Introduction
At present, gastric cancer is an extremely common
malignancy. Although the incidence of the disease has been
declining for the past few decades, each year ∼798,000 people are
diagnosed with gastric cancer worldwide (9.9% of total cancer
cases) and 628,000 people succumb to the disease (12.1% of cancer
mortalities) (1). Globally, the
majority of cases of gastric cancer have been in East Asia, in
countries such as China, Japan and Korea. In China, the overall
prognosis for this cancer remains poor since the majority of
patients are diagnosed with advanced disease due to the lack of an
effective screening method or strategy. Consequently, the 1- and
5-year survival rates of patients suffering from gastric cancer are
extremely low (2). Although
adjuvant treatments, including chemotherapy and radiotherapy are
frequently used, their overall impact on the prognoses of these
patients remains limited and their adverse reactions frequently
affect the patients’ quality of life. A new therapeutic strategy
may be an urgent requirement for gastric cancer therapy.
Cellular immunotherapy against solid tumors has
become a promising treatment following a number of preclinical and
clinical trials. Of the numerous effector cells, cytokine-induced
killer (CIK) cells have received considerable attention. CIK cells
are expanded in vitro from peripheral blood mononuclear
cells (PBMCs) by the addition of interferon-γ (IFN-γ), interleukin
(IL)-2, IL-1 and a monoclonal antibody (McAb) against CD3 (3,4). CIK
cells are highly efficient cytotoxic effector cells with the
co-expression of CD3 and CD56 and NK activity (5). CIK cells have been used as effector
cells in the adoptive cell therapy against certain cancers and have
shown a promising effect (6,7).
Bispecific antibodies (BsAbs) are antibodies which
are made by chemical or biological methods and not observed in
natural conditions. They are typically designed to recognize a
specific epitope on effector cells and a target epitope on tumor
cells simultaneously. In comparison with conventional mAbs, BsAbs
are able to link immune effector cells to tumor cells directly and
have a more powerful ability to activate the immune-mediated
destruction of cancer cells. In 1994, Beun et al(8) reported that BsAbs directed to a
target-cell surface antigen and the T-cell-receptor
(TCR)/CD3-complex mediated the activation of T cells and the
induction of target-cell lysis by the activated cells. Therapeutic
strategies using BsAbs have been performed in a number of basal and
preclinical studies (9–13). However, a therapeutic strategy using
a combination of CIK cells and BsAbs against gastric cancer has not
yet been reported. The present study investigated the cytotoxic
activity of CIK cells targeted by epidermal growth factor receptor
(EGFR)/CD3 BsAbs against the gastric cancer cell line SGC7901 to
analyze the effect of this therapeutic strategy against gastric
cancer.
Materials and methods
Materials
EGFR McAb was purchased from Beijing Zhongshan
Biotechnology Co., Ltd. (Beijing, China). Rabbit anti-mouse HPR
(1:1,000; Dako, Copenhagen, Denmark) was used to recognize the
corresponding proteins. Common cell culture plates were purchased
from Orange Scientific (Braine-l’Alleud, Belgium). EGFR, CD3 and
CD56 McAbs were purchased from Biosynthesis Co., Ltd. (Beijing,
China). A total of 50 8–10-week-old female nude mice weighing 18–22
g (NU/NU; Iffa Credo, l’Arbresle, France) were purchased from
Biotechnology Co., Ltd. (Beijing, China) and housed in
self-contained filter-top cages (5 mice/cage). Approval of the
human blood collection protocol was obtained from The Beijing Blood
Center the study was approved by the Experimental Animal
Investigation Committee of The 309 Hospital of PLA, Beijing,
China.
Cell lines and culture
The human gastric adenocarcinoma cell line (SGC7901)
was obtained from the Shanghai Cell Research Institute of the
Chinese Scientific Academy (Shanghai, China) and stored and
transfer cultured in The Laboratory of the Gastroenterology
Department at The 309 Hospital. DMEM containing 10% calf serum, 100
IU/ml penicillin and 100 IU/ml streptomycin was used as a
conventional culture medium. The culture procedures were performed
at 37°C, 5% CO2 and saturation humidity.
Investigation of EGFR expression in the
SGC7901 cell line
The expression of EGFR in SGC7901 cells was detected
to determine whether the cell lines were suitable for use in the
study. An immunocytochemical assay and RT-PCR were performed and
the results showed that EGFR was expressed in the cell line.
Production of crosslinked anti-CD3 and
anti-EGFR BsAb
The anti-CD3 × anti-EGFR (EGFR/CD3) BsAb was
produced using the chemical heteroconjugation technique described
previously by Sen et al(14). Anti-CD3 McAb (1 mg/ml) in 50 mM NaCl
and 1 mM EDTA at pH 8.0 was added to a 5-fold molar excess of
Traut’s reagent (2-iminothiolane HCl; Pierce, Rockford, IL, USA) at
room temperature for 1 h. Anti-EGFR (1 mg/ml) in 0.1 mol/l sodium
phosphate and 0.15 mol/l NaCl (pH 7.2) were then added to a 4-fold
excess of sulfosuccinimidyl-4-(N-maleimidomethyl)
cyclohexane-1-carboxylate (Pierce) at room temperature for 1 h. The
two antibodies were purified on PD-10 columns (Pharmacia, Uppsala,
Sweden) in PBS to remove the unbound cross-linker. The cross-linked
McAbs were mixed immediately in equimolar ratios and conjugated at
4°C overnight. The product was analyzed using SDS-PAGE (2–15%
gradient; OWL Scientific, Woburn, MA, USA) and detected using
Coomassie blue staining.
Preparation of CIK cells
The CIK cells were prepared according to the method
in the study by Tita-Nwa et al(15). PBMCs were isolated from healthy
donors via the blood bank of Beijing. PBMCs were isolated by Ficoll
density gradient centrifuging, washed with RPMI-1640 and then
resuspended in RPMI-1640 containing 10% calf serum, 100 IU/ml
penicillin, 100 IU/ml streptomycin and INF-γ (500 U/ml). On day 1,
IL-2 (400 U/ml), IL-1α (100 U/ml) and anti-CD3 McAb (20 ng/ml) were
added. The culture media were changed every 2 days and the
concentration of the cells was regulated to 2×106/ml.
The cells were cultured for 21 days and detected using a flow
cytometry assay.
Binding of EGFR/CD3 BsAb to effector
cells and target cells
The binding of the EGFR/CD3 BsAb to SGC7901 cells
and CIK cells was detected by indirect immunofluorescence methods.
The binding of the EGFR/CD3 BsAb to SGC7901 cells was detected
using FITC-labeled CD3 antigen, while the binding of the EGFR/CD3
BsAb to CIK cells was detected with FITC-labeled EGFR antigen. The
binding rates were detected using a flow cytometry assay.
Cytotoxicity assays
51Cr release assays were performed to
detect the cytotoxicity of CIK cells to gastric cancer cells
targeted by EGFR/CD3 BsAb as described previously by Hoyle et
al(16) and Chan et
al(17). SGC7901 cells
(1×106) were labeled with 300 μCi sodium chromate
(Dupont-NEM, Boston, MA, USA). The labeled cells were then washed
twice with PBS, suspended in RPMI-1640 and plated in 96-well plates
at 1×104 cells per well, in triplicate. Effector cells
and antibodies were added to form groups as follows: group A:
EGFR/CD3 BsAb (20 μg/ml) + CIK cells (5×106/ml) +
SGC7901 cells (1×105/ml); group B: CD3 McAb (20
μg/ml) + CIK cells (5×106/ml) + SGC7901 cells
(1×105/ml); group C: EGFR McAb (20 μg/ml) + CIK
cells (5×106/ml) + SGC7901 cells (1×105/ml);
group D: CD3 McAb (20 μg/ml) + EGFR McAb (20 μg/ml) +
CIK cells (5×106/ml) + SGC7901 cells
(1×105/ml); and group E: CIK cells (5×106/ml)
+ SGC7901 cells (1×105/ml). CIK cells were added as
effector cells at effector:target (E:T) cell ratios of 50:1 and
incubated at 37°C, 5% CO2 for 4 h. The radioactivity of
the supernatant was measured in a γ counter (Cobra/AII; Packard
BioScience, Meriden, CT, USA). The lysis rate was calculated
according to the following formula: Lysis rate (%) = [(sample
release) − (spontaneous release) / (maximum release) − (spontaneous
release)] × 100.
Cytotoxicity assays at various E:T cell
ratios
The cytotoxicity of CIK cells targeted by the
EGFR/CD3 BsAb to gastric cancer cells was analyzed by
51Cr release assays at various E:T cell ratios. The
assays were performed as described previously. CIK cells were added
as effector cells at E:T cell ratios (1:1, 10:1, 20:1, 30:1, 40:1,
50:1, 60:1,70:1, 80:1, 90:1 and 100:1) and incubated at 37°C, 5%
CO2 for 4 h, then the lysis rates of the groups were
measured. The results reported are the mean values of three
independent experiments performed in triplicate and a curve was
established based on the mean lysis rates.
Cytotoxicity analysis in vivo
The cytotoxicity of CIK cells targeted by the
EGFR/CD3 BsAb to gastric cancer cells was investigated by
therapeutic experiments in a xenograft mouse model. Each mouse was
injected s.c. in the right flank with 5×106 SGC7901
cells in 0.2 ml PBS. After 10 days, the mice with tumors (diameter
≥0.5 cm) were selected for the therapeutic experiment. The selected
mice were assigned to 8 treatment groups (5 mice per group), as
follows: group A: EGFR/CD3 BsAb (1 mg) + CIK cells
(1×109/ml); group B: CD3 McAb (1 mg) + CIK cells
(1×109/ml); group C: EGFR McAb (1 mg) + CIK cells
(1×109/ml); group D: CD3 McAb (1 mg) + EGFR McAb (1 mg)
+ CIK cells (1×109/ml); group E: CD3 McAb (1 mg); group
F: EGFR McAb (1 mg) group G: EGFR/CD3 BsAb (1 mg); and group H:
0.9% NaCl i.v. injection alone (0.2 ml/injection). All antibodies
and effector cells were injected i.v. into the heat-dilated tail
vein and the first day of treatment was day 0. The treatments were
performed with twice weekly measurements of tumor dimensions and
estimation of tumor volumes (mm3) using the formula: V =
a × b2/2, where a is the length and b is the width. A
tumor growth curve of each group was established based on the mean
tumor volume at various times. At day 35, the tumors were dissected
and weighed, then the tumor inhibition rates were calculated by the
following formula: Tumor inhibition rate (%) = [(mean tumor weight
of group G) − (mean tumor weight of each experimental group) /
(mean tumor weight of group G)] × 100.
Investigation of EGFR and CD3 expression
in tumor tissues following treatment
The expression levels of EGFR and CD3 in the tumor
tissues of the xenograft mice following treatment were detected to
determine whether the effector cells were able to migrate into the
tumor tissues. The tumors were dissected and embedded in paraffin.
Tissue sections of 5-μm thickness were cut from
paraffin-embedded tissue blocks, then H&E staining and
immunohistochemical assays were performed.
Statistical analysis
In present study, the data are expressed as mean ±
SD. One-way ANOVA test and the Student’s t-test were performed. All
data were analyzed with the SPSS 11.0 (SPSS Inc., Chicago, IL, USA)
statistical software package. P<0.05 was considered to indicate
a statistically significant difference.
Results
Expression of EGFR in SGC7901 cells
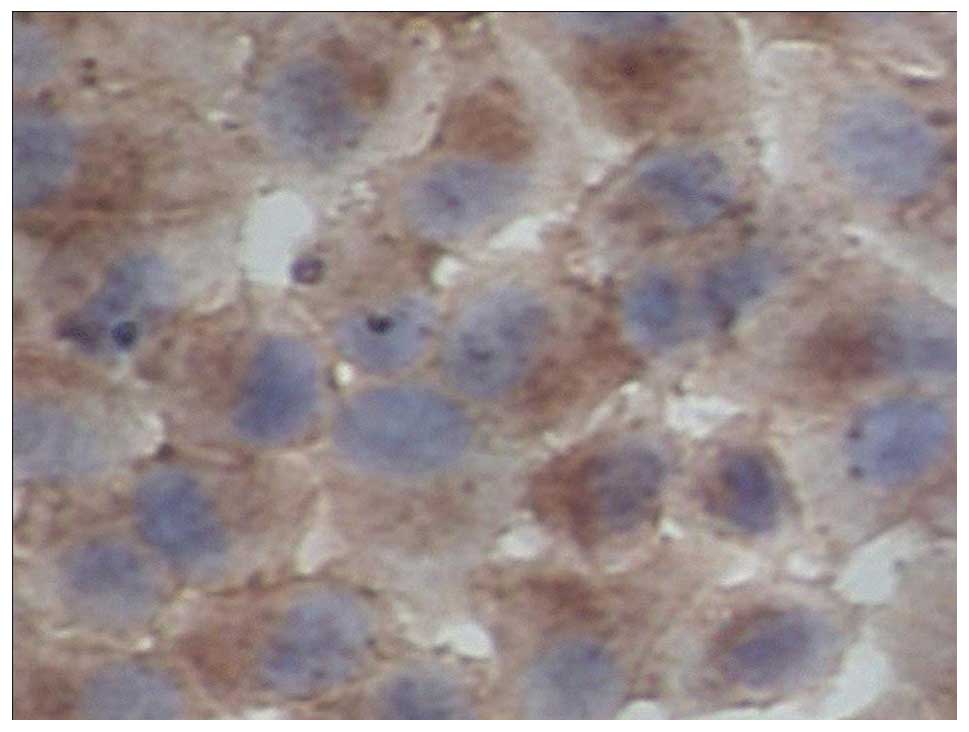
The expression of EGFR in the gastric adenocarcinoma
cell strain SGC7901 was detected by RT-PCR and immunocytochemical
analysis. The results in the SGC7901 cells were positive. The
immunohistochemical assay showed that the expression of EGFR in
SGC7901 cells was mainly distributed in the cell membrane and
cytoplasm and there was no clear positive signal in the cell
nucleus (Figs. 1 and 2).
Investigation and characterization of CIK
cells
The populations of cells in expanded CIK cells were
analyzed using flow cytometric analysis at day 21, to evaluate the
cytotoxicity of the CIK cells. The results suggested that
populations of CD3+CD8+ T cells accounted for
∼64.9% and CD3+CD56+ T cells accounted for
∼33.2% (Fig. 3).
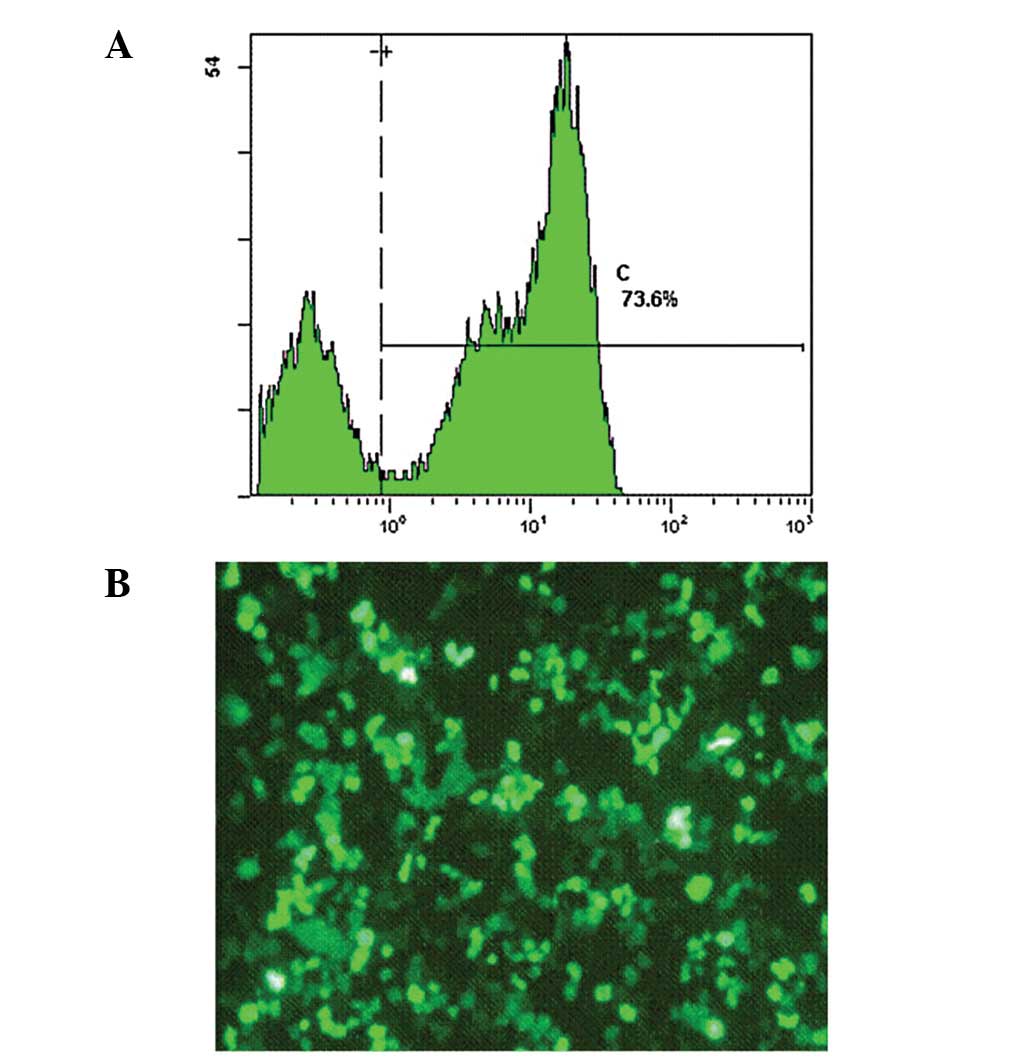
Binding of EGFR/CD3 BsAb
The binding activity of the EGFR/CD3 BsAb to SGC7901
and CIK cells was studied using an indirect immunofluorescence
method and the binding rates were analyzed with a flow cytometry
assay. The results showed that the EGFR/CD3 BsAb had a high binding
activity with regard to CIK and SGC7901 cells, which revealed the
dual affinity of the BsAb for the target and effector cells. The
binding rates of the EGFR/CD3 BsAb to SGC7901 and CIK cells were
73.6 and 71.2%, respectively (Figs.
4 and. 5).
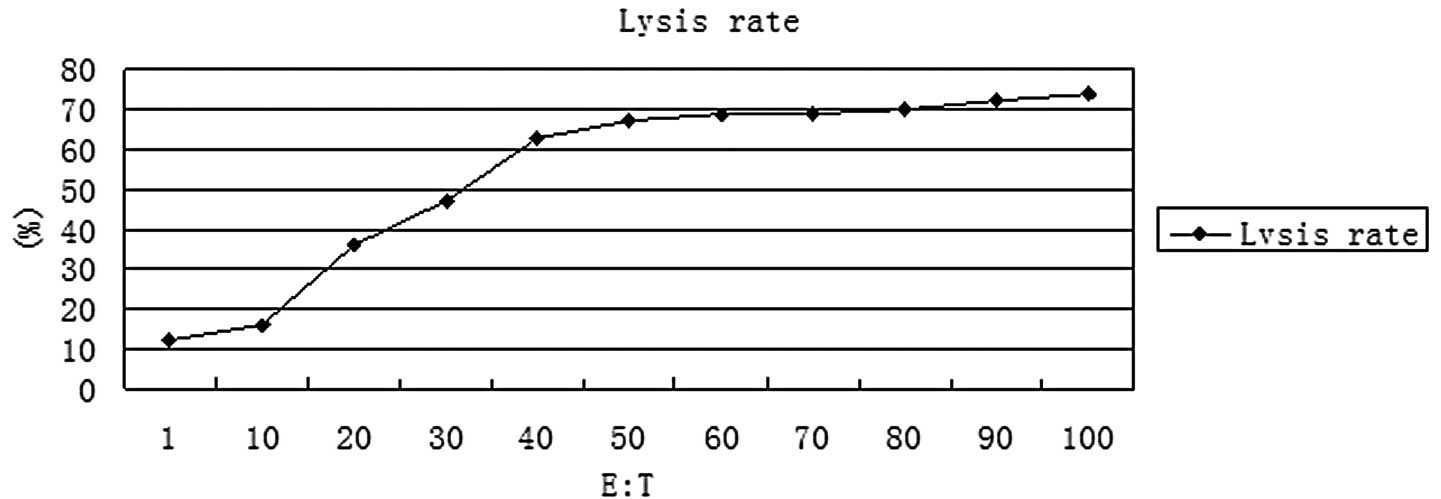
Cytotoxicity at various E:T cell
ratios
The cytotoxicity of CIK cells targeted by the
EGFR/CD3 BsAb at various E:T cell ratios was tested by
51Cr release assays. The results of the assays are shown
in Fig. 6. At an E:T cell ratio of
∼50:1, CIK cells targeted by the EGFR/CD3 BsAb exhibited
considerable cytotoxicity. The cytotoxicity of CIK cells did not
increase significantly when the E:T cell ratio increased beyond
50:1.
Enhancement of CIK cell cytotoxicity by
EGFR/CD3 BsAb
The cytotoxicity of CIK cells directed by the
EGFR/CD3 BsAb was tested by a cell lysis assay and compared with
that of CIK cells directed by EGFR or CD3 McAbs. The results of the
assay are shown in Fig. 7. At an
E:T cell ratio of 50:1, CIK cells directed by EGFR/CD3 BsAb
exhibited a cell lysis rate of 71.4%, which was higher than those
of CIK cells directed by EGFR or CD3 McAbs and the other control
groups (P<0.05).
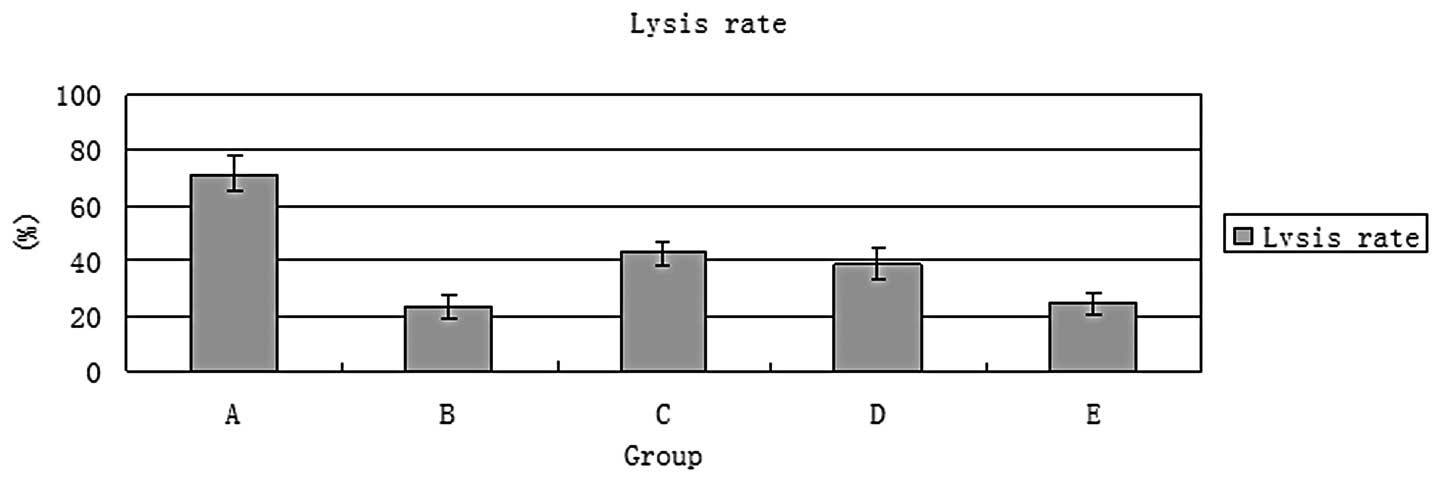 | Figure 751Cr release assays. At an
E:T cell ratio of 50:1, CIK cells directed by EGFR/CD3 BsAb had a
cell lysis rate of 71.4% which was higher than those of the other
groups. E:T, effector:target; CIK, cytokine-induced killer; EGFR,
epidermal growth factor receptor; BsAb, bispecific antibody; McAb,
monoclonal antibody; A, EGFR/CD3 BsAb (20 μg/ml) + CIK cells
(5×106/ml) + SGC7901 cells (1×105/ml); B, CD3
McAb (20 μg/ml) + CIK cells (5×106/ml) + SGC7901
cells (1×105/ml); C, EGFR McAb (20 μg/ml) + CIK
cells (5×106/ml) + SGC7901 cells (1×105/ml);
D, CD3 McAb (20 μg/ml) + EGFR McAb (20 μg/ml) + CIK
cells (5×106/ml) + SGC7901 cells (1×105/ml);
E, CIK cells (5×106/ml) + SGC7901 cells
(1×105/ml) . |
Effect of CIK cells directed by EGFR/CD3
BsAb on tumor growth
The antineoplastic activity of CIK cells directed by
the EGFR/CD3 BsAb was analyzed with tumor growth and tumor
reduction assays. The results are shown in Table I and Fig. 8. The EGFR/CD3 BsAb-redirected CIK
cells showed a significant enhancement of antitumor activity in the
mice. The results of the tumor growth curve assay showed that the
tumors in mice treated with CIK cells directed by the EGFR/CD3 BsAb
grew significantly slower than those in the other groups
(P<0.05; Fig. 8). The
administration of CIK cells directed by the EGFR/CD3 BsAb caused a
mean tumor reduction of 69.8%, which was higher than those of other
groups (P<0.05; Table I). The
administration of CD3 McAb alone without CIK cells had no
significant antineoplastic effect compared with that in the control
group treated with 0.9% NaCl. CD3 McAb did not enhance the
cytotoxicity of CIK cells.
 | Figure 8Tumor growth assay. The assay showed
that the tumors in mice treated with CIK cells directed by the
EGFR/CD3 BsAb grew significantly slower than those in the other
groups (P<0.05). EGFR, epidermal growth factor receptor; CIK,
cytokine-induced killer; BsAb, bispecific antibody; McAb,
monoclonal antibody; A, EGFR/CD3 BsAb (1 mg) + CIK cells
(1×109/ml); B, CD3 McAb (1 mg) + CIK cells
(1×109/ml); C, EGFR McAb (1 mg) + CIK cells
(1×109 1ml); D, CD3 McAb (1 mg) + EGFR McAb (1 mg) + CIK
cells (1×109/ml); E, CD3 McAb (1 mg); F, EGFR McAb (1
mg); G, EGFR/CD3 BsAb (1 mg); H: 0.9% NaCl i.v. injection alone
(0.2 ml/injection). |
 | Table ITumor inhibition rate of each group
(mean ± SD). |
Table I
Tumor inhibition rate of each group
(mean ± SD).
| Group | Tumor mass
(mg) | Tumor inhibition
rate (%) |
|---|
| A |
422.4±37.7abc |
69.8±7.8abc |
| B | 947.9±86.6a | 32.2±5.1 |
| C |
724.3±61.3ab |
48.2±6.5ab |
| D |
731.9±40.8ab |
47.6±5.5ab |
| E | 1357.6±114.2 | 2.8±1.3 |
| F | 1012.3±91.1a | 27.6±6.3a |
| G | 989.4±87.5a | 29.2±7.6a |
| H | 1397.4±131.6 | 0 |
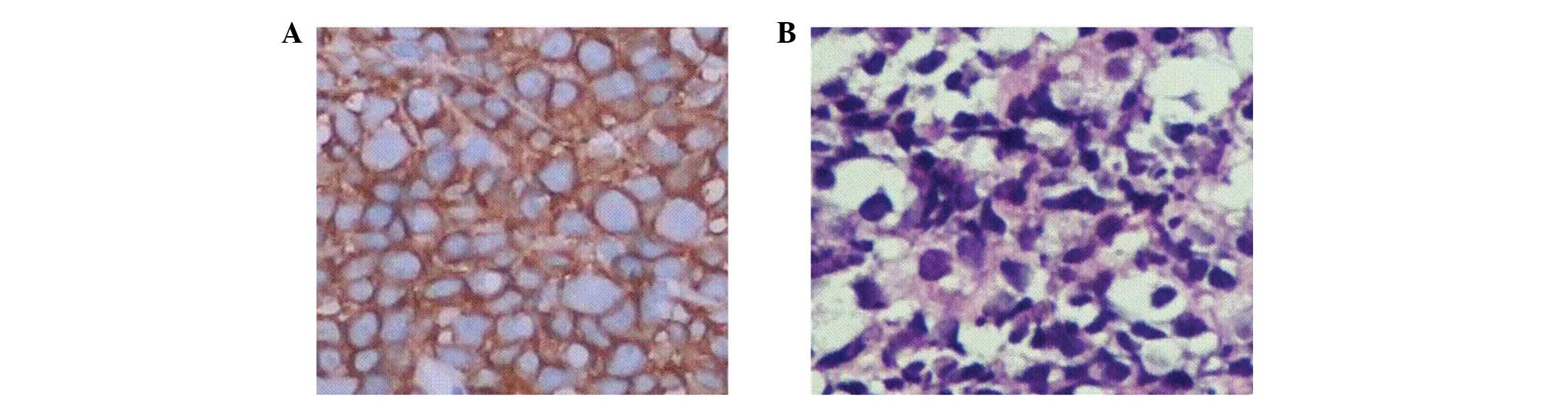
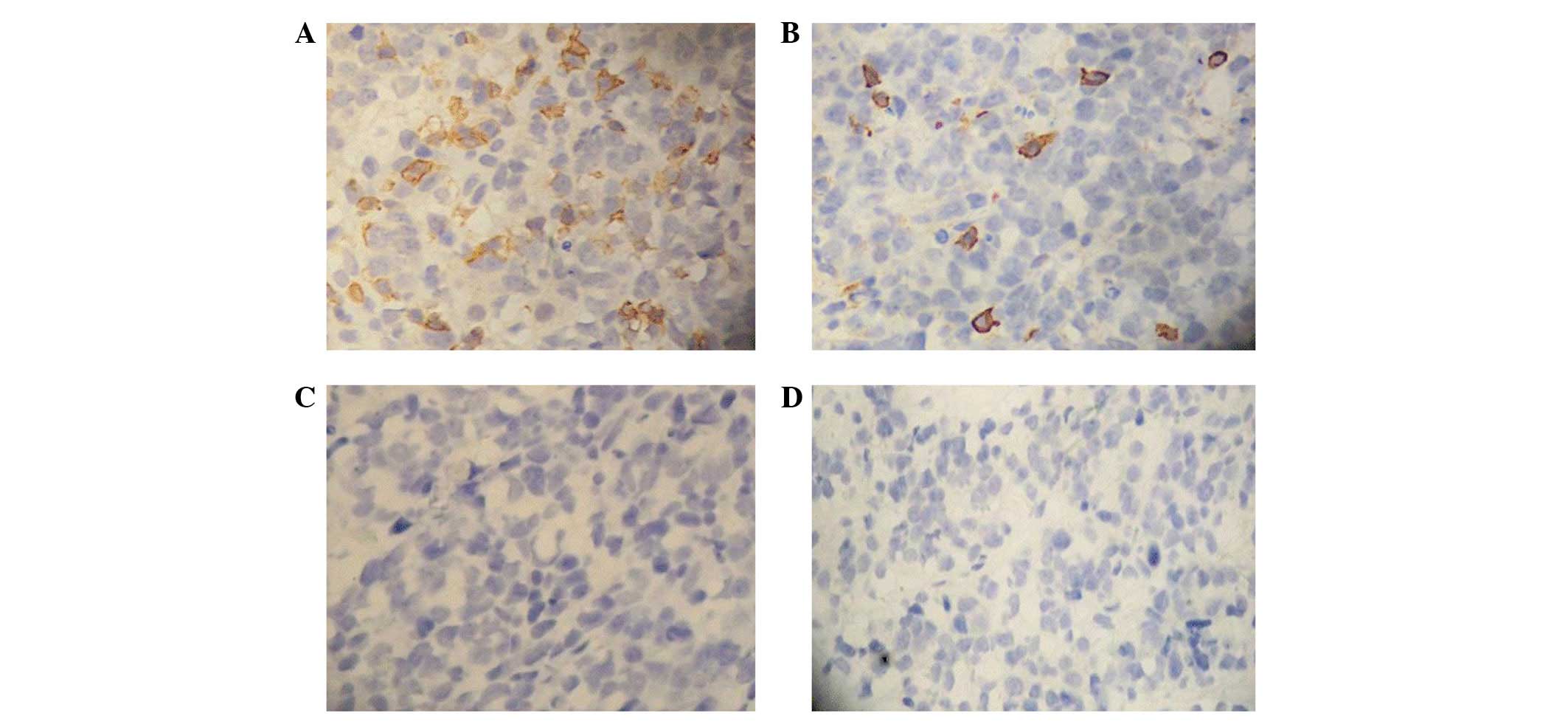
Expression of EGFR and CD3 in tumor
tissues following treatment
The immunohistochemical analysis revealed marked
positive expression of EGFR in the tumor tissues of all groups and
the positive signals were distributed in the cytoplasm and membrane
(Fig. 9). The immunohistochemical
assay showed that the expression of CD3 in the tumor tissues of the
mice treated with the EGFR/CD3 BsAb-redirected CIK cells was
increased significantly and the positive signals were mainly
distributed in the membrane and cytoplasm (Fig. 10). No positive signals were
observed in the tumor tissues of the mice treated with antibodies
alone without CIK cells. These results indicate that the EGFR/CD3
BsAb was able to direct CIK cells to tumor tissues and enhanced
their cytotoxicity to gastric cancer cells in vivo.
Discussion
Although the morbidity and mortality of gastric
cancer has been reduced, gastric cancer remains a common disease in
East Asia, with a poor prognosis and low survival rates. In China,
the prognosis of gastric cancer is poor since mass screening and
early diagnosis are not feasible. The therapeutic effects of
traditional strategies are not satisfactory as the pathogenesis and
molecular control of gastric cancer are poorly understood, although
the involvement of a number of affecting factors has been proposed.
Among these factors, cellular immune defects are common in patients
with gastric adenocarcinoma, as well as other malignancies, which
may have diverse effects in regulating critical antitumor
functions, including immunological recognition and regulation and
cytotoxicity. Therefore, cellular immunotherapy has been proposed
to be an important adjunctive therapy for gastric cancer and has
yielded promising effects. As the most common type of effector
cell, CIK cells express the T cell marker CD3 and NK cell marker
CD56 and develop cytotoxic activity against various cancer cells,
including those of gastric cancer. To enhance cytotoxity and
antitumor specificity, certain artificial BsAbs have been used to
direct CIK cells to tumor cells in previous studies (18–20).
The EGFR/CD3 BsAb directs CIK cells to tumor cells
by connecting CD3 on effector cells with EGFR on tumor cells. It
has been demonstrated that CD3 is an important triggering molecule
that activates T cells (21–24).
However, EGFR McAb was able to block the EGFR and restrain tumor
cell growth in a number of preclinical and clinical studies
(25,26). Theoretically, the EGFR/CD3 BsAb
binds to EGFR and CD3 simultaneously, so it should not only
activate T cells/CIK cells, but also restrict tumor cell growth. In
the present study, a EGFR/CD3 BsAb was produced and the
cytotoxicity of CIK cells targeted by this BsAb to gastric cancer
cells was investigated.
The results of cytotoxicity assays suggested that
redirected lysis was increased significantly when CIK cells were
combined with the EGFR/CD3 BsAb. CD3 McAb did not enhance tumor
cell lysis compared with the other groups. It was observed in the
present study that cytotoxicity increased to some extent when CIK
cells were combined with EGFR McAb. With regard to the expression
of Fc receptors (CD16, CD32 or CD64) on CIK cells, EGFR McAb also
directed effector cells to EGFR positive target cells by antibody
dependent cellular cytotoxicity (ADCC). However, the ability of
EGFR McAb to enhance cytotoxicity was more limited compared with
that of the EGFR/CD3 BsAb. Tita-Nwa et al demonstrated that
CD19/CD5 bsAb enhanced the cytotoxicity of CIK cells against
CD19+ B cell lymphoma lines in vitro and
suggested that these results supported the experimental use of the
in vivo bsAb models (15).
Chan et al investigated the cell killing ability of CIK
cells against primary ovarian carcinoma cells with and without
BsAbs and revealed that a CA125/Her2 BsAb significantly enhanced
the cytotoxicity of CIK cells in primary ovarian cancer cells in a
mouse model (17). These results
provide more support for the role of BsAbs in enhancing the
cytotoxicity of CIK cells. No studies on this therapeutic strategy
for gastric cancer have been reported previously, so the present
study is the first to report that a EGFR/CD3 BsAb is able to
enhance the cytotoxicity of CIK cells to gastric cancer cells in
vitro and in vivo.
The results of the cytotoxicity assays in
vivo showed that the antitumor activity of CIK cells was
enhanced significantly by the EGFR/CD3 BsAb when compared with EGFR
or CD3 McAbs in a nude mouse model. The present results are in
agreement with those of other studies which used different BsAbs to
target T cells or CIK cells to malignant cells (27,28).
These findings have demonstrated the promising therapeutic effect
and clinical potential of this strategy in a number of malignant
tumors. Certain researchers (29)
have used bioluminescent imaging to serially observe the response
to therapy without the need for sacrificing the experimental
animals. We were unable to perform bioluminescent imaging due to a
lack of equipment, but we considered that it was possible to obtain
credible data using the present test methods.
The immunohistochemical assays revealed that EGFR
expression was common in tumor tissues and the positive signals
were distributed in the cell membrane and cytoplasm. EGFR is the
most important member of the EGFR family. The overexpression of
EGFR occurs in numerous human malignancies, including lung, breast,
colon and gastric carcinomas (30–32).
It is well known that EGFR is associated with malignant
transformation and tumorigenesis, so this molecule has been
regarded as an important target in numerous preclinical and
clinical studies. The overexpression of EGFR in gastric cancer has
been confirmed in a number of studies (33,34).
Certain EGFR McAbs have been generated and used against a variety
of malignancies. One such antibody, cetuximab, has yielded a
promising therapeutic effect in delaying gastric cancer progression
(35,36). Consequently, EGFR was selected as
the target in gastric cancer cells in the present study.
The present results showed that the positive
immunohistochemical staining of CD3 was distributed in the cell
membrane and cytoplasm of certain cells in the tumor tissues of
groups A, C and D. However, there were no clear positive signals in
the tumor tissues of the other groups. The CD3 positive cells in
the mice treated with EGFR/CD3 BsAb-redirected CIK cells were
significantly more abundant than those in the mice treated with
EGFR McAb-redirected CIK cells. The results suggested that the
EGFR/CD3 BsAb and EGFR McAb were able to direct CD3 positive CIK
cells to tumor cells and the ability of the EGFR/CD3 BsAb to target
CIK cells to tumor cells was higher than that of EGFR McAb.
Given the unfavorable prognosis of patients with
advanced gastric cancer, there is a marked impetus to investigate
new therapeutic strategies to improve the outcome for these
patients. The results of the present investigation demonstrated
that the EGFR/CD3 BsAb significantly enhanced the cytotoxic
activity of CIK cells to gastric cancer cells in vitro and
in vivo, so this BsAb may potentially aid cellular
therapy.
Acknowledgements
The authors would like to thank Dr
Haili Huang, Dr Gangshi Wang, Weidi You, Weihua Wang, et al,
for handling patient contacts, and the Fourth Military Medical
University of the PLA for providing the means to perform the
present study.
References
|
1
|
Parkin DM, Poisani P and Ferlay: Global
cancer statistics. CA Cancer J Clin. 49:33–64. 1999. View Article : Google Scholar
|
|
2
|
Stein HJ, Sendler A, Fink U and Siewert
JR: Multidisciplinary approach to esophageal and gastric cancer.
Surg Clin North Am. 80:659–682. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lu PH and Negrin RS: A novel population of
expanded human CD3+CD56+ cells derived from T
cells with potent in vivo antitumor activity in mice with severe
combined immunodeficiency. J Immunol. 153:1687–1696.
1994.PubMed/NCBI
|
|
4
|
Schmidt-Wolf IG, Lefterova P, Mehta BA, et
al: Phenotypic characterization and identification of effector
cells involved in tumor cell recognition of cytokine-induced killer
cells. Exp Hematol. 21:1673–1679. 1993.PubMed/NCBI
|
|
5
|
Märten A, Ziske C, Schöttker B, et al:
Interactions between dendritic cells and cytokine-induced killer
cells lead to an activation of both populations. J Immunother.
24:502–510. 2001.PubMed/NCBI
|
|
6
|
Hui D, Qiang L, Jian W, et al: A
randomized, controlled trial of postoperative adjuvant
cytokine-induced killer cells immunotherapy after radical resection
of hepatocellular carcinoma. Dig Liver Dis. 41:36–41. 2009.
View Article : Google Scholar
|
|
7
|
Li H, Wang C, Yu J, et al: Dendritic
cell-activated cytokine-induced killer cells enhance the anti-tumor
effect of chemotherapy on non-small cell lung cancer in patients
after surgery. Cytotherapy. 11:1076–1083. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Beun GD, van de Welde CJ and Fleuren GJ:
T-cell based cancer immunotherapy: direct or redirected tumor-cell
recognition? Immunol Today. 15:11–15. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fury MG, Lipton A, Smith KM, Winston CB
and Pfister DG: A phase-I trial of the epidermal growth factor
receptor directed bispecic antibody MDX-447 without and with
recombinant human granulocytecolony stimulating factor in patients
with advanced solid tumors. Cancer Immunol Immunother. 57:155–163.
2008. View Article : Google Scholar
|
|
10
|
Gall JM, Davol PA, Grabert RC, Deaver M
and Lum LG: T cells armed with anti-CD3 × anti-CD20 bispecific
antibody enhance killing of CD20+ malignant B cells and
bypass complement-mediated rituximab resistance in vitro. Exp
Hematol. 33:452–459. 2005.
|
|
11
|
Stanglmaier M, Faltin M, Ruf P,
Bodenhausen A, Schröder P and Lindhofer H: Bi20 (fBTA05), a novel
trifunctional bispecific antibody (anti-CD20 3 anti-CD3), mediates
efficient killing of B-cell lymphoma cells even with very low CD20
expression levels. Int J Cancer. 123:1181–1189. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wimberger P, Xiang W, Mayr D, Diebold J,
Dreier T, Baeuerle PA and Kimmig R: Efficient tumor cell lysis by
autologous, tumor-resident T-lymphocytes in primary ovarian cancer
samples by an EP-CAM-/CD3-bispecific antibody. Int J Cancer.
105:241–248. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Morecki S, Lindhofer H, Yacovlev E,
Gelfand Y, Ruf P and Slavin S: Induction of long-lasting antitumor
immunity by concomitant cell therapy with allogeneic lymphocytes
and trifunctional bispecific antibody. Exp Hematol. 36:997–1003.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sen M, Wankowski DM, Garlie NK, et al: Use
of anti-CD3 × anti-HER2/neu bispecific antibody for redirecting
cytotoxicity of activated T cells toward HER2/neu+
tumors. J Hematother Stem Cell Res. 10:247–260. 2001.
|
|
15
|
Tita-Nwa F, Moldenhauer G, Herbst M,
Kleist C, Ho AD and Kornacker M: Cytokine-induced killer cells
targeted by the novel bispecific antibody CD19xCD5 (HD37xT5.16)
efficiently lyse B-lymphoma cells. Cancer Immunol Immunother.
56:1911–1920. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hoyle C, Bangs CD, Chang P, et al:
Expansion of Philadelphia chromosome-negative CD3(+) CD56(+)
cytotoxic cells from chronic myeloid leukemia patients: in vitro
and in vivo efficacy in severe combined immunodeficiency disease
mice. Blood. 92:3318–33127. 1998.PubMed/NCBI
|
|
17
|
Chan JK, Hamilton CA, Cheung MK, Karimi M,
Baker J, Gall JM, Schulz S, Thorne SH, Teng NN, Contag CH, Lum LG
and Negrin RS: Enhanced killing of primary ovarian cancer by
retargeting autologous cytokine-induced killer cells with
bispecific antibodies: a preclinical study. Clin Cancer Res.
12:1859–1867. 2006. View Article : Google Scholar
|
|
18
|
Mehta BA, Schmidt-Wolf IG, Weissman IL and
Negrin RS: Two pathways of exocytosis of cytoplasmic granule
contents and target cell killing by cytokine-induced
CD3+ CD56+ killer cells. Blood. 86:3493–3499.
1995.PubMed/NCBI
|
|
19
|
Schmidt-Wolf GD, Negrin RS and
Schmidt-Wolf IG: Activated. T cells and cytokine-induced
CD3+CD56+ killer cells. Ann Hematol.
74:51–56. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schmidt-Wolf IG, Lefterova P, Johnston V,
Huhn D, Blume KG and Negrin RS: Propagation of large numbers of T
cells with natural killer cell markers. Br J Haematol. 87:453–458.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Canevari S, Stoter G, Arienti F, Bolis G,
Colnaghi MI, Di Re EM, Eggermont AM, Goey SH, Gratama JW, Lamers
CH, et al: Regression of advanced ovarian carcinoma by
intraperitoneal treatment with autologous T lymphocytes retargeted
by a bispecific monoclonal antibody. J Natl Cancer Inst.
87:1463–1469. 1995. View Article : Google Scholar
|
|
22
|
de Gast GC, Haagen IA, van Houten AA,
Klein SC, Duits AJ, de Weger RA, Vroom TM, Clark MR, Phillips J and
van Dijk AJ: CD8 T cell activation after intravenous administration
of CD3 × CD19 bispecific antibody in patients with non-Hodgkin
lymphoma. Cancer Immunol Immunother. 40:390–396. 1995.
|
|
23
|
Kroesen BJ, Bakker A, van Lier RA, et al:
Bispecific antibody-mediated target cell-specific costimulation of
resting T cells via CD5 and CD28. Cancer Res. 55:4409–4415.
1995.PubMed/NCBI
|
|
24
|
Marmé A, Strauss G, Bastert G, Grischke EM
and Moldenhauer G: Intraperitoneal bispecific antibody
(HEA125xOKT3) therapy inhibits malignant ascites production in
advanced ovarian carcinoma. Int J Cancer. 101:183–189.
2002.PubMed/NCBI
|
|
25
|
Jonker DJ, O’Callaghan CJ, Karapetis CS,
et al: Cetuximab for the treatment of colorectal cancer. N Engl J
Med. 357:2040–2048. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fakih MG, Wilding G and Lombardo J:
Cetuximab-induced hypomagnesemia in patients with colorectal
cancer. Clin Colorectal Cancer. 6:152–156. 2006. View Article : Google Scholar
|
|
27
|
Katzenwadel A, Schleer H, Gierschner D,
Wetterauer U and Elsässer-Beile U: Construction and in vivo
evaluation of an anti-PSA x anti-CD3 bispecific antibody for the
immunotherapy of prostate cancer. Anticancer Res. 20:1551–1555.
2000.PubMed/NCBI
|
|
28
|
de Leij L, Molema G, Helfrich W and
Kroesen BJ: Bispecific antibodies for treatment of cancer in
experimental animal models and man. Adv Drug Deliv Rev. 31:105–129.
1998.PubMed/NCBI
|
|
29
|
Verneris MR, Arshi A, Edinger M, Kornacker
M, Natkunam Y, Karami M, Cao YA, Marina N, Contag CH and Negrin RS:
Low levels of Her2/neu expressed by Ewing's family tumor cell lines
can redirect cytokine-induced killer cells. Clin Cancer Res.
11:4561–4570. 2005.PubMed/NCBI
|
|
30
|
Scambia G, Benedetti Panici P, Ferrandina
G, et al: Significance of epidermal growth factor receptor
expression in primary human endometrial cancer. Int J Cancer.
56:26–30. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mukohara T, Kudoh S, Yamauchi S, et al:
Expression of epidermal growth factor receptor (EGFR) and
downstream-activated peptides in surgically excised non-small-cell
lung cancer (NSCLC). Lung Cancer. 41:123–130. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xie FH and Wang RX: The research progress
of the relationship between the way of EGFR family’s signal
transfer and breast cancer. Int J Lab Med. 27:1004–1008. 2006.
|
|
33
|
Kameda T, Yasui W, Tsujino T, et al:
Tyrosine kinase activity of epidermal growth factor receptor in
human gastric carcinomas. Pathol Res Pract. 188:37–43. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yasui W, Hata T, Yokozaki H, et al:
Interaction between epidermal growth factor and its receptor in
progression of human gastric carcinoma. Int J Cancer. 41:211–217.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pinto C, Difabio F, Siena S, et al: Phase
II study of cetuximab in combination with FOLFIRI in patients with
untreated advanced gastric or gastroesophageal junction
adenocarcinoma (FOLCETUX study). Ann Oncol. 18:510–517. 2007.
View Article : Google Scholar
|
|
36
|
Han SW, Oh DY, Im SA, et al: Phase II
study and biomarker analysis of cetuximab combined with modified
FOLFOX6 in advanced gastric cancer. Br J Cancer. 100:298–304. 2009.
View Article : Google Scholar : PubMed/NCBI
|