Introduction
Globally, endometrial carcinoma is the fourth most
common type of malignant tumor affecting women (1,2). In the
past few decades, as longevity has increased and lifestyle has
changed, the incidence of endometrial cancer has significantly
increased and it has become the most common gynecological cancer in
Western countries (3). In 2010,
43,470 individuals were newly diagnosed with endometrial cancer,
whereas there were only 12,200 estimated new cases of cervical
cancer in the United States during the same period (1). Although the prognosis of early-stage
endometrial cancer is favorable when using surgical resection or
adjuvant chemotherapy, no promising treatment is available for
advanced-stage or/and metastatic endometrial cancer. Therefore, it
is crucial that a novel viable treatment strategy is developed in
this field. As in the majority of sarcomas, the phosphoinositide
3-kinase (PI3K)/AKT/mammalian target of rapamycin (mTOR) signaling
transduction pathway plays a critical role in endometrial carcinoma
progression. Numerous studies have suggested that several genetic
mutations in this pathway, including loss of function of the main
negative regulator, phosphatase and tensin homolog (PTEN), directly
contribute to its constant activation, which further leads to tumor
progression. Therefore, the study of the PI3K/AKT/mTOR signaling
transduction pathway may provide novel insights into the drug
development for endometrial cancer. mTOR is one important
downstream target in the PI3K/AKT signaling pathway. Drugs that
target mTOR are able to inhibit cancer cell proliferation, induce
call apoptosis and reverse the drug resistance of cancer cells.
Autophagy is a basic biological process that occurs
in response to physiological or pathological stress, such as
starvation or energy depletion (4).
The process is essential for maintaining intercellular homeostasis
and is thus a key player in tumorigenesis. On one hand, autophagy
promotes tumor cell survival and induces drug resistance (5). On the other hand, certain drug-induced
autophagy can suppress tumor growth and contribute to tumor cell
apoptosis (6,7). Although the exact mechanism involved has
not yet been elucidated, autophagy is considered to exhibit dual
roles in cancer development depending on the cell type and the
stimuli received (6,7). Several studies have been performed
analyzing the role of mTOR in autophagy (8). These studies suggested that mTOR complex
1 (mTORC1) interacted with the unc-51 like autophagy-activating
kinase 1 (ULK1) complex via Raptor (9), and that under nutrient-rich conditions,
mTOR suppressed autophagy through the direct phosphorylation of
ULK1 and mAtg13. Additionally, under glucose-rich conditions, mTOR
could mediate the dissociation of adenosine monophosphate-activated
protein kinase and ULK1 via the direct phosphorylation of ULK1,
thereby inhibiting autophagy (10).
Upon starvation or rapamycin stimulation, mTOR dissociated from the
ULK1 complex provoking the dephosphorylation of ULK1 and mAtg13,
which further initiated autophagy (11). Therefore, mTOR, as the main regulator
in autophagy, is also an attractive target for the future
development of cancer treatment.
RAD-001 (also known as everolimus), a derivative of
rapamycin, is a newly developed oral mTOR inhibitor that is
currently undergoing clinical trials as an antitumor drug (12,13).
Similar to the rapamycin, RAD001 suppresses tumor proliferation by
inhibiting mTOR function and blocking the mTOR signaling pathway
(14). The half maximal inhibitory
concentration (IC50) for RAD001 ranges between 5 and
1,800 µmol/l among different cell types, including melanoma, lung
cancer, breast cancer, lymphoma, pancreatic and colon cancer cells
(15). Preclinical studies indicated
that RAD001 alone could suppress tumor growth, and that it also
displayed a synergistic effect in combination with either hormones
or other cytotoxic agents (16). It
was reported that RAD001 could induce autophagy in testicular
cancer cells with PTEN mutation and that it sensitized the cells to
radiation therapy (17,18). For certain drug-resistant cancer
cells, RAD001 in combination with certain anticancer drugs could
effectively inhibit cell proliferation and promote cell apoptosis.
Clinical study results suggested that RAD001 showed a synergistic
effect in combination with other anticancer agents, and no marked
toxic effect was observed (19,20). A
recent phase IB clinical study conducted on patients with
advanced-stage carcinoma reveled that the combination of RAD001 and
paclitaxel exhibited a less toxic effect than either drug alone
(21).
The present study therefore aimed to investigate the
effect of the mTOR inhibitor, RAD001, on autophagy in endometrial
cancer cells and the mechanism involved.
Materials and methods
Materials
The human endometrial cancer Ishikawa and HEC-1A
cell lines were obtained from the Shanghai Institute of Cellular
Biology of the Chinese Academy of Sciences (Shanghai, China) and
cultivated in Dulbecco' modified Eagle's medium (DMEM) (Invitrogen;
Thermo Fisher Scientific Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (Gibco; Thermo Fisher Scientific Inc.), 50
µg/ml penicillin (Invitrogen; Thermo Fisher Scientific Inc.), 50
µg/ml streptomycin (Invitrogen; Thermo Fisher Scientific Inc.) and
2 mmol/l glutamine (Gibco; Thermo Fisher Scientific Inc.) at 37°C
in 5% CO2. All experiments were performed with cells in
the logarithmic phase.
RAD001 (catalogue no. 07741), MTT and chloroquine
(CQ) were purchased from Sigma-Aldrich (Merck Millipore, Darmstadt,
Germany). RAD001 was formulated in 1 mmol/l dimethyl sulfoxide
(DMSO) diluent. Polyclonal rabbit anti-phospho-AKT (Ser473)
antibody (#9271; 1:1,000), monoclonal rabbit anti-mTOR antibody
(#2983; 1:1,000), monoclonal rabbit anti-phospho-mTOR (Ser2448)
antibody (#5536; 1:1,000), monoclonal rabbit anti-p70S6K antibody
(#2708; 1:1,000) and polyclonal rabbit anti-phospho-p70S6K
(Thr421/Ser424) antibody (#9204; 1:1,000) were acquired from Cell
Signaling Technology Inc. (Danvers, MA, USA). Polyclonal rabbit
anti-LC3 antibody (#NB100-2220; 1:1,000) was obtained from Novus
Biological LLC (Littleton, CO, USA). Goat anti-rabbit
immunoglobulin G horseradish peroxidase-conjugated secondary
antibody (#sc-2004; 1:2,000) was purchased from Santa Cruz
Biotechnology Inc. (Dallas, TX, USA) and monoclonal mouse
anti-GAPDH antibody (KC-5G4; 1:5,000) was purchased from Kangcheng
Bio-tech Inc. (Shanghai, China).
MTT assay
Cells in the logarithmic phase were seeded in
96-well plates at a density of 8,000 cells/well in 195 µl DMEM and
cultivated at 37°C overnight. The cells were then treated with CQ
(10 mg/ml in DMSO), paclitaxel (1, 2, 4, 8 and 16 µM or the
indicated concentrations in DMSO), 3-methyladenine (3-MA) (10 µM in
H2O, which was used as an autophagy inhibitor) and DAPI
(1 µg/ml in H2O) for the indicated time points at 37°C
in 5% CO2. Solvent was used as the control in each
experiment. Experiments were terminated by adding 10 µl of 5 mg/ml
MTT and incubated at 37°C for 4 h. Following complete removal of
the medium, 100 µl of DMSO (Sigma-Aldrich; Merck Millipore) was
added to each well to dissolve the purple formazan product.
Absorbance values of the resultant purple solution were obtained
with a test wavelength of 570 nm. The IC50 values were
calculated by the Bliss method: Inhibitory rate (%)= [1 - the
average optical density (OD) value of the treatment group / the
average OD value of the control group] × 100. Growth inhibition was
calculated according to the results of the MTT assay, and the
combination index was determined using CalcuSyn software version
2.0 (Biosoft, Cambridge, UK).
Fluorescence microscopy
For the microscopic examination, 1 day prior to
transfection, the cells were plated in 6-well plates with
antibiotic-free RPMI 1640 growth medium at a density of
1.5×105 cells/well. When the cells grew to a confluence
of ~50% on the second day, green fluorescent protein
microtubule-associated protein 1 light chain 3α
(GFP-LC3)-expressing plasmids were transfected into the cells using
Lipofectamine 2000 (Thermo Fisher Scientific Inc.), and cells
stably transfected with the GFP-LC3 plasmid (kindly provided by
Professor Beth Levine), which stably expressed GFP-LC3, were
selected with growth medium containing G418 antibiotic. Subsequent
to treatment with the experimental drugs, the cells were washed
with 1X PBS three times and then evaluated under a confocal
fluorescence microscope (LSM 710 Meta; Carl Zeiss AG, Oberkochen,
Germany).
Immunoblotting analysis
Prior to the drug treatment, the cells were plated
in 6-well plates at a density of 4×105 cells/well and
incubated at 37°C in 5% CO2 overnight. Following
treatment, the cells were collected and washed with 1X PBS three
times and lysed in 100 µl/well of lysis buffer. Cell lysates were
centrifuged at 13,600 × g for 10 min at 4°C and the protein
concentrations were determined by Bio-Rad protein assay (Bio-Rad
Laboratories Inc., Hercules, CA, USA). SDS-PAG loading buffer was
added to the cell lysate, which was then heated at 95°C for 10 min.
Each sample containing 40 µg of protein was then loaded into each
well of the SDS-PAGE gels and the resolved proteins were
transferred to a polyvinylidene difluoride membrane
electrophoretically. Subsequent to blocking with 5% skimmed milk,
the membranes were probed sequentially with primary and secondary
antibodies overnight at 4°C. Following washing three times with TBS
plus Tween 20 [10 mmol/l Tris-HCl (pH 7.4), 150 mmol/l NaCl and
0.1% Tween 20] (TBST), the proteins were detected using enhanced
chemiluminescence reagent (GE Healthcare Life Sciences, Chalfont,
UK) and XAR film (Kodak, New York, NY, USA).
Flow cytometry
Pre-treated cells were collected and washed twice
with 1X PBS. The cells were then re-suspended in 1X PBS at a
density of 1×106 cells/ml. Next, 10 µl of 10 mg/l
propidium iodide was added to 1-ml cell suspension, which was then
incubated in the dark for 10 min. The samples were placed on ice
prior to being analyzed by flow cytometer (BD Biosciences, Franklin
Lakes, NJ, USA).
RNA interference
Protein depletion through RNA-mediated interference
was mediated using the pSUPER small hairpin (sh)RNA system.
Retroviruses were generated by co-transfection of pSUPER-shRNA
plasmids (#30519; Addgene, Inc., Cambridge, MA, USA) with
retrovirus plasmid PIK (Ecopac: M. Finer Cell Genosys, Redwood
City, CA, USA) into 293T cells by liposome. Retroviruses were
collected in high-serum media at 48 and 72 h post-transfection.
Ishikawa and HEC-1A cells were transduced with retroviruses and 8
µg/ml Polybrene (hexadimethrine bromide; Sigma-Aldrich; Merck
Millipore) followed by incubation with virus at 37°C for 4–6 h.
shRNA-transduced cells were selected for with 1 µg/ml puromycin for
72 h. To confirm the efficiency of Atg5 shRNA, puromycin-selected
cells transfected with a specific shRNA targeting human Atg5
(5′-GCAACUCUGGAUGGGAUUG-3′) were cultured three-dimensionally in
vitro. Cells were then subjected to western blot detection with
anti-Atg5 polyclonal rabbit antibody (1:1,000; #2630; Cell
Signaling Technology Inc.) and anti-GAPDH antibody (1:5,000;
KC-5G4; Zhejiang Kangchen Biotech Co., Ltd., Shanghai, China) for
12 h at 4°C. Following three washes with TBST, the proteins were
detected using an enhanced chemiluminescence reagent and BioMax XAR
Film (Kodak, Rochester, NY, USA).
Statistical analysis
Data were presented as the mean ± standard
deviation, and analyzed with a one-way analysis of variance and
Student-Newman-Keuls-q test (22) by
SPSS 16.0 statistical software (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
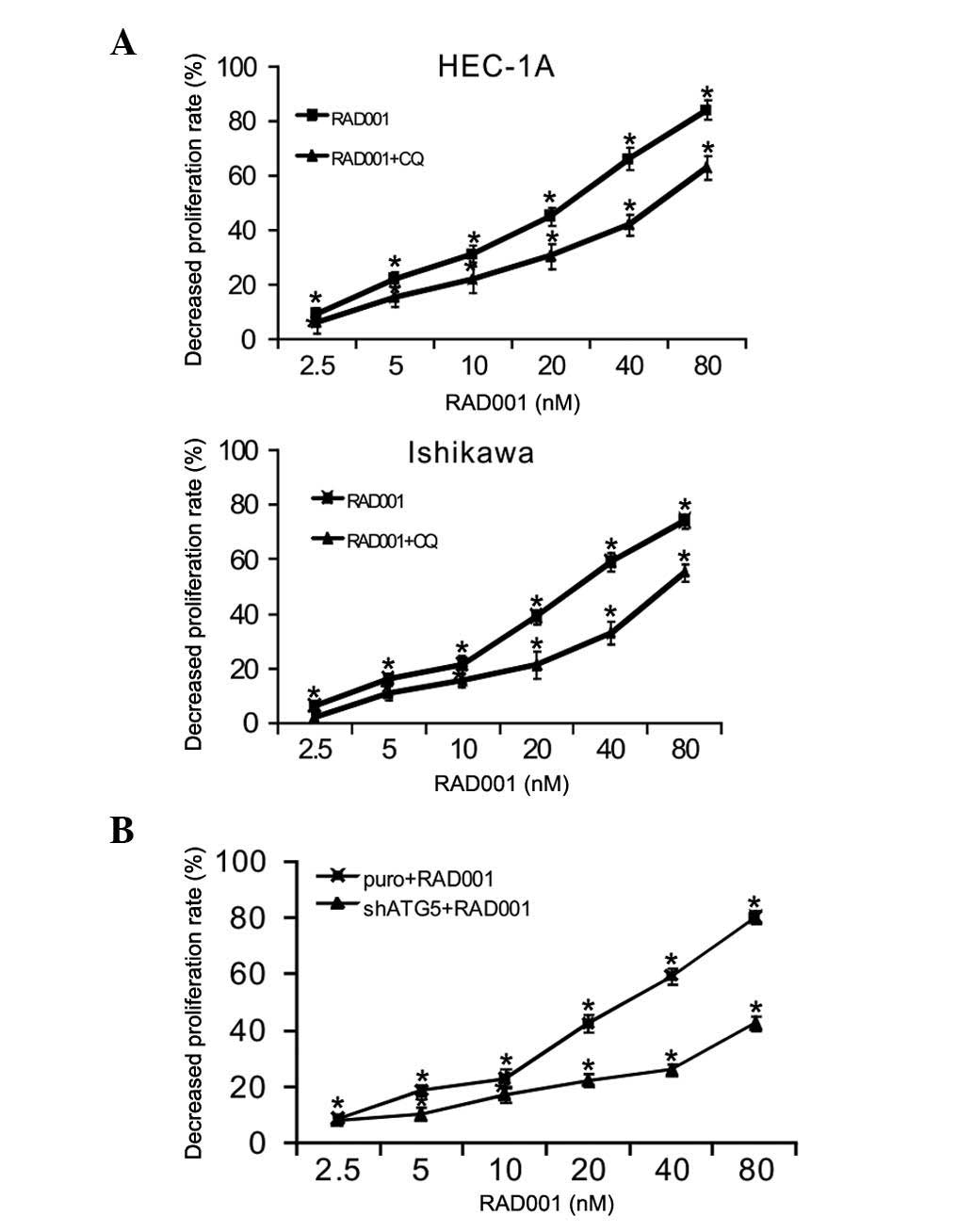
RAD001 inhibits human endometrial
cancer Ishikawa and HEC-1A cell proliferation
The inhibitory effect of RAD001 on Ishikawa and
HEC-1A cells was demonstrated using MTT assay. The soluble yellow
compound of MTT was reduced to insoluble formazan, which produced a
purple color in living cells. As the amount of formazan produced
was proportional to the number of viable cells, after dissolving it
in DMSO, the absorbance values of the resultant purple solution
were used to calculate the inhibitory rate of cell proliferation
and thus evaluate the cytotoxicity of RAD001. Following treatment
with different concentrations (0, 5, 10, 20, 40 and 80 nM) of
RAD001 for 72 h, the proliferation of the Ishikawa and HEC-1A cells
was suppressed in a dose-dependent manner, and all results were
significant compared with the control group (P<0.01) (Fig. 1). The group treated with 0 nM PAD001
was considered as the control group, and the decreased
proliferation rate of this group was normalized to 0% (which
coincides with the origin of coordinates in Fig. 1). The IC50 values were
36.80±1.64 and 25.72±1.16 nM for the Ishikawa and HEC-1A cells,
respectively. The results suggested that RAD001 alone could
effectively inhibit the proliferation of the Ishikawa and HEC-1A
cells.
RAD001 sensitizes endometrial cancer
Ishikawa and HEC-1A cells to paclitaxel treatment
Following treatment with RAD001 in combination with
different concentrations of paclitaxel, the proliferation of the
Ishikawa and HEC-1A cells was significantly inhibited in a
dose-dependent manner. The IC50 values for the Ishikawa
and HEC-1A cells treated with paclitaxel alone were 7.91 and 9.27
µM, respectively. The corresponding combination index was <1 for
the two cell lines, which was statistically significant (Fig. 2A). Apoptosis was observed in the
Ishikawa cells treated with paclitaxel, as indicated by the
presence of cleaved caspase3 and cleaved poly ADP ribose
polymerase; when RAD001 was also added, the cleaved band became
markedly more intense (Fig. 2B).
According to flow cytometry, the apoptotic cell count increased
from 15.2 to 45% (Fig. 2C). These
results indicated that the combination treatment of RAD001 and
paclitaxel is synergistic for suppressing the human endometrial
cancer cell proliferation.
RAD001 induces autophagy in
endometrial cancer Ishikawa and HEC-1A cells
LC3 is an autophagic marker protein whose lipidated
form, LC3II, is recruited to the autophagosomal membranes at a late
stage of autophagy (23). As shown in
Fig. 3A, upon treatment with RAD001,
green puncta were detected in the cytosol of GFP-LC3-expressing
Ishikawa and HEC-1A cells, denoting the formation of
autophagosomes. Western blot analysis results further confirmed
this, as the expression level of LC3II protein increased in a
dose-dependent manner for the two cell lines following treatment
with different concentrations of RAD001 for 24 h (Fig. 3B). This suggested that RAD001 induced
autophagy in the Ishikawa and HEC-1A cells.
RAD001 induces autophagic cell death
in endometrial cancer Ishikawa and HEC-1A cells
MTT results confirmed that the inhibitory effect of
RAD001 on Ishikawa and HEC-1A cell proliferation was decreased when
autophagy was suppressed by CQ, suggesting that RAD001 induced
autophagic cell death in the Ishikawa and HEC-1A cells (Fig. 4A). Furthermore, inhibition of
autophagy by shRNA knockdown of Atg5 also resulted in the reduced
inhibition of cell death as induced by RAD001 (Fig. 4B). The levels of Atg5 were effectively
reduced by Atg5 shRNA (data not shown). Together, these results
showed that RAD001 induced autophagic cell death in the endometrial
cancer Ishikawa and HEC-1A cells.
RAD001 suppresses the AKT/mTOR/p70S6K
signaling transduction pathway in endometrial cancer Ishikawa and
HEC-1A cells
To further investigate whether the AKT/mTOR/p70S6K
signaling pathway plays a role in RAD001-induced autophagy, the
expression levels of the key proteins in this signaling pathway
were examined in Ishikawa and HEC-1A cells by western blot
analysis. The results indicated that RAD001 inhibited serine 473
phosphorylation and mTOR phosphorylation in a dose-dependently
manner in the two cell lines, with no significant change in the
expression level of mTOR (Fig. 5).
Furthermore, it was observed that the phosphorylation of one of the
essential substrates of mTORC1, p70S6 kinase, was also
significantly suppressed. These results suggested that the
AKT/mTOR/p70S6K signaling transduction pathway was suppressed upon
treatment with RAD001 in the Ishikawa and HEC-1A cells.
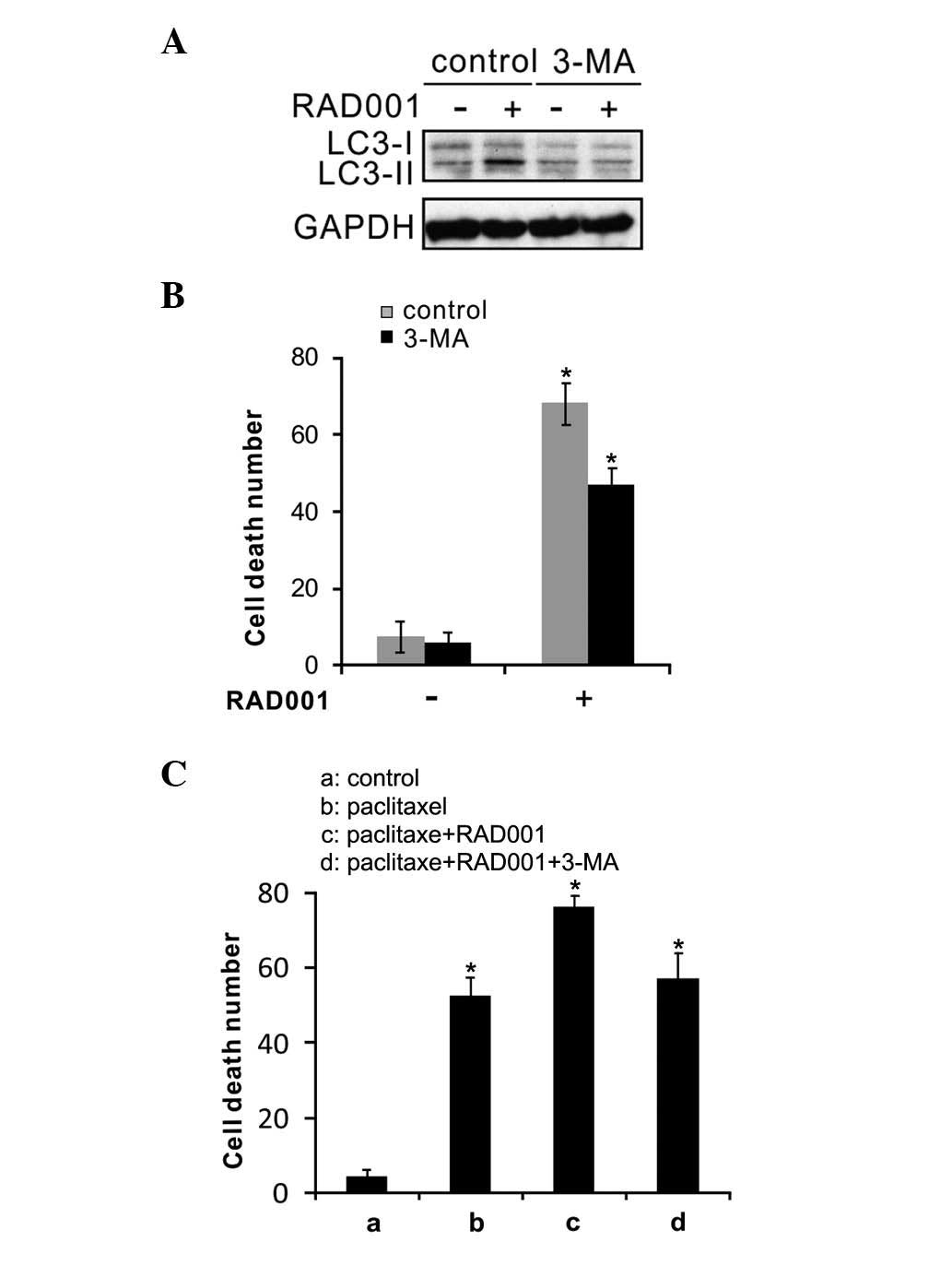
RAD001 sensitizes Ishikawa cells to
paclitaxel via the induction of autophagy
To establish whether RAD001 sensitized endometrial
cancer Ishikawa and HEC-1A cells to paclitaxel treatment via the
induction of autophagy, Ishikawa cells were treated with 3-MA, the
autophagy inhibitor, together with RAD001. Consistent with
aforementioned results, RAD001 triggered autophagy, as shown by the
presence of LC3II, and 3-MA inhibited this change (Fig. 6A). As shown in Fig. 6B, the effect of autophagy inhibition
with 3-MA plus RAD001 was tested on cell death in the Ishikawa
cells; it was found that RAD001-induced autophagic cell death was
efficiently suppressed when autophagy was blocked. Also, the result
from the flow cytometric apoptosis assay indicated that Ishikawa
cell apoptosis was significantly enhanced when the cells were
treated with RAD001 in combination with paclitaxel, and this effect
was repressed upon inhibition of autophagy (Fig. 6C). These results suggested that RAD001
sensitized the Ishikawa cells to paclitaxel via the induction of
autophagy.
Discussion
Endometrial cancer is the most common gynecological
cancer, accounting for 20–30% of cases. In certain countries, it is
even the most common gynecological malignancy (1).
Abnormal signal transduction is one of the
contributing factors for cancer development (24,25). As a
deeper understanding has been gained in the field of signal
transduction over the past decade, targeted cancer therapy has
become an innovative approach for future cancer treatment (24–26). mTOR
is a key downstream protein kinase of the PI3K/Akt signaling
pathway, and drugs that target mTOR can effectively inhibit cancer
cell proliferation, induce cancer cell apoptosis and reverse the
drug-resistant effect in cancer chemotherapy (27,28). Since
mTOR has become a novel target for research in cancer therapy, its
inhibitor, RAD001, has received much attention as a potential
targeted cancer drug (12,13).
Paclitaxel is a common anticancer drug that is used
in combination chemotherapy (29–32).
Although a high dose of paclitaxel has proven to be more effective,
its clinical toxicity is severe (33,34). The
present study investigated the effect of the combination of RAD001
and paclitaxel on Ishikawa and HEC-1A cells. The results showed
that RAD001 sensitized the cells to paclitaxel, suggesting that
using a combination treatment involving RAD001 could reduce the
cytotoxic effect of paclitaxel, as it was effective at a lower
dose. More importantly, RAD001 only targets malignant cancer cells,
with few side effects on normal cells, which makes it an ideal
targeted drug in the synergistic therapy for future clinical cancer
treatment (35).
Autophagy is a highly-conserved metabolic process
that removes damaged organelles or proteins, and recycles
cytoplasmic contents in response to cytotoxic and metabolic
stresses (36). Although autophagy is
essential for maintaining intracellular homeostasis and promoting
cell survival, it can also be involved in the cell death process
(37). Therefore, the precise role of
autophagy in cancer should be the first consideration in the study
of autophagy-inducing antitumor agents. To further illustrate the
role of RAD001-induced autophagy in endometrial cancer cells, the
present study pre-treated Ishikawa and HEC-1A cells with the
autophagy inhibitor CQ. The results showed that cell survival was
significantly increased, suggesting that RAD001 induced autophagic
cell death in endometrial cancer cells.
Previous studies confirmed that, under nutrient
deprivation or rapamycin stimulation, the function of mTOR kinase
was suppressed. Also, Atg13 was dephosphorylated, which allowed it
to have higher affinity to Atg1 and thus bind to Atg17-29-31 to
form the Atg1 complex. Atg1 was further activated by Atg13/Atg17
and re-localized to the phagosome assembly site for the initiation
of autophagy (38–40). Hence, mTOR kinase is the negative
regulator in autophagy. The present results demonstrated that
RAD001 significantly inhibited the AKT/mTOR phosphorylation in
Ishikawa and HEC-1A cells, which further suppressed its downstream
substrate, p70S6 kinase.
In conclusion, the present study demonstrated that
RAD001 sensitizes endometrial cancer Ishikawa and HEC-1A cells to
paclitaxel by inducing autophagic cell death via suppression of the
mTOR signaling pathway. The study provides experimental evidence
for the future clinical study of combination chemotherapy and
introduces the possibility of using RAD001 as a potential targeted
drug in the synergistic therapy for future clinical cancer
treatments.
Acknowledgements
The authors would like to thank Professor Beth
Levine (Center for Autophagy Research, University of Texas
Southwestern Medical Center, Dallas, TX, USA) for kindly providing
the GFP-LC3 plasmid (41). The
present study was supported by grants from the National Nature
Science Foundation of China (grant nos. 81201772 and 31200806), the
Fundamental Research Funds for the Central Universities (Sun
Yat-Sen University Young Teachers Plan; grant nos. 12ykpy50 and
12ykpy05), the Pearl River Nova Program of Guangzhou (grant no.
2014J2200039) and the PhD Start-up Fund of Natural Science
Foundation of Guangdong Province (grant no. S2012040007502).
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sankaranarayanan R and Ferlay J: Worldwide
burden of gynaecological cancer: The size of the problem. Best
Pract Res Clin Obstet Gynaecol. 20:207–225. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boya P, González-Polo RA, Casares N,
Perfettini JL, Dessen P, Larochette N, Métivier D, Meley D,
Souquere S, Yoshimori T, et al: Inhibition of macroautophagy
triggers apoptosis. Mol Cell Biol. 25:1025–1040. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qi XF, Kim DH, Lee KJ, Kim CS, Song SB,
Cai DQ and Kim SK: Autophagy contributes to apoptosis in A20 and
EL4 lymphoma cells treated with fluvastatin. Cancer Cell Int.
13:1112013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kondo Y, Kanzawa T, Sawaya R and Kondo S:
The role of autophagy in cancer development and response to
therapy. Nat Rev Cancer. 5:726–734. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mathew R, Karantza-Wadsworth V and White
E: Role of autophagy in cancer. Nat Rev Cancer. 7:961–967. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jung CH, Ro SH, Cao J, Otto NM and Kim DH:
mTOR regulation of autophagy. FEBS Lett. 584:1287–1295. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ganley IG, Lam du H, Wang J, Ding X, Chen
S and Jiang X: ULK1 ATG13. FIP200 complex mediates mTOR signaling
and is essential for autophagy. J Biol Chem. 284:12297–12305. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guertin DA and Sabatini DM: Defining the
role of mTOR in cancer. Cancer Cell. 12:9–22. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hippert MM, O'Toole PS and Thorburn A:
Autophagy in cancer: Good, bad, or both? Cancer Res. 66:9349–9351.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lévy A, Sauvin LA, Massard C and Soria JC:
Everolimus (RAD001) and solid tumours: A 2008 summary. Bull Cancer.
95:1205–1211. 2008.(In French). PubMed/NCBI
|
|
13
|
Agarwala SS and Case S: Everolimus
(RAD001) in the treatment of advanced renal cell carcinoma: A
review. Oncologist. 15:236–245. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Beuvink I, Boulay A, Fumagalli S,
Zilbermann F, Ruetz S, O'Reilly T, Natt F, Hall J, Lane HA and
Thomas G: The mTOR inhibitor RAD001 sensitizes tumor cells to
DNA-damaged induced apoptosis through inhibition of p21
translation. Cell. 120:747–759. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Weigelt B and Downward J: Genomic
determinants of PI3K pathway inhibitor response in cancer. Front
Oncol. 2:1092012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dengler J, von Bubnoff N, Decker T,
Peschel C and Duyster J: Combination of imatinib with rapamycin or
RAD001 acts synergistically only in Bcr-Abl-positive cells with
moderate resistance to imatinib. Leukemia. 19:1835–1838. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cao C, Subhawong T, Albert JM, Kim KW,
Geng L, Sekhar KR, Gi YJ and Lu B: Inhibition of mammalian target
of rapamycin or apoptotic pathway induces autophagy and
radiosensitizes PTEN null prostate cancer cells. Cancer Res.
66:10040–10047. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shinohara ET, Cao C, Niermann K, Mu Y,
Zeng F, Hallahan DE and Lu B: Enhanced radiation damage of tumor
vasculature by mTOR inhibitors. Oncogene. 24:5414–5422. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Haritunians T, Mori A, O'Kelly J, Luong
QT, Giles FJ and Koeffler HP: Antiproliferative activity of RAD001
(everolimus) as a single agent and combined with other agents in
mantle cell lymphoma. Leukemia. 21:333–339. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Panner A, Parsa AT and Pieper RO: Use of
APO2L/TRAIL with mTOR inhibitors in the treatment of glioblastoma
multiforme. Expert Rev Anticancer Ther. 6:1313–1322. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mayer IA, Burris H, Bendel J, Means-Powell
J, Arteaga C, Shyr Y and Pietenpol J: A phase Ib trial of RAD001,
an mTOR inhibitor, with weekly cisplatin and paclitaxel in patients
with HER2-negative metastatic breast cancer. Abstract 3093. 32nd
Annual San Antonio Breast Cancer Symposium. San Antonio, TX.
December 9–13, 2009;
|
|
22
|
Mason RL, Gunst RF and Hess JJ:
Statistical Design and Analysis of ExperimentsWith Applications to
Engineering and Science. 2nd. John Wiley and Sons Inc.; Hoboken,
NJ: 2003
|
|
23
|
Tanida I, Ueno T and Kominami E: LC3 and
autophagy. Methods Mol Biol. 445:77–88. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sever R and Brugge JS: Signal transduction
in cancer. Cold Spring Harb Perspect Med. 5:a0060982015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kolch W, Halasz M, Granovskaya M and
Kholodenko BN: The dynamic control of signal transduction networks
in cancer cells. Nat Rev Cancer. 15:515–527. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ranjan A, Fofaria NM, Kim SH and
Srivastava SK: Modulation of signal transduction pathways by
natural compounds in cancer. Chin J Nat Med. 13:730–742.
2015.PubMed/NCBI
|
|
27
|
Zhou HY and Huang SL: Current development
of the second generation of mTOR inhibitors as anticancer agents.
Chin J Cancer. 31:8–18. 2012.PubMed/NCBI
|
|
28
|
Tan HK, Moad AI and Tan ML: The mTOR
signalling pathway in cancer and the potential mTOR inhibitory
activities of natural phytochemicals. Asian Pac J Cancer Prev.
15:6463–6475. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ozols RF: Paclitaxel (Taxol)/carboplatin
combination chemotherapy in the treatment of advanced ovarian
cancer. Semin Oncol 27 (Suppl 7). 3–7. 2000.
|
|
30
|
Okamoto I, Moriyama E, Fujii S, Kishi H,
Nomura M, Goto E, Kiyofuji C, Imamura F, Mori T and Matsumoto M:
Phase II study of carboplatin-paclitaxel combination chemotherapy
in elderly patients with advanced non-small cell lung cancer. Jpn J
Clin Oncol. 35:188–194. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pentheroudakis G, Razis E, Athanassiadis
A, Pavlidis N and Fountzilas G: Paclitaxel-carboplatin combination
chemotherapy in advanced breast cancer: Accumulating evidence for
synergy, efficacy, and safety. Med Oncol. 23:147–160. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yardley DA, Brufsky A, Coleman RE, Conte
PF, Cortes J, Glück S, Nabholtz JM, O'Shaughnessy J, Beck RM, Ko A,
et al: Phase II/III weekly nab-paclitaxel plus gemcitabine or
carboplatin versus gemcitabine/carboplatin as first-line treatment
of patients with metastatic triple-negative breast cancer (the
tnAcity study): Study protocol for a randomized controlled trial.
Trials. 16:5752015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yamazaki S, Sekine I and Saijo N:
Paclitaxel (taxol): A review of its antitumor activity and toxicity
in clinical studies. Gan To Kagaku Ryoho. 25:605–615. 1998.(In
Japanese). PubMed/NCBI
|
|
34
|
Liu H, Chen X, Sun J, Gao P, Song Y, Zhang
N, Lu X, Xu H and Wang Z: The efficacy and toxicity of paclitaxel
plus S-1 compared with paclitaxel plus 5-FU for advanced gastric
cancer: A PRISMA systematic review and meta-analysis of randomized
controlled trials. Medicine (Baltimore). 93:e1642014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dufour M, Dormond-Meuwly A, Demartines N
and Dormond O: Targeting the Mammalian Target of Rapamycin (mTOR)
in Cancer Therapy: Lessons from Past and Future Perspectives.
Cancers (Basel). 3:2478–2500. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mizushima N: Autophagy: Process and
function. Genes Dev. 21:2861–2873. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Berry DL and Baehrecke EH: Autophagy
functions in programmed cell death. Autophagy. 4:359–360. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Alonso S, Pethe K, Russell DG and Purdy
GE: Lysosomal killing of Mycobacterium mediated by
ubiquitin-derived peptides is enhanced by autophagy. Proc Natl Acad
Sci USA. 104:6031–6036. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lum JJ, Bauer DE, Kong M, Harris MH, Li C,
Lindsten T and Thompson CB: Growth factor regulation of autophagy
and cell survival in the absence of apoptosis. Cell. 120:237–248.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Park HJ, Lee SJ, Kim SH, Han J, Bae J, Kim
SJ, Park CG and Chun T: IL-10 inhibits the starvation induced
autophagy in macrophages via class I phosphatidylinositol 3-kinase
(PI3K) pathway. Mol Immunol. 48:720–727. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pattingre S, Tassa A, Qu X, Garuti R,
Liang XH, Mizushima N, Packer M, Schneider MD and Levine B: Bcl-2
antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell.
122:927–939. 2005. View Article : Google Scholar : PubMed/NCBI
|