Introduction
Ribosomal S6 kinases (RSKs) are a family of
serine/threonine protein kinases that are directly regulated by
extracellular signal-regulated kinase (ERK) signaling. RSK1, a
member of the RSK family, was initially identified as an X-linked
gene in patients with mental retardation (1–3).
Typically, RSK is expressed in the cerebellum during embryogenesis
and silenced postnatally. Aberrant RSK signaling is integral for
various types of cancer, including breast, colon and renal cancer,
as well as melanoma. Li et al (4) observed that the p90 RSK2-cAMP response
element-binding protein (CREB) pathway is commonly activated in
diverse metastatic human cancer cells. Degen et al (5) also demonstrated this phenomenon, and
further observed that overexpression of RSK3 and RSK1 supports
cellular proliferation under the PI3K signaling pathway blockade.
This occurs through the inhibition of apoptosis and regulation of
cellular translation in squamous carcinoma cell through
phosphorylation of RSK and eukaryotic translation initiation factor
4B. It is thus reported that RSK1 serves a role in squamous
carcinoma cell growth and proliferation. In addition, Cohen et
al (6) observed that RSK1
overexpression is associated with sunitinib resistance in renal
cell carcinoma cell lines. Elf et al (7) also reported that, although RSK1 is
dispensable for BCR-ABL-induced myeloid leukemia, it may be
required for the pathogenesis and lineage determination in
FLT3-ITD-induced hematopoietic transformation. However, the
expression of RSK1 in leukemia remains uncertain.
In the past decades, numerous studies have attempted
to identify natural compounds with potential anti-leukemic activity
(8–10). Luteolin, also known as
3′,4′,5′,7′-tetrahydroxyflavone, is a flavonoid compound that has a
C6-C3-C6 structure and possesses two benzene rings, an
oxygen-containing ring and a 2–3 carbon double bond. This compound
has been found to possess various beneficial properties, including
antioxidant (11), anti-inflammatory
(12), anti-bacterial (13) and anti-cancer activities (14–17).
Recently, Reipas et al (18)
reported that luteolin is a novel p90 RSK inhibitor that can be
considered as a promising candidate agent for the treatment of
liver, lung, breast, colon, prostate, gastric and ovarian cancer,
as well as melanoma. In the present study, the effect of Luteolin
on AML was investigated.
Materials and methods
Materials
Luteolin was purchased from Sigma-Aldrich (Oakville,
ON, USA) and dissolved in dimethyl sulfoxide to stock concentration
of 100 mM at −20°C. The Cell Counting kit-8 (CCK-8) and Hoechst
33258 were purchased from Beyotime Institute of Biotechnology
(Beijing, China). Lipofectamine RNAiMAX, TRIzol and RT reagents
were purchased from Thermo Fisher Scientific, Inc. (Waltham, MA,
USA). SYBR Green Supermix was purchased from Takara Bio, Inc.
(Otsu, Japan). An Annexin V-FITC/propidium iodide (PI) double
staining kit was purchased from Nanjing KeyGen Biotech Co., Ltd.
(Nanjing, China). Antibodies were purchased against the
phosphorylated forms of RSK1 (Ser221; ab10695; Abcam, Cambridge,
MA, USA), B-cell lymphoma (Bcl)-2-associated death promoter (BAD)
(Ser112; 9291; Cell Signaling Technology, Inc., Danvers, MA, USA),
kidney/brain protein (KIBRA) (Ser947; ab107637; Abcam) and GAPDH
(ab8245; Abcam), and were diluted at 1:1,000.
Patients and specimens
A total of 30 patients with primary newly-diagnosed
acute myeloid leukemia (AML) were enrolled into the present study.
The patients underwent consecutive chemotherapy at the Department
of Hematology of the Sun Yat-Sen Memorial Hospital (Sun Yat-Sen
University, Guangzhou, China) between July 2011 and July 2014. The
samples were obtained by bone marrow aspiration prior to initiation
of the therapy and after finishing 6 cycles of daunorubicin and
cytarabine (DA) therapy, which was defined as the first complete
remission (CR) stage (CR1). Each chemotherapy cycle included
standard dose cytarabine (100–200 mg/m2) as a continuous
infusion for 7 days with daunorubicin hydrochloride (40–60
mg/m2) ×3 once every 28 days. Informed consent was
obtained from all patients following a protocol approved by the
Ethics Committee of the Sun Yat-Sen Memorial Hospital, Sun Yat-Sen
University. AML was evaluated by the World Health Organization
classification (19). CR was defined
as a bone marrow sample with <5% blast cells and a neutrophil
count of >10,000 cells.
RT-qPCR
RSK1 level was detected in MOLM-13, Kasumi-1 and
primary AML cells. The primary AML cells were isolated from from 2
ml bone marrow using lymphocyte separation liquid by a density
gradient centrifugation method (20).
For each clinical sample, mononuclear cells were isolated from 2 ml
bone marrow using lymphocyte separation liquid by a density
gradient centrifugation method. In order to compare the expression
of RSK1 in leukemic cell line, mononuclear cells were isolated from
2 ml peripheral blood of 3 cases using the seam method, and these
samples were used as the control group. Total RNA was extracted
from the samples using TRIzol reagent, according to the
manufacturer's instructions. The cDNA was synthesized using the
M-MLV reverse transcriptase kit (Invitrogen; Thermo Fisher
Scientific, Inc.). Quantitative analysis of RSK1 mRNA expression
was evaluated by qPCR using SYBR® Green One-Step qRT-PCR
kit (11736059; Invitrogen; Thermo Fisher Scientific, Inc.), and
β-actin was used as an endogenous control. The sequences of the
qPCR primers were as follows: RSK1 forward,
5′-GGTGGTCCTATGGGGTGTTG-3′, and reverse,
5′-TCGCCTTCAGAATCAGTGTCA-3′; and β-actin forward,
5′-TGAAGTGTGACGTGGACATC-3′, and reverse,
5′-GGAGGAGCAATGATCTTGAT-3′. All the reactions were performed in a
20 µl reaction volume in triplicate. The thermal cycling conditions
were as follows: A denaturation step of 31 cycles at 90°C for 20
sec, an annealing step at 52°C for 25 sec and synthesis at 72°C for
20 sec. The fold changes were calculated through relative
quantification with the 2−∆∆Cq method.
Cell culture
MOLM-13 and Kasumi-1 cells (Type Culture Collection
of the Chinese Academy of Sciences, Shanghai, China) were
maintained in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.) containing 10% heat-inactivated fetal bovine serum (10437028;
Gibco; Thermo Fisher Scientific, Inc.) in a 95% humidified
incubator with 5% CO2 at 37°C. The cultures were split
every second day by dilution to a concentration of 2×105
cells/ml.
Construction of the RSK1 plasmid and
blocking RSK1 expression with small interfering RNA (siRNA)
The full-length human RSK1 cDNA sequence was
amplified from the cDNA library of MOLM-13 cells by PCR using the
following two primers: Sense, 5′-AAGGTACCACCATGGAGCAGGATCCCAAGC-3′,
and antisense, 5′-CTCTCGAGTCACAGGGTGGTGGATGGC-3′. Taq DNA
polymerase (Thermo Fisher Scientific, Inc.) was used for
amplification. The RSK1 cDNA sequencing was inserted into the
pcDNA3.1 plasmid (Invitrogen; Thermo Fisher Scientific, Inc.). In
order to silence RSK1 expression by siRNA, 21 nt complementary RNA
with symmetrical 2 nt overhangs was obtained from Shanghai
GenePharma Co., Ltd. (Shanghai, China). The DNA sequence was
5′-CCCAACATCATCACTCTGAAA-3′. The siRNA-RSK1 NC was also transferred
into leukemic cells, which had the same number and type of bases as
the siRNA-RSK1, but the arrangement was different. The DNA sequence
was 5′-CCCAACCTAATCTACCTGAAA-3′. The plasmid and siRNA was
transferred into MOLM-13 and Kasumi-1 cells, respectively.
Cell viability assay
MOLM-13 and Kasumi-1 cells were seeded into 96-well
culture plates at a density of 1.5×104 cells/well in 100
µl medium and were treated with different concentrations (15–60 µM)
of luteolin for 24, 48 and 72 h. The pcDNA3.1-RSK1 plasmid was
initially constructed, which overexpressed RSK1. Following
transfection of cells with pcDNA3.1-RSK1 or empty plasmid for 24 h,
the cells were treated with 30 µM luteolin, which was the
representable value near the half maximal inhibitory concentration
(IC50), for 24 h. Viable cells were then evaluated with
the CCK-8 assay according to the manufacturer's instructions.
Briefly, CCK-8 solution (50 µl/well) was added to the cells in
24-well plates (0.6×104 cells/well), and the samples
were incubated at 37°C for 4 h. Subsequently, the optical density
of each well was read at 450 nm using a microplate reader (ELx800
absorbance reader; BioTek Instruments, Inc., Winooski, VT, USA).
The viability was assessed with the following equation: Viability
(%) = Experimental / Control × 100%.
Cell apoptosis assay
Apoptotic and dead cell counts were performed using
a FITC-labeled Annexin V and PI staining by flow cytometry. The
cells were collected and resuspended in binding buffer at a
concentration of 3×106/ml. Next, 100 µl cell suspension
was added to 5 µl Annexin V-FITC and 10 µl PI, and the sample was
mixed for 15 min in the dark at room temperature. Subsequently, 400
µl phosphate-buffered saline was added to the solution. A FACScan
instrument (BD Biosciences, Franklin Lakes, NJ, USA) was used to
count the cells (1×103) at an excitation wavelength of
490 nm and determine the cell apoptosis. CellQuest software version
5.1 (BD Biosciences) was used for data collection and
processing.
Western blot analysis
SDS Lysis Buffer (P0013G; Beyotime Institute of
Biotechnology) was added to MOLM-13 and Kasumi-1 cells to isolate
the proteins. Protein concentration was detected using a
bicinchoninic acid kit (P0009; Beyotime Institute of
Biotechnology). For each western blot sample, 20 µg protein was
load onto each lane and resolved through 12% SDS-PAGE, and then
electrophoretically transferred to a polyvinylidene difluoride
membrane (EMD Millipore, Billerica, MA, USA). Next, the membrane
was probed with primary antibodies against human RSK1 (Ser221;
ab10695, Abcam), BAD (Ser112; 9291; Cell Signaling Technology,
Inc.) and KIBRA (Ser947; ab107637; Abcam), with GAPDH (ab8245;
Abcam) used as the internal control. The antibodies were diluted
1:1,000 and incubated at room temperature for 2 h. Next, the
membrane was incubated with horseradish peroxidase-conjugated goat
anti-rabbit secondary antibody (ab6721; Abcam; dilution, 1:5,000)
at room temperature for 1.5 h. The membranes were visualized with
SignalFire™ ECL Reagent (#6883; Cell Signaling Technology, Inc.),
and the band intensity was measured by Quantity One software 3.0
(Bio-Rad Laboratories, Inc. Hercules, CA, USA).
Migration assay
The migration of MOLM-13 and Kasumi-1 cells was
assayed using a Boyden chamber (Qilinbeier Bio Co., Haimen, China).
Cells were treated with different concentrations of luteolin and
then plated at 1×106 cells/ml in the upper chamber, with
10% RPMI-1640 medium added to the lower chamber. After incubating
for 24 h, non-migration cells were removed from the top well, while
the bottom cells were collected and counted by trypan blue
exclusion assay in triplicate.
Statistical analysis
All computations were carried out using the SPSS
v18.0 for Windows (SPSS, Inc., Chicago, IL, USA). Data were
expressed as mean ± standard deviation. One-way analysis of
variance was used to analyze the differences, and Kaplan-Meier
survival analysis was used to analyze patient survival rate.
P<0.05 was considered to indicate a statistically significant
difference.
Results
High expression of RSK1 is observed in
newly diagnosed AML cases
Aberrant RSK signaling is integral for several types
of cancer, such as breast cancer and melanoma. In the present
study, bone marrow samples from 30 AML patients (12 males and 18
females) at the newly-diagnosed and CR stages were analysed for
RSK1 expression using the RT-qPCR method. The clinicopathological
characteristics of the 30 AML patients are shown in Table I. The expression of RSK1 was found to
be associated with the French-American-Britain (FAB) subtype and
the percentage of minimal residual disease (MRD) cells. There were
no statistically significant differences in other clinical
features, including the patient gender and age, as well as the
leukocyte, hemoglobin and platelet counts, between individuals with
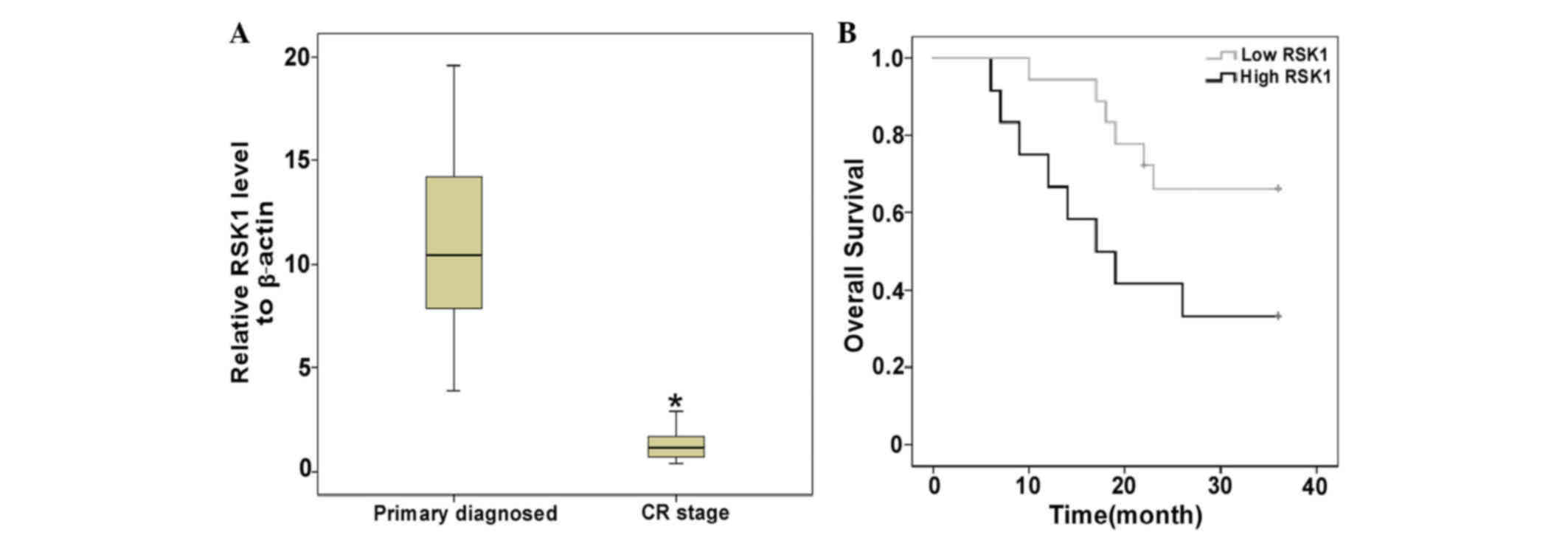
high and low RSK1 expression. Fig. 1A
shows a markedly higher expression of RSK1 in newly diagnosed
patients compared with that in patients at the CR1 stage.
Furthermore, the prognostic significance of RSK1 expression was
assessed in the 30 adult AML patients according to the clinical
follow-up records. Kaplan-Meier survival analysis indicated that
the high RSK1 expression group tended to have a shorter overall
survival (OS) compared with that of the low RSK1 expression group
(P=0.038; Fig. 1B).
 | Table I.Association of RSK1 expression with
clinicopathological characteristics of 30 acute myeloid leukemia
patients. |
Table I.
Association of RSK1 expression with
clinicopathological characteristics of 30 acute myeloid leukemia
patients.
|
|
| RSK1 expression
(n) |
|
|---|
|
|
|
|
|
|---|
| Clinical
variables | Number | Low (<10) | High (≥10) | P-value |
|---|
| Gender |
|
|
| 0.559 |
|
Male | 12 | 8 | 4 |
|
|
Female | 18 | 10 | 8 |
|
| Age (years) |
|
|
| 0.884 |
|
<60 | 17 | 10 | 7 |
|
|
≥60 | 13 | 8 | 5 |
|
| Leukocytes
(cells/µl) |
|
|
| 0. 146 |
|
>10,000 | 15 | 7 | 8 |
|
|
≤10,000 | 15 | 11 | 4 |
|
| Hemoglobin
(g/dl) |
|
|
| 0.667 |
|
>80 | 16 | 9 | 7 |
|
|
≤80 | 14 | 9 | 5 |
|
| Platelet
(cells/µl) |
|
|
| 0.447 |
|
>25,000 | 20 | 11 | 9 |
|
|
≤25,000 | 10 | 8 | 2 |
|
| FAB |
|
|
| 0.227 |
|
M1-6 | 29 | 18 | 11 |
|
| M7 | 1 | 0 | 1 |
| MRD cells (%) |
|
|
| 0.002 |
|
<0.25 | 14 | 12 | 2 |
|
|
≥0.25 | 16 | 6 | 10 |
|
Effect of luteolin on viability,
chemosensitivity and apoptosis in leukemia cells
RSKs serve a role in various cellular processes,
including gene expression, cell survival, apoptosis and
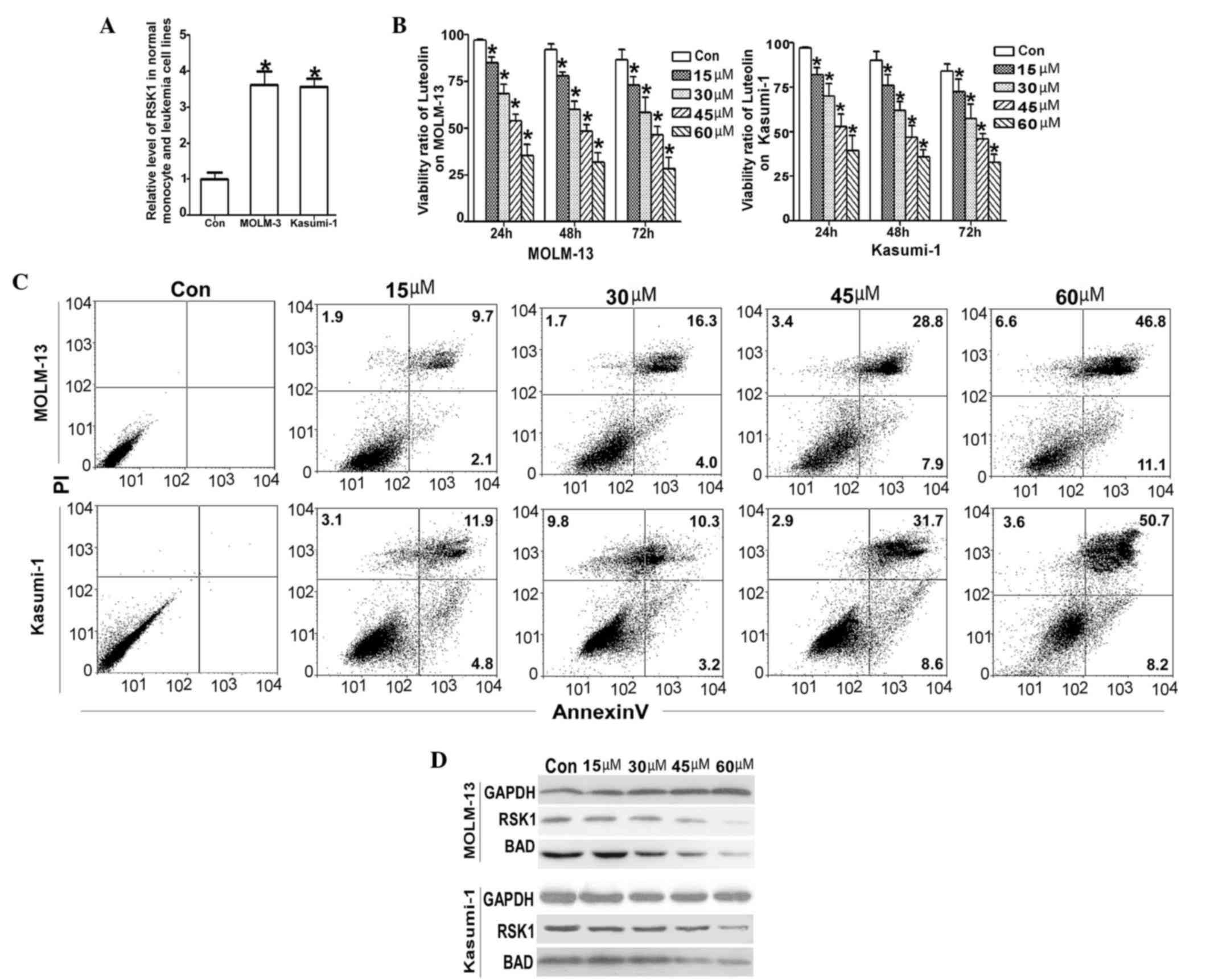
proliferation. The present study verified that, compared with
normal mononuclear cells, RSK1 overexpression was observed in the
leukemic cell lines MOLM-13 and Kasumi-1 (Fig. 2A). In order to investigate whether
luteolin effects leukemic cells through RSK1, the cell viability of
MOLM-13 and Kasumi-1 were first investigated upon treatment with
luteolin. The luteolin concentration range used in the present
experiment was 15–60 nM for 24, 48 and 72 h. The IC50
for each concentration was determined and averaged for the
triplicate experiments, and the final IC50 values of
luteolin on MOLM-13 and Kasumi-1 cells were found to be 34.75 and
34.05 µM, respectively. Luteolin treatment resulted in a marked
reduction of cell proliferation in a dose-dependent manner
(Fig. 2B).
To understand the mechanism by which luteolin caused
viability loss in MOLM-13 and Kasumi-1 cells, apoptosis experiments
were performed (Fig. 2C). The
resultant viability loss was accompanied by decreased
phosphorylation of proteins targeted by RSKs, such as BAD (21,22), which
is a proapoptotic protein (Fig. 2D).
Collectively, the current findings indicate that luteolin was able
to induce the viability loss, while apoptosis of MOLM-13 and
Kasumi-1 cells was associated with RSK1-associated apoptotic
injury.
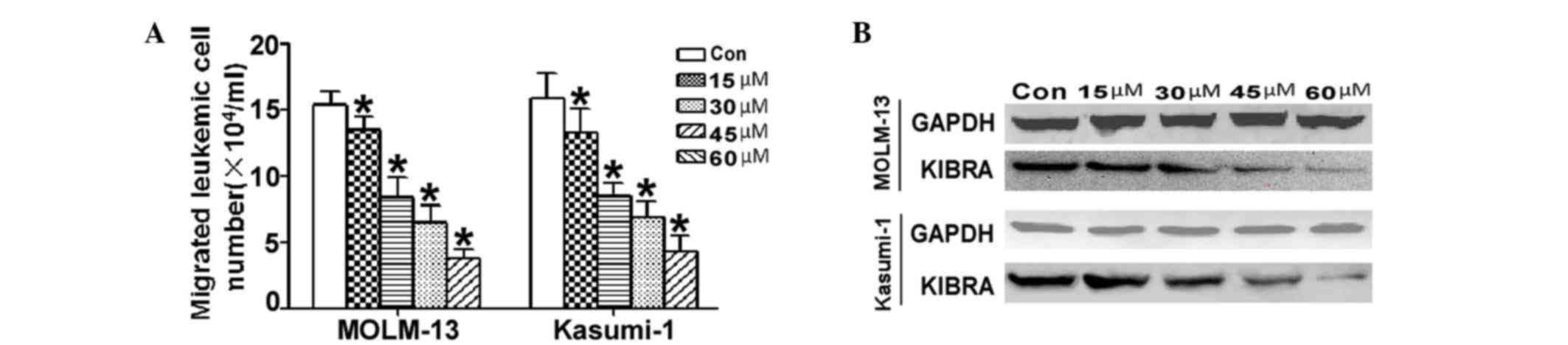
Effect of luteolin on migration of
leukemia cells
Leukocytes migrate into and out of blood vessels at
multiple points during their development and maturation, and during
immune surveillance. Migration is also an important part of
leukemia progression. To determine the influence of luteolin on
in vitro cell migration, the Boyden chamber assay was
performed in MOLM-13 and Kasumi-1 cells. As shown in Fig. 3A, the results indicated that there was
significantly less migration through the basement membrane
following treatment with luteolin, in a dose-dependent manner. Cell
migration of MOLM-13 and Kasumi-1 cells drove us to examine the
possible biological functions of luteolin in leukemic cells,
particularly the effect of this treatment on cell migration
(Fig. 3B). The results demonstrated
that the phosphorylated level of KIBRA also decreased in a
dose-dependent manner.
Overexpression of RSK1 reverses the
effects of luteolin treatment on leukemic cells
To examine the role of RSK1 in the generation of
luteolin-induced functional responses, MOLM-13 and Kasumi-1
leukemia cells were treated in the presence of luteolin with the
RSK1-overexpressing plasmid or control plasmid. The overexpression
of RSK1 mRNA and protein expression levels were evaluated by
RT-qPCR and western blot analysis, respectively (Fig. 4A and B). As determined by RT-qPCR, the
RSK1 mRNA level was upregulated by 15-fold, and the protein level
in the RSK1 overexpression group was 3-fold greater than that of
the control. The cell viability was also determined by CCK-8
assays, as shown in Fig. 4C. For the
luteolin with RSK1-overexpressing plasmid group, transfection with
the RSK1-overexpressing plasmid significantly counteracted the
effect of luteolin on cell viability when compared with the
luteolin alone group (Fig. 4C).
Similarly, when induction of apoptosis was assessed, the apoptotic
ratio of the luteolin and RSK1-overexpressing group was found to be
significantly decreased (Fig. 4D),
with a similar effect observed in the migration ability of cells
(Fig. 4E).
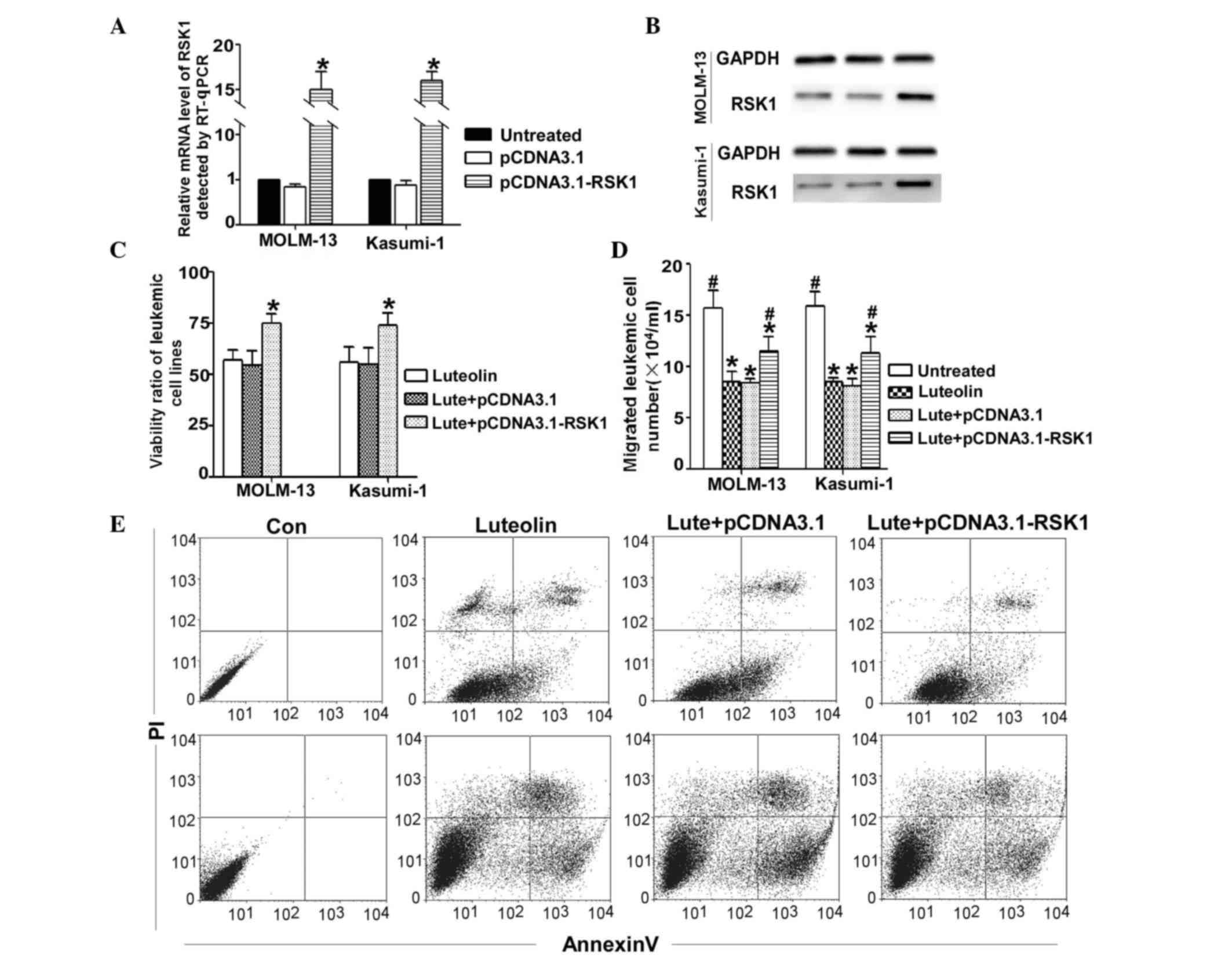 | Figure 4.Transfection with an
RSK1-overexpressing plasmid for 24 h reverses the luteolin-induced
apoptosis in MOLM-13 and Kausmi-1 cells. Cells were treated with
luteolin alone (30 µM), or combined with a RSK1-overexpressing
plasmid or empty vector for 24 h. (A) mRNA and (B) protein
expression levels of RSK1 were evaluated by RT-qPCR and western
blot analysis, respectively. (C) Cell viability detected by the
cell counting kit-8 method indicated that RSK1 overexpression
reversed the effects of luteolin on the cells, with a significantly
higher viability ratio detected (*P<0.01 vs. luteolin alone).
(D) Cell migration assay indicated that transfection with the
RSK1-overexpressing plasmid reversed the effects of luteolin
(*P<0.01 vs. untreated group; #P<0.01 vs. luteolin
alone group). (E) Changes in the apoptotic ratio of MOLM-13 and
Kasumi-1, as detected by flow cytometry. In the RSK1-overexpressing
group, the apoptotic ratio was markedly reduced compared with the
other two groups. RSK1, ribosomal S6 kinase 1; Lute, luteolin; Con,
control; PI, propidium iodide; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction. |
Knocking down of RSK1 has the same
effects on leukemic cells as luteolin treatment
To further verify whether RSK1 serves an important
role in the luteolin-induced functional responses, the expression
of RSK1 was blocked by siRNA transfection in MOLM-13 and Kasumi-1
leukemia cells. The siRNA silenced the expression of RSK1 on the
mRNA and protein levels, as evaluated by RT-qPCR and western blot
analysis, respectively (Fig. 5A and
B). Following transfection with the siRNA-RSK1 in the leukemic
cell lines, the cell viability was determined by CCK-8 assay. The
results demonstrated that the viability of the siRNA-RSK1 group was
significantly lower compared with that of the untreated control and
NC groups (Fig. 5C). Similarly, when
induction of apoptosis was assessed, the results identified that
the apoptotic ratio of the siRNA group was significantly increased
(Fig. 5D). By contrast, the migration
ability decreased markedly upon silencing of RSK1 with siRNA
(Fig. 5E).
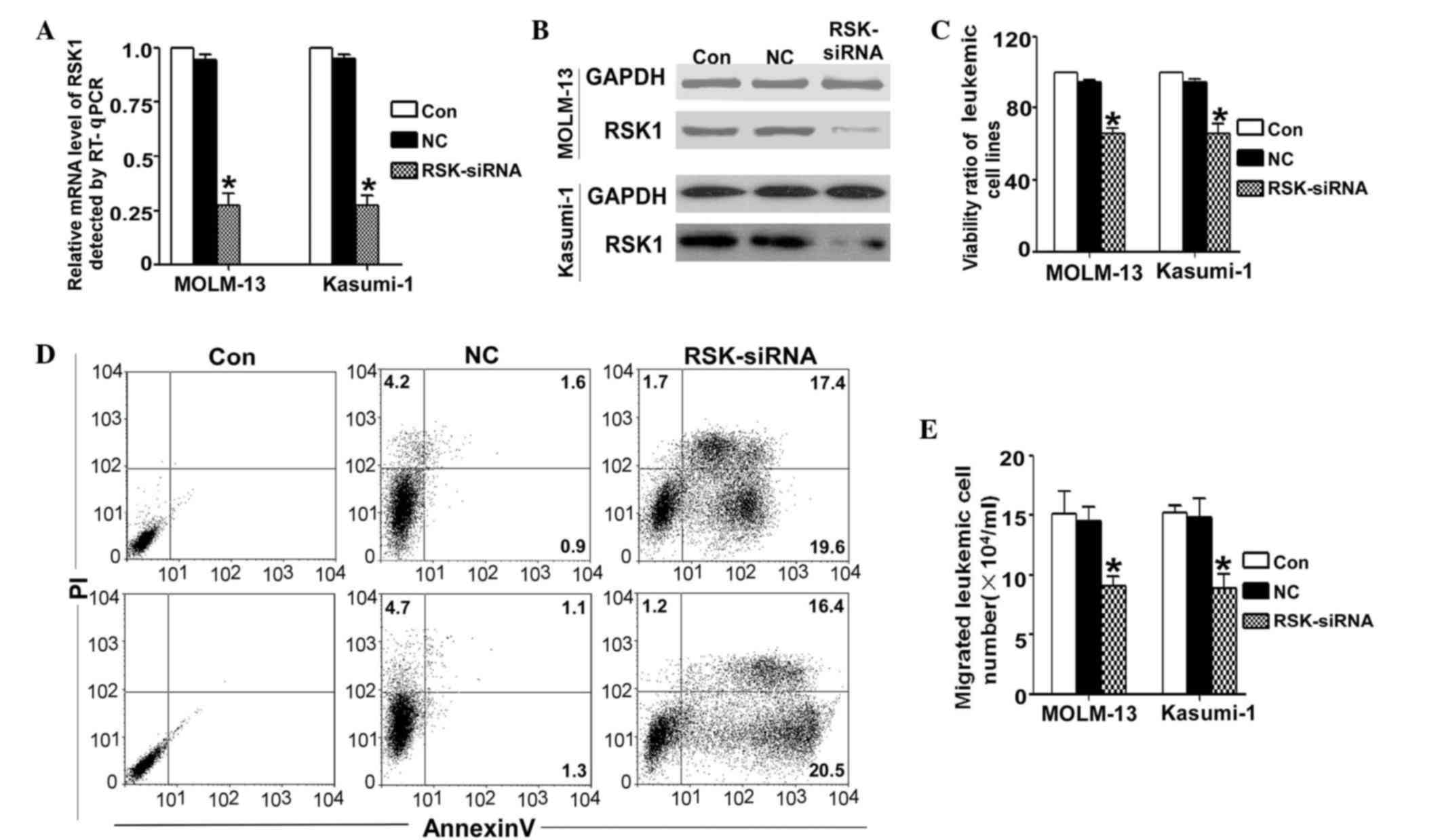 | Figure 5.Silencing of RSK1 by transfection
with siRNA-RSK1 for 24 h had a similar effect to that of luteolin
treatment on MOLM-13 and Kausmi-1 cells. (A) mRNA and (B) protein
expression levels of RSK1 were RT-qPCR and western blot analysis,
respectively. (C) Cell viability was detected by the cell counting
kit-8 method following transfection with siRNA-RSK or NC for 24 h.
The viability ratio was significant lower compared with that of the
two control groups. (D) Changes in the apoptotic ratio of MOLM-13
and Kasumi-1 cells, as detected by flow cytometry. In the
siRNA-RSK1 group, the apoptotic ratio was markedly higher compared
with that of the other two groups. (E) Cell migration ability of
the two cell lines, showing that the siRNA-RSK1 group presented
markedly lower migration compared with the control group.
*P<0.001 vs. the untreated control group. RSK1, ribosomal S6
kinase 1; Con, untreated control; NC, negative control; PI,
propidium iodide; siRNA, small interfering RNA; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction. |
Discussion
Proliferation, survival and migration of cells are
regulated by different intracellular signaling pathways. Among
these, the Ras/mitogen-activated protein kinase kinase (MEK)/ERK
signaling pathway has been established (23). Novel treatments targeting the RSK,
PI3K/Akt and MEK/ERK signaling pathways are currently under
evaluation in numerous types of tumors in clinical trials (24–26).
AML is an aggressive hematological disorder
characterized by an abnormal increase of immature hematopoietic
progenitor cells known as ‘blasts’, which lose their ability to
differentiate normally and continue to proliferate. The downstream
effectors MEK and Erk1/2 are constitutively activated in the
majority of AML cases. The present study focused on the members of
the RSK family of Ser/Thr kinases, which are downstream effectors
of the MAPK/ERK pathway, whose activities regulate cellular
effectors that promote cell growth, survival and migration
(27). This kinase family consists of
four human isoforms (RSK1-4), which possess a 73–80% amino acid
identity and have the same general structure, and two structurally
associated homologues (1). RSKs
function downstream of MEK, ERK and protein kinase C, and are
frequently activated in various types of cancer, including
leukemia, and this activation may be triggered by steroids,
insulin, epidermal growth factor and estrogen. Previous studies
have demonstrated that RSK1 is overexpressed in prostate (28), breast (29) and colon cancer tissues (30).
In the present study, the association between the
expression of RSK1 and the outcome of 30 cases AML was analyzed.
Bone marrow mononuclear cells from 30 patients with AML were
isolated, and the mRNA level of RSK1 was detected by RT-qPCR. The
percentage of MRD cells is the main independent prognostic factor
for leukemia, and the expression of RSK1 was found to be closely
associated with the MRD percentage in the present study. Regarding
the FAB subtypes of patients in the current study, only one patient
had an M7 subtype. Future studies should thus include a larger
number of cases and samples. According to the present results, the
expression of RSK1 was not associated with the clinical
characteristics of the AML patients, such as hyperleukocytosis,
severe anemia, thrombocytopenia and older age (>60 years). High
expression of RSK1 was also observed in the newly diagnosed AML
samples compared with the CR stage (Fig.
1A). Kaplan-Meier survival analysis described that a higher
RSK1 level was correlated with a shorter OS of patients (Fig. 1B).
AML is an extremely heterogeneous disease with a
variable long term survival rate ranging between 20 and 90%. The
major interventions of conventional medicine failed to have its
effects because of its side effects. An increasing number of
studies have been focusing on Chinese herb extracts, with certain
compounds recently reported as molecular targeted anti-cancer
drugs, such as luteolin, a novel inhibitor of RSK. Luteolin has
been the focus of clinical-translational interest in anticancer
treatment (31–33). In the present study, the effects of
pharmacological inhibition of the RSK1 inhibitor, luteolin, on
leukemic cell lines were investigated. As shown in Fig. 2A, luteolin exhibited potent
suppressive effects on MOLM-13 and Kasumi-1 cells in a
dose-dependent manner, with IC50 values of 34.75 and
34.05 µM, respectively.
Cancer cells acquire alternations for enhanced
survival and become apoptosis-resistant to anticancer therapies.
Therefore, the induction of apoptosis can be an effective approach
for inhibiting uncontrolled cell proliferation and survival of
malignant tumor cells. Compared with the untreated control cells,
luteolin treatment led to cell apoptosis in the present study
(Fig. 1B). A mechanism through which
RSK interacts to cause apoptosis is the alteration of BAD
phosphorylation, thus triggering its proapoptotic functions. BAD is
a unique BH3-only protein, since its functions are tightly
regulated by serine phosphorylation (34). In the hypophosphorylated form, BAD
interacts with Bcl-2 or Bcl-extra large to neutralize their
anti-apoptotic functions. RSK has been shown to phosphorylate BAD
on the Ser112 site. The present study observed that luteolin was
able to induce BAD dephosphorylation in a dose-dependent manner.
Therefore, it is likely that luteolin inhibited RSK1 with the
hypophosphorylated form of BAD.
It has been reported that RSKs serve important roles
in cell migration as well (35). Yang
et al (36) reported that RSK1
specifically phosphorylates KIBRA, a memory performance-associated
protein, and also modulates its migratory activity in MDA-MB-231
breast cancer cells. Recently, KIBRA was demonstrated to be a
positive regulator of the Hippo signaling pathway activity in
mammalian cells (37). The present
study demonstrated that luteolin, an RSK1 inhibitor, impaired the
migration of MOLM-13 and Kasumi-1 cells in a dose-dependent manner
through dephosphorylation of KIBRA. Thus, there results indicate
that KIBRA, the downstream factor of RSK1, serves an important role
in the mobility of leukemia cells. The current study next explored
whether RSK1 expression is a key factor in the effect of luteolin
treatment on leukemic cell proliferation and migration. An
RSK1-overexpressing plasmid was transfected in leukemic cell lines
for 24 h, and cells were then treated with luteolin. The viability,
apoptosis and migration analysis results supported that RSK1 serves
an important role in luteolin-modulated cell proliferation and
migration of AML cells.
In conclusion, the current pooled analysis
demonstrated that RSK1 mRNA expression was significantly higher in
primary AML samples compared with the expression at the CR stage
following standard chemotherapy. In addition, a shorter survival
time was correlated with high RSK1 expression. Luteolin treatment
triggered RSK-dependent antileukemic responses with
dephosphorylation of BAD or KIBRA. Therefore, luteolin, a novel RSK
inhibitor, has significant anti-leukemia effects in vitro
and may function as a possible therapeutic in the future; however,
further investigation in clinical trials is first required to
confirm these results.
Acknowledgements
The present study was supported by grants from the
Natural Science Foundation of Guangdong Province (grant nos.
S2012010008748 and S2013010014715) and the National Natural Science
Foundation of China (grant no. 81400103).
References
|
1
|
Lara R, Seckl MJ and Pardo OE: The p90 RSK
family members: Common functions and isoform specificity. Cancer
Res. 17:5301–5308. 2013. View Article : Google Scholar
|
|
2
|
Romeo Y, Zhang X and Roux PP: Regulation
and function of the RSK family of protein kinases. Biochem J.
441:553–569. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kostenko S, Shiryaev A, Dumitriu G, Gerits
N and Moens U: Cross-talk between protein kinase A and the
MAPK-activated protein kinases RSK1 and MK5. J Recept Signal
Transduct Res. 31:1–9. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li D, Jin L, Alesi GN, Kim YM, Fan J, Seo
JH, Wang D, Tucker M, Gu TL, Lee BH, et al: The prometastatic
ribosomal S6 kinase 2-cAMP response element-binding protein
(RSK2-CREB) signaling pathway up-regulates the actin-binding
protein fascin-1 to promote tumor metastasis. J Biol Chem.
288:32528–32538. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Degen M, Barron P, Natarajan E, Widlund HR
and Rheinwald JG: RSK activation of translation factor eIF4B drives
abnormal increases of laminin γ2 and MYC protein during neoplastic
progression to squamous cell carcinoma. PLoS One. 8:e789792013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cohen JD, Babiarz JE, Abrams RM, Guo L,
Kameoka S, Chiao E, Taunton J and Kolaja KL: Use of human stem cell
derived cardiomyocytes to examine sunitinib mediated cardiotoxicity
and electrophysiological alterations. Toxicol Appl Pharmacol.
257:74–83. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Elf S, Blevins D, Jin L, Chung TW,
Williams IR, Lee BH, Lin JX, Leonard WJ, Taunton J, Khoury HJ and
Kang S: p90RSK2 is essential for FLT3-ITD-but dispensable for
BCR-ABL-induced myeloid leukemia. Blood. 117:6885–6894. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sak K and Everaus H: Chemomodulating
effects of flavonoids in human leukemia cells. Anticancer Agents
Med Chem. 15:1112–1126. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Morceau F, Chateauvieux S, Orsini M,
Trecul A, Dicato M and Diederich M: Natural compound and
pharmaceuticals reprogram leukemia cell differentiation pathways.
Biotechnol Adv. 33:(6 Pt 1). 785–797. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Humisto A, Herfindal L, Jokela J, Karkman
A, Bjornstad R, Choudhury RR and Sivonen K: Cyanobacteria as a
source for novel anti-leukemic compound. Curr Pharm Biotechnol.
17:78–91. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu Y, Zhang J, Liu J, Li S, Li C, Wang W,
Ma R and Liu Y: Luteolin attenuate the d-galactose-induced renal
damage by attenuation of oxidative stress and inflammation. Nat
Prod Res. 29:1078–1082. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sung J and Lee J: Anti-inflammatory
activity of butein and luteolin through suppression of NFκB
activation and induction of heme oxygenase-1. J Med Food.
18:557–564. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Eumkeb G, Siriwong S and Thumanu K:
Synergistic activity of luteolin and amoxicillin combination
against amoxicillin-resistant Escherichia coli and mode of action.
J Photochem Photobiol B. 117:247–253. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim YS, Kim SH, Shin J, Harikishore A, Lim
JK, Jung Y, Lyu HN, Baek NI, Choi KY, Yoon HS and Kim KT: Luteolin
suppresses cancer cell proliferation by targeting vaccinia-related
kinase 1. PLoS One. 9:e1096552014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Park SH, Ham S, Kwon TH, Kim MS, Lee DH,
Kang JW, Oh SR and Yoon DY: Luteolin induces cell cycle arrest and
apoptosis through extrinsic and intrinsic signaling pathways in
MCF-7 breast cancer cells. J Environ Pathol Toxicol Oncol.
33:219–231. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chian S, Li YY, Wang XJ and Tang XW:
Luteolin sensitizes two oxaliplatin-resistant colorectal cancer
cell lines to chemotherapeutic drugs via inhibition of the Nrf2
pathway. Asian Pac J Cancer Prev. 15:2911–2916. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen KC, Chen CY, Lin CR, Yang TY, Chen
TH, Wu LC and Wu CC: Luteolin attenuates TGF-β1-induced
epithelial-mesenchymal transition of lung cancer cells by
interfering in the PI3K/Akt-NF-κB-Snail pathway. Life Sci.
93:924–933. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Reipas KM, Law JH, Couto N, Islam S, Li Y,
Li H, Cherkasov A, Jung K, Cheema AS, Jones SJ, et al: Luteolin is
a novel p90 ribosomal S6 kinase (RSK) inhibitor that suppresses
Notch4 signaling by blocking the activation of Y-box binding
protein-1 (YB-1). Oncotarget. 4:329–345. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Falini B, Tiacci E, Martelli MP, Ascani S
and Pileri SA: New classification of acute myeloid leukemia and
precursor-related neoplasms: Changes and unsolved issues. Discov
Med. 10:281–292. 2010.PubMed/NCBI
|
|
20
|
Flygare J, Kiefer T, Miyake K, Utsugisawa
T, Hamaguchi I, Da Costa L, Richter J, Davey EJ, Matsson H, Dahl N,
et al: Deficency of ribosomal protein S19 in CD34+ cells generated
by siRNA blocks erythroid development and mimics defected seen in
Diamond-Blackfan anemia. Blood. 105:4627–4634. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun D, Huang J, Zhang Z, Gao H, Li J, Shen
M, Cao F and Wang H: Luteolin limits infarct size and improves
cardiac function after myocardium ischemia/reperfusion injury in
diabetic rats. PLoS One. 7:e334912012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee WR, Shen SC, Lin HY, Hou WC, Yang LL
and Chen YC: Wogonin and fisetin induce apoptosis in human
promyeloleukemic cells, accompanied by a decrease of reactive
oxygen species, and activation of caspase 3 and
Ca2+-dependent endonuclease. Biochem Pharmacol.
63:225–236. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Friday BB and Adjei AA: Advances in
targeting the Ras/Raf/MEK/Erk mitogen-activated protein kinase
cascade with MEK inhibitors for cancer therapy. Clin Cancer Res.
14:342–346. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Crudden C, Ilic M, Suleymanova N, Worrall
C, Girnita A and Girnita L: The dichotomy of the Insulin-like
growth factor 1 receptor: RTK and GPCR: Friend or foe for cancer
treatment? Growth Horm IGF Res. 25:2–12. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sabine VS, Crozier C, Brookes CL, Drake C,
Piper T, van de Velde CJ, Hasenburg A, Kieback DG, Markopoulos C,
Dirix L, et al: Mutational analysis of PI3K/AKT signaling pathway
in tamoxifen exemestane adjuvant multinational pathology study. J
Clin Oncol. 32:2951–2958. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Johnson AS, Crandall H, Dahlman K and
Kelley MC: Preliminary results from a prospective trial of
preoperative combined BRAF and MEK-targeted therapy in advanced
BRAF mutation-positive melanoma. J Am Coll Surg. 220:581–593.e1.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dent P: Arsenic in leukemia: A RSKy
business. Cancer Biol Ther. 14:871–872. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu G, Lee YC, Cheng CJ, Wu CF, Song JH,
Gallick GE, Yu-Lee LY, Kuang J and Lin SH: RSK promotes prostate
cancer progression in bone through ING3, CKAP2, and PTK6-mediated
cell survival. Mol Cancer Res. 13:348–357. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Stratford AL, Reipas K, Hu K, Fotovati A,
Brough R, Frankum J, Takhar M, Watson P, Ashworth A, Lord CJ, et
al: Targeting p90 ribosomal S6 kinase eliminates tumor-initiating
cells by inactivating Y-box binding protein-1 in triple-negative
breast cancers. Stem Cells. 30:1338–1348. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Goulet AC, Watts G, Lord JL and Nelson MA:
Profiling of selenomethionine responsive genes in colon cancer by
microarray analysis. Cancer Biol Ther. 6:494–503. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sato Y, Sasaki N, Saito M, Endo N, Kugawa
F and Ueno A: Luteolin attenuates doxorubicin-induced cytotoxicity
to mcf-7 human breast cancer cells. Biol Pharm Bull. 38:703–709.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bumke-Vogt C, Osterhoff MA, Borchert A,
Guzman-Perez V, Sarem Z, Birkenfeld AL, Bähr V and Pfeiffer AF: The
flavones apigenin and luteolin induce FOXO1 translocation but
inhibit gluconeogenic and lipogenic gene expression in human cells.
PLoS One. 9:e1043212014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chian S, Thapa R, Chi Z, Wang XJ and Tang
X: Luteolin inhibits the Nrf2 signaling pathway and tumor growth in
vivo. Biochem Biophys Res Commun. 447:602–608. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Thimmaiah KN, Easton JB and Houghton PJ:
Protection from rapamycin-induced apoptosis by insulin-like growth
factor-I is partially dependent on protein kinase C signaling.
Cancer Res. 70:2000–2009. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Czaplinska D, Turczyk L, Grudowska A,
Mieszkowska M, Lipinska AD, Skladanowski AC, Zaczek AJ, Romanska HM
and Sadej R: Phosphorylation of RSK2 at Tyr529 by FGFR2-p38
enhances human mammary epithelial cells migration. Biochim Biophys
Acta. 1843:2461–2470. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang S, Ji M, Zhang L, Chen Y, Wennmann
DO, Kremerskothen J and Dong J: Phosphorylation of KIBRA by the
extracellular signal-regulated kinase (ERK)-ribosomal S6 kinase
(RSK) cascade modulates cell proliferation and migration. Cell
Signal. 26:343–351. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xiao L, Chen Y, Ji M and Dong J: KIBRA
regulates Hippo signaling activity via interactions with large
tumor suppressor kinases. J Biol Chem. 286:7788–7796. 2011.
View Article : Google Scholar : PubMed/NCBI
|