Introduction
MicroRNAs (miRNAs), which are a conserved class of
short, non-coding, endogenous RNAs comprising ~22 nucleotides, are
involved in post-transcriptional gene regulation by inhibiting
translation or cleaving specific target mRNAs (1). It has been indicated that miRNAs play
important regulatory roles in various biological processes, such as
cell proliferation, developmental timing, apoptosis, metabolism and
carcinogenesis (2). miRNAs have
been classified as oncogenes or tumor suppressor genes according to
their function in carcinogenesis and ectopic expression in tumors
(3). For example, miR-27a is
overexpressed and functions as an oncogene in the development of
renal cell carcinoma (RCC) (4), and
Let-7a may inhibit cell growth by targeting c-myc in RCC (5).
MicroRNA-34a (miR-34a) is a direct target of p53
(6) and is commonly downregulated
in several types of cancer. For example, miR-34a was downregulated
in 10 out of 10 prostate cancer tissues (7). Gallardo et al (8) demonstrated that miR-34a expression was
significantly reduced in non-small-cell lung cancer compared with
normal tissues. The ectopic expression of miR-34a may induce G1
arrest (9–11), apoptosis and senescence in tumors
(6,12,13).
Moreover, miR-34a directly regulates numerous target genes that
mediate its diverse antitumor effects, and several target genes of
miR-34a have been reported, including Notch1 (14,15),
cyclin-dependent kinase 6 (CDK6) (11), E2F transcription factor 3 (E2F3)
(12), cyclin E2 (CCNE2) (16) and silent information regulator 1
(SIRT1) (17).
The Notch signaling pathway is highly conserved and
regulates the biological processes involved in cell fate
specification, differentiation, proliferation, apoptosis, adhesion,
migration and angiogenesis (18).
Abnormal Notch signaling has been found in the tumorigenesis of
cancers (19–21) and ectopic expression of Notch
signaling is also involved in the carcinogenesis of RCC (22,23).
Previous studies have shown that miR-34a regulates the expression
of Notch1 in a number of human cancers (14,15).
Thus, this study hypothesized that there is a significant
correlation between antineoplastic miR-34a and oncogenic Notch1 in
the development of RCC. This study aimed to investigate the
biological function and molecular mechanisms of miR-34a in human
renal cell carcinoma cells.
Materials and methods
Tissue samples
Patient-matched RCC (10 pairs) and normal renal
tissues were obtained from patients who underwent radical
nephrectomy at the Department of Urology, Shanghai First People’s
Hospital, School of Medicine, Shanghai Jiaotong University
(Shanghai, China) between 2006 and 2010. The normal renal tissues
were obtained from a distance of ≥5 cm from the tumor tissues.
Paired tissue specimens (n=10) were identified as clear cell RCCs
with four, five and one specimen classified as stages I, II and
III, respectively, according to clinical tumor-node-metastasis
staging (American Joint Committee on Cancer); whereas one, seven
and two RCCs were classified as grades I, II and III, respectively,
according to their differentiation status. The clinical and
pathological characteristics of the renal samples are presented in
Table I. All the specimens were
snap-frozen in liquid nitrogen immediately and stored at −80°C
following surgery, until RNA extraction. The histological diagnosis
was confirmed by examining hematoxylin and eosin-stained original
sections simultaneously by two pathologists. The The Research and
Ethics Committees of Shanghai First People’s Hospital (Shanghai,
China) approved the study, and patient consent was obtained prior
to tissue collection.
 | Table IClinicopathological characteristics
and miRNA-34a expression in renal cell carcinoma tissues. |
Table I
Clinicopathological characteristics
and miRNA-34a expression in renal cell carcinoma tissues.
| No. | Age (years) | Gender | Clinical stage | Pathological
grade | Relative expression
of miR-34a |
|---|
| 1 | 59 | M | II | G2 | Downregulated |
| 2 | 56 | M | I | G2 | Downregulated |
| 3 | 71 | F | II | G2 | Upregulated |
| 4 | 66 | M | I | G2 | Downregulated |
| 5 | 61 | F | I | G1 | Downregulated |
| 6 | 64 | M | III | G3 | Upregulated |
| 7 | 62 | F | II | G2 | Downregulated |
| 8 | 57 | M | II | G2 | Downregulated |
| 9 | 57 | M | I | G3 | Downregulated |
| 10 | 74 | M | II | G2 | Downregulated |
Cell culture
The 786-O and Caki-1 human RCC cell lines were
obtained from the Cell Resource Center of Shanghai Institutes for
Biological Sciences, Chinese Academy of Sciences (Shanghai, China).
The 786-O cells were cultured in RPMI-1640 medium (Gibco-BRL,
Rockville, MD, USA) supplemented with 10% fetal bovine serum (FBS;
Gibco-BRL) and 100 U/ml penicillin/streptomycin (Invitrogen Life
Technologies, Carlsbad, CA, USA). The Caki-1 cells were cultured in
McCoy’s 5A medium (Gibco-BRL) supplemented with 10% FBS and 100
U/ml penicillin/streptomycin. The cells were cultured in a
humidified incubator in an atmosphere of 5% CO2 at
37°C.
Cell transfection
The 786-O and Caki-1 cells were seeded at
1×105 cells per well in 6-well plates and transfected
with 50 nM of miR-34a mimics or the negative control (GenePharma,
Co., Ltd., Shanghai, China) using Lipofectamine® RNAiMAX
(Invitrogen Life Technologies) according to the manufacturer’s
instructions. The cells were then harvested 48 h after
transfection.
RNA extraction
Total RNA from cultured cells and tissues was
extracted using the TRIzol reagent (Invitrogen Life Technologies)
according to the manufacturer’s instructions. RNA quantity and
quality were determined by spectrophotometry at 260 nm (SmartSpec
Plus, Bio-Rad, Hercules, CA, USA) and agarose gel electrophoresis
(SantaiBio, Shanghai, China).
Quantitative polymerase chain reaction
(qPCR)
Reverse transcription for miRNAs was performed using
the TaqMan MicroRNA Reverse Transcription kit (Applied Biosystems,
Foster City, CA, USA). TaqMan MicroRNA expression assays (Applied
Biosystems) were used to provide specific primers for the reverse
transcription and quantitation of mature miR-34a and RNU6B. Thermal
cycling conditions for qPCR were as follows: 95°C for 10 min,
followed by 40 cycles of 95°C for 15 sec and 60°C for 60 sec. RNU6B
expression was used as an internal control for miR-34a expression.
In order to detect mRNA, 500 ng of RNA was reverse-transcribed
using a PrimeScript Reverse Transcriptase reagent kit (Takara,
Dalian, China) according to the manufacturer’s instructions, and
was amplified by qPCR using specific primers (Table II).
Glyceraldehyde-3-phosphate-dehydrogenase (GAPDH) was used as an
internal control to normalize for differences in the input RNA. The
PCR conditions were as follows: One cycle at 95°C for 1 min and 34
cycles at 94°C for 10 sec, 60°C for 30 sec and 72°C for 15 sec. All
the measurements were performed in triplicate. Amplification was
analyzed using the ΔΔCt method (24).
| Table IIPrimer sequences of target genes of
microRNA-34a. |
Table II
Primer sequences of target genes of
microRNA-34a.
| Target genes | Forward primer | Reverse primer |
|---|
| SIRT1 |
5′-TCCTGGACAATTCCAGCCATCTCT-3′ |
5′-TCCAGCGTGTCTATGTTCTGGGTA-3′ |
| Notch1 |
5′-TGGAGAAGGGAAGTTGAACGAGCA-3′ |
5′-CAAATTAATCCGCGTGCGGAAGGT-3′ |
| CDK6 |
5′-ATTCACTGCCTGGGACACAGTCTT-3′ |
5′-ACAGGCCACTGTGGTAACTCTCAA-3′ |
| E2F3 |
5′-TGCAGTGTTGTCCCTTCCTACCTT-3′ |
5′-GCCTGCAACTGTGCGTTTAGACAA-3′ |
| CCNE2 |
5′-ATGACACCACCGAAGAGCACTGAA-3′ |
5′-TGGCTAGGGCAATCAATCACAGCA-3′ |
| GAPDH |
5′-TCGACAGTCAGCCGCATCTTCTTT-3′ |
5′-ACCAAATCCGTTGACTCCGACCTT-3′ |
Cell proliferation analysis by Cell
Counting Kit-8 (CCK-8)
The cells were seeded into 96-well plates at 1,000
cells per well 24 h after transfection. The effects of miR-34a on
cell proliferation were detected 0, 24 and 48 h after seeding using
CCK-8 (Dojindo Molecular Technologies, Inc., Kumamoto, Japan)
according to the manufacturer’s instructions. Each assay was
performed in five replicates on three independent experiments.
Cell cycle analysis by flow
cytometry
After transfection (48 h) and prior to cell
counting, the cells were collected, washed with cold
phosphate-buffered saline (PBS), fixed in cold 70% ethanol in PBS
for at least 24 h and labeled with propidium iodide. After
staining, cells were counted on FACSCalibur using CellQuest Pro
software (BD Biosciences, Franklin Lakes, NJ, USA). The cell cycle
fractions were analyzed using ModFit software, version 3.0 (BD
Biosciences).
Western blot analysis
Total protein was extracted from the cells using
whole cell lysates (Beyotime, Jiangsu, China). The protein
concentrations of individual samples were assessed using a standard
bicinchoninic acid assay (Beyotime). For each sample, 60 μg of
protein was separated on 10% SDS-PAGE gel (Bio-Rad), transferred
onto a polyvinylidene difluoride membrane (catalogue number:
3010040001; Roche Applied Science, Mannheim, Germany)and blocked
with 5% skimmed milk and 0.1% Tris-buffered saline-Tween 20 (TBST)
at room temperature for 1.5 h. The membranes were washed in TBST
three times and incubated overnight at 4°C with monoclonal rabbit
anti-Notch1 antibody (1/1,000 dilution) and monoclonal rabbit
anti-GAPDH antibody (1/8,000 dilution). The primary antibodies
recognize human protein and were purchased from Cell Signaling
Technology, Inc. (Beverly, MA, USA). The membranes were then washed
with 1X TBST (Jiuzhou Tech, Beijing, China) and incubated with
anti-rabbit IgG horseradish peroxidase-conjugated secondary
antibody (Cell Signaling Technology, Inc.). The protein expression
was evaluated using chemiluminescence and exposure to Kodak
film.
Statistical analysis
Data are presented as the means ± standard deviation
from at least three separate experiments and significance was
analyzed using the Student’s t-test. All the analyses were
performed using SPSS software, version 17.0 (SPSS Inc., Chicago,
IL, USA). P<0.05 is considered to indicate a statistically
significant difference.
Results
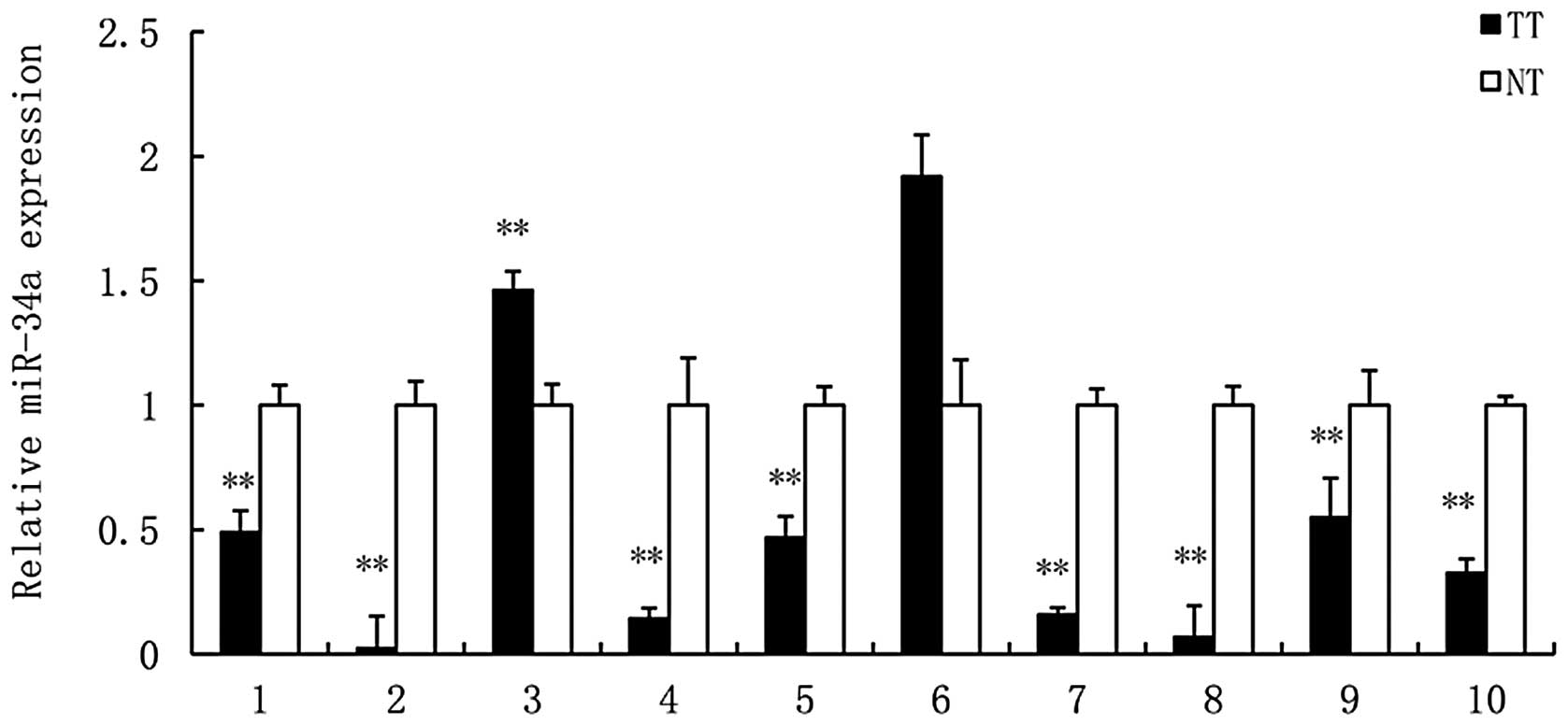
miR-34a expression is downregulated in
renal tumors
To investigate the underlying mechanisms of miR-34a
in the development of RCC, the expression levels of miR-34a in
human RCC tissues and adjacent normal tissues were examined by
qPCR. miR-34a was significantly downregulated in eight of the 10
(80%) human RCC tissues compared with the adjacent normal tissues
(Fig. 1). These findings suggest
that the downregulation of miR-34a may be involved in human RCC
carcinogenesis.
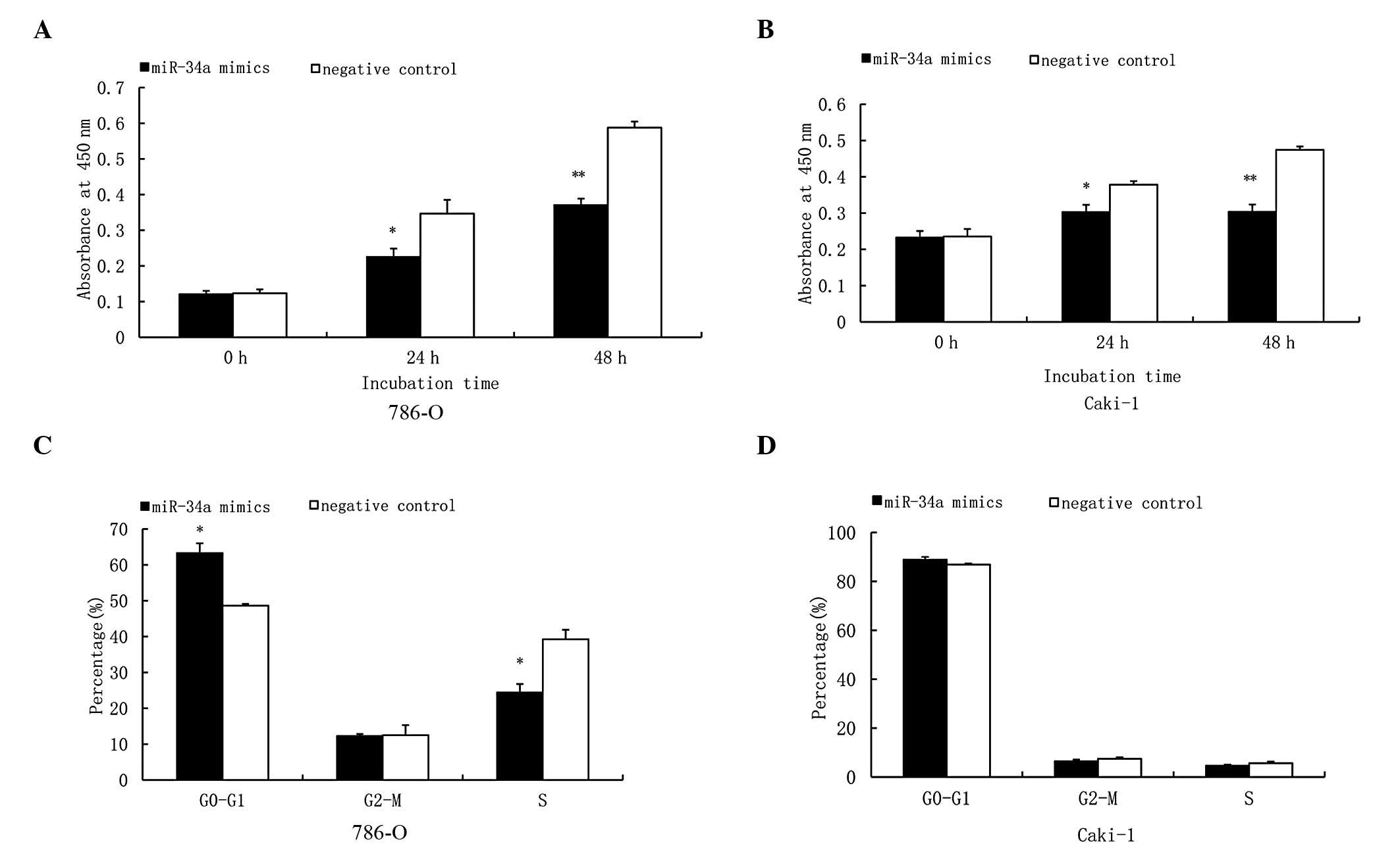
Functional effects of miR-34a on
proliferation and cell cycle progression in RCC cell lines
The effects of miR-34a on cell proliferation in two
RCC cell lines was investigated. To validate the antiproliferative
function of miR-34a in RCCs, the RCC cell lines were transfected
with miR-34a mimics and the negative control. The proliferation
rate of each RCC cell line 24 h after transfection and over the
following three days was examined using CCK-8. Our findings
suggested that the overexpression of miR-34a significantly
inhibited the cell proliferation rate compared with the negative
control in the two RCC cell lines (Fig.
2A and B). Flow cytometry examined the functional effects of
miR-34a on cell cycle progression. Different alterations in the
cell cycle were observed in the two RCC cell lines (Fig. 2C and D). A significant decrease in
the percentage of cells in S phase and a significant arrest in the
G0–G1 phase was observed in the 786-O cells (Fig. 2C); whereas no significant decrease
in the percentage of cells in S phase was observed in Caki-1 cells,
but a significant arrest in the G0–G1 phase was identified
(Fig. 2D).
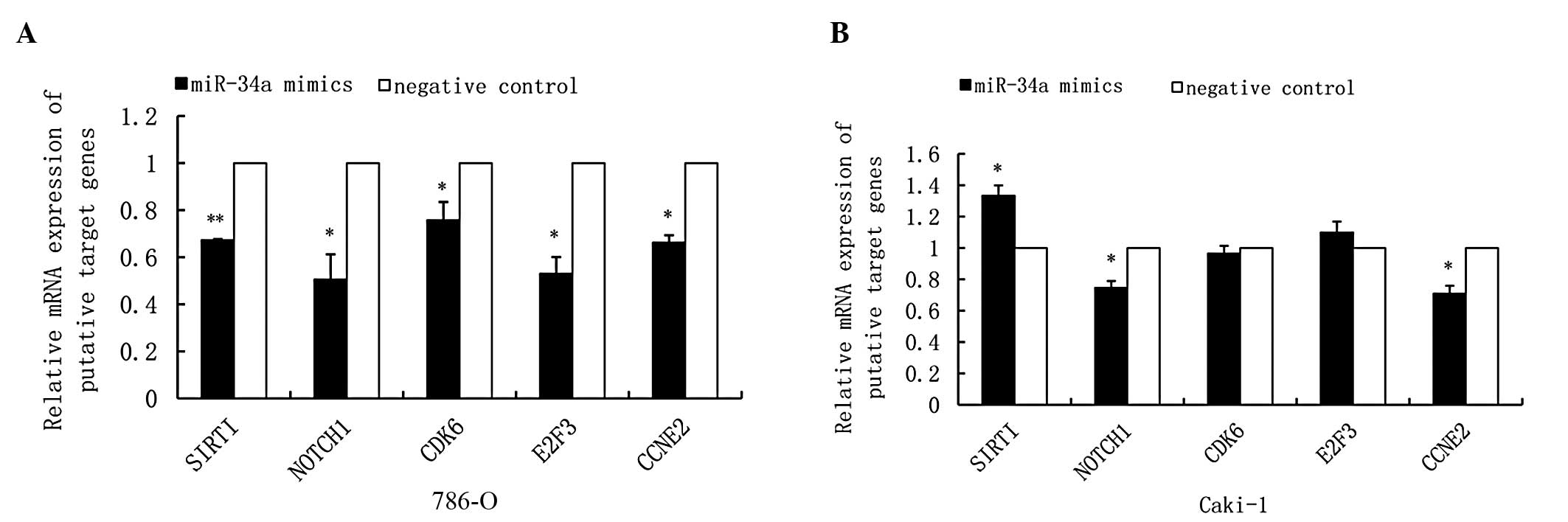
Relative mRNA expression levels of
putative target genes of miR-34a in RCC cell lines
The molecular mechanisms of miR-34a in the
development of RCC were further investigated. miR-34a exerts its
diverse inhibitory effects by regulating a number of target genes.
According to the bioinformatic analysis on TargetScan (http://www.targetscan.org/), SIRT1, Notch1, CDK6, E2F3
and CCNE2 are putative target genes of miR-34a and have been found
in different tumors. In the present study, the mRNA expression
levels of these genes following transfection were examined by qPCR.
Our data showed that the mRNA expression levels of SIRT1, Notch1,
CDK6, E2F3 and CCNE2 in the 786-O cell line were significantly
reduced in the miR-34a mimics group compared with the negative
control group (Fig. 3A). In the
Caki-1 cell line, Notch1 and CCNE2 were downregulated and miR-34a
exerted almost no significant effects on the mRNA levels of SIRT1,
CDK6 and E2F3 (Fig. 3B).
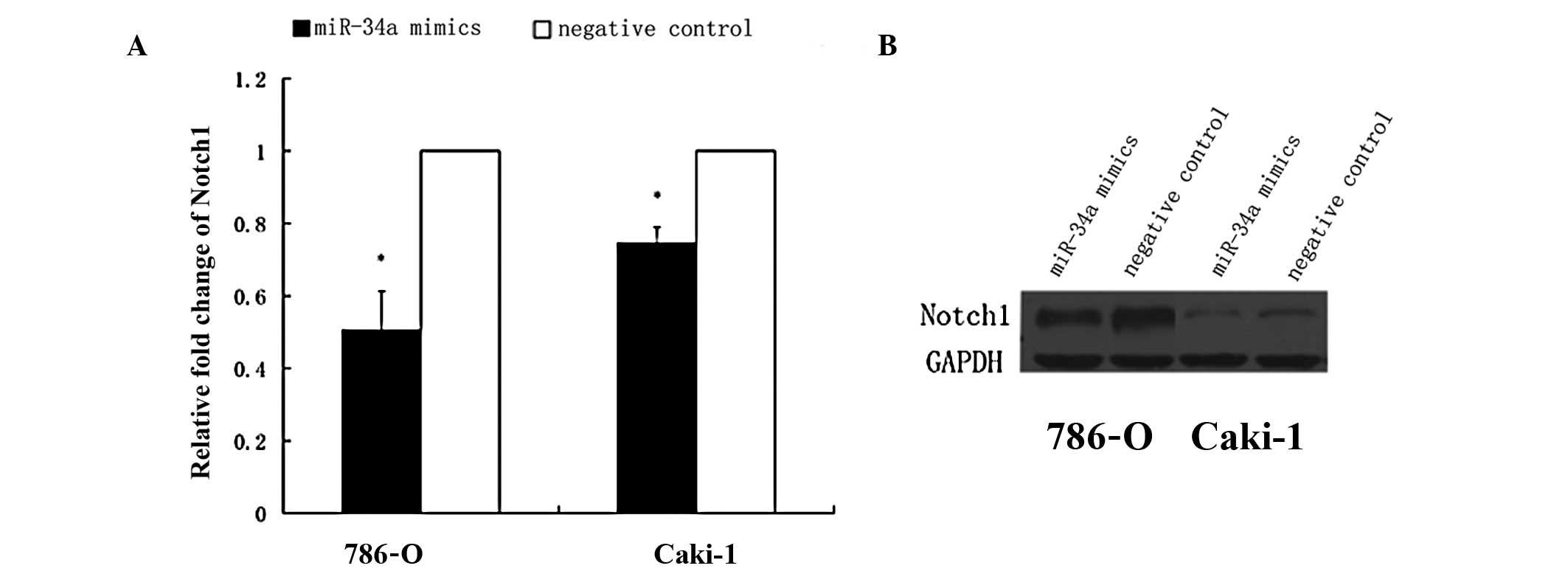
Notch1 is downregulated by miR-34a in the
RCC cell lines
Among all the putative and reported genes in the
current study, we found that miR-34a significantly reduced the mRNA
expression of Notch1 in the 786-O and Caki-1 RCC cell lines. Thus,
our study focused on Notch1 and investigated whether the forced
expression of miR-34a could downregulate the expression of Notch1
in RCC. The RCC cell lines were transfected with miR-34a mimics and
the negative control; after 48 h, the cells transfected with
miR-34a mimics displayed decreased Notch1 protein and mRNA levels
in comparison to those transfected with the negative control
(Fig. 4A and B).
Discussion
A number of studies have reported a relatively low
expression level of miR-34a in human tumors and cancer cell lines
(12,18). In accordance with these
observations, qPCR examination revealed that the expression of
miR-34a was reduced in eight of the 10 (80%) human RCC tissues
compared with the adjacent normal tissues (Fig. 1), which suggested that dysregulation
of miR-34a may be involved in the development of human RCC.
However, a limitation of the present study was the small number of
tissue samples. Our study measured the expression of miR-34a in RCC
tissue to investigate whether miR-34a is involved in the
development of RCC; although, the main purpose of the study was to
explore the biological function and molecular mechanism of miR-34a
in RCC cell lines. The decreased expression of miR-34a in tumors
may be correlated with the loss of chromosome 1p36, p53 mutations
and CpG methylation of the miR-34a promoters in cancer. The 1p36
genomic region is frequently deleted in a number of tumors
(25) and miR-34a was reported to
be located within chromosome 1p36 (12,26).
Furthermore, previous studies have reported that miR-34a is the
target of p53 tumor suppressor gene, underlying the downregulation
of miR-34a in tumors (6,9), and p53 mutation is involved in the
development of RCC (27). In
addition, miR-34a was inactivated by aberrant CpG methylation in
various types of cancer, including RCC (28). Thus, the downregulation of miR-34a
may be a common event involved in the tumorigenesis of RCC.
Our study focused on the antiproliferative effects
of miR-34a in RCC. Forced expression of miR-34a was observed to
reduce the proliferation rate of the RCC cell lines (Fig. 2A and B) compared with the negative
control. In addition, Notch signaling plays important roles in the
carcinogenesis of RCC. Xu et al (29) showed that Notch1 functions as an
oncogene, promoting the growth of RCC. It has also been reported
that Notch1 is associated with the metastasis of RCC (22,23).
Previous studies have shown that Notch1 is the target of miR-34a in
a number of tumors (14,15,30).
However, to the best of our knowledge, it had not previously been
demonstrated that miR-34a downregulates the expression of Notch1 in
RCC. Our findings suggested that miR-34a repressed the expression
of Notch1 at the protein and mRNA level (Fig. 4). Thus, the suppressive function of
miR-34a on the proliferation of RCC cells may be partly due to the
regulation of Notch1 and miR-34a may affect the Notch signaling
pathway.
Previous studies have reported that miR-34a induces
cell cycle arrest in tumors (11,14,15);
however, in our study, a significant decrease in the percentage of
cells in S phase and a significant G0–G1 arrest was observed in
786-O cells, while no significant decrease in the percentage of
cells in S phase, but a significant G0–G1 arrest was identified in
Caki-1 cells (Fig. 2C and D).
Therefore, there may be alternative mechanisms involved in cell
cycle distribution in the Caki-1 RCC cell line, which require
further investigation. The control of cell proliferation by miRNAs
may be via the regulation of critical cell cycle regulators
(31). Our data suggested that
miR-34a may, in part, act through additive or synergistic effects
of multiple cell-cycle-related proteins, such as CDK6, E2F3 and
CCNE2, mediating the dysregulation of cell growth and proliferation
in RCCs (Fig. 3).
In conclusion, the expression of miR-34a was lower
in RCC tumors compared with the adjacent normal tissues. Several
putative target genes of miR-34a were also dysregulated at the mRNA
level when the expression of miR-34a was elevated in the RCC cell
lines. Our findings suggest that miR-34a could suppress
carcinogenesis by modulating the expression of Notch1 at the mRNA
and protein level, thus inhibiting cell proliferation and inducing
cell cycle arrest at G0 phase in the RCC cell lines. In conclusion,
miR-34a expression is associated with the malignant behavior of RCC
cells and, therefore, may be a candidate molecular therapeutic
target for RCC in the future (32).
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (no. 81372753) and the Program
of Science and Technology Commission of Shanghai Municipality (no.
124119a2200). The authors would like to thank Youyi Lu for
assistance with the study.
Abbreviations:
|
RCC
|
renal cell carcinoma
|
|
miR-34a
|
microRNA-34a
|
|
CCK-8
|
Cell Counting Kit-8
|
|
qPCR
|
quantitative polymerase chain
reaction
|
|
GAPDH
|
glyceraldehyde-3-phosphate-dehydrogenase
|
|
SIRT1
|
silent information regulator 1
|
|
CDK6
|
cyclin-dependent kinase 6
|
|
CCNE2
|
cyclin E2
|
References
|
1
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kloosterman WP and Plasterk RH: The
diverse functions of microRNAs in animal development and disease.
Dev Cell. 11:441–450. 2006. View Article : Google Scholar
|
|
3
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar
|
|
4
|
Zhou L, Yin B, Liu Y, Hong Y, Zhang C and
Fan J: Mechanism and function of decreased FOXO1 in renal cell
carcinoma. J Surg Oncol. 105:841–847. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu Y, Yin B, Zhang C, Zhou L and Fan J:
Hsa-let-7a functions as a tumor suppressor in renal cell carcinoma
cell lines by targeting c-myc. Biochem Biophys Res Commun.
417:371–375. 2012. View Article : Google Scholar
|
|
6
|
Chang TC, Wentzel EA, Kent OA, et al:
Transactivation of miR-34a by p53 broadly influences gene
expression and promotes apoptosis. Mol Cell. 26:745–752. 2007.
View Article : Google Scholar
|
|
7
|
Yamamura S, Saini S, Majid S, et al:
MicroRNA-34a modulates c-Myc transcriptional complexes to suppress
malignancy in human prostate cancer cells. PLoS One. 7:e297222012.
View Article : Google Scholar
|
|
8
|
Gallardo E, Navarro A, Viñolas N, et al:
miR-34a as a prognostic marker of relapse in surgically resected
non-small-cell lung cancer. Carcinogenesis. 30:1903–1909. 2009.
View Article : Google Scholar
|
|
9
|
He L, He X, Lim LP, et al: A microRNA
component of the p53 tumour suppressor network. Nature.
447:1130–1134. 2007. View Article : Google Scholar
|
|
10
|
Hermeking H: p53 enters the microRNA
world. Cancer Cell. 12:414–418. 2007. View Article : Google Scholar
|
|
11
|
Sun F, Fu H, Liu Q, et al: Downregulation
of CCND1 and CDK6 by miR-34a induces cell cycle arrest. FEBS Lett.
582:1564–1568. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Welch C, Chen Y and Stallings RL:
MicroRNA-34a functions as a potential tumor suppressor by inducing
apoptosis in neuroblastoma cells. Oncogene. 26:5017–5022. 2007.
View Article : Google Scholar
|
|
13
|
Tazawa H, Tsuchiya N, Izumiya M and
Nakagama H: Tumor-suppressive miR-34a induces senescence-like
growth arrest through modulation of the E2F pathway in human colon
cancer cells. Proc Natl Acad Sci USA. 104:15472–15477. 2007.
View Article : Google Scholar
|
|
14
|
Pang RT, Leung CO, Ye TM, et al:
MicroRNA-34a suppresses invasion through downregulation of Notch1
and Jagged1 in cervical carcinoma and choriocarcinoma cells.
Carcinogenesis. 31:1037–1044. 2010. View Article : Google Scholar
|
|
15
|
Li WB, Ma MW, Dong LJ, Wang F, Chen LX and
Li XR: MicroRNA-34a targets notch1 and inhibits cell proliferation
in glioblastoma multiforme. Cancer Biol Ther. 12:477–483. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar
|
|
17
|
Yamakuchi M, Ferlito M and Lowenstein CJ:
miR-34a repression of SIRT1 regulates apoptosis. Proc Natl Acad Sci
USA. 105:13421–13426. 2008. View Article : Google Scholar
|
|
18
|
Bolós V, Grego-Bessa J and de la Pompa JL:
Notch signaling in development and cancer. Endocr Rev. 28:339–363.
2007.
|
|
19
|
Tohda S and Nara N: Expression of Notch1
and Jagged1 proteins in acute myeloid leukemia cells. Leuk
Lymphoma. 42:467–472. 2001. View Article : Google Scholar
|
|
20
|
Reedijk M, Odorcic S, Chang L, et al:
High-level coexpression of JAG1 and NOTCH1 is observed in human
breast cancer and is associated with poor overall survival. Cancer
Res. 65:8530–8537. 2005. View Article : Google Scholar
|
|
21
|
Wang Z, Zhang Y, Li Y, Banerjee S, Liao J
and Sarkar FH: Down-regulation of Notch-1 contributes to cell
growth inhibition and apoptosis in pancreatic cancer cells. Mol
Cancer Ther. 5:483–493. 2006. View Article : Google Scholar
|
|
22
|
Ai Q, Ma X, Huang Q, et al: High-level
expression of Notch1 increased the risk of metastasis in T1 stage
clear cell renal cell carcinoma. PLoS One. 7:e350222012. View Article : Google Scholar
|
|
23
|
Sjölund J, Boström AK, Lindgren D, et al:
The notch and TGF-β signaling pathways contribute to the
aggressiveness of clear cell renal cell carcinoma. PLoS One.
6:e230572011.
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
|
|
25
|
Kaghad M, Bonnet H, Yang A, et al:
Monoallelically expressed gene related to p53 at 1p36, a region
frequently deleted in neuroblastoma and other human cancers. Cell.
90:809–819. 1997. View Article : Google Scholar
|
|
26
|
Wei JS, Song YK, Durinck S, et al: The
MYCN oncogene is a direct target of miR-34a. Oncogene.
27:5204–5213. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Reiter RE, Anglard P, Liu S, Gnarra JR and
Linehan WM: Chromosome 17p deletions and p53 mutations in renal
cell carcinoma. Cancer Res. 53:3092–3097. 1993.
|
|
28
|
Lodygin D, Tarasov V, Epanchintsev A, et
al: Inactivation of miR-34a by aberrant CpG methylation in multiple
types of cancer. Cell Cycle. 7:2591–2600. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu L, Zhu Y, Xu J, et al: Notch1
activation promotes renal cell carcinoma growth via PI3K/Akt
signaling. Cancer Sci. 103:1253–1258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Du R, Sun W, Xia L, et al: Hypoxia-induced
down-regulation of microRNA-34a promotes EMT by targeting the Notch
signaling pathway in tubular epithelial cells. PLoS One.
7:e307712012. View Article : Google Scholar
|
|
31
|
Bueno MJ and Malumbres M: MicroRNAs and
the cell cycle. Biochim Biophys Acta. 1812:592–601. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nana-Sinkam SP and Croce CM: MicroRNAs as
therapeutic targets in cancer. Transl Res. 157:216–225. 2011.
View Article : Google Scholar : PubMed/NCBI
|