Introduction
In the majority of regions of the world, thyroid
cancer incidence has increased over the last few decades (1). Papillary thyroid carcinoma (PTC) is the
most common thyroid cancer, accounting for 85–95% of thyroid cancer
(1–4).
Potential risk factors for PTC include a family history of thyroid
cancer, radiation exposure and abnormal iodine intake (4–7). In recent
years, studies have demonstrated that genomic instability,
alterations in epigenetic events and subsequent inappropriate gene
expression are key in the regulation of PTC (8,9). However,
the mechanisms underlying PTC pathogenesis have not been
elucidated.
Recent studies have revealed a complex involvement
of epigenetic mechanisms in the regulation of gene expression,
including the diverse activities of noncoding RNAs (ncRNAs)
(10,11). The roles of microRNAs have been
relatively well studied in PTC, but other studies have revealed
that alterations in PTC-associated long noncoding RNAs (lncRNAs)
may lead to an altered regulation of key genes (12,13).
lncRNAs are transcript RNA molecules, >200 nucleotides in
length, that do not encode proteins. lncRNAs are among the least
well-understood of ncRNA species; however, they cannot be dismissed
as transcriptional ‘noise’. Thousands of lncRNAs have recently been
described in mammals (14). Studies
have revealed that an aberrant expression of lncRNAs results in
abnormalities in gene expression and tumorigenesis. In addition, an
altered expression of lncRNAs is a characteristic of numerous types
of cancers, which have been revealed to promote the development,
invasion and metastasis of tumors through various mechanisms
(15–17). However, study concerning lncRNA
involvement in PTC is in its infancy, and the identification of
additional PTC-associated lncRNAs and their mechanisms of action
require additional investigation.
The present study profiled the expression of lncRNAs
and mRNAs in three paired PTC and adjacent noncancerous samples via
microarray analysis, and identified a series of novel
PTC-associated lncRNAs. In addition, the present study demonstrated
that genes associated to these lncRNAs were associated with risk
factors for tumorigenesis. The present results suggest that lncRNA
expression patterns may provide novel molecular biomarkers for the
diagnosis of and methods for treating PTC.
Materials and methods
Patient samples
PTC samples and corresponding noncancerous tissue
samples were prospectively obtained from 36 patients at Tangshan
Workers' Hospital (Tangshan, China) between April 2014 and November
2014. The samples were snap-frozen in liquid nitrogen immediately
following resection. Of these samples, three pairs were used for
lncRNA microarray analysis and the remaining samples were used for
additional validation in the present study. Patient characteristics
are presented in Table I. The present
study was approved by the Institutional Ethics Review Board of
Tangshan Workers' Hospital, and all patients provided written
informed consent.
 | Table I.Clinicopathological characteristics
of patients with papillary thyroid carcinoma. |
Table I.
Clinicopathological characteristics
of patients with papillary thyroid carcinoma.
| Characteristic | n |
|---|
| Total | 36 |
| Age, years |
|
|
<45 | 15 |
|
≥45 | 21 |
| Gender |
|
|
Male | 14 |
|
Female | 22 |
| Tumor stage |
|
| I +
II | 24 |
| III +
IV | 12 |
| Tumor size, cm |
|
|
<1 | 28 |
| ≥1 | 8 |
| Lymph node
involvement |
|
|
Yes | 15 |
| No | 21 |
| Multifocal |
|
|
Yes | 16 |
| No | 20 |
RNA extraction
Total RNA was extracted using TRIzol®
Reagent (Invitrogen™; Thermo Fisher Scientific, Inc. Waltham, MA,
USA), according to the manufacturer's protocol. RNA quantification
and quality were assured by NanoDrop ND-1000 spectrophotometer
(Thermo Fisher Scientific, Inc. Wilmington, DE, USA). RNA integrity
and gDNA contamination test were assessed by denatured agarose gel
electrophoresis.
Microarray and computational
analysis
Sample labeling and array hybridization were
performed using the Agilent One-Color Microarray-Based Gene
Expression Analysis (Agilent Technologies, Inc., Santa Clara, CA,
USA), according to the manufacturer's protocol, with minor
modifications. Briefly, mRNA was purified from total RNA following
the removal of rRNA using a mRNA-ONLY™ Eukaryotic mRNA Isolation
kit (Epicentre, Madison, WI, USA). Subsequently, each sample was
amplified and transcribed into fluorescent cRNA along the entire
length of the transcripts without 3′ bias utilizing a random
priming method (Flash RNA Labeling kit; Arraystar, Rockville, MD,
USA). The labeled cRNAs were purified by RNeasy Mini kit (Qiagen,
Inc., Valencia, CA, USA). The concentration and specific activity
of the labeled cRNAs (pmol Cy3/µg cRNA) was measured by NanoDrop
ND-1000. The labeled cRNAs were hybridized onto a Human LncRNA
Microarray V3.0 (Arraystar), which is designed for 30,586 lncRNAs
and 26,109 coding transcripts. The lncRNAs were carefully
constructed using the following public transcriptome databases:
RefSeq (NCBI Reference Seqeunce Database; www.ncbi.nlm.nih.gov/refseq/); UCSC Known Genes and
Gencode (genome.ucsc.edu/index.html); and Gencode (www.gencodegenes.org/); as well as landmark
publications (18–20). Each transcript was accurately
identified by a specific exon or splice junction probe, and
subsequently the hybridized arrays were washed, fixed and scanned
using the Agilent DNA Microarray Scanner (model no., G2505C) and
Feature Extraction software version 11.0.1.1 (Agilent Technologies,
Inc.), which was used to analyze acquired array images. Quantile
normalization and subsequent data processing were performed using
the GeneSpring GX version 12.1 software (Agilent Technologies,
Inc.). The microarray analysis was performed by KangChen Bio-tech,
Inc. (Shanghai, China).
Gene ontology (GO) and pathway
analysis
GO and pathway analysis were performed to identify
the function and associated pathways of differentially expressed
mRNAs. GO analysis was derived from Gene Ontology (www.geneontology.org), which covers the following
three domains: Biological process, cellular component and molecular
function. The P-value denotes the significance of the GO term
enrichment in the differentially expressed mRNA list (cut-off,
P<0.05). Pathway analysis for the differentially expressed mRNAs
was based on the latest Kyoto Encyclopedia of Genes and Genomes
(KEGG) pathways (www.genome.jp/kegg/). This analysis was performed in
order to locate the significant enrichment pathways. The P-value
denotes the significance of the pathway associated with the latest
data from the KEGG database (cut-off, P<0.05).
Classification and subgroup
analysis
LncRNAs may be classified into different subgroups,
including lncRNAs with enhancer-like function, antisense lncRNAs
and large intergenic noncoding RNAs (lincRNAs).
To investigate the association between lncRNAs and
their coding genes, lncRNAs with enhancer-like function were
identified using the Gencode annotation of human genes (18). The selection of lncRNAs with
enhancer-like function involved the exclusion of transcripts mapped
to the exons and introns of annotated protein coding genes. These
enhancer lncRNAs and the corresponding mRNAs (<300 kb) were
integrated in order to study the potential function of the
lncRNAs.
The function of antisense lncRNAs were identified
based on the study by Faghihi and Wahlestedt (19). The present study integrated the
differentially expressed antisense lncRNAs and corresponding sense
mRNAs in order to deduce the function of the lncRNAs.
LincRNAs is a current hotspot of research. The
function of lincRNAs were identified by the present study based on
the study by Cabili et al (20). In order to deduce the function of
these differentially expressed lincRNAs, the lincRNAs and
corresponding nearby mRNAs underwent conjoint analysis.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was performed to confirm the expression of
lncRNAs identified by microarray analysis. In total, 8 lncRNAs
(upregulated: ENST00000503723, ENST00000423539, uc003tab.3,
NR_073085; downregulated: ENST00000515275, ENST00000570022,
uc003qef.1, ENST00000427243) were selected to undergo qPCR analysis
in 33 paired PTC and adjacent noncancerous samples. Total RNA was
extracted from 33 frozen tumor specimens using TRIzol Reagent and
subsequently reverse transcribed into cDNA using PrimeScript RT
reagent kit (Invitrogen™), according to the manufacturer's
protocol. DNase (1 µl; 1 U/µl) was purchased from Invitrogen™. qPCR
was performed using SYBR Green Real-Time PCR Master Mix
(Invitrogen™), according to the manufacturer's protocol, on a
MasterCycler RealPlex (Eppendorf, Hamburg, Germany). The following
cycling conditions were used: 95°C for 2 min; and 95°C for 15 sec,
57°C for 15 sec, 72°C for 1 min for a total 40 cycles. The primers
used for the 8 lncRNAs are listed in Table II and were synthesized by
Invitrogen™. Glyceraldehyde 3-phosphate dehydrogenase was used as a
reference gene with the following primers: Forward,
5′-GAAAGCCTGCCGGTGACTAA-3′ and reverse, 5′-AGGAAAAGCATCACCCGGAG-3′.
For quantitative results, the expression of each lncRNA was
represented as a fold change (FC) using 2−ΔΔCq (21). The experiment was repeated three
times.
| Table II.Long noncoding RNA gene primers used
for quantitative polymerase chain reaction. |
Table II.
Long noncoding RNA gene primers used
for quantitative polymerase chain reaction.
| Primer name | Source | Primer sequence,
5′-3′ | Product size,
bp |
|---|
|
ENST00000503723 | Gencode | F,
TTTGAGTGATGACAGTTATATG | 141 |
|
|
| R,
AACTTCTTAACCTACACCTTCT |
|
|
ENST00000423539 | Gencode | F,
CAAAGGTAATACAACTGTGAAG | 87 |
|
|
| R,
TCTCTGCAAAAGGTTACATC |
|
| uc003tab.3 | UCSC_knowngene | F,
TGCTTCTTCCATCCCTGCTTCC | 156 |
|
|
| R,
TGGGAAGTTTGCGGCTCTGAA |
|
| NR_073085 | RefSeq | F,
TTCTGGGGCTGCACCTTTGACT | 155 |
|
|
| R,
TCACCTTCTCGGCTGCCTCTG |
|
|
ENST00000515275 | Gencode | F,
GGCAGTGTTCCAAATTAGATAT | 146 |
|
|
| R,
AAGCAATTCTCAAAAGTCATG |
|
|
ENST00000570022 | Gencode | F,
CCGAGTGGAAGATGTCAGAGGAAACC | 177 |
|
|
| R,
AACCGCTCGCACCTCAGGACTG |
|
| uc003qef.1 | UCSC_knowngene | F,
CCCTCCATCTTGCCAAACTGCTGT | 180 |
|
|
| R,
GGGTGGTGGTGAAAGGCAAAAGC |
|
|
ENST00000427243 | Gencode | F,
GCCAGTGTCTTCCAATGAGCTC | 151 |
|
|
| R,
GCAGGTCTCAGGTCATCACAGG |
|
Statistical analysis
Statistical analysis was performed using Student's
t-test for the comparison of two groups in the microarray. FC and
the t-test were used to analyze the statistical significance of the
microarray results. The false discovery rate was calculated to
correct the P-value. The threshold value used to designate
differentially expressed lncRNAs and mRNAs was a fold change of
≥2.0 or ≤0.5, P<0.05. P<0.05 was considered to indicate a
statistically significant difference. Statistical analyses were
performed with SPSS version 13 software (SPSS, Inc., Chicago, IL,
USA).
Results
Expression profile of lncRNAs and
mRNAs in PTC
Using microarray analysis, the present study
examined the lncRNA and mRNA expression profiles in human paired
PTC and adjacent noncancerous tissue samples (Exp vs. Ctrl). FC
(T/N) and P-value were calculated from the normalized expression.
Microarray analysis revealed that 675 lncRNAs and 751 mRNAs were
significantly differentially expressed in PTC samples compared with
their adjacent noncancerous tissue samples (FC ≥2.0; P<0.05).
Among the lncRNAs, 312 were upregulated and 363 were downregulated.
Among the mRNAs, 499 were upregulated and 252 were downregulated.
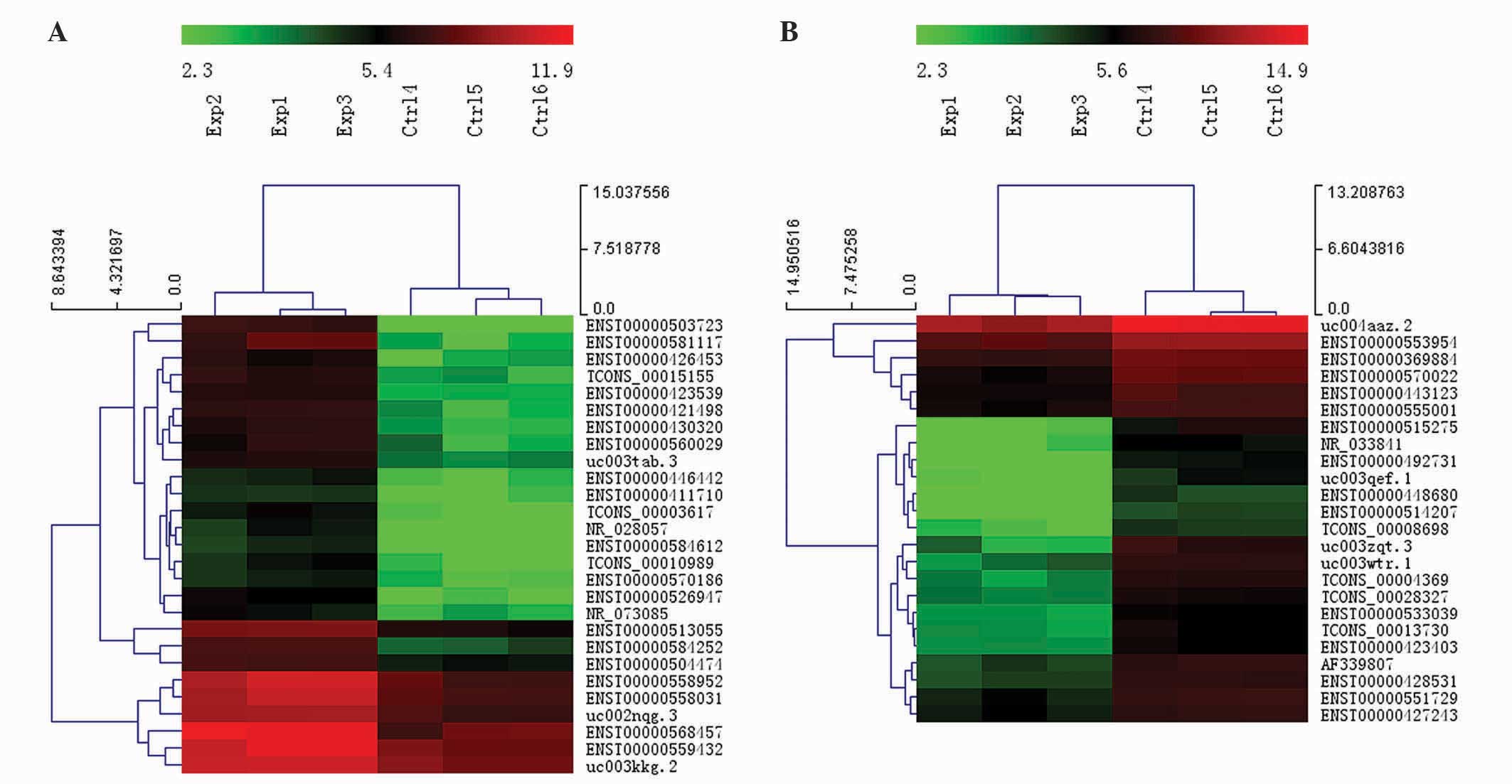
Hierarchical clustering analysis was used to arrange the
differentially expressed lncRNAs into groups based on their
expression levels. A dendrogram revealed the lncRNA expression
patterns among samples (FC ≥4.0; P<0.01; Fig. 1). ENST00000503723 (FC=24.501809) was
the most upregulated lncRNA, and ENST00000515275 (FC=14.349726) was
the most downregulated lncRNA.
Gene ontology analysis
Corresponding to the downregulated mRNAs, the total
number of differentially expressed genes consisted of 159 genes
involved in biological processes, 187 genes involved in cellular
components and 170 genes involved in molecular functions. The
highest enriched GOs targeted by downregulated transcripts were
negative regulation of transporter activity (biological process),
myofibril (cellular component) and oxygen transporter activity
(molecular function).
Corresponding to the upregulated mRNAs, the total
number of differentially expressed genes consisted of 362 genes
involved in biological processes, 376 genes involved in cellular
components and 359 genes involved in molecular functions. The
highest enriched GOs targeted by the upregulated transcripts were
response to stimulus (biological process), plasma membrane
(cellular component) and protein binding (molecular function).
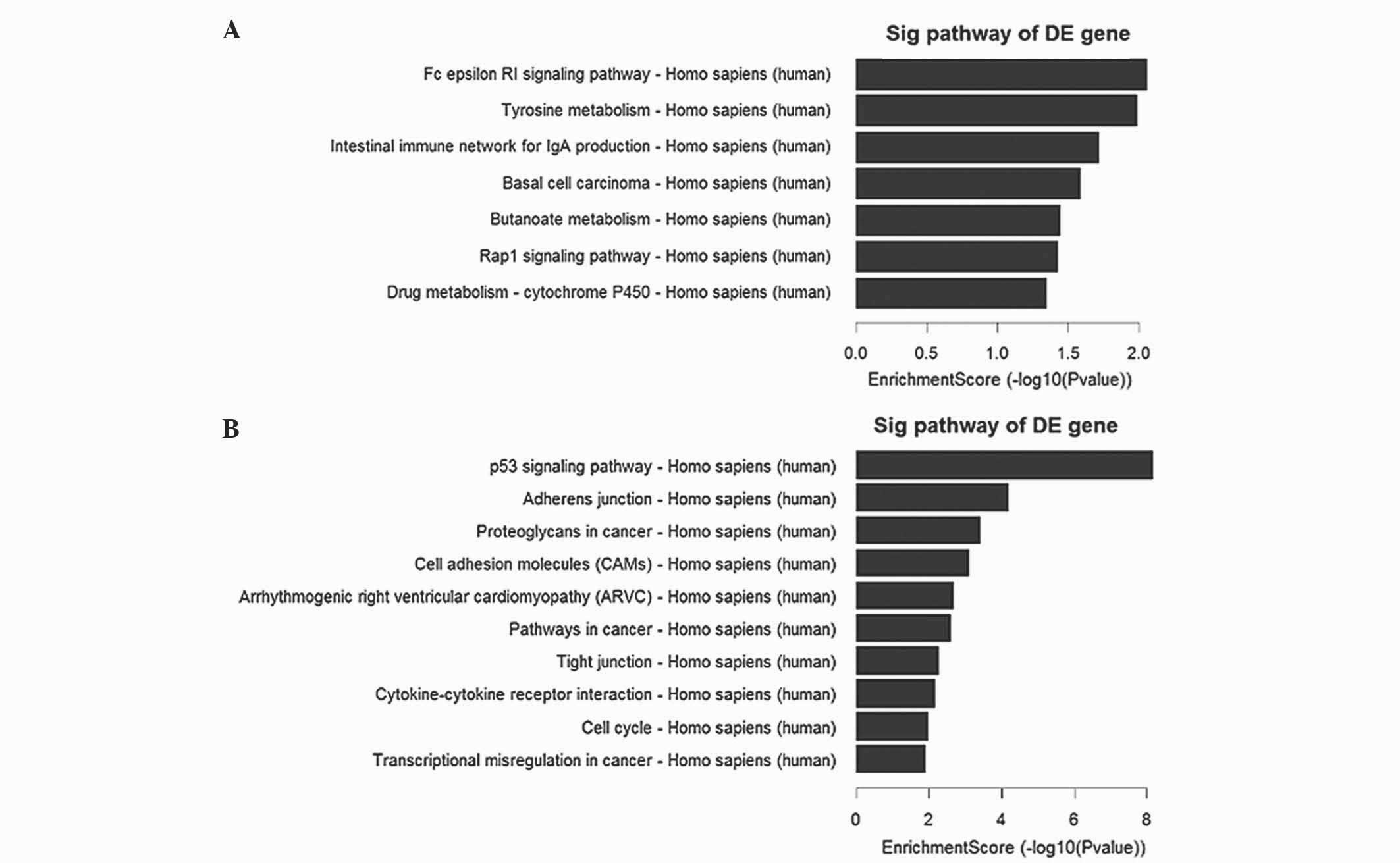
Pathway analysis
Pathway analysis indicated that there were 7
downregulated pathways (Fig. 2A), and
the most enriched network was ‘Fc epsilon RI signaling
pathway-Homo sapiens (human)’, which was composed of 4
targeted genes. Furthermore, the analysis revealed that of the 29
upregulated pathways (Fig. 2B)
identified, the most enriched network was ‘p53 signaling
pathway-Homo sapiens (human)’, which was composed of 13
targeted genes.
Classification and subgroup
analysis
LncRNA classification and subgroup analysis
indicated that 7 pairs of enhancer-like lncRNA-mRNA (Table III), 9 pairs of antisense
lncRNA-mRNA (Table IV) and 45 pairs
of lincRNA-mRNA (Table V) were
differentially expressed between PTC and their paired adjacent
noncancerous samples.
| Table III.Enhancer-like lncRNA-mRNA and nearby
genes. |
Table III.
Enhancer-like lncRNA-mRNA and nearby
genes.
| Seqname | Regulation,
lncRNAs | Genome
assocation | Nearby gene
seq. | Nearby gene | Regulation,
mRNAs |
|---|
|
ENST00000418242 | Up | Downstream | NM_001122674 | ABCD3 | Up |
|
ENST00000423539 | Up | Upstream | NM_020651 | PELI1 | Up |
|
ENST00000429328 | Up | Upstream | NM_178134 | CYP4Z1 | Down |
|
ENST00000430945 | Up | Downstream | NM_014057 | OGN | Down |
|
ENST00000430945 | Up | Upstream | NM_004560 | ROR2 | Down |
|
ENST00000442526 | Down | Upstream | NM_018590 | CSGALNACT2 | Up |
| NR_024160 | Down | Upstream | NM_001937 | DPT | Down |
| Table IV.Antisense lncRNA-mRNA and nearby
genes. |
Table IV.
Antisense lncRNA-mRNA and nearby
genes.
| Seqname | Regulation,
lncRNAs | Genome
assocaition | Nearby gene
seq. | Nearby gene | Regulation,
mRNAs |
|---|
|
ENST00000506160 | Down | Intronic
antisense | NM_000667 | ADH1A | Down |
|
ENST00000541892 | Up | Intronic
antisense | NM_001001395 | LMO3 | Up |
|
ENST00000550214 | Up | Intronic
antisense | NM_001651 | AQP5 | Up |
| NR_036580 | Up | Intronic
antisense | NM_001178036 | DPP10 | Up |
|
ENST00000418335 | Up | Natural
antisense | NM_001935 | DPP4 | Up |
|
ENST00000419425 | Up | Natural
antisense | NM_006449 | CDC42EP3 | Up |
|
ENST00000512486 | Up | Natural
antisense | NM_014899 | RHOBTB3 | Up |
| NR_033252 | Down | Natural
antisense | NM_001482 | GATM | Down |
| uc001pnn.3 | Down | Natural
antisense | NM_181351 | NCAM1 | Down |
| Table V.Large intergenic noncoding RNA-mRNA
and nearby genes. |
Table V.
Large intergenic noncoding RNA-mRNA
and nearby genes.
| Seq. name | Regulation,
lncRNAs | Genome
association | Nearby gene
seq. | Nearby gene | Regulation,
mRNAs |
|---|
| TCONS_00001038 | Down | Upstream | NM_001122674 | ABCD3 | Up |
| TCONS_00003617 | Up | Upstream | NM_022823 | FNDC4 | Up |
| TCONS_00003757 | Up | Upstream | NM_001042437 | ST3GAL5 | Up |
| TCONS_00003757 | Up | Upstream | NM_198843 | SFTPB | Up |
| TCONS_00004186 | Up | Downstream | NM_022823 | FNDC4 | Up |
| TCONS_00004538 | Up | Downstream | NM_012223 | MYO1B | Up |
| TCONS_00004570 | Up | Downstream | NM_001079526 | IKZF2 | Up |
| TCONS_00005191 | Down | Downstream | NM_001042437 | ST3GAL5 | Up |
| TCONS_00005191 | Down | Downstream | NM_198843 | SFTPB | Up |
| TCONS_00005192 | Down | Downstream | NM_001042437 | ST3GAL5 | Up |
| TCONS_00005192 | Down | Downstream | NM_198843 | SFTPB | Up |
| TCONS_00005258 | Down | Upstream | NM_000888 | ITGB6 | Up |
| TCONS_00005258 | Down | Upstream | NM_001007267 | PLA2R1 | Down |
| TCONS_00005258 | Down | Upstream | NM_007366 | PLA2R1 | Down |
| TCONS_00005259 | Down | Upstream | NM_000888 | ITGB6 | Up |
| TCONS_00005259 | Down | Upstream | NM_001007267 | PLA2R1 | Down |
| TCONS_00005259 | Down | Upstream | NM_007366 | PLA2R1 | Down |
| TCONS_00006023 | Up | Upstream | NM_004635 | MAPKAPK3 | Up |
| TCONS_00006284 | Down | Upstream | NM_014996 | PLCH1 | Down |
| TCONS_00006733 | Down | Downstream |
ENST00000072516 | IL1RAP | Up |
| TCONS_00006733 | Down | Downstream | NM_134470 | IL1RAP | Up |
| TCONS_00008649 | Down | Downstream |
ENST00000265000 | GALNT7 | Up |
| TCONS_00008698 | Down | Downstream |
ENST00000448662 | SORBS2 | Down |
| TCONS_00008698 | Down | Downstream | NM_001145670 | SORBS2 | Down |
| TCONS_00008698 | Down | Downstream | NM_003603 | SORBS2 | Down |
| TCONS_00008738 | Down | Downstream | NM_006527 | SLBP | Up |
| TCONS_00008739 | Down | Downstream | NM_006527 | SLBP | Up |
| TCONS_00010526 | Up | Upstream | NM_001112724 | STK32A | Up |
| TCONS_00010526 | Up | Upstream | NM_145001 | STK32A | Up |
| TCONS_00010794 | Up | Upstream | NM_004887 | CXCL14 | Up |
| TCONS_00012533 | Up | Upstream | NM_001122769 | LCA5 | Up |
| TCONS_00012535 | Up | Upstream | NM_001122769 | LCA5 | Up |
| TCONS_00014681 | Up | Downstream | NM_020130 | C8orf4 | Up |
| TCONS_00016073 | Down | Upstream | NM_014057 | OGN | Down |
| TCONS_00016117 | Up | Upstream | NM_021224 | ZNF462 | Up |
| TCONS_00016354 | Up | Upstream | NM_000700 | ANXA1 | Up |
| TCONS_00018333 | Up | Upstream | NM_004419 | DUSP5 | Up |
| TCONS_00018333 | Up | Upstream | NM_005962 | MXI1 | Down |
| TCONS_00019600 | Down | Upstream | NM_001584 | MPPED2 | Down |
| TCONS_00019602 | Down | Downstream | NM_001584 | MPPED2 | Down |
| TCONS_00020396 | Down | Downstream | NM_181847 | AMIGO2 | Up |
| TCONS_00024026 | Down | Downstream | NM_024708 | ASB7 | Up |
| TCONS_00027184 | Down | Upstream | NM_031209 | QTRT1 | Down |
| TCONS_00027673 | Down | Upstream | NM_031209 | QTRT1 | Down |
| TCONS_00029753 | Up | Upstream | NM_001193414 | TUBA8 | Down |
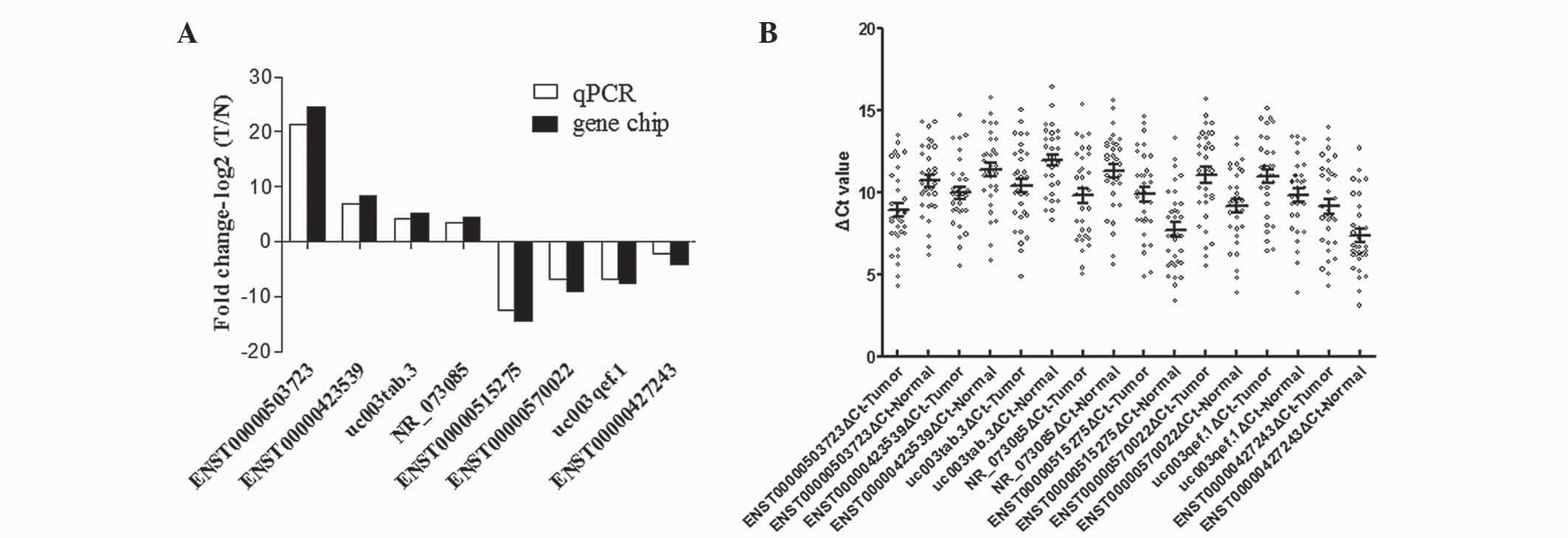
RT-qPCR results
To verify the microarray data, 8 differentially
expressed lncRNAs, including 4 upregulated (ENST00000503723,
ENST00000423539, uc003tab.3 and NR_073085) and 4 downregulated
(ENST00000515275, ENST00000570022, uc003qef.1 and ENST00000427243)
lncRNAs, were randomly selected for RT-qPCR in 33 pairs of PTC and
adjacent noncancerous tissues. The results were consistent with the
microarray data (Fig. 3).
Discussion
PTC is a common malignant tumor of the endocrine
system that occurs more often in women than men (22). The incidence of PTC has increased
globally for several decades. The tumorigenesis of PTC is a complex
biological process characterized by various molecular
abnormalities. Over the past few decades, the molecular mechanism
of PTC has been extensively investigated; however, the exact
pathogenesis of this disease remains unclear. Increasing evidence
suggests that lncRNAs may be significant in regulating gene
expression. LncRNA have gained considerable attention and have
become the frontiers of ncRNA research. Although lncRNAs were
initially indicated to be transcriptional noise, studies suggest
that they may be key in cellular development and human diseases
(23,24), including lnc-IL7R in the inflammatory
response (25), and HOTAIR and Gas5
in breast tumors and metastases (16,26,27).
Aberrant expression of lncRNAs were also identified in the present
study in PTC using microarray data, including HOTAIR, which was
downregulated; however, this was not statistical significant. Of
all the functions of lncRNAs, the most important is their
involvement in tumorigenesis. However, the role of lncRNAs in tumor
pathogenesis and development is less well characterized.
The present study investigated the lncRNA expression
profiles of PTC using microarray analysis and revealed that lncRNA
expression levels were altered compared to adjacent noncancerous
tissues. From the microarray expression profiles, 312 upregulated
lncRNAs and 363 downregulated lncRNAs were significantly
differentially expressed in all PTC samples. The most upregulated
lncRNA was ENST00000503723, and the most downregulated lncRNA was
ENST00000515275. This result suggests that ENST00000503723 and
ENST00000515275 may contribute to the development of PTC; however,
additional study concerning the biological function of these two
lncRNAs requires confirmation. To validate the microarray profiling
expression data, the expression of 8 randomly selected lncRNAs were
examined in PTC and adjacent noncancerous samples by RT-qPCR. The
RT-qPCR results confirmed the data from the microarray.
The present results demonstrated an altered
expression profile of lncRNAs in PTC. Additional analysis by the
present study highlighted that numerous genes associated with these
lncRNAs were associated with various types of cancer, including
ENST00000427243, uc003qef.1 and NR_073085. ENST00000427243 was
significantly downregulated in PTC and was identified to be
associated with breast cancer anti-estrogen resistance protein 3
(BCAR3) gene. BCAR3 overexpression results in enhanced cancer cell
migration and colocalization (28).
By contrast, BCAR3 depletion inhibits cancer cell migration and
invasion (29). In addition, BCAR3
functions as a positive regulator in invasive breast cancer cells
(30) and acts as a putative
suppressor of breast cancer progression (31). Therefore, there may be an association
between ENST00000427243 lncRNA and BCAR3 in the development of PTC,
and BCAR3 may aid in the understanding of PTC and lncRNAs at the
transcriptional level.
uc003qef.1 was also identified to be significantly
downregulated in the present results. Its associated gene was eyes
absent homolog 4 (EYA4). Decreased EYA4 expression is not only
associated with poor survival in sporadic lung cancer, but also
EYA4 single-nucleotide polymorphisms have been associated with an
increased familial cancer risk (32).
The present study concludes that uc003qef.1 may play an important
role with EYA4 in the familial cancer risk of PTC.
NR_073085 was identified to be significantly
upregulated in PTC samples by the present study. NR_073085 was
associated with cysteine-rich protein 2 (CRIP2), which was
identified as an important candidate gene capable of functionally
suppressing tumor formation (33).
CRIP2 is significantly downregulated in tumors, and an
overexpression of CRIP2 may induce cancer cell apoptosis (33). Furthermore, CRIP2 acts as a
transcriptional repressor of the nuclear factor-κB-mediated
proangiogenic cytokine expression, and thereby functionally
inhibits tumor formation and angiogenesis (34). NR_073085 may be significant in the
regulation of CRIP2 during tumor progression. The association
between NR_073085 and PTC requires additional experimental
verification.
The microarray-based profiling performed by the
present study identified 751 mRNAs with a differential expression
in PTC. GO analysis and pathway analysis indicated that there were
7 downregulated and 29 upregulated pathways in PTC compared with
adjacent non-tumorous tissues. Among the upregulated pathways, the
p53 signaling pathway was the most enriched network. Lan et
al (35) also demonstrated that
the p53 signaling pathway was differentially expressed in PTC in a
mRNA expression profile study. It is known that p53 plays an
important tumor suppressive role during thyroid carcinoma
progression (36). Signaling pathways
that use lncRNAs to regulate the growth and metastasis of tumors
require additional study to be identified.
Furthermore, lncRNA classification and subgroup
analysis performed by the present study indicated 61 pairs of
lncRNA-mRNA were differentially expressed in PTC. Certain nearby
proteins of these lncRNAs were associated with inflammation. It is
commonly accepted that cancer is associated with inflammation, and
an intense immune infiltrate is often observed with PTC, which may
be critical in the regulation of carcinogenesis and carcinoma
progression (37). The present study
identified ENST00000423539 as a significantly upregulated enhancer
lncRNA in PTC, and the nearby protein was E3 ubiquitin-protein
ligase pellino homolog 1 (Peli1). Peli1 regulates Toll-like
receptor pathway signaling by promoting the degradation of tumor
necrosis factor receptor-associated factor 3, which is a potent
inhibitor of mitogen-activated protein kinase activation and gene
induction (38). In addition, Peli1
is a target of microRNA-21 (39),
which is significantly downregulated in PTC tissues (40). ENST00000423539 may exert its functions
through interactions with coding transcripts in PTC. The
inflammatory pathways of cancer-associated inflammation is critical
for the identification of novel target molecules that may lead to
improved diagnosis and treatment.
In conclusion, the present study demonstrated the
existence of a significantly altered lncRNA and mRNA expression
profile in PTC. Dysregulation of these lncRNAs may be important in
the development, invasion and metastasis of tumors. Additional
study is required to determine whether these alterations are
indirectly due to genotype, familial cancer risk, inflammatory
states or a combination of these cancer risk factors. Further
molecular mechanism studies may aid in the elucidation of the exact
roles of these lncRNAs in PTC.
Acknowledgements
The authors would like to thank all the donors that
donated to the Microarray Service at KangChen Bio-tech, Inc.
References
|
1
|
La Vecchia C, Malvezzi M, Bosetti C,
Garavello W, Bertuccio P, Levi F and Negri E: Thyroid cancer
mortality and incidence: A global overview. Int J Cancer.
136:2187–2195. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang L, Zheng R, Wang N, Zhang S and Chen
W: Analysis of incidence and mortality of thyroid cancer in China,
2010. Zhonghua Yu Fang Yi Xue Za Zhi. 48:663–668. 2014.(In
Chinese). PubMed/NCBI
|
|
4
|
Pelizzo MR, Boschin Merante I, Toniato A,
Pagetta C, Casal Ide E, Mian C and Rubello D: Diagnosis, treatment,
prognostic factors and long-term outcome in papillary thyroid
carcinoma. Minerva Endocrinol. 33:359–379. 2008.PubMed/NCBI
|
|
5
|
Richards ML: Familial syndromes associated
with thyroid cancer in the era of personalized medicine. Thyroid.
20:707–713. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Galleani J, Miranda C, Pierotti MA and
Greco A: H2AX phosphorylation and kinetics of radiation-induced DNA
double strand break repair in human primary thyrocytes. Thyroid.
19:257–264. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Leux C, Truong T, Petit C,
Baron-Dubourdieu D and Guénel P: Family history of malignant and
benign thyroid diseases and risk of thyroid cancer: A
population-based case-control study in New Caledonia. Cancer Causes
Control. 23:745–755. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Santos JC, Bastos AU, Cerutti JM and
Ribeiro ML: Correlation of MLH1 and MGMT expression and promoter
methylation with genomic instability in patients with thyroid
carcinoma. BMC Cancer. 13:792013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Neta G, Brenner AV, Sturgis EM, Pfeiffer
RM, Hutchinson AA, Aschebrook-Kilfoy B, Yeager M, Xu L, Wheeler W,
Abend M, et al: Common genetic variants related to genomic
integrity and risk of papillary thyroid cancer. Carcinogenesis.
32:1231–1237. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Taft RJ, Pang KC, Mercer TR, Dinger M and
Mattick JS: Non-coding RNAs: Regulators of disease. J Pathol.
220:126–139. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Djebali S, Davis CA, Merkel A, Dobin A,
Lassmann T, Mortazavi A, Tanzer A, Lagarde J, Lin W, Schlesinger F,
et al: Landscape of transcription in human cells. Nature.
489:101–108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jendrzejewski J, He H, Radomska HS, Li W,
Tomsic J, Liyanarachchi S, Davuluri RV, Nagy R and de la Chapelle
A: The polymorphism rs944289 predisposes to papillary thyroid
carcinoma through a large intergenic noncoding RNA gene of tumor
suppressor type. Proc Natl Acad Sci USA. 109:8646–8651. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Y, Guo Q, Zhao Y, Chen J, Wang S, Hu
J and Sun Y: BRAF-activated long non-coding RNA contributes to cell
proliferation and activates autophagy in papillary thyroid
carcinoma. Oncol Lett. 8:1947–1952. 2014.PubMed/NCBI
|
|
14
|
Dinger ME, Pang KC, Mercer TR, Crowe ML,
Grimmond SM and Mattick JS: NRED: A database of long noncoding RNA
expression. Nucleic Acids Res. 37(Database Issue): D122–D126. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu M, Chen Q, Liu X, Sun Q, Zhao X, Deng
R, Wang Y, Huang J, Xu M, Yan J and Yu J: lncRNA H19/miR-675 axis
represses prostate cancer metastasis by targeting TGFBI. FEBS J.
281:3766–3775. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ørom UA, Derrien T, Beringer M, Gumireddy
K, Gardini A, Bussotti G, Lai F, Zytnicki M, Notredame C, Huang Q,
et al: Long noncoding RNAs with enhancer-like function in human
cells. Cell. 143:46–58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Faghihi MA and Wahlestedt C: Regulatory
roles of natural antisense transcripts. Nat Rev Mol Cell Biol.
10:637–643. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cabili MN, Trapnell C, Goff L, Koziol M,
Tazon-Vega B, Regev A and Rinn JL: Integrative annotation of human
large intergenic noncoding RNAs reveals global properties and
specific subclasses. Genes Dev. 25:1915–1927. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu G, Chen J, Pan Q, Huang K, Pan J, Zhang
W, Chen J, Yu F, Zhou T and Wang Y: Long noncoding RNA expression
profiles of lung adenocarcinoma ascertained by microarray analysis.
PLoS One. 9:e1040442014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kweon SS, Shin MH, Chung IJ, Kim YJ and
Choi JS: Thyroid cancer is the most common cancer in women, based
on the data from population-based cancer registries, South Korea.
Jpn J Clin Oncol. 43:1039–1046. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wilusz JE, Sunwoo H and Spector DL: Long
noncoding RNAs: Functional surprises from the RNA world. Genes Dev.
23:1494–1504. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cui H, Xie N, Tan Z, Banerjee S,
Thannickal VJ, Abraham E and Liu G: The human long noncoding RNA
lnc-IL7R regulates the inflammatory response. Eur J Immunol.
44:2085–2095. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bhan A, Hussain I, Ansari KI, Bobzean SA,
Perrotti LI and Mandal SS: Bisphenol-A and diethylstilbestrol
exposure induces the expression of breast cancer associated long
noncoding RNA HOTAIR in vitro and in vivo. J Steroid Biochem Mol
Biol. 141:160–170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mourtada-Maarabouni M, Pickard MR, Hedge
VL, Farzaneh F and Williams GT: GAS5, a non-protein-coding RNA,
controls apoptosis and is downregulated in breast cancer. Oncogene.
28:195–208. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wallez Y, Riedl SJ and Pasquale EB:
Association of the breast cancer antiestrogen resistance protein 1
(BCAR1) and BCAR3 scaffolding proteins in cell signaling and
antiestrogen resistance. J Biol Chem. 289:10431–10444. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Schrecengost RS, Riggins RB, Thomas KS,
Guerrero MS and Bouton AH: Breast cancer antiestrogen resistance-3
expression regulates breast cancer cell migration through promotion
of p130Cas membrane localization and membrane ruffling. Cancer Res.
67:6174–6182. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wilson AL, Schrecengost RS, Guerrero MS,
Thomas KS and Bouton AH: Breast cancer antiestrogen resistance 3
(BCAR3) promotes cell motility by regulating actin cytoskeletal and
adhesion remodeling in invasive breast cancer cells. PLoS One.
8:e656782013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guo J, Canaff L, Rajadurai C, Fils-Aimé N,
Tian J, Dai M, Korah J, Villatoro M, Park M, Ali S and Lebrun JJ:
Breast cancer anti-estrogen resistance-3 inhibits transforming
growth factor-β/Smad signaling and associates with favorable breast
cancer disease outcomes. Breast Cancer Res. 16:4762014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wilson IM, Vucic EA, Enfield KS, Thu KL,
Zhang YA, Chari R, Lockwood WW, Radulovich N, Starczynowski DT,
Banáth JP, et al: EYA4 is inactivated biallelically at a high
frequency in sporadic lung cancer and is associated with familial
lung cancer risk. Oncogene. 33:4464–4473. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lo PH, Ko JM, Yu ZY, Law S, Wang LD, Li
JL, Srivastava G, Tsao SW, Stanbridge EJ and Lung ML: The LIM
domain protein, CRIP2, promotes apoptosis in esophageal squamous
cell carcinoma. Cancer Lett. 316:39–45. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cheung AK, Ko JM, Lung HL, Chan KW,
Stanbridge EJ, Zabarovsky E, Tokino T, Kashima L, Suzuki T, Kwong
DL, et al: Cysteine-rich intestinal protein 2 (CRIP2) acts as a
repressor of NF-kappaB-mediated proangiogenic cytokine
transcription to suppress tumorigenesis and angiogenesis. Proc Natl
Acad Sci USA. 108:8390–8395. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lan X, Zhang H, Wang Z, Dong W, Sun W,
Shao L, Zhang T and Zhang D: Genome-wide analysis of long noncoding
RNA expression profile in papillary thyroid carcinoma. Gene.
569:109–117. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
McFadden DG, Vernon A, Santiago PM,
Martinez-McFaline R, Bhutkar A, Crowley DM, McMahon M, Sadow PM and
Jacks T: p53 constrains progression to anaplastic thyroid carcinoma
in a Braf-mutant mouse model of papillary thyroid cancer. Proc Natl
Acad Sci USA. 111:E1600–E1609. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liotti F, Visciano C and Melillo RM:
Inflammation in thyroid oncogenesis. Am J Cancer Res. 2:286–297.
2012.PubMed/NCBI
|
|
38
|
Xiao Y, Jin J, Chang M, Chang JH, Hu H,
Zhou X, Brittain GC, Stansberg C, Torkildsen Ø, Wang X, et al:
Peli1 promotes microglia-mediated CNS inflammation by regulating
Traf3 degradation. Nat Med. 19:595–602. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Marquez RT, Wendlandt E, Galle CS, Keck K
and McCaffrey AP: MicroRNA-21 is upregulated during the
proliferative phase of liver regeneration, targets Pellino-1, and
inhibits NF-kappaB signaling. Am J Physiol Gastrointest Liver
Physiol. 298:G535–G541. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Huang Y, Liao D, Pan L, Ye R, Li X, Wang
S, Ye C and Chen L: Expressions of miRNAs in papillary thyroid
carcinoma and their associations with the BRAFV600E mutation. Eur J
Endocrinol. 168:675–681. 2013. View Article : Google Scholar : PubMed/NCBI
|