Introduction
Wilms' tumor (WT), also known as nephroblastoma, is
the most common pediatric renal tumor, arising mainly in children
<5 years of age. The tumor is believed to have an embryonic
kidney cell origin (1). The current
treatment protocol for WTs, as indicated by the International
Society of Pediatric Oncology, involves the use of pre-operative
chemotherapy followed by nephrectomy, however, such chemotherapy is
not of equal benefit to all patients (2). It has been noted that
differentially-expressed genes and microRNA (miRNA/miR) have a
large impact on the development and therapy of WTs, and that
associated genes and miRNA have less effect on WTs.
Genes include transcription factors (TFs), target
genes and host genes, which play different roles in the network
process. TFs and miRNAs are prominent gene expression regulators;
gene regulatory factors are mainly formed from TFs and miRNAs,
which control the developmental processes of genomic information in
multicellular genomes by specifying when and where the genes are to
be expressed (3,4). Camas et al indicated that TFs are
proteins that bind specific short DNA sequences adjacent to the
genes whose transcription they regulate (5). miRNAs are a class of small non-coding
RNA sequences that contribute to cancer development and
metastasis.
miRNAs pair to the messages of protein-coding genes
to target these genes for post-transcriptional repression (6).
miRNA locates at host genes. Rodriguez et al
suggested that the transcriptional patterns of all miRNA host genes
were selected from a range of sources, highlighting the spatial,
temporal and physiological regulation of miRNA expression, which
strongly suggested parallel transcription of the miRNA and the host
transcripts, and the fact that the two differing miRNA
transcription classes (exonic and intronic) may require slightly
different biogenesis mechanisms (7).
A large amount of data on genes and miRNAs has been
obtained from the experimentation of biologists and scientists.
However, the majority of the experiments were based on one factor
(gene or miRNA), and the regulatory association among the data
remains unclear. In order to research WTs from a global
perspective, the present study focused on the networks of TFs,
miRNAs, target genes and host genes, and their control mechanisms.
The differentially-expressed network involves
differentially-expressed genes and miRNAs in the development and
progression of WTs; the host genes where differentially-expressed
miRNAs locate were considered as differentially expressed. The
related network contains the associated genes and miRNAs in WT. The
association between the genes and miRNA was analyzed in detail,
comparing the similarities and differences. The vital linkages, in
particular, were extracted for selective analysis. The study aimed
to provide a greater understanding with regard to the pathogenesis
of WTs.
Materials and methods
Extraction of TFs and miRNA regulatory
associations
An experimentally validated dataset on TFs and the
miRNAs that are regulated by them were gathered from TransmiR
(8). The TransmiR dataset is a
manually collated collection of regulatory associations between TFs
and miRNAs. This complete data was considered as dataset S1.
Extraction of miRNAs and target gene
regulatory associations
An experimentally validated dataset on miRNAs and
their target genes was gathered from Tarbase5.0 (http://diana.imis.athena-innovation.gr/DianaTools/index.php?r=tarbase/index)
and miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/). To unify symbols,
the official National Center for Biotechnology Information (NCBI)
symbols of genes and miRNAs were used. The complete dataset was
considered as dataset S2.
Extraction of miRNAs and host gene
regulatory relations
miRNAs and their host genes were collected from the
NCBI and miRBase (9). Official NCBI
symbols were used for miRNAs and host genes. This dataset was
regarded as dataset S3.
Extracting data of genes and
miRNAs
Differentially-expressed miRNAs and associated
miRNAs with regard to WT were gathered from the pertinent
literature. The differentially-expressed miRNAs and associated
miRNAs were considered as dataset S4 and S5, respectively.
Differentially-expressed genes were manually
collected from the NCBI Single Nucleotide Polymorphism database
(http://www.ncbi.nlm.nih.gov/snp/) and
from the pertinent literature. Related genes were gathered from the
GeneCards database (http://www.genecards.org/) and from the pertinent
literature in the Web of Science (http://wokinfo.com/). The related genes were those to
take part in the development, progression, radical therapy and
clinical outcome of WT. In addition, P-match algorithm (10) was used to extract predicted TFs. The
University of California Santa Cruz database (11) indicates that target genes of
differentially-expressed miRNAs have 1,000 nt promoter region
sequences. P-match combining pattern and weight matrix approaches
was used to identify TF binding sites (TFBSs) in 1,000- and
5,000-nt promoter region sequences, and the TFBSs were mapped onto
the promoter regions of targets. The matrix library of P-match, as
well as sets of known TFBSs collected in TRANSFAC (www.gene-regulation.com/pub/databases.html)
provide the possibility of searching for a large variety of
different TFBSs. Differentially-expressed genes and related genes
were placed into the S6 and S7 datasets separately.
Three levels of network
construction
All the regulatory associations in three levels of
network (the differentially-expressed network, the related network
and the global network) are based on TFs, miRNAs, target genes and
host genes. Specifically, the datasets of S1, S2 and S3 contain all
the associations among them, and furthermore, the global network is
the combination of the three datasets.
The related network is a network that contains
related genes and related miRNAs, and the regulatory relations
between them. This was obtained by mapping S4 and S6 onto S1, S2
and S3.
The differentially-expressed network is the smallest
and most important of the three networks. The factors it contains
are differentially expressed. Differentially-expressed genes and
miRNAs acted as primary factors to build up the network. The
regulatory pathways were obtained by mapping S5 and S7 onto S1, S2
and S3.
Results
Differentially-expressed network of
WTs
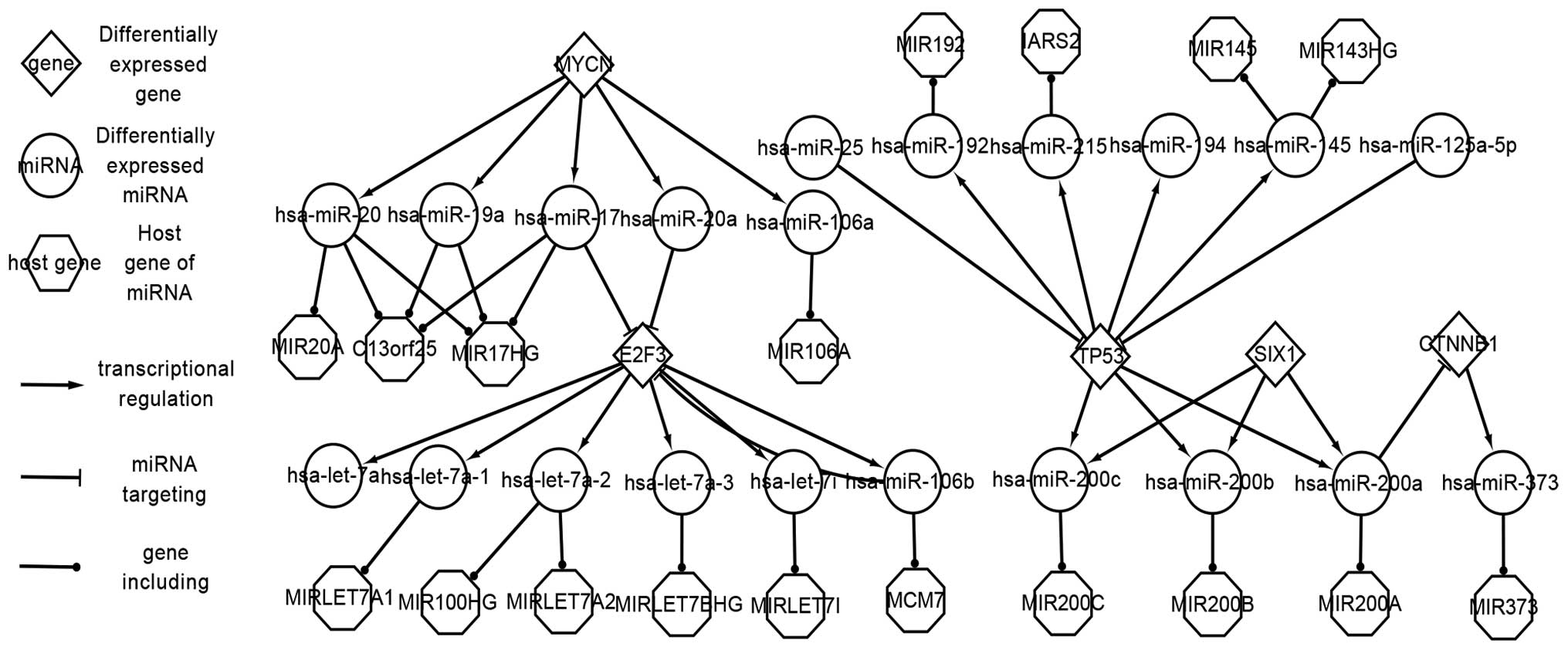
Numerous significant regulatory pathways and factors
are shown in Fig. 1; 5 TFs (E2F3,
MYCN, SIX1, TP53 and CTNNB1), 38 differentially-expressed miRNAs,
33 host genes and 18 target genes (TFs targeted by miRNAs were also
regarded as target genes) were determined. A number of regulatory
pathways were highlighted in the complex network.
The associations of factors can be classified into
three classes: i) miRNAs are regulated by TFs; ii) miRNAs target
the target genes; and iii) miRNAs locate at host genes. The network
is so complex for the following reasons: Firstly, certain TFs also
act as target genes. For example, TP53 regulates hsa-miR-145,
hsa-miR-192, hsa-miR-194, hsa-miR-215, hsa-miR-200a, hsa-miR-200b
and hsa-miR-200c, but is also targeted by hsa-miR-25 and
hsa-miR-125a-5p. Secondly, miRNAs and genes may form feed-forward
loops (FFLs); for example, hsa-miR-106b targets E2F3, but is also
regulated by E2F3. Thirdly, a single miRNA can locate at more than
one host gene, such as hsa-let-7c, which locates at MIRLET7C,
LINC00478 and C21orf34. Multiple different miRNAs can also locate
at one host gene; for example, hsa-miR-19a, hsa-miR-17 and
hsa-miR-20 are all located at C13orf26. In addition, IGF2 is
overexpressed in WT (12). However,
IGF2 acts as a target gene and a host gene; hsa-let-7a targets IGF2
and hsa-miR-483 locates at IGF2. Host genes together with their
miRNAs assist in our comprehension of the development of WTs.
Associations among genes can be summarized as
follows: Genes affect other genes through miRNAs. For instance,
hsa-miR-145 targets IGF1R and is also regulated by TP53. This
indicates that TP53 could produce an effect on IGF1R through
hsa-miR-145. Combined experimentally validated factors and their
actions reveal the pathogenesis of WTs.
Transcriptional network of TFs and
miRNAs in WT
The transcriptional network is part of a
differentially-expressed network. TFs are proteins that regulate
miRNAs. The present study focused upon TFs, miRNAs and the
regulatory associations between them. In total, 5 TFs (E2F3, TP53,
SIX1, CTNNB1 and MYCN) play a vital role in WTs (Fig. 2).
E2F3 is the core factor in the transcriptional
network. Cooper et al indicated that the E2F3 TF has an
established role in controlling cell cycle progression and
proliferation (13). Furthermore,
E2F3 is overexpressed in WTs, implying that it may be a therapeutic
target against the tumors (14).
Hsa-miR-106b, hsa-miR-17 and hsa-miR-20a target E2F3, and E2F3
regulates 6 miRNAs (hsa-miR-106b, hsa-let-7a, has-let-7i,
has-let-7a-1, hsa-let-7a-2 and hsa-let-7a-3). In other words, these
upstream miRNAs affect downstream miRNAs through E2F3, which means
that certain TFs can be regarded as bridges to connect predecessors
and successors of miRNAs, and the expression of miRNAs in the
important regulatory pathways when they have changed. E2F3 is such
a vital factor that it should be a critical focus of attention.
SIX1 has certain differences to E2F3. SIX1 has been
shown to be mainly highly expressed in the blastemal tissue
components of WT (15). SIX1 was
included in the network with other TFs in order to analyze WT in a
systematic manner. No differentially-expressed miRNAs target SIX1,
and SIX1 regulates hsa-miR-200a, hsa-miR-200b and hsa-miR-200c,
which indicates that it may be the first factor to affect the
expression of miRNAs in this regulatory pathway.
Related network of WTs
Numerous regulatory associations are shown in
Fig. 3. The data in the related
network include associated genes and associated miRNAs, including
differentially-expressed genes and miRNAs. The related genes and
miRNAs that have no associations with other factors are not shown
in the figure. The present study focused on analyzing the pathways
between the genes and miRNAs. The differentially-expressed network
is clearly contained within the related network. Fig. 3 lists 18 TFs (such as LIN28B, MYC and
MYCN), including five differentially-expressed TFs and three
predicted TFs (ZEB1, NFKB1 and YY1), as well as 64 miRNAs (such as
hsa-miR-100, hsa-miR-125 and hsa-miR-106a).
The related TFs LIB28B and MYC play prominent roles
in the related network; they have the most regulatory associations.
LIN28B regulates 13 miRNAs, including hsa-let-7a, hsa-let-7b and
hsa-let-7c, and these miRNAs target certain other target genes,
which means that LIN28B affects other target genes by regulating
miRNAs. It is significant that LIN28B is targeted by no miRNAs. MYC
regulates 23 related miRNAs, including hsa-let-7a, hsa-let-7b,
hsa-let-7c and hsa-miR-19a, and is also targeted by 9 related
miRNAs, including hsa-let-7a, hsa-let-7c and hsa-let-7g. The STAT1
gene is represented using a unique square; this gene was obtained
from GeneCards and P-match.
ZEB1, NFKB1 and YY1 are three related TFs that were
obtained from P-match. ZEB1 is targeted by 5 miRNAs (hsa-miR-200a,
hsa-miR-200b, hsa-miR-200c, hsa-miR-141 and hsa-miR-429) and
regulates 19 miRNAs (including hsa-let-7a, hsa-let-7b, hsa-let-7c,
hsa-miR-200a, hsa-miR-200b and hsa-miR-200c). The pathways
concerning ZEB1 are suggested to play a significant role in the
development and pathogenesis of WTs. Detailed information on ZEB1,
NFKB1 and YY1 is introduced in the following text.
Global network of WTs
The global network is too complex and large to be
represented in this study. It contains a large number of
associations between S1, S2 and S3 datasets. Certain genes and
miRNAs in the global network are also involved with other tumors,
such as renal cell carcinoma (16),
which indicates that the global network affects more than one type
of tumor. Therefore, the global network provides a novel
perspective for research regarding the pathogenesis of WT.
Comparison and analysis of features of
differentially-expressed genes in the WT networks
The pathways of the genes and miRNAs has been
described as aforementioned; in this section, the study focuses on
differentially-expressed genes. The total 84
differentially-expressed genes concerned with WT can be classified
into 4 classes.
The first class of genes (E2F3, CTNNB1 and TP53) has
6 types of adjacency nodes. Those adjacency nodes are viewed as 3
successors and 3 predecessors. The following text focuses on
E2F3.
Table I shows the
miRNAs that target E2F3 and are regulated by E2F3. In total, 3
differentially-expressed miRNAs (hsa-miR-17, hsa-miR-20a and
hsa-miR-106b) target E2F3 in the differentially-expressed network,
while 4 and 11 miRNAs target E2F3 in the related and global
networks, respectively. E2F3 regulates 6 differentially-expressed
miRNAs (hsa-let-7a, hsa-let-7a-1, hsa-let-7a-2, hsa-let-7a-3,
hsa-let-7i and hsa-miR-106b) in the differentially-expressed
network. While in the related and global networks, E2F3 regulates 6
and 13 miRNAs, respectively.
 | Table I.Regulatory association between E2F3
and miRNAs. |
Table I.
Regulatory association between E2F3
and miRNAs.
| Gene association |
Differentially-expressed network | Related network | Global network |
|---|
| E2F3 |
|
|
|
| miRNAs
targeting the gene | miR-17, miR-20a,
miR-106b | miR-17, miR-20a,
miR-106b, miR-200b-3p | miR-17, miR-20a,
miR-106b, miR-200b-3p, miR-125b, miR-128, miR-195, miR-203a,
miR-210, miR-34a, miR-34c-5p |
| miRNAs
regulated by the gene | let-7a, let-7a-1,
let-7a-2, let-7a-3, let-7i, miR-106b | let-7a, let-7a-1,
let-7a-2 let-7a-3, let-7i, miR-106b | let-7a, let-7a-1,
let-7a-2, let-7a-3, let-7i, miR-106b, miR-15a, miR-15b, miR-16,
miR-16-2, miR-195, miR-34, miR-34a |
The second class of genes includes EZH2 only. EZH2
has three successors and two predecessors. Differentially-expressed
miRNAs hsa-miR-199a-5p and hsa-miR-217 target EZH2 in the
differentially-expressed network, while 4 and 16 miRNAs target EZH2
in the related and global networks, respectively. No
differentially-expressed miRNAs are regulated by EZH2 in the
differentially-expressed network. Related miRNA hsa-miR-137 is
regulated by EZH2 in the related network and 3 miRNAs are regulated
by EZH2 in the global network.
The third class of genes (TP73, MYCN, SIX1 and EGFR)
has 4 types of adjacency nodes. Despite the fact that they all have
4 adjacency nodes, these genes are not the same. TP73 has no
adjacency node in the differentially-expressed network. TP73
regulates hsa-miR-145 and is targeted by hsa-miR-193a-5p in the
related network. MYCN and SIX1 are targeted by no miRNAs in the
differentially-expressed and related networks. EGFR regulates no
miRNAs in the differentially-expressed and related networks.
The fourth class of genes (including DICER1, BBC3,
RARA, IGF1R and ACVR2B) has 3 types of adjacency nodes. All of
these differentially-expressed genes have no successor, but have 3
predecessors. For example, DICER1 is targeted by hsa-let-7a and
hsa-let-7d in the differentially-expressed network, while 9 and 17
miRNAs target DICER1 in the related and global networks,
respectively. No miRNA is regulated by DICER1 in the three levels
of network.
Comparison and analysis of features of
differentially-expressed miRNAs in the WT networks
The same method was used to compare every
differentially-expressed miRNA, and 87 validated
differentially-expressed miRNAs were extracted from the permanent
literature. These can be classified into four classes.
The first class of miRNAs (hsa-miR-17, hsa-miR-19a,
hsa-miR-20a, hsa-miR-145, hsa-miR-200c and hsa-miR-373) has 6 types
of adjacency nodes. All 6 miRNAs have 3 successors and 3
predecessors. hsa-miR-20a is used here as an example to fully
understand the regulatory mechanisms between genes and miRNAs
(Table II). MYCN regulates
hsa-miR-20a in the differentially-expressed network, 3 and 10 genes
regulate hsa-miR-20a in the related and global networks,
respectively. E2F3 is targeted by hsa-miR-20a in the
differentially-expressed network. Furthermore, 7 and 29 genes are
targeted by hsa-miR-20a in the related and global networks,
respectively.
| Table II.Regulatory relation between
hsa-miR-20a and genes. |
Table II.
Regulatory relation between
hsa-miR-20a and genes.
| Regulatory
relation |
Differentially-expressed network | Related network | Global network |
|---|
| hsa-miR-20a |
|
|
|
| Genes
that regulate miRNA | MYCN | MYCN, CCND1, MYC
E2F1, ESR1, NKX2-5, SPI1, STAT5B, TLX1, TLX3 | MYCN, CCND1,
MYC, |
| Genes
targeted by miRNA | E2F3 | E2F3, CCND1, CCND2
HIF1A, MYC, THBS1 VEGFA | E2F3, CCND1, CCND2,
HIF1A, MYC, THBS1, VEGFA, APP, BCL2, BIM, BMPR2, BNIP2, CDKN1A,
E2F1, IL8, JAK1, MAP3K12, MAPK9, MEF2D, MUC17, NRAS, PTEN, RB1,
RBL1, RBL2, RUNX1, WEE1, SMAD4, TGFBR2 |
The second class of miRNAs (hsa-let-7d, hsa-let-7g
and hsa-miR-106a) has 5 types of adjacency nodes. Despite the same
amount of adjacency nodes, hsa-miR-106a has 2 successors and 3
predecessors, while the other miRNAs (hsa-let-7d and hsa-let-7g)
have 3 successors and 2 predecessors. Hsa-miR106a is regulated by
MYCN in the differentially-expressed network, while 4 and 6 genes
regulate hsa-miR106a in the related and global networks,
respectively. There is no miRNA targeted by hsa-miR106a in the
differentially-expressed network. In the related network, VEGFA is
targeted by hsa-miR106a. In addition, hsa-miR106a targets 22
genes.
The third class of miRNAs (hsa-miR-224, hsa-miR-25,
hsa-miR-24, hsa-miR-192, hsa-miR-194 and hsa-miR-200b) has 4 types
of adjacency nodes. While 3 of them (hsa-miR-224, hsa-miR-25 and
hsa-miR-24) have 3 types of successors and 1 type of predecessor,
the others have 1 type of successor and 3 types of predecessors.
Hsa-miR-224 targets NCOA6 in the differentially-expressed and
related networks, and 18 genes are targeted by hsa-miR-224 in the
global network.
The fourth class of miRNAs (hsa-miR-126,
hsa-miR-154, hsa-miR-197, hsa-miR-22, hsa-miR-27a and hsa-miR-302a)
has 3 types of adjacency nodes. The 3 types of adjacency nodes can
be regard as 2 types of 2 successors and 1 type of predecessor. For
example, none of the genes regulate or are targeted by hsa-miR-126
in the differentially-expressed network; VEGFA is targeted by
hsa-miR-126 in the related network.
Comparison and analysis of features of
predicted TFs in the WT networks
In total, 17 predicted TFs (including NFKB1, ZEB1,
CREB1, HLF, ELK, NR2F2, STAT1, CUX1 and YY1) were extracted by
P-match, and the same method as aforementioned was used to analyze
them. Among the 17 TFs, 13 have no adjacency nodes in the
differentially-expressed and related networks. Only NFKB1, ZEB1,
STAT1 and YY1 were analyzed in this section. These genes can be
classified into 2 types of genes.
The first type of TF (ZEB1 and NFKB1) has 6 types of
adjacency nodes. Table III shows
the regulatory mechanism for ZEB1. The hsa-miR-200a, hsa-miR-200b,
hsa-miR-200c and hsa-miR-141 differentially-expressed miRNAs target
ZEB1. In the related network, ZEB1 is also targeted by these 4
miRNAs. While 6 miRNAs target ZEB1 in the global network, 17 miRNAs
(including hsa-let-7, hsa-let-7a, hsa-let-7a-1, hsa-let-7a-2 and
hsa-let-7a-3) regulate ZEB1 in the differentially-expressed
network. A total of 18 and 20 miRNAs regulate ZEB1 in the related
and global networks, respectively.
| Table III.Regulatory relation between ZEB1 and
miRNAs. |
Table III.
Regulatory relation between ZEB1 and
miRNAs.
| Regulatory
relation |
Differentially-expressed network | Related network | Global network |
|---|
| ZEBI |
|
|
|
| miRNAs
targeting the gene | miR-200a, miR-200b,
miR-200c, miR-141 | miR-200a, miR-200b
miR-200c, miR-141 | miR-200a, miR-200b,
miR-200c, miR-141, miR-205, miR-429 |
| miRNAs
regulated by the gene | let-7, let-7a,
let-7a-1, let-7a-2, let-7a-3, let-7b, let-7c, let-7d, let-7e,
let-7f, let-7f-1, let-7f-2, let-7g, miR-141, miR-200a, miR-200b,
miR-200c | let-7, let-7a,
let-7a-1, let-7a-2, let-7a-3, let-7b, let-7c, let-7d, let-7e,
let-7f, let-7f-1, let-7f-2, let-7g, miR-141, miR-200a, miR-200b,
miR-200c, miR-34b | let-7, let-7a,
let-7a-1, let-7a-2, let-7a-3, let-7b, let-7c, let-7d, let-7e,
let-7f, let-7f-1, let-7f-2, let-7g, miR-141, miR-200a, miR-200b,
miR-200c, miR-34b, miR-34, miR-34a |
The second class of TFs (YY1) has 4 types of
adjacency nodes. Hsa-miR-29b-2 is regulated by YY1 in the related
network; and 8 miRNAs are regulated by YY1 in the global network.
Hsa-miR-31 targets YY1 in the related network, and 2 miRNAs
(hsa-miR-31 and hsa-miR-34a) target YY1 in the global network.
Discussion
In the present study, validated data were collected
from authoritative databases and the permanent literature. These
data were composed of genes and miRNAs. Three levels of network
(the differentially-expressed network, the related network and the
global network) were constructed from genes, miRNAs and the
regulatory mechanisms between them. The intricate association
between genes and miRNAs was the most significant point to analyze
in the study.
After fully researching the networks and comparing
the similarities and differences among the pathways, certain
important factors and pathways were found. There are 5 TFs (E2F3,
MYCN, TP53, SIX1 and CTNNB1) in the differentially-expressed
network. Notably, SIX1 regulates hsa-miR-200a, hsa-miR-200b and
hsa-miR-200c in the differentially-expressed network. However, no
miRNA targets SIX1 in the differentially-expressed and related
networks, which suggests that SIX1 may be the first factor in the
pathogenesis of WT. In addition, E2F3 together with hsa-miR-106b
forms an FFL in the differentially-expressed network; furthermore,
LIN28B and hsa-let-7d also form an FFL in the related network.
These FFLs make a difference in the pathogenesis and progression of
WT as they have been identified to serve a key role in increasing
tumor growth and regulation of WT mechanisms (17). In order to investigate the
pathogenesis and determine better methods for curing patients with
WTs, further comprehensive and authoritative data and pathways will
be used to construct the three level networks, which can be used to
explain the interaction between genes and miRNAs more clearly.
The method proposed by the present study may be
useful in preventing the occurrence of WT. Core genes and miRNAs
not only affect WT, but exhibit an important function in other
tumors. CTNNB1 exhibits a significant role in WT, while it is also
involved in hepatoblastoma (18).
Furthermore, hsa-let-7a is differentially expressed in WT, and is
also overexpressed in retinoblastoma (19). The present study provides a novel
perspective for biologists with regard to WT treatment. The same
differentially-expressed genes and miRNAs exhibit various functions
in different tumors. Therefore, future studies will focus on the
similarities and differences among similar studies on other
cancers.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no. 60905022).
Glossary
Abbreviations
Abbreviations:
|
WT
|
wilms' tumor
|
|
miRNA
|
microRNA
|
|
TF
|
transcription factor
|
|
NCBI
|
National Center for Biotechnology
Information
|
|
TFBS
|
transcription factor binding site
|
|
FFL
|
feed-forward loop
|
References
|
1
|
Beckwith JB, Kiviat NB and Bonadio JF:
Nephrogenic rests, nephroblastomatosis and the pathogenesis of
Wilms' tumor. Pediatr Pathol. 10:1–36. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Watson JA, Bryan K, Williams R, Popov S,
Vujanic G, Coulomb A, Boccon-Gibod L, Graf N, Pritchard-Jones K and
O'Sullivan M: miRNA Profiles as a predictor of chemoresponsiveness
in Wilms' tumor blastema. PLoS One. 8:e534172013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hobert O: Gene regulatory by transcription
factors and microRNAs. Science. 319:W1785–W1786. 2008. View Article : Google Scholar
|
|
4
|
Levine M and Davidson EH: Gene regulatory
networks for development. Sci USA. 102:4936–4942. 2005. View Article : Google Scholar
|
|
5
|
Camas FM, Alm EJ and Poyatos JF: Local
gene regulation details a recognition code within the laci
transcriptional factor family. PLoS Comput Biol. 6:e10009892010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rodriguez A, Griffiths-Jones S, Ashurst JL
and Bradley A: Identification of mammalian microRNA host genes and
transcription units. Genome Res. 14:1902–1910. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang J, Lu M, Qiu C and Cui Q: TransmiR: A
transcription factor-microRNA regulation database. Nucleic Acids
Res. 38:D119–D122. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kozomara A and Griffiths-Jones S: miRBase
integrating microRNA annotation and deep-sequencing data. Nucleic
Acids Res. 39:D152–D157. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chekmenev DS, Haid C and Kel AE: P-Match:
Transcription factor binding site search by combining patterns and
weight matrices. Nucleic Acids Res. 33:W432–W437. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fujita PA, Rhead B, Zweig AS, Hinrichs AS,
Karolchik D, Cline MS, Goldman M, Barber GP, Clawson H, Coelho A,
et al: The UCSC genome browser database: Update. Nucleic Acids Res.
39:D876–D882. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zim B, Hartmann O, Samans B, Krause M,
Wittmann S, Mertens F, Graf N, Eilers M and Gessler M: Expression
profiling of Wilms tumors reveals new candidate genes for different
clinical parameters. Int J Cancer. 118:1954–1962. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cooper CS, Nicholson AG, Foster C, Dodson
A, Edwards S, Fletcher A, Roe T, Clark J, Joshi A, Norman A, et al:
Nuclear overexpression of the E2F3 transcription factor in human
lung cancer. Lung Cancer. 54:155–162. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
An Q, Wang Y, An R, Li Y, Yao T, Zhai B
and Sun X: Association of E2F3 expression with clinicopathological
features of Wilms' tumors. J Pediatr Surg. 48:2187–2193. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sehic D, Karlsson J, Sandstedt B and
Gisselsson D: SIX1 protein expression selectively identifies
blastemal elements in Wilms' tumor. Pediatr Blood Cancer. 59:62–68.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Song CL, Xu ZW, Jin Y, Zhu M, Wang K and
Wang N: The network of microRNAs, transcription factors, target
genes and host genes in human renal cell carcinoma. Oncol Lett.
9:498–506. 2015.PubMed/NCBI
|
|
17
|
Wang L, Che XJ, Wang N, Li J and Zhu MH:
Regulatory network analysis of microRNAs and genes in
neuroblastoma. Asian Pac J Cancer Prev. 10:7645–7652. 2014.
View Article : Google Scholar
|
|
18
|
Koch A, Denkhaus D, Albrecht S, Leuschner
I, von Schweinitz D and Pietsch T: Childhood hepatoblastomas
frequently carry a mutated degradation targeting box of the
beta-catenin gene. Cancer Res. 59:269–273. 1999.PubMed/NCBI
|
|
19
|
Li J, Xu ZW, Wang KH, Wang N, Li DQ and
Wang S: Networks of microRNAs and genes in retinoblastomas. Asian
Pac J Cancer Prev. 14:6631–6636. 2014. View Article : Google Scholar : PubMed/NCBI
|