitro. Conversely, in the present
in vivo study, sTRAIL resulted in the best single-gene
treatment in tumor-reducing effects (with an average volume of 1.4
cm3 leg-tumor size as opposed to flTRAIL with 1.5
cm3) and survivability (50% compared with 37.5% for
flTRAIL). This finding could be due to inherent differences between
in vitro and in vivo models and protein presentation,
as multimers of flTRAIL are more likely as they are suspended in a
controlled environment. MSCs administered intravenously or in
another form of distant administration require a large number of
cells due to loss and death on the way to the tumor site (4,11);
the majority get trapped in capillaries and in the lungs owing to
their large size and expression of adhesion molecules (2). Despite this, the ones that arrive at
the tumor site could develop activity, which in turn is enhanced
through genetic modification to express cytokines and pro-apoptotic
proteins (11–13). By contrast,
intratumoral-inoculated cells are significantly more restricted,
relying only on cells close to the tumor parenchyma puncture site;
additionally, the soluble TRAIL construct was designed to be
secreted abundantly and perfused more efficiently through the
tumor. Furthermore, to reduce variation during the administration
of the MSCs in each of the experimental groups, the inoculation of
cells was performed less than 1 h after cell counting further
avoiding any reduction in viability.
IFNβ has been intensively studied in several models
(21,42,43) with abundant evidence of its
anticancer activity, namely, immune system stimulation,
angiogenesis, and malignant cell inhibition using S-phase
accumulation (44–46). In the present study, IFNβ
treatment dealt relatively poor tumor-reducing activity while
administered alone, as it exhibited only 25% of survival rate.
Notably, combined treatment with sTRAIL showed a 62.5% survival
rate, which surpasses both single-gene treatments (sTRAIL and
IFNβ), suggesting an additive effect.
IFNβ was unexpectedly ineffective. Interferons type
I have direct effects on T-cells; for instance, IFNβ-based
treatment is the first choice to treat multiple sclerosis by
modulation of T cell activity (47); therefore, malignant T cells are
inherently influenced by it. However, the specific nature of such
effects is unclear as current information is notably paradoxical
due to the complex effect IFNβ exerts on T cells (48). To elucidate this subject goes
beyond the aim of the present study. On the other hand, IFNβ can
induce apoptosis by upregulation and enhanced sensitivity of
endogenous TRAIL and upregulation of TRAIL-R1 and TRAIL-R2 in
cancer cell lines; moreover, there is evidence that IFNβ induces a
lengthened S-phase which also potencies TRAIL's apoptosis-inducing
effect in nasopharyngeal (49),
and cervical carcinomas (50).
It is well known that more than caspase cascade
activation, TRAIL can also modulate NF-κB signaling (51,52), which is controversial as NF-κB can
upregulate the expression of survival factors such as members of
the apoptosis family inhibitor and Bcl-xL (53). Conversely, TRAIL-dependent NF-κB
activation triggers apoptosis rather than survival in epithelial
cell lines and T cells (54,55). In addition, TRAIL sensitivity can
be upregulated by IFNs by overexpression of the TRAIL-R2 DR5
(56–58). IFNβ induces TRAIL-R2 expression in
ME-180 cells in the S phase (50), and in melanoma and breast cancer
cells, IFNβ induces apoptosis by an extrinsic pathway dependent on
TRAIL expression. TRAIL overexpression is a Stat1 dependent process
associated with activation of the TRAIL promoter (59).
Related to the MSC treatment, first, 50% survival
was observed, but in a second experiment, recurrence and tumor
growth after day 26 was identified. This finding showed that MSCs
present immunoregulatory properties, as they can inhibit T-cell
proliferation and migratory properties and are resistant to natural
killer cell-mediated cytolysis. Those attributes contribute to the
application of MSC-based delivery of therapeutic genes to solid
tumors (60). However, MSC could
improve cancer development by increasing the expression of
immunosuppressive cytokines such as IL-10 when activated with
signals from the tumor microenvironment (22,61,62). In addition, it was observed that
the saline solution (placebo) group showed a 25% survival rate,
which was reasonably expected considering all mice were
immunocompetent. Mice reached immune maturity, which could be
explained as a local immunological reaction due to intramuscular
solution inoculation that resulted in mild bleeding and muscle
inflammation, acting as a local adjuvant.
By day 41 PI, the surviving mice reduced the tumor
to the point of being imperceptive. The histological analysis of
the untreated mice showed almost no proper muscle tissue but
instead was completely saturated with lymphoid cells; by contrast,
treated mice showed seemingly healthy muscle tissue, with
histologically normal muscle fibers and few scattered lymphoid
cells. This finding bifurcates into two rather simplistic yet
actual possibilities: i) The main component of the tumor
microenvironment is tumor infiltrating lymphocytes of different
phenotypes as helper and cytotoxic T cells; these cells could be
normal, healthy lymphoid cells due to the immune surveillance
induced by the malignant cells, which in this scenario, are
seemingly depleted; or ii) Remaining transformed lymphoma cells
within the muscle tissue represent an ongoing immunological process
rather than a terminated one (63,64). Both scenarios could be elucidated
by finer cell characterization, a longer-term observation period to
detect relapse, and larger subject groups. In addition, the
combined treatment of sTRAIL and IFNβ did recover the tissue
morphology mainly due to extended tumor damage. Thus, the mechanism
of tissue repair after cancer elimination needs improvement.
In conclusion, the present findings demonstrated
that MSCs are effective anti-lymphoma vehicles, as non-treated mice
developed massive tumor masses and succumbed. Moreover, in this
model, sTRAIL had improved tumor-reducing effects, and IFNβ
enhanced it.
Acknowledgements
The authors would like to thank Dr Sergio Lozano
(Autonomous University of Nuevo Leon, Monterrey, Mexico) for
reviewing the manuscript.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
AGQR and CAGV contributed to designing, performing
experiments, analyis and discussion of results. HMR helped to
design of experiments, wrote, analyzed and corrected the
manuscript. SSF designed the experiments, performed literature
analysis and discussion of results. MCSC and AYLF helped in the
management and in the care with murine model. ASD helped in the
standardization of immunohistochemistry and with the software of
the fluorescence microscope. GPR helped with statistical analysis.
RMDOL contributed to designing of transgenes of lentivirus. JFI
edited the text, analyzed the results and reviewed the final
manuscript. ENGT contributed to the design of experiments, analysis
of the results, discussion and correction of the manuscript. AGQR,
CAGV and ENGT confirm the authenticity of all the raw data. All
authors have read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The present study was approved (approval no.
BI15-005) by the Scientific Research and Bioethics Committee and
the Institutional Animal Care Committees of the School of Medicine
of the Autonomous University of Nuevo Leon (Monterrey, México).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Urrutiocoechea A, Alemany R, Balart J,
Villanueva A, Viñals F and Capellá G: Recent advances in cancer: An
Overview. Curr Pharm Des. 16:3–10. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Herskovic A, Martz K, Al-Sarraf M,
Leichman L, Brindle J, Vaitkevicius V, Cooper J, Byhardt R, Davis L
and Emami B: Combined chemotherapy and radiotherapy compared with
radiotherapy alone in patients with cancer of the esophagus. N Engl
J Med. 326:1593–1598. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Singh P, Pandit S, Mokkapati VRSS, Garg A,
Ravikumar V and Mijakovic I: Gold nanoparticles in diagnostics and
therapeutics for human cancer. Int J Mol Sci. 19:19792018.
View Article : Google Scholar
|
|
4
|
Baetke SC, Lammers T and Kiessling F:
Applications of nanoparticles for diagnosis and therapy of cancer.
Br J Radiol. 88:201502072015. View Article : Google Scholar
|
|
5
|
Rupaimoole R and Slack FJ: MicroRNA
therapeutics: Towards a new era for the management of cancer and
other diseases. Nat Rev Drug Discov. 16:203–221. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Iorio MV and Croce CM: MicroRNA
dysregulation in cancer: Diagnostics, monitoring and therapeutics.
A comprehensive review. EMBO Mol Med. 4:143–159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ling H, Fabbri M and Calin GA: MicroRNAs
and other non-coding RNAs as targets for anticancer drug
development. Nat Rev Drug Discov. 176:139–148. 2013.
|
|
8
|
Loebinger M, Sage E, Davies D and Janes S:
TRAIL-expressing mesenchymal stem cells kill the putative cancer
stem cell population. Br J Cancer. 103:1692–1697. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu X, Hu J, Li Y, Cao W, Wang Y, Ma Z and
Li F: Mesenchymal stem cells expressing interleukin-18 inhibit
breast cancer in a mouse model. Oncol Lett. 15:6265–6274. 2018.
|
|
10
|
Matosevic S: Viral and nonviral
engineering of natural killer cells as emerging adoptive cancer
immunotherapies. J Immunol Res. 2018:40548152018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Friedenstein AJ, Gorskaja JF and Kulagina
NN: Fibroblast precursors in normal and irradiated mouse
hematopoietic organs. Exp Hematol. 4:267–274. 1976.
|
|
12
|
Pittenger MF, Mackay AM, Beck SC, Jaiswal
RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S and
Marshak DR: Multilineage potential of adult human mesenchymal stem
cells. Science. 284:143–147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Igura K, Zhang X, Takahashi K, Mitsuru A,
Yamaguchi S and Takahashi TA: Isolation and characterization of
mesenchymal progenitor cells from chorionic villi of human
placenta. Cytotherapy. 6:543–553. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Karp JM and Leng Teo GS: Mesenchymal stem
cell homing: The devil is in the details. Cell Stem Cell.
4:206–216. 2009. View Article : Google Scholar
|
|
15
|
Karnoub AE, Dash AB, Vo AP, Sullivan A,
Brooks MW, Bell GW, Richardson AL, Polyak K, Tubo R and Weinberg
RA: Mesenchymal stem cells within tumour stroma promote breast
cancer metastasis. Nature. 449:557–563. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shinagawa K, Kitadai Y, Tanaka M, Sumida
T, Kodama M, Higashi Y, Tanaka S, Yasui W and Chayama K:
Mesenchymal stem cells enhance growth and metastasis of colon
cancer. Int J Cancer. 127:2323–2333. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu JM, Jun ES, Bae YC and Jung JS:
Mesenchymal stem cells derived from human adipose tissues favor
tumor cell growth in vivo. Stem Cells Dev. 17:463–473. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pan G, O'Rourke K, Chinnaiyan A, Gentz R,
Ebner R, Ni J and Dixit V: The receptor for the cytotoxic ligand
TRAIL. Science. 276:111–113. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wong SHM, Kong WY, Fang CM, Loh HS, Chuah
LH, Abdullah S and Ngai SC: The TRAIL to cancer therapy: Hindrances
and potential solutions. Crit Rev Oncol Hematol. 143:81–94. 2019.
View Article : Google Scholar
|
|
20
|
Nicolini A, Carpi A and Rossi G: Cytokines
in breast cancer. Cytokine Growth Factor Rev. 17:325–337. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Studeny M, Marini FC, Champlin RE,
Zompetta C, Fidler IJ and Andreeff M: Bone marrow-derived
mesenchymal stem cells as vehicles for interferon-β delivery into
tumors. Cancer Res. 62:3603–3608. 2002.PubMed/NCBI
|
|
22
|
Gonzalez-Villarreal C, Rodriguez M and
Said-Fernández S: How desirable and undesirable features of naïve
or genetically reengineered mesenchymal stem cells are being
considered in preclinical or clinical assays. J BUON. 22:812–830.
2017.PubMed/NCBI
|
|
23
|
Qin JY, Zhang L, Clift KL, Hulur I, Xiang
AP, Ren BZ and Lahn BT: Systematic comparison of constitutive
promoters and the doxycycline-inducible promoter. PLoS One. 5:3–6.
2010. View Article : Google Scholar
|
|
24
|
Kim MH, Billiar TR and Seol DW: The
secretable form of trimeric TRAIL, a potent inducer of apoptosis.
Biochem Biophys Res Commun. 321:930–935. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yan C, Li S, Li Z, Peng H, Yuan X, Jiang
L, Zhang Y, Fan D, Hu X, Yang M and Xiong D: Human umbilical cord
mesenchymal stem cells as vehicles of CD20-specific TRAIL fusion
protein delivery: A double-target therapy against non-Hodgkin's
lymphoma. Mol Pharm. 10:142–151. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Stern B, Olsen LC, Tröße C, Ravneberg H
and Pryme IF: Improving mammalian cell factories: The selection of
signal peptide has a major impact on recombinant protein synthesis
and secretion in mammalian cells. Trends Cell Mol Biol. 10:19–24.
2007.
|
|
27
|
Yuan Z, Kolluri KK, Sage EK, Gowers KH and
Janes SM: Mesenchymal stromal cell delivery of full-length tumor
necrosis factor-related apoptosis-inducing ligand is superior to
soluble type for cancer therapy. Cytotherapy. 17:885–896. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Langford DJ, Bailey AL, Chanda ML, Clarke
SE, Drummond TE, Echols S, Glick S, Ingrao J, Klassen-Ross T,
Lacroix-Fralish ML, et al: Coding of facial expressions of pain in
the laboratory mouse. Nat Methods. 7:447–449. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Committee on Rodents, Institute of
Laboratory Animal Resources, Commission on Life Sciences and the
National Research Council, . Laboratory Animal Managent Rodents.
National Academies Press; Washington, D.C., USA: pp. 991996
|
|
30
|
Bankert RB, Egilmez NK and Hess SD:
Human-SCID mouse chimeric models for the evaluation of anti-cancer
therapies. Trends Immunol. 22:386–393. 2001. View Article : Google Scholar
|
|
31
|
Haouzi P: Murine models in critical care
research. Crit Care Med. 39:2290–2293. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Frese K and Tuveson D: Maximizing mouse
cancer models. Nat Rev Cancer. 7:654–658. 2007. View Article : Google Scholar
|
|
33
|
Boxall SA and Jones E: Markers for
characterization of bone marrow multipotential stromal cells. Stem
Cells Int. 2012:9758712012. View Article : Google Scholar
|
|
34
|
Darzi S, Werkmeister JA, Deane JA and
Gargett CE: Identification and characterization of human
endometrial mesenchymal stem/stromal cells and their potential for
cellular therapy. Stem Cells Transl Med. 5:1127–1132. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li Q, Hou H, Li M, Yu X, Zuo H, Gao J,
Zhang M, Li Z and Guo Z: CD73+ Mesenchymal stem cells
ameliorate myocardial infarction by promoting angiogenesis. Front
Cell Dev Biol. 9:6372392021. View Article : Google Scholar
|
|
36
|
Song N, Gao L, Qiu H, Huang C, Cheng H,
Zhou H, Lv S, Chen L and Wang J: Mouse bone marrow-derived
mesenchymal stem cells inhibit leukemia/lymphoma cell proliferation
in vitro and in a mouse model of allogeneic bone marrow
transplant. Int J Mol Med. 36:139–149. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Secchiero P, Zorzet S, Tripodo C,
Corallini F, Melloni E, Caruso L, Bosco R, Ingrao S, Zavan B and
Zauli G: Human bone marrow mesenchymal stem cells display
anti-cancer activity in SCID mice bearing disseminated
non-hodgkin's lymphoma xenografts. PLoS One. 5:e111402010.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tu E, Chia PZC and Chen W: TGFβ in T cell
biology and tumor immunity: Angel or devil? Cytokine Growth Factor
Rev. 25:423–435. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yuen GJ, Demissie E, Pillai S and Burnet
M: B lymphocytes and cancer: A love-hate relationship. Trends
Cancer. 2:747–757. 2017. View Article : Google Scholar
|
|
40
|
Soussi T: p53 Antibodies in the sera of
patients with various types of cancer: A review. Cancer Res.
60:1777–1788. 2000.PubMed/NCBI
|
|
41
|
Sharonov GV, Serebrovskaya EO, Yuzhakova
DV, Britanova OV and Chudakov DM: B cells, plasma cells and
antibody repertoires in the tumour microenvironment. Nat Rev
Immunol. 20:294–307. 2020. View Article : Google Scholar
|
|
42
|
Lei H, Furlong PJ, Jin HR, Mullins D,
Cantor R, Fraker DL and Spitz FR: AKT activation and response to
interferon-β in human cancer cells. Cancer Biol Ther. 4:709–715.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ren C, Kumar S, Chanda D, Kallman L, Chen
J, Mountz JD and Ponnazhagan S: Cancer gene therapy using
mesenchymal stem cells expressing interferon-β in a mouse prostate
cancer lung metastasis model. Gene Ther. 15:1446–1453. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dong Z, Greene G, Pettaway C, Dinney CP,
Eue I, Lu W, Bucana CD, Balbay MD, Bielenberg D and Fidler IJ:
Suppression of angiogenesis, tumorigenicity, and metastasis by
human prostate cancer cells engineered to produce interferon-beta.
Cancer Res. 59:872–879. 1999.PubMed/NCBI
|
|
45
|
Pestka S and Langer JA: Interferons and
their actions. Ann Rev Biochem. 56:727–777. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Qin X, Runkel L, Deck C, Dedios C and
Barsoum J: Interferon-beta induces S phase accumulation selectively
in human transformed cells. J Interf Cytokine Res. 17:355–367.
1997. View Article : Google Scholar
|
|
47
|
Kavrochorianou N, Markogiannaki M and
Haralambous S: IFN-β differentially regulates the function of T
cell subsets in MS and EAE. Cytokine Growth Factor Rev. 30:47–54.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Teige I, Liu Y and Issazadeh-Navikas S:
IFN-β Inhibits T cell activation capacity of central nervous system
APCs. J Immunol. 177:3542–3553. 2006. View Article : Google Scholar
|
|
49
|
Makowska A, Wahab L, Braunschweig T,
Kapetanakis NI, Vokuhl C, Denecke B, Shen L, Busson P and Kontny U:
Interferon beta induces apoptosis in nasopharyngeal carcinoma cells
via the TRAIL-signaling pathway. Oncotarget. 9:14228–14250. 2018.
View Article : Google Scholar
|
|
50
|
Vannucchi S, Chiantore MV, Fiorucci G,
Percario ZA, Leone S, Affabris E and Romeo G: TRAIL is a key target
in S-phase slowing-dependent apoptosis induced by interferon-β in
cervical carcinoma cells. Oncogene. 24:2536–2546. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang J, LeBlanc FR, Dighe SA, Hamele CE,
Olson TL, Feith DJ and Loughran TP: TRAIL mediates and sustains
constitutive NF-kB activation in LGL leukemia. Blood.
131:2803–2815. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mühlenbeck F, Schneider P, Bodmer JL,
Schwenzer R, Hauser A, Schubert G, Scheurich P, Moosmayer D,
Tschopp J and Wajant H: The tumor necrosis factor-related
apoptosis-inducing ligand receptors TRAIL-R1 and TRAIL-R2 have
distinct cross-linking requirements for initiation of apoptosis and
are non-redundant in JNK activation. J Biol Chem. 275:32208–32213.
2000. View Article : Google Scholar
|
|
53
|
Wang CY, Guttridge DC, Mayo MW and Baldwin
AS: NF-κB induces expression of the Bcl-2 homologue A1/Bfl-1 to
preferentially suppress chemotherapy-induced apoptosis. Mol Cell
Biol. 19:5923–5929. 1999. View Article : Google Scholar
|
|
54
|
Baetu TM, Kwon H, Sharma S, Grandvaux N
and Hiscott J: Disruption of NF-kappaB signaling reveals a novel
role for NF-kappaB in the regulation of TNF-related
apoptosis-inducing ligand expression. J Immunol. 167:3164–3173.
2001. View Article : Google Scholar
|
|
55
|
Shetty S, Brown Gladden J, Henson ES, Hu
X, Villanueva J, Haney N and Gibson SB: Tumor necrosis
factor-related apoptosis inducing ligand (TRAIL) up-regulates death
receptor 5 (DR5) mediated by NFkappaB activation in epithelial
derived cell lines. Apoptosis. 7:413–420. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Sedger LM, Shows DM, Blanton RA, Peschon
JJ, Goodwin RG, Cosman D and Wiley SR: IFN-gamma mediates a novel
antiviral activity through dynamic modulation of TRAIL and TRAIL
receptor expression. J Immunol. 163:920–926. 1999.
|
|
57
|
Meng RD and El-Deiry WS: P53-independent
upregulation of KILLER/DR5 TRAIL receptor expression by
glucocorticoids and interferon-gamma. Exp Cell Res. 262:154–169.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Shigeno M, Nakao K, Ichikawa T, Suzuki K,
Kawakami A, Abiru S, Miyazoe S, Nakagawa Y, Ishikawa H, Hamasaki K,
et al: Interferon-alpha sensitizes human hepatoma cells to
TRAIL-induced apoptosis through DR5 upregulation and NF-kappa B
inactivation. Oncogene. 22:1653–1662. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Choi EA, Lei H, Maron DJ, Wilson JM,
Barsoum J, Fraker DL, El-Deiry WS and Spitz FR: Stat1-dependent
induction of tumor necrosis factor-related apoptosis-inducing
ligand and the cell-surface death signaling pathway by interferon
beta in human cancer cells. Cancer Res. 63:5299–5307.
2003.PubMed/NCBI
|
|
60
|
Bao Q, Zhao Y, Niess H, Conrad C, Schwarz
B, Jauch KW, Huss R, Nelson PJ and Bruns CJ: Mesenchymal stem
cell-based tumor-targeted gene therapy in gastrointestinal cancer.
Stem Cells Dev. 21:2355–2363. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Taraballi F, Pasto A, Bauza G and Varner
C: Immunomodulatory potential of mesenchymal stem cell role in
diseases and therapies: A bioengineering prospective. J Immunol
Regen Med. 4:1000172019.
|
|
62
|
Monteran L and Erez N: The Dark side of
fibroblasts: Cancer-Associated fibroblasts as mediators of
immunosuppression in the tumor microenvironment. Front Immunol.
10:18352019. View Article : Google Scholar
|
|
63
|
Yu P and Fu YX: Tumor-infiltrating T
lymphocytes: Friends or foes? Lab Investig. 86:231–245. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Badalamenti G, Fanale D, Incorvaia L,
Barraco N, Listì A, Maragliano R, Vincenzi B, Calò V, Iovanna JL,
Bazan V and Russo A: Role of tumor-infiltrating lymphocytes in
patients with solid tumors: Can a drop dig a stone? Cell Immunol.
343:1037532019. View Article : Google Scholar
|