Introduction
Gastric cancer is the most frequent type of cancer
and remains the second leading cause of cancer mortality worldwide
(1). It is estimated that 360,000
individuals succumb to gastric cancer each year in China. Gastric
cancer is a heterogeneous disease with various subtypes. In total,
>95% of all cancers of the stomach are adenocarcinomas (2). No effective targeting therapy is
available for gastric cancer, mainly due to a lack of complete
understanding of the molecular mechanisms underlying gastric cancer
development (3). Data are now
emerging on the potential affect of the disease subtype on the
treatment outcome. For example, HER2 amplification and
overexpression is far more prevalent in proximal/gastro-esophageal
junction (GEJ) adenocarcinoma than in diffuse gastric cancer. In an
exploratory analysis, these proximal/GEJ tumors appeared to be less
sensitive to bevacizumab therapy than the diffuse and distal
non-diffuse gastric cancers (4).
Thus, disease biology may affect patient outcomes with specific
treatments.
MicroRNAs (miRNAs) are ~17–24 nucleotides long and
act as negative regulators of gene expression by inhibiting mRNA
translation or promoting mRNA degradation (5,6). This
class of regulators has been described as playing a significant
role in a vast range of biological processes, including
proliferation, differentiation and apoptosis (7). Recent progress in cancer biology has
revealed that miRNAs are frequently deregulated in various types of
human cancer, indicating that they may play a role as a novel class
of oncogenes or tumor suppressor genes in gastric cancer (8). For example, aberrant miR-106a
expression was detected in gastric carcinoma tissues and it was
revealed that this may promote gastric carcinogenesis (9). miR-21 has been shown to be correlated
with Helicobacter pylori infection and gastric cancer
progression (10). These data
indicate that miRNAs are involved in gastric carcinogenesis via a
variety of patterns and pathways and that it may be possible to
manipulate miRNA to achieve therapeutic effects.
Previously, it was observed that miR-372 and miR-373
act as oncogenes in the tumorigenesis of human testicular germ cell
tumors (Tera-1 and 833KE cells), through the direct inhibition of
LATS2 expression (11). In
addition, miR-373 was noted to affect esophageal cancer cell growth
through the inhibition of LATS2 expression (12). Moreover, we previously reported that
miR-372 maintains oncogene characteristics by targeting tumor
necrosis factor, α-induced protein 1 (TNFAIP1), and that it
regulates AGS cell growth (13).
However, it is not clear whether miRNA-373 affects the growth of
gastric cancer cells through the inhibition of TNFAIP1 expression.
The present study demonstrated that miR-373 is upregulated in
gastric adenocarcinoma tissue and gastric carcinoma cell lines when
compared to normal gastric tissues. The relative quantification of
miR-373 in gastric cancer was 0.5243 times that of normal gastric
tissue (P<0.01). Moreover, the bioinformatics prediction and
experimental results indicated that miR-373 is able to target
TNFAIP1. The study further demonstrated that miR-373 induces the
cell proliferation of gastric cancer cells in vitro, via the
downregulation of TNFAIP1.
Materials and methods
Cell culture and transfection
The human gastric carcinoma (AGS, HGC-27 and GES-1)
and human embryonic kidney (HEK)293 cell lines were obtained from
the Cell Bank of the Chinese Academy of Sciences (Shanghai, China).
The cells were cultured in DMEM or F12 media (both from Gibco,
Carlsbad, CA, USA) that was supplemented with 10% fetal bovine
serum, 100 μg/ml streptomycin and 100 IU/ml penicillin, and
maintained at 37°C in a humidified atmosphere with 5%
CO2. All miRNA mimics and inhibitors that were used in
this study were purchased from GenePharma (Shanghai, China). All
transfections were carried out using Lipofectamine® 2000
(Invitrogen, Carlsbad, CA, USA), according to the manufacturer’s
instructions.
Human tissue samples
All human gastric adenocarcinoma and adjacent normal
tissue samples were obtained from the Department of Pathology at
Xiangya Hospital (Changsha, China). The samples were classified
according to the World Health Organization criteria that were
published in 2000 (14). Informed
consent for the use of the samples was obtained from all patients
prior to surgery. All patients signed consent forms approved by the
Committee on Human Rights in Research of the Ethics Committee at
the College of Life Science, Hunan Normal University (Changsha,
China).
RNA extraction and real-time
quantification of miRNA-373 and TNFAIP1
Total RNA was extracted using an
E.Z.N.A.® FFPE RNA isolation kit (Omega Bio-Tek, Inc.,
Norcross, GA, USA), according to the manufacturer’s instructions.
For miRNA detection, 2 μg small RNA was reverse transcribed to cDNA
using an miRNA First-Strand cDNA Synthesis kit (Invitrogen),
according to the manufacturer’s instructions. Quantitative (q)PCR
analyses for miR-373 and U6 were performed in triplicate with the
SYBR-Green PCR Master mix (Perkin-Elmer Applied Biosystems, Foster
City, CA, USA), according to the manufacturer’s instructions. U6
RNA was used to normalize expression. To detect the target genes, 2
μg total RNA was reverse transcribed to cDNA using oligo(dT)
primers and Moloney murine leukemia virus reverse transcriptase
(Promega, Madison, WI, USA). qPCR was used to determine the
expression levels of TNFAIP1 and miR-373 using the primers
presented in Table I. β-actin
levels were used to normalize expression. The data analysis was
performed using the 2−ΔΔCt method (15).
 | Table IOligonucleotides used in the
study. |
Table I
Oligonucleotides used in the
study.
| Name | Sequence (5′→3′) |
|---|
| pmirGLO-TNFAIP1 |
| Forward | CGAGCTCGTGCTGCCTGGGTCTCTGC |
| Reverse | GCTCTAGAGCAGCTGCTCTGTCGGATGTTT |
|
pmirGLO-TNFAIP1-mut1 |
| Forward | CCTGCTCATGTCTGGAGAC |
| Reverse | CAGACATGAGCAGGGGCAA |
|
pmirGLO-TNFAIP1-F2 |
TGTGCAGAAGGGCTACTGC |
|
pmirGLO-TNFAIP1-R2 |
GCAGTAGCCCTTCTGCACA |
|
pmirGLO-TNFAIP1-mut2 |
| Forward | ATTCTCATGTACATGACAATAAG |
| Reverse | TCAGTACAACATGAGTTAAAGAA |
| PCMV-TNFAIP1-CDS |
| Forward | AGTCGACGATGTCAGGGGACACCTGT |
| Reverse | GGGTACCTCAGTCACGATGAGTGGA |
|
PCMV-TNFAIP1-3′UTR |
| Forward | AGTCGACGATGTCAGGGGACACCTGT |
| Reverse | GCTCTAGAGCAGCTGCTCTGTCGGATGTTT |
|
pCMV-TNFAIP1-3′UTRMT1 |
| Forward | CCTGCTCATGTCTGGAGAC |
| Reverse | CAGACATGAGCAGGGGCAA |
|
pCMV-TNFAIP1-3′UTRMT2 |
| Forward | ATTCTCATGTACATGACAATAAG |
| Reverse |
TCAGTACAACATGAGTTAAAGAA |
| RT-TNFAIP1 |
| Forward |
GCACTTTGGGCACCATTTTGA |
| Reverse |
CGGTTCTGAGGGAGGGTGAT |
| U6 |
TGCGGGTGCTCGCTTCGGCAGC |
| β-actin |
| Forward |
CCTGTACGCCAACACAGTGC |
| Reverse |
ATACTCCTGCTTGCTGATCC |
Construction of TNFAIP1 3′ untranslated
region (UTR)reporter plasmid
The 3′UTR of TNFAIP1 was amplified from HeLa cDNA
using RT-PCR and was inserted into the 3′-end of the firefly
luciferase gene of the dual-luciferase miRNA target expression
vector pmirGLO (Promega) between the SacI and XbaI
sites. Equally, the TNFAIP1-3′UTR mutant vectors, which contained
mutated miR-373 binding sites, were cloned to the pmirGLO between
the same sites. Similarly, the following TNFAIP1 expression vectors
were obtained from HeLa cDNA: pCMV-TNFAIP1, which did not contain
the 5′ and 3′UTR of TNFAIP1; pCMV-TNFAIP1-3′UTR, which contained
the 3′UTR of TNFAIP1; pCMV-TNFAIP1-3′UTRMT, which contained the
3′UTR mutation sites of TNFAIP1. The cells were transfected at 70%
confluence using Lipofectamine 2000 transfection reagent, according
to the manufacturer’s instruction. The primer sequences used for
the RT-PCR amplification are shown in Table I. All primers were synthesized by
Sangon Biotech (Shanghai, China).
Luciferase assay
The dual-luciferase reporter plasmids were
co-transfected with miRNA mimics (GenePharma) into HEK293 cells. At
24 h post-transfection, the cells were assayed for luciferase
activity using the Dual-Glo Luciferase assay system (Promega),
according to the manufacturer’s instructions. The firefly
luciferase activities were normalized to Renilla luciferase
activity. The firefly luciferase activity of the cells that were
transfected with miRNA mimics was represented as the percentage
activity relative to that of the cells that were transfected with
negative control miRNA mimics. For each transfection, the
luciferase activity was averaged from three replicate experiments
(16).
Western blot analysis
At 24 h post-transfection, the cells were harvested
and lysed in RIPA buffer [50 mM Tris-HCl (pH 7.2), 150 mM NaCl, 1%
(v/v) Triton X-100, 1% (w/v) sodium deoxycholate, 0.1% (w/v) SDS]
with protease inhibitors. Proteins were separated on 10%
SDS-polyacrylamide gel and transferred to PVDF membranes. A
PageRuler prestained protein ladder was used as a molecular marker.
Polyclonal TNFAIP1 (ProteinTech Group Inc., Chicago, IL, USA),
monoclonal c-myc (Santa Cruz Biotechnology, Santa Cruz, CA, USA)
and polyclonal β-actin (endogenous control; CWBIO, Beijing, China)
antibodies, were incubated with the blot overnight at 4°C. Goat
anti-mouse and goat anti-rabbit secondary antibodies (BA1010 and
BA1011; Wuhan Boster Biological Technology, Ltd., Wuhan, China)
were added after the membrane was washed three times with TBST. The
protein was detected using a HRP-conjugated secondary antibody and
a Chemilucent ECL Detection system (Millipore, Billerica, MA,
USA).
MTT assay
Briefly, the AGS or GES-1 cells were seeded in
24-well plates at 16,000 cells/well. At 24 h post-transfection, the
cells were incubated with 80 μl MTT at 37°C for another 4 h. The
medium was then removed and the precipitated formazan was dissolved
in 400 μl DMSO. Subsequent to being shaken for 15 min, the
absorbance at 420 nm was detected using a microplate
spectrophotometer.
Statistical analysis
All the results are presented as the mean ± standard
deviation from at least three separate experiments. The differences
among groups were analyzed using the double-sided Student’s t-test,
and P<0.05 was used to indicate a statistically significant
difference.
Results
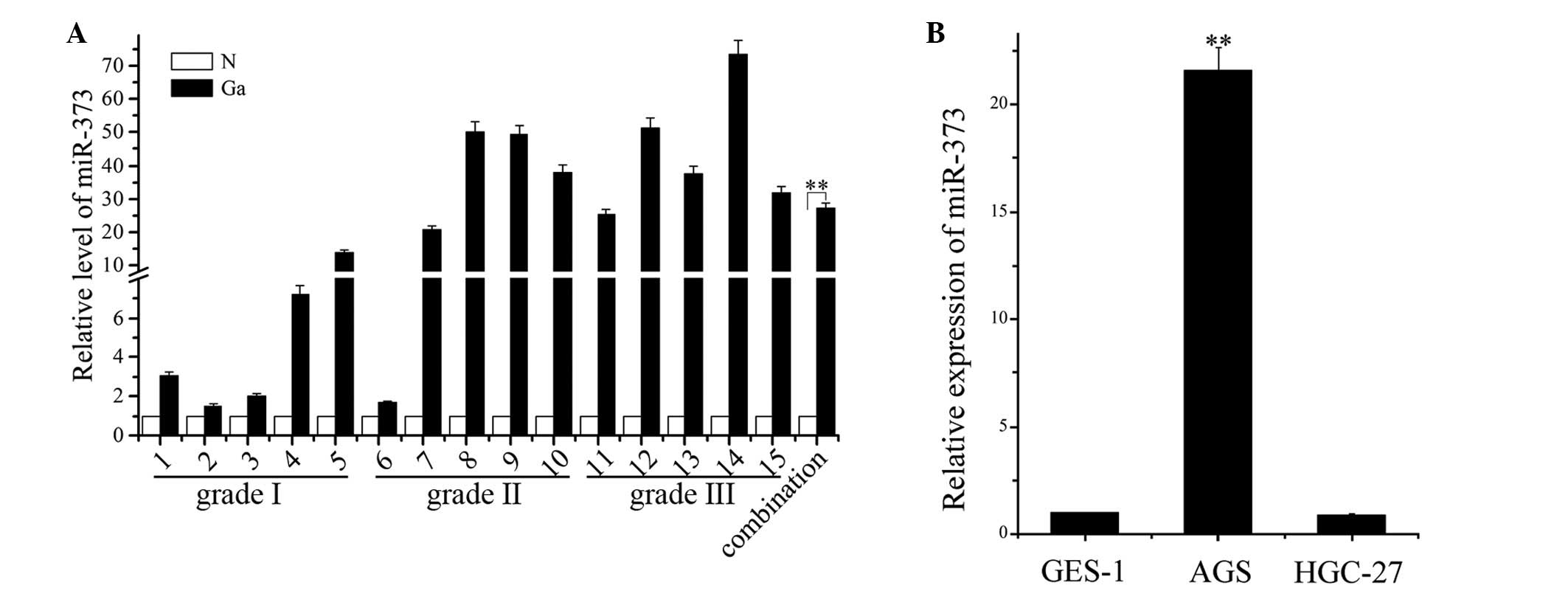
miR-373 is upregulated in human gastric
adenocarcinoma tissue and AGS cells
To confirm the expression level of miR-373 in
gastric adenocarcinoma tissue and normal gastric tissues, total RNA
was extracted from 15 gastric adenocarcinoma samples, consisting of
five grade I, five grade II and five grade III gastric
adenocarcinoma tissues, 15 matched normal tissues and three human
gastric carcinoma cell lines, and then qPCR was performed to
analyze the expression profile. Abnormal upregulation of miR-373
was observed in the gastric adenocarcinoma tissue and AGS cells,
while low expression was observed in the matched normal tissue,
indicating that miR-373 expression is upregulated in gastric cancer
(Fig. 1A). The relative
quantification of miR-373 in gastric cancer was 26.861 times that
of miR-373 in the normal gastric tissue (P<0.05). Previously,
Cho et al(17) analyzed the
expression of the miRNA-371–373 cluster in four human gastric
cancer cell lines, SNU-1, SNU-638, SNU-719 and AGS, and one normal
human gastric cell line, Hs677.sT, and identified that miR-372
expression was markedly elevated in the AGS cells. The present
study has shown the expression of miR-373 in three gastric cancer
cell lines to be consistent with a previous study (16) (Fig.
1B), indicating that this may play a possible role as an
oncogene.
miR-373 negatively regulates TNFAIP1
expression by targeting the 3′UTR
Two putative miR-373 binding sites were predicted to
have greater specificity to the TNFAIP1 3′UTR, ranging from
dinucleotide 170–192 bp or 570–593 bp, as predicted by four
algorithms (TargetScan, PicTar, miRanda and miRBase Target).
Moreover, this putative miRNA response element (MRE) was highly
conserved in vertebrates (Fig. 2A).
The present study validated whether the predicted MRE could be
recognized by the miR-372/miR-373 family using the dual-luciferase
vector pmirGLO. The predicted MRE, wild-type 3′UTR of TNFAIP1, was
cloned downstream of the firefly luciferase of the pmirGLO vector
and co-transfected with miR-372 or miR-373 mimics (double-stranded
processed miRNA) into HEK293 cells, which do not express miR-372 or
miR-373. The expression of miR-372 or miR-373 suppressed the
firefly luciferase activities of the MRE-containing 3′UTR of
TNFAIP1 (Fig. 2B). Three mutant
vectors of the 3′UTR of TNFAIP1 were also constructed: Two single
site mutation vectors, with one binding site with miR-373 and a
whole site mutation vector, with no binding site with miR-373.
Then, a luciferase reporter assay was performed with the mutated
constructs in the HEK293 cells. Once the conserved targeting
regions for miR-373 recognition had been mutated, the relative
luciferase activity of the TNFAIP1 gene was also restored. The
results demonstrated that the luciferase activity was elevated in
all the mutated vectors and that the relative luciferase activity
of the two single site mutation vectors increased by a lower
percentage compared with the whole site mutation vector. These
observations indicate that the predicted complementary sequence in
TNFAIP1 3′UTR is a functional element of miR-373.
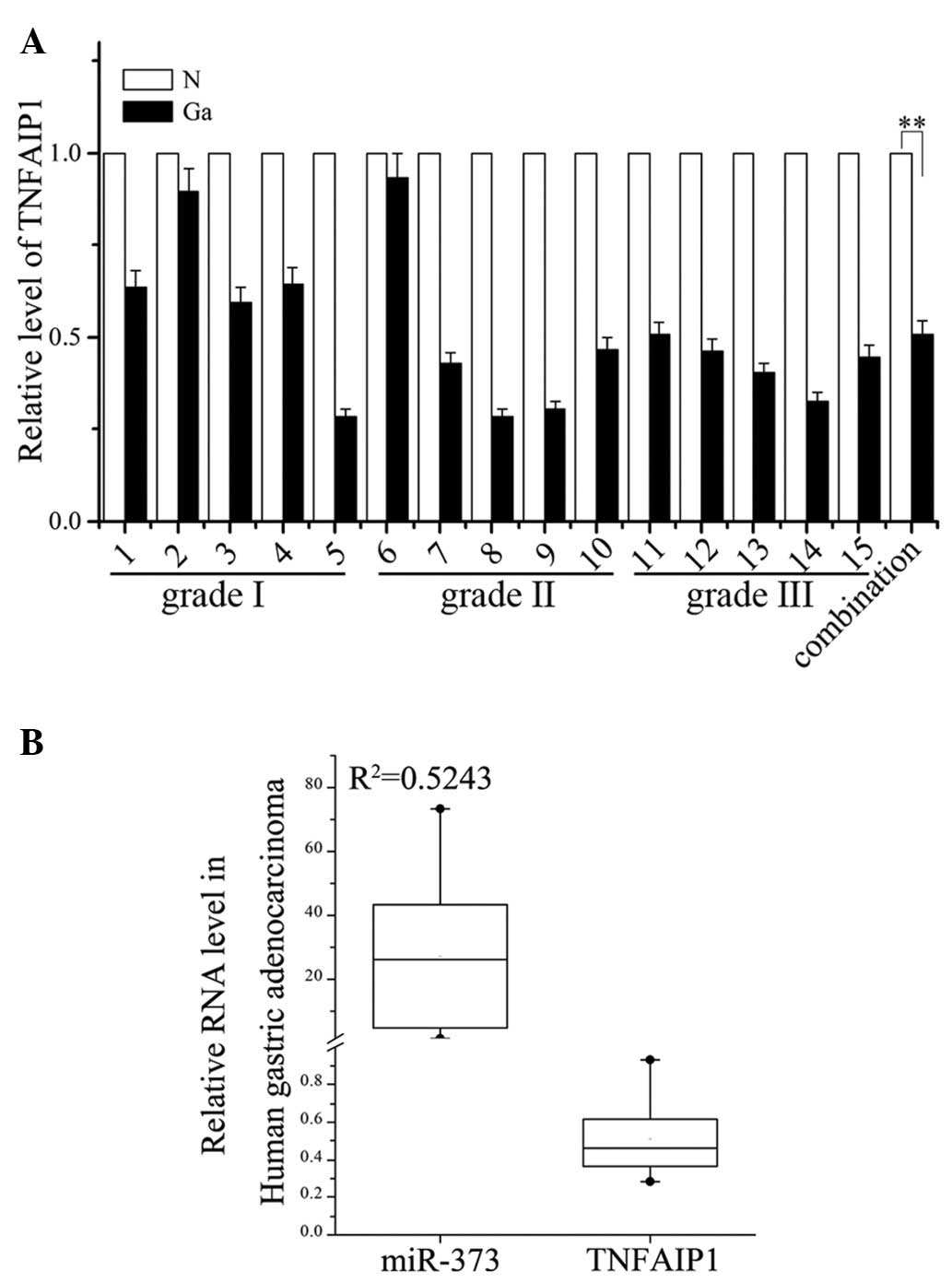
TNFAIP1 is downregulated in human gastric
adenocarcinoma tissue
The high expression of miR-373 was observed in human
gastric adenocarcinoma tissue and AGS cells, while the potential
expression levels of TNFAIP1 were not clear. Although TNFAIP1 has
been widely reported to be involved in tumorigenesis, there are no
studies on its role in human gastric adenocarcinoma tissue. To
investigate whether miR-373 affects the TNFAIP1 expression in
vivo, the mRNA levels of TNFAIP1 in gastric adenocarcinoma
tissue and normal gastric tissues were determined by RT-PCR. A
correlation was observed between the level of miR-373 and the mRNA
levels of TNFAIP1 in gastric adenocarcinoma tissue. This showed
that, in grades I, II and III gastric adenocarcinoma tissues, the
upregulation of miR-373 was more marked than that in 15 matched
normal tissues, while the mRNA levels of TNFAIP1 in all
investigated gastric adenocarcinoma tissues were lower than those
in matched normal tissues (Fig.
3A). High expression of miR-373 inversely correlated with lower
expression of TNFAIP1 in human gastric cancer (Fig. 3B). In conclusion, these results
showed an inverse correlation between miR-373 and TNFAIP1 mRNA
levels.
miR-373 represses endogenous TNFAIP1
expression
To examine the effect of miR-373 on endogenous
TNFAIP1 expression, miR-373 mimics were transfected into HGC-27
cells, which are known to express high levels of TNFAIP1 protein.
The enhanced expression of miR-373 in the HGC-27 cells
significantly decreased the amount of TNFAIP1 protein expression
compared with mock transfection (Fig.
4A). However, when transected with miR-373 inhibitors
(miR-373-ASO) in AGS cells, which are 2′OMe chemically modified,
single-stranded nucleic acids designed to specifically bind to and
inhibit endogenous miR-373 molecules, the observations indicated
that TNFAIP1 was negatively regulated by miR-373 at the protein
level (Fig. 4B).
miR-373 regulates growth of gastric cell
lines
Gain-of-function experiments are widely used for
functional studies of miRNAs. To further investigate whether the
correlation between miR-373 and gastric cell lines only existed in
gastric cancer cells, GES-1 cells, which are normal human gastric
mucosal cells, were introduced into the experiment. The relative
expression of miR-373 measured using RT-PCR demonstrated that
miR-373 expression levels in the AGS cells were markedly higher
than in the GES-1 cells, where the level was almost undetected. The
growth of gastric cell lines affected by miR-373 was then studied
using an MTT assay. To characterize the effects of miR-373 on cell
growth, the AGS cells were transfected with miR-373-ASO, and growth
was monitored using a cell growth curve, which indicated that the
knockdown of miR-373 inhibited cell growth. The growth rate of the
AGS cells transfected with the miR-373 inhibitor was lower than in
those transfected with the miR-NC, miR-NC-ASO and miR-373 mimics
(Fig. 5A). To determine whether
increasing miR-373 reversed the inhibition of cellular
proliferation observed due to overexpression, the GES-1 cells were
transfected with miR-373 mimics and compared with the results for
miR-373-ASO, miR-NC-ASO and miR-NC. An increase in proliferation
was observed in the cells transfected with miR-373 mimics as
compared with cells transfected with the control (Fig. 5B). These studies indicate that AGS
and GES-1 cell growth may be positively modulated by miR-373.
miR-373 induces AGS cell proliferation by
targeting TNFAIP1
In order to further investigate the effect of
miR-373 on the regulation of TNFAIP1 expression in the AGS cells,
AGS cells grown on a 6 cm plate were co-transfected with miR-373
mimics and TNFAIP1 expression vector pCMV-TNFAIP1,
pCMV-TNFAIP1-3′UTR and pCMV-TNFAIP1-3′UTRMT, respectively. Western
blotting assays revealed an inverse correlation between miR-373 and
TNFAIP1 protein levels (Fig. 5C).
MTT assays showed that the proliferation rate was reduced following
the overexpression of TNFAIP1, which was able to produce the same
effect as miR-373 inhibitor treatment in the AGS cells (Fig. 5A and D). This supports the
hypothesis that miR-373 induces the proliferation of gastric cancer
cell lines by negatively regulating the expression of TNFAIP1 at
the protein level.
Discussion
Documented evidence has demonstrated that miRNAs may
function as a novel class of tumorigenetic and tumor suppressing
genes. A more direct link between miRNA function and oncogenesis is
supported by studies examining the expression of miRNAs in clinical
samples (18–21). For example, the profiling of miRNA
expression showed that the majority of miRNAs are downregulated in
tumors compared to normal tissues, such as miR-128 in glioma
tissues (22) and miR-145 in human
breast cancer (23). However,
miR-17–92 is significantly increased in small-cell lung cancers and
human B-cell lymphomas and plays a key role in tumorigenesis
(24). Recent evidence suggests
that miR-373 is tumorigenetic in human reproductive system cancers
and human embryonic stem cells (ESCs) by targeting the tumor
suppressor LATS2 (25). In the
present study, the expression of miR-373 was detected in human
gastric adenocarcinoma tissue samples and gastric cancer cell lines
and observed to be upregulated compared to levels in normal gastric
tissues. This conclusion was established by results showing that
the downregulation of miR-373 expression by miR-373-ASO suppressed
the growth of AGS cells and increased apoptosis. Overexpression of
miR-373 in the HGC-27 cells increased cell growth. These findings
are in agreement with a previous report demonstrating that miR-373
has oncogenic properties (11).
Although a large number of miRNAs have been
discovered, only a few targets have been identified. The
identification of miRNA target genes remains a great challenge.
Computational algorithms showed >500 targets of miR-373.
Considering that miR-373 overexpression is able to increase the
proliferation of gastric cancer cells, several predicted target
genes associated with tumorigenesis or cell proliferation were
subjected to a luciferase reporter assay in the present study. The
results showed that, among these selected genes, TNFAIP1 was
negatively regulated by miR-373. These findings are in agreement
with a previous study demonstrating that miR-372 targets TNFAIP1.
To determine whether miR-373 has a direct effect on TNFAIP1
expression, miR-373 mimics or miR-373-ASO were transfected into the
HGC-27 or AGS cells to alter the level of miR-373. When the AGS
cells were transfected with miR-373 inhibitors (miR-373-ASO), which
are 2′OMe chemically modified, single-stranded nucleic acids
designed to specifically bind to and inhibit endogenous miR-373
molecules, the observations indicated that TNFAIP1 was negatively
regulated by miR-373 at the protein and mRNA levels. Conversely,
treatment with miR-373 mimics or control (miR-NC) was performed in
the HGC-27 cells. The results showed that the overexpression of
miR-373 markedly decreased the expression of TNFAIP1 at the protein
and mRNA levels. These observations indicate that TNFAIP1 is
negatively regulated by miR-373.
The expression levels of TNFAIP1 were measured in 15
pairs of human gastric adenocarcinoma tissue and adjacent normal
tissues using RT-PCR. The results showed that TNFAIP1 expression
levels were generally lower in the human gastric adenocarcinoma
tissues than in the matched normal gastric tissues, indicating that
TNFAIP1 expression is downregulated in gastric cancer. One notable
point observed in this study was that the upregulation of miR-373
and the downregulation of TNFAIP1 was all in grade I, II and III
gastric adenocarcinoma tissues. Moreover, a high expression of
miR-373 inversely correlated with a lower expression of TNFAIP1 in
these gastric adenocarcinoma tissues. Gastric cancer is a
heterogeneous disease, with subtypes in existence. There is,
however, a limitation within the present data interpretation with
regard to the correlated expression of miR-373 and TNFAIP1, since
human gastric adenocarcinoma tissue and normal gastric tissues were
used to detect their expression levels. Nevertheless, the stomach
is a mixture of different cell types and detection of miR-373 and
TNFAIP1 by other methods may be required. Whether miR-373 and
TNFAIP1 may be used as molecular markers for the diagnosis of
gastric adenocarcinoma requires more evidence.
TNFAIP1, also termed B12, is highly conserved in a
wide range of species, including humans, mice, rats (26) and C. elegans(27). A high expression of TNFAIP1 has been
observed in normal cell lines, while a low level of expression has
been detected in cancer cell lines (28). TNFAIP1 is an immediate-early
response gene of the endothelium induced by TNF-α and IL-6
(29). TNFAIP1 may play roles in
DNA synthesis, DNA repair, cell apoptosis and human diseases
(30). A previous study (28) demonstrated that the TNFAIP1 gene was
weakly expressed in several cancer-derived cell lines, while it was
expressed at a high level in normal cells and also modulated the
cellular proliferation rate. Conversely, reducing miR-373
expression using miRNA inhibitors greatly sensitized the cells to
apoptosis. miR-373 overexpression affected the cell growth and
correlated with the level of TNFAIP1. Thus, we conclude that
miRNA-373 accelerates gastric cancer cell differentiation by
negatively regulating TNFAIP1.
In conclusion, the present study established the
important role played by miR-373 in gastric cancer. Following the
observation of the elevated expression of miR-373 in human gastric
adenocarcinoma tissue samples and AGS cells, the study demonstrated
that: i) Overexpression of miR-373 is associated with accelerated
cell differentiation; ii) miR-373 directly targets and
downregulates the expression of TNFAIP1; and iii) the knockdown of
miR-373 by miRNA inhibitors recapitulates the differentiation
phenotype created by the overexpression of TNFAIP1. Collectively,
these findings demonstrate an oncogenic role for miR-373 in
controlling cell growth and apoptosis through the downregulation of
TNFAIP1.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant nos. 81071696, 31071150 and
81071656), the Project of Hunan Provincial Masters Innovation Fund
(CX2012B227) and the Project of Changsha Science and Technology
Plan (K1109006-31 and K1207014-31).
References
|
1
|
Kamangar F, Dores GM and Anderson WF:
Patterns of cancer incidence, mortality, and prevalence across five
continents: defining priorities to reduce cancer disparities in
different geographic regions of the world. J Clin Oncol.
24:2137–2150. 2006. View Article : Google Scholar
|
|
2
|
Catalano V, Labianca R, Beretta GD, Gatta
G, de Braud F and Van Cutsem E: Gastric cancer. Crit Rev Oncol
Hematol. 71:127–164. 2009. View Article : Google Scholar
|
|
3
|
Gao C, Zhang Z, Liu W, Xiao S, Gu W and Lu
H: Reduced microRNA-218 expression is associated with high nuclear
factor kappa B activation in gastric cancer. Cancer. 116:41–49.
2010.PubMed/NCBI
|
|
4
|
Shah MA, Jhawer M, Ilson DH, et al: Phase
II study of modified docetaxel, cisplatin, and fluorouracil with
bevacizumab in patients with metastatic gastroesophageal
adenocarcinoma. J Clin Oncol. 29:868–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mukherji S, Ebert MS, Zheng GX, Tsang JS,
Sharp PA and van Oudenaarden A: MicroRNAs can generate thresholds
in target gene expression. Nat Genet. 43:854–859. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pichinuk E, Broday L and Wreschner DH:
Endogenous RNA cleavages at the ribosomal SRL site likely reflect
miRNA (miR) mediated translational suppression. Biochem Biophys Res
Commun. 414:706–711. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang B, Pan X, Cobb GP and Anderson TA:
microRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xiao B, Guo J, Miao Y, et al: Detection of
miR-106a in gastric carcinoma and its clinical significance. Clin
Chim Acta. 400:97–102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Z, Li Z, Gao C, et al: miR-21 plays
a pivotal role in gastric cancer pathogenesis and progression. Lab
Invest. 88:1358–1366. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Voorhoeve PM, le Sage C, Schrier M, et al:
A genetic screen implicates miRNA-372 and miRNA-373 as oncogenes in
testicular germ cell tumors. Cell. 124:1169–1181. 2006. View Article : Google Scholar
|
|
12
|
Lee KH, Goan YG, Hsiao M, et al:
MicroRNA-373 (miR-373) post-transcriptionally regulates large tumor
suppressor, homolog 2 (LATS2) and stimulates proliferation in human
esophageal cancer. Exp Cell Res. 315:2529–2538. 2009. View Article : Google Scholar
|
|
13
|
Zhou C, Li X, Zhang X, et al: microRNA-372
maintains oncogene characteristics by targeting TNFAIP1 and affects
NFκB signaling in human gastric carcinoma cells. Int J Oncol.
42:635–642. 2013.PubMed/NCBI
|
|
14
|
Hamilton SR and Aaltonen LA: Pathology and
genetics Tumours of the digestive system. WHO Classification of
Tumours. Kleihues P and Sobin LH: 2. IARC Press; Lyon: 2000
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
|
|
16
|
Zeng Y and Cullen BR: Sequence
requirements for micro RNA processing and function in human cells.
RNA. 9:112–123. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cho WJ, Shin JM, Kim JS, et al: miR-372
regulates cell cycle and apoptosis of ags human gastric cancer cell
line through direct regulation of LATS2. Mol Cells. 28:521–527.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chan SH, Wu CW, Li AF, Chi CW and Lin WC:
miR-21 microRNA expression in human gastric carcinomas and its
clinical association. Anticancer Res. 28:907–911. 2008.PubMed/NCBI
|
|
19
|
Lai KW, Koh KX, Loh M, et al:
MicroRNA-130b regulates the tumour suppressor RUNX3 in gastric
cancer. Eur J Cancer. 46:1456–1463. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Motoyama K, Inoue H, Nakamura Y, Uetake H,
Sugihara K and Mori M: Clinical significance of high mobility group
A2 in human gastric cancer and its relationship to let-7 microRNA
family. Clin Cancer Res. 14:2334–2340. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ueda T, Volinia S, Okumura H, et al:
Relation between microRNA expression and progression and prognosis
of gastric cancer: a microRNA expression analysis. Lancet Oncol.
11:136–146. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Chao T, Li R, et al: MicroRNA-128
inhibits glioma cells proliferation by targeting transcription
factor E2F3a. J Mol Med (Berl). 87:43–51. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Spizzo R, Nicoloso M, Lupini L, et al:
miR-145 participates with TP53 in a death-promoting regulatory loop
and targets estrogen receptor-alpha in human breast cancer cells.
Cell Death Differ. 17:246–254. 2010. View Article : Google Scholar
|
|
24
|
Hayashita Y, Osada H, Tatematsu Y, et al:
A polycistronic microRNA cluster, miR-17–92, is overexpressed in
human lung cancers and enhances cell proliferation. Cancer Res.
65:9628–9632. 2005.
|
|
25
|
Martinez NJ and Gregory RI: MicroRNA gene
regulatory pathways in the establishment and maintenance of ESC
identity. Cell Stem Cell. 7:31–35. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou J, Hu X, Xiong X, et al: Cloning of
two rat PDIP1 related genes and their interactions with
proliferating cell nuclear antigen. J Exp Zool A Comp Exp Biol.
303:227–240. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Link CD, Taft A, Kapulkin V, et al: Gene
expression analysis in a transgenic Caenorhabditis elegans
Alzheimer’s disease model. Neurobiol Aging. 24:397–413. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang L, Zhou A, Li H, et al: Expression
profile in the cell lines of human TNFAIP1 gene. Yi Chuan.
28:918–922. 2006.(In Chinese).
|
|
29
|
Wolf F, Marks R, Sarma V, et al:
Characterization of a novel tumor necrosis factor-alpha-induced
endothelial primary response gene. J Biol Chem. 267:1317–1326.
1992.PubMed/NCBI
|
|
30
|
Yang L, Liu N, Hu X, et al: CK2
phosphorylates TNFAIP1 to affect its subcellular localization and
interaction with PCNA. Mol Biol Rep. 37:2967–2973. 2010. View Article : Google Scholar : PubMed/NCBI
|