Introduction
Osteosarcoma (OS), the most common malignant bone
tumor, occurs mainly in adolescents and young adults, with a
morbidity of ∼5 cases per million (1). OS develops from a common mesenchymal
stem cell progenitor exhibiting a disruption of normal osteoblast
differentiation (2). Although the
5-year survival rate has increased to 60–70%, a significant
proportion of patients respond poorly to chemotherapy and are at
high risk of relapse or metastasis, even following curative
resection (3). The majority of OSs
exhibit numerous and highly variable genomic aberrations. DNA
methylation is an important mechanism for the downregulation of
gene expression, more specifically at CpG islands in the promoter
region of genes. It was previously reported that epigenetic
modifications may be more prevalent than mutations in certain types
of childhood cancer, such as retinoblastoma (4). A number of study groups have reported
comparisons of alterations in DNA methylation/deacetylation and
microRNA (miRNA) expression for other types of cancer (5), although only a limited number of studies
have investigated the association of these types of mechanisms with
OS. The identification of molecular markers and pathways
contributing to OS development and progression may facilitate
earlier diagnosis and the development of novel treatment
strategies.
miRNAs are short endogenous non-coding RNAs that
post-transcriptionally regulate the expression of target genes
(oncogenes or tumor suppressor genes) involved in several types of
cancer, including OS (6). A single
miRNA may silence a large number of genes, allowing these molecules
extensive control over numerous cellular functions (7). Evidence of individual miRNAs affecting
developmental biology, cellular differentiation and oncogenesis is
continuously emerging (8). Of note,
it was recently reported that miRNAs may be able to cause heritable
changes in gene expression by epigenetic mechanisms, such as
altering DNA methyltransferase levels, whereas miRNAs themselves
may be regulated by epigenetic mechanisms (9). Certain alterations in miRNAs and the
possible mechanisms through which miRNAs affect cell proliferation
and migration in OS were recently found to be correlated with
methylation epigenetic mechanisms (10,11). The
majority of the downregulated miRNAs were found to be silenced
through epigenetic mechanisms in solid malignancies, including
breast cancer (12) and neuroblastoma
(13). In OS, the expression levels
of tumor suppressor miRNAs, including miRNA-34 (miR-34) and
miR-143, were recently found to be downregulated. The miR-34 gene
was hypermethylated in OS, which was found to be associated with
cancer cell metastasis (14).
However, there is currently no sufficient evidence supporting the
correlation of miRNA overexpression in OS with epigenetic events.
It was suggested that miR-142 may play an important role in
maintaining the self-renewal capacity of bronchioalveolar stem
cells (15). It was also reported
that patients with esophageal squamous cell carcinoma (ESCC)
displaying a high expression of miR-142 had poorer survival rates
compared with those with low expression of miR-142, suggesting that
miR-142 may act as a tumor suppressor (16).
Although miR-142 has been found to be downregulated
in OS cell lines (17), the role and
epigenetic mechanisms of miR-142 in OS remain unknown. The aim of
the present study was to compare the expression of miR-142 between
OS and pericancerous tissues and investigate the role of epigenetic
regulation in the decreased expression of miR-142 in OS.
Materials and methods
Tissue samples
All the patients signed an informed consent,
approved by the Independent Ethics Committee of Central South
University (Changsha, China). The OS and matched adjacent normal
tissues used in this study were collected from 6 patients who were
treated between 2011 and 2014 in Xiangya Hospital (Changsha,
China). All the samples were stored at −80°C until tissue
analysis.
Cell culture
hFOB1.19, Saos-2, MG63, U2OS and HOS cells were
obtained from American Type Culture Collection (Manassas, VA, USA).
All the cells were cultured in RPMI-1640 basic medium (Gibco Life
Technologies, Carlsbad, CA, USA) supplemented with 10% fetal bovine
serum (FBS; Invitrogen Life Technologies, Carlsbad, CA, USA). All
the cells were cultured under conditions of 95% air and 5%
CO2 at 37°C.
Epigenetic drug treatment of
cells
The Saos-2 and MG63 cells were divided in four
groups as follows: i) Control cells; ii) cells treated with 15.55
nM of the demethylation drug 5-Aza-2′-deoxycytidine (Aza;
Sigma-Aldrich, St. Louis, MO, USA); iii) cells treated with 1.5 nM
of the histone deacetylase inhibitor 4-phenylbutyric acid (PBA;
Sigma-Aldrich); and iv) cells co-treated with 15.55 nM Aza and 1.5
nM PBA (Aza + PBA).
Quantitative polymerase chain reaction
(qPCR) analysis
Total RNA was extracted from the indicated cells
using TRIzol® reagent (Invitrogen Life Technologies) following the
manufacturer's instructions. The specific primers for miRNA-142 and
U6 were purchased from GeneCopoeia (Rockville, MD, USA). The
relative expression of miR-142 was measured using the miScript
SYBR® Green PCR Kit (Qiagen, Venlo, The Netherlands). The
expression of U6 was used as an endogenous control. Data were
processed using the 2−ΔΔCt method.
Cell counting kit-8 (CCK-8) cell
proliferation assay
CCK-8 was used to evaluate cell proliferation. A
total of 5×103 cells were seeded in each 96-well plate
for 24 h, treated with the indicated drugs and further incubated
for 0, 24, 48 and 72 h. One hour prior to the completion of the
incubation, 10 µl CCK-8 reagent was added to each well. The optical
density (OD) 450 nm value in each well was determined by an enzyme
immunoassay analyzer (Thermo Fisher Scientific, Inc., Waltham, MA,
USA).
Transwell™ assay
Following treatment with the indicated drugs for 72
h, the cells were starved in serum-free medium (Invitrogen Life
Technologies) for 24 h and resuspended in serum-free medium. The
cells were added to the upper chamber, while the lower chamber was
filled with basic medium containing 10% FBS. Following incubation
for 24 h, the cells attached to the bottom were fixed and stained
with crystal violet (Amresco, LLC, Solon, OH, USA) for 20 min. The
redundant crystal violet was washed by 0.1 M phosphate-buffered
saline (PBS; Amresco, LLC), and dried in air. The OD of crystal
violet dissolved in 10% acetic acid at 570 nm was detected by an
enzyme immunoassay analyzer.
Flow cytometric analysis of the cell
cycle
The cells were treated with the indicated drugs for
72 h. Following trypsinization and washing with ice-cold PBS, the
cell suspensions were stained using BD Cycletest™ Plus (BD
Biosciences, Franklin Lakes, NJ, USA) according to the
manufacturer's instructions and the cell cycle was then analyzed by
flow cytometry (Beckman Coulter, Brea, CA, USA). The experiments
were performed in triplicate.
Measurement of miR-142 promoter CpG
island methylation status by bisulfite genomic sequencing PCR (BSP)
and methylation-specific PCR (MSP)
Genomic DNA was extracted using the Takara Genomic
DNA Extraction kit (Takara Co., Dalian, China). Genomic DNA (1 µg
per sample) was modified with bisulfite using the Epitect Bisulfite
kit (Qiagen) according to the manufacturer's instructions. The
modified DNA was amplified using the following primers: miR-142
forward, TTTAGGGTATGAGAGAGGGTTTTTAG and reverse,
AATTCCCTTCAAAAAAAACAAAACT. The PCR products were gel-extracted
(Qiagen) to confirm that a single band had been obtained and were
then sequenced by Invitrogen Life Technologies.
MSP was performed on bisulfate-treated DNA. The
primers used were as follows: Unmethylated miR-142 forward,
TTAAGGTTTGTGTGGGTTTAAATG and reverse, AAATAAACAAAAAACAACATCAAC;
methylated miR-142 forward, TTTTAAGGTTTGTGTGGGTTTAAAC and reverse,
AAAATAAACAAAAAACAACGTCGAC. The annealing temperature was 60°C for
methylated-PCR and 55°C for unmethylated-PCR, with 27 cycles used
for each.
Statistical analysis
Student's t-tests or one-way analysis of variance
were used to analyze data using GraphPad Prism 5 software (GraphPad
Software, Inc., La Jolla, CA, USA), depending on the experimental
conditions. All the data are presented as mean ± standard
deviation. Compared with the respective controls, P<0.05 was
considered to indicate a statistically significant difference.
Results
Methylation of CpG islands and
expression of miR-142 in OS tissues and cell lines
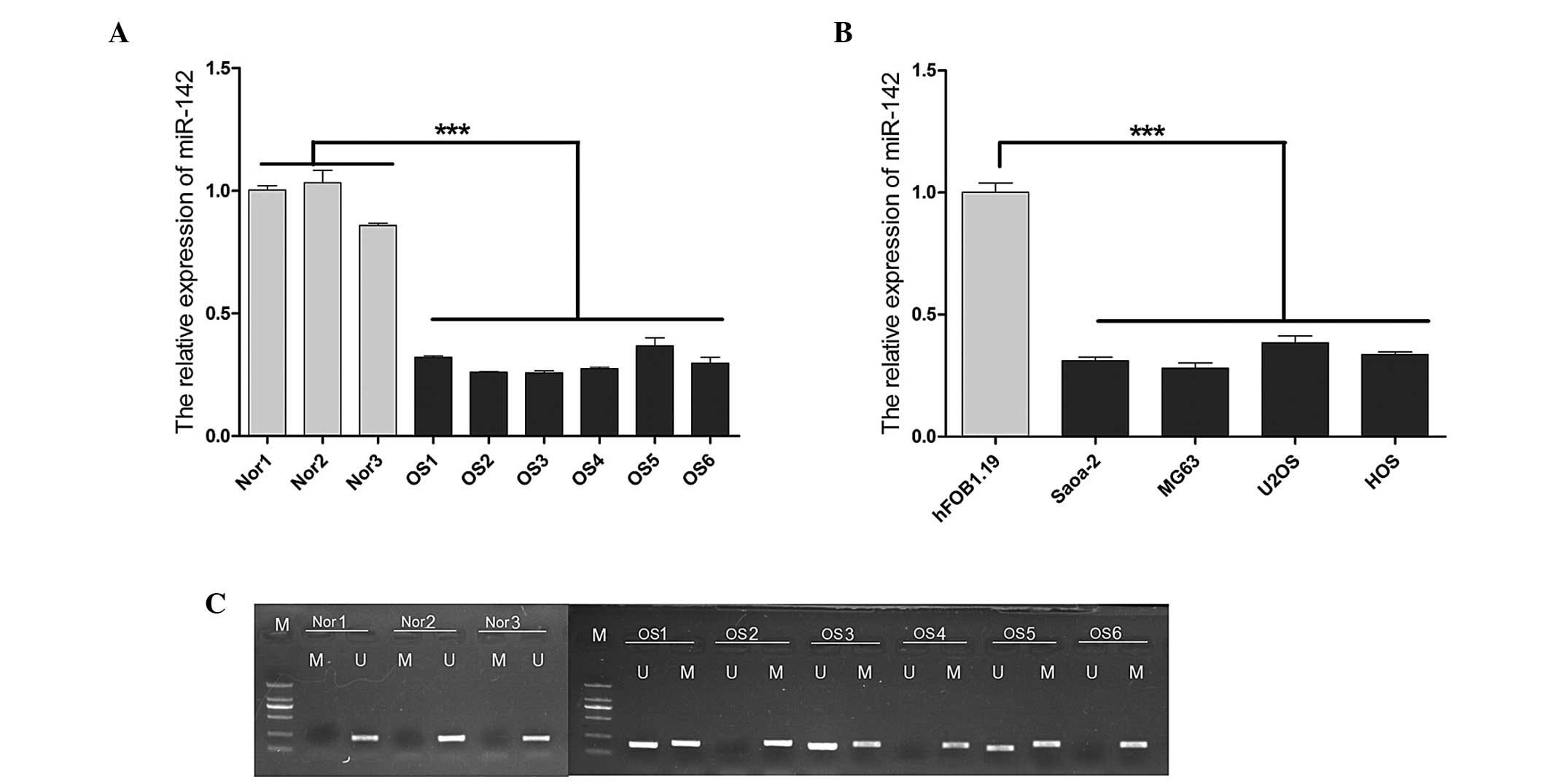
The average expression level of miR-142 was found to
be significantly lower (P<0.001) in the OS samples obtained from
the 6 OS patients compared with the normal controls, as indicated
by the reverse transcription-qPCR (Fig.
1A). Similar results were observed in the OS cell lines,
particularly in Saos-2 and MG63 cells (Fig. 1B). In light of these findings, the
role of epigenetic mechanisms that may be involved in the silencing
of miR-142 was investigated. MSP was used to detect miR-142
methylation in tissues obtained from the 6 patients with OS and the
normal tissue samples (Fig. 1C). A
significantly higher level of methylation was observed in OS
compared with that in normal tissues. There was no methylation
detected in any of the normal tissue samples.
Upregulated expression of miR-142 was
induced by treatment with demethylation agents
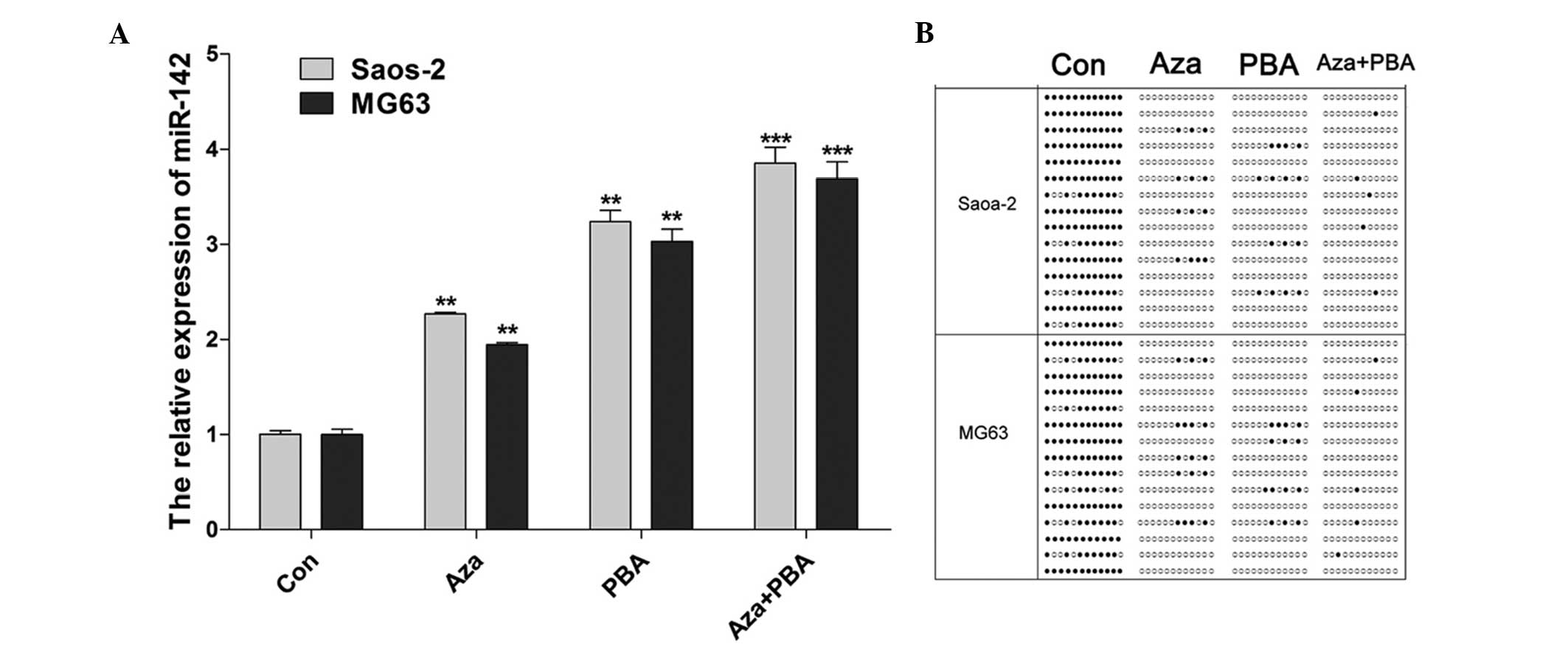
An increase in the expression of miR-142 was
observed in Saos-2 and MG63 cells treated with 15.55 nM Aza, 1.5 nM
PBA and 15.55 nM Aza + 1.5 nM PBA (Fig.
2A). The miR-142 CpG island methylation status was examined
using BSP following treatment with demethylation agents. The
results indicated that methylation was decreased by all the
treatments, particularly by the combined Aza + PBA treatment
(Fig. 2B).
Effects of demethylation agents on
cell proliferation and invasion
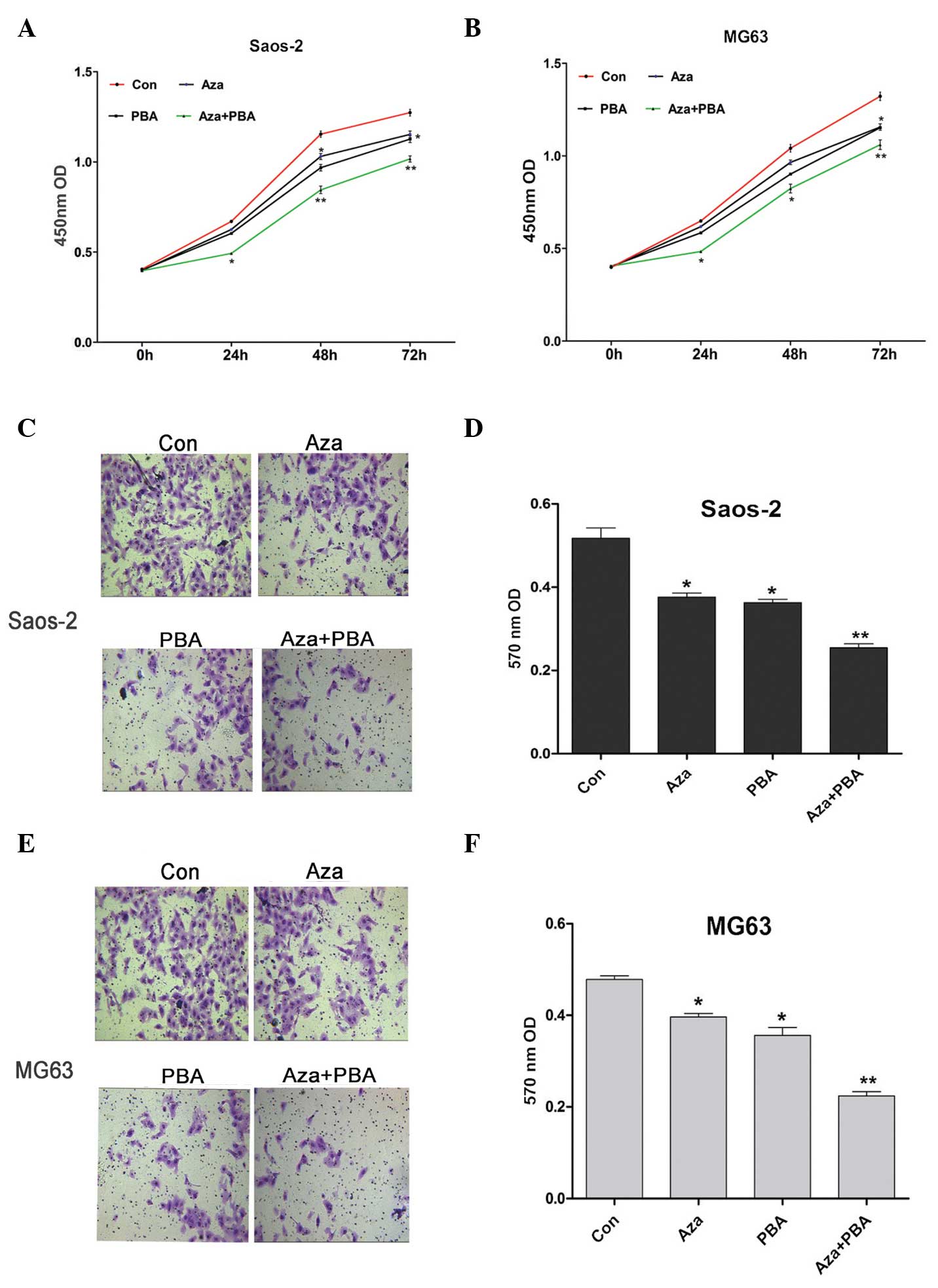
CCK-8 was used to assess the effects of
demethylation agents on Saos-2 and MG63 cell proliferation. It was
observed that inhibition of proliferation was induced by Aza or PBA
treatment alone, whereas a significantly higher level of inhibition
was observed in cells treated with Aza + PBA (Fig. 3A and B). The Transwell™ assay was used
to measure the invasive ability of Saos-2 and MG63 cells following
treatment with demethylation agents. The results indicated that
their invasive ability was significantly decreased by all the
treatments, particularly by the combined Aza + PBA treatment
(Fig. 3C-F).
Demethylation agents induce the
retardation of the S phase in Saos-2 and MG63 cells
Flow cytometric analysis was used to analyze cell
cycle alterations following treatment with demethylation agents. In
the cell cycle analysis, the ratio of cells in different phases of
the cell cycle was as shown in Fig.
4A and B. The results revealed that demethylation agents
induced cell cycle arrest, thus inducing an increase in
G2 phase cells, with a concomitant decrease in S phase
cells (Fig. 4C).
Discussion
The survival rates of patients with OS have improved
considerably following multiagent chemotherapy, with a 5-year
survival rate of ∼60% for patients without metastases (18). However, the survival rates have
reached a plateau and novel biology-based therapies are imperative
to achieve further improvement. At the molecular level, the
majority of OSs exhibit complex genomic aberrations and highly
variable patterns of gene expression. Emerging evidence has
demonstrated that several miRNAs were altered in OS, some of which
play a critical role in carcinogenesis (6,10,19,20).
Detailed understanding of miRNAs and their regulation mechanisms
may aid the development of novel strategies to improve OS
treatment.
The function of miR-142 has been primarily described
in the hematopoietic and immune systems. Increased expression of
miR-142 was observed in bronchioalveolar stem cells (BASCs),
suggesting that miR-142 may play an important role in maintaining
the self-renewal capacity of BASCs (15). In addition, ESCC patients with high
expression levels of miR-142 exhibited poorer survival rates
compared with those with low expression levels of miR-142 (16), whereas upregulated levels of miR-142
were involved in aggressive non-small-cell lung carcinoma (NSCLC)
(21,22). However, a recent study has suggested
that miR-142 may be associated with the suppression of NSCLC cells
(23). Wang et al (24) reported that miR-142 was upregulated by
the NGX6 metastasis suppressor gene in colon cancer cells.
Furthermore, their findings indicated that miR-142 plays a vital
role in suppressing the proliferation of colon cancer cells and
increasing the sensitivity of these cells to chemotherapeutic
agents by targeting CD133, ABCG2 and Lgr5. It was also found that
miR-142-3p acts as a tumor suppressor by targeting CD133, ABCG2 and
Lgr5 in colon cancer cells (25).
In the present study, we confirmed that miR-142 was
significantly decreased in OS tissues and cell lines. Although
previous data demonstrated that miR-142 was downregulated in OS
cell lines, as detected by global microarray analyses (17), the role of miR-142 in OS remains
unknown.
Mature miRNAs are generated from primary miRNA
transcripts (26), through an
intermediate precursor (27).
Increasing evidence indicates that, similar to protein-coding
genes, epigenetic mechanisms, such as DNA methylation and histone
acetylation, may be associated with the regulation of miRNA
expression (28). DNA methylation at
the dinucleotide CpG is one of the most common epigenetic
modifications in eukaryotic genomes and plays a significant role in
various biological processes (29).
The majority of human miRNA genes have been found to be associated
with CpG islands (30). However, the
epigenetic mechanisms underlying the tissue- and cell type-specific
regulation of miR-142 have not been clearly determined in the
majority of cases.
The present study revealed that a significantly
higher level of miR-142 methylation was observed in OS compared
with normal tissues. There was no methylation detected in any of
the normal tissue samples. The findings suggested that the
methylation mechanism of miR-142 may be involved in OS development.
To investigate the involvement of epigenetic silencing, Saos-2 and
MG63 cells were initially treated with the demethylating agent Aza
or/and PBA, which resulted in a concomitant increase of miR-142
transcripts, particularly in the group co-treated with Aza + PBA.
To provide further evidence, BSP was used to demonstrate that the
methylated CpG islands of miR-142 were significantly decreased by
Aza and PBA treatment in Saos-2 and MG63 cells. Furthermore, a
significant inhibition of proliferation in Saos-2 and MG63 cells by
expression of miR-142 was induced by demethylation agents. It was
also observed that increased miR-142 expression by Aza or PBA
repressed cell migration. The overexpression of miR-142 has been
shown to inhibit the proliferation and colony-forming ability of
primitive hematopoietic cells (31).
Some of these functions appear to depend on the cellular type and
context, since it was reported that miR-142 acts as an
anti-migratory factor in hepatocellular carcinoma cells (32) and has also been reported to be
deregulated in mesenchymal tumors (33). In the present study, demethylation
agents induced Saos-2 and MG63 cell cycle arrest, thus inducing an
increase in G2 phase cells with a concomitant decrease
in S phase cells. The exact mechanisms through which miR-142
inhibits cell proliferation and decreases S phase arrest remain
unknown. One possible explanation is that multiple genes associated
with the cell cycle are targeted by the miRNA signal. Therefore,
other genes regulated by miR-142 that affect cell growth, the cell
cycle and invasion should be further investigated.
In conclusion, the present study revealed that the
expression level of miR-142 was significantly lower in OS tissues
and cells due to hypermethylation. These findings suggested that
miR-142 played an important role in the inhibition of cell
proliferation and invasiveness of OS cell lines induced by
demethylation agents. Of note, this study investigated the role of
miR-142 only in vitro and the tumor-suppressive role of
miR-142 in OS requires further confirmation by studies performed
in vivo.
References
|
1
|
Kobayashi E, Hornicek FJ and Duan Z:
MicroRNA involvement in osteosarcoma. Sarcoma.
2012.3597392012.PubMed/NCBI
|
|
2
|
Cao ZQ, Shen Z and Huang WY: MicroRNA-802
promotes osteosarcoma cell proliferation by targeting p27. Asian
Pac J Cancer Prev. 14:7081–7084. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Diao CY, Guo HB, Ouyang YR, et al:
Screening for metastatic osteosarcoma biomarkers with a DNA
microarray. Asian Pac J Cancer Prev. 15:1817–1822. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Benavente CA, McEvoy JD, Finkelstein D, et
al: Cross-speciesgenomic and epigenomic landscape of
retinoblastoma. Oncotarget. 4:844–859. 2013.PubMed/NCBI
|
|
5
|
Jin J, Cai L, Liu ZM and Zhou XS:
miRNA-218 inhibits osteosarcoma cellmigration and invasion by
down-regulating of TIAM1, MMP2 and MMP9. Asian Pac J Cancer Prev.
14:3681–3684. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maire G, Martin JW, Yoshimoto M,
Chilton-MacNeill S, Zielenska M and Squire JA: Analysis of
miRNA-gene expression-genomic profiles reveals complex mechanisms
of microRNA deregulation in osteosarcoma. Cancer Genet.
204:138–146. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lewis BP, Burge CB and Bartel DP:
Conserved seedpairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kelly TK, De Carvalho DD and Jones PA:
Epigenetic modifications as therapeutic targets. Nat Biotechnol.
28:1069–1078. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jones KB, Salah Z, Del Mare S, et al:
miRNA signatures associate with pathogenesis and progression of
osteosarcoma. Cancer Res. 72:1865–1877. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Poos K, Smida J, Nathrath M, Maugg D,
Baumhoer D and Korsching E: HowmicroRNA and transcription factor
co-regulatory networks affect osteosarcoma cell proliferation. PLoS
Comput Biol. 9:e10032102013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Png KJ, Yoshida M, Zhang XH, et al:
MicroRNA-335 inhibits tumorreinitiation and is silenced
throughgenetic and epigenetic mechanisms in human breast cancer.
Genes Dev. 25:226–231. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Das S, Bryan K, Buckley PG, et al:
Modulation of neuroblastoma disease pathogenesis by an extensive
network of epigenetically regulated microRNAs. Oncogene.
32:2927–2936. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
He C, Xiong J, Xu X, et al: Functional
elucidation of MiR-34 in osteosarcomacells and primary tumor
samples. Biochem Biophys Res Commun. 388:35–40. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Qian S, Ding JY, Xie R, et al: MicroRNA
expression profile of bronchioalveolar stem cells from mouse lung.
Biochem Biophys Res Commun. 377:668–673. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin RJ, Xiao DW, Liao LD, et al:
MiR-142-3p as a potential prognostic biomarker for esophageal
squamous cell carcinoma. J Surg Oncol. 105:175–182. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Namløs HM, Meza-Zepeda LA, Barøy T, et al:
Modulation of the osteosarcoma expression phenotype by microRNAs.
PLoS One. 7:e480862012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Posthuma De Boer J, Witlox MA, Kaspers GJ
and van Royen BJ: Molecular alterations as target for therapy in
metastatic osteosarcoma: a review of literature. Clin Exp
Metastasis. 28:493–503. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Baumhoer D, Zillmer S, Unger K, et al:
MicroRNA profiling with correlation to gene expression revealed the
oncogenic miR-17-92 cluster to be up-regulated in osteosarcoma.
Cancer Genet. 205:212–219. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lulla RR, Costa FF, Bischof JM, et al:
Identification of differentially expressed microRNAs in
osteosarcoma. Sarcoma. 2011.7326902011.PubMed/NCBI
|
|
21
|
Boeri M, Verri C, Conte D, et al: MicroRNA
signatures intissues and plasma predictdevelopment and prognosis of
computed tomography detected lung cancer. In: Proc Natl Acad Sci
USA. 108. pp. 3713–3718. 2011; View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kaduthanam S, Gade S, Meister M, et al:
Serum miR-142-3p is associated with early relapse in operable lung
adenocarcinoma patients. Lung Cancer. 80:223–227. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lei Z, Xu G, Wang L, et al: MiR-142-3p
represses TGF-β-induced growth inhibition through repression of
TGFβR1 in non-small cell lung cancer. FASEB J. 28:2696–2704. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang XY, Wu MH, Liu F, et al: Differential
miRNAexpression and their target genes between NGX6-positive and
negative colon cancer cells. Mol Cell Biochem. 345:283–290. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shen WW, Zeng Z, Zhu WX and Fu GH:
MiR-142-3p functions as a tumor suppressor by targeting CD133,
ABCG2, and Lgr5 in colon cancer cells. J Mol Med (Berl).
91:989–1000. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee Y, Ahn C, Han J, et al: The nuclear
RNase III Drosha initiates microRNA processing. Nature.
425:415–419. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee Y, Jeon K, Lee JT, Kim S and Kim VN:
MicroRNA maturation: stepwiseprocessing and subcellular
localization. EMBO J. 21:4663–4670. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vrba L, Garbe JC, Stampfer MR and Futscher
BW: Epigenetic regulation of normal human mammary cell
type-specific miRNAs. Genome Res. 21:2026–2037. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hackett JA and Surani MA: DNA methylation
dynamics during the mammalian life cycle. Philos Trans R Soc Lond B
Biol Sci. 368:201103282013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Weber B, Stresemann C, Brueckner B and
Lyko F: Methylation of human microRNA genes innormal and neoplastic
cells. Cell Cycle. 6:1001–1005. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bissels U, Wild S, Tomiuk S, et al:
Combined characterization ofmicroRNA and mRNA profiles delineates
early differentiation pathways of CD133+ and
CD34+ hematopoieticstem and progenitor cells. Stem
Cells. 29:847–857. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu L, Cai C, Wang X, Liu M, Li X and Tang
H: MicroRNA-142-3p, a new regulator of RAC1, suppresses
themigration and invasion of hepatocellular carcinoma cells. FEBS
Lett. 585:1322–1330. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang P, Bill K, Liu J, et al: MiR-155 is
a liposarcoma oncogene that targets casein kinase-1α and enhances
β-catenin signaling. Cancer Res. 72:1751–1762. 2012. View Article : Google Scholar : PubMed/NCBI
|