Introduction
Gastric cancer (GC) is the second leading cause of
cancer-associated mortality worldwide (1). Despite substantial advances in surgery,
chemo- and radiotherapy, and targeted molecular therapy, the 5-year
survival rate of patients with advanced GC remains low. Therefore,
identifying novel diagnostic and prognostic markers, and
elucidating the mechanisms underlying disease progression are
essential for preventing and treating gastric tumorigenesis.
The Wnt/β-catenin signaling pathway is implicated in
oncogenesis, and contributes to the initiation and progression of
GC (2). In the absence of Wnt
ligands, cytoplasmic β-catenin is phosphorylated by a multi-protein
complex consisting of adenomatous polyposis coli (APC), axin,
casein kinase 1 and glycogen synthase kinase (GSK)-3β, leading to
its ubiquitination and degradation (3,4). This is
inhibited upon activation of Wnt signaling; β-catenin accumulates
in the cytoplasm and translocates to the nucleus, where it
initiates the transcription of a variety of target genes. The
accumulation of nuclear β-catenin is a hallmark of Wnt signaling
activation (5). Somatic mutations in
APC, β-catenin and axin, among other factors, are responsible for
activating the Wnt/β-catenin pathway (6–8). However,
the mechanisms by which the Wnt/β-catenin pathway is activated in
GC remain to be fully elucidated.
The tripartite motif (TRIM) family, identified as a
subfamily of the RING-type E3 ubiquitin ligase family, is involved
in a broad range of biological processes, including cell growth and
apoptosis, development and tumorigenesis (9). TRIM24, formerly known as transcriptional
intermediary factor 1α, is a member of the TRIM family, which is
characterized by a RING domain, two B-box zinc fingers and a
coiled-coin region (10). The
aberrant overexpression of TRIM24 is a prognostic factor in several
types of cancer, and promotes tumor development and progression
through various mechanisms. TRIM24 ubiquitinates and induces the
proteasome-mediated degradation of p53 (11,12), and
can also bind to chromatin and the estrogen receptor to activate
target genes associated with cell proliferation and tumor
development in breast cancer (13).
TRIM24 also promotes tumorigenesis by activating aerobic glycolysis
(14) and serves as a target of
chromosomal translocations leading to the formation of oncogenic
fusion proteins (15–17). However, the role of TRIM24 in the
development of GC and the underlying molecular mechanisms remain to
be fully elucidated. Previous studies have indicated that TRIM
proteins regulate Wnt/β-catenin signaling. TRIM29 promotes
proliferation and metastasis via Wnt/β-catenin pathway activation
in pancreatic cancer (18), whereas
TRIM24 knockdown in human HepG2 liver cancer cells downregulates
β-catenin and cyclinD1, two major downstream genes of the Wnt
pathway (19). Therefore, the present
study hypothesized that TRIM24 promotes the aggressiveness of GC by
activating Wnt/β-catenin signaling.
To confirm this hypothesis, the present study
examined the expression and functions of TRIM24 in GC cell lines
and tissue samples. It was found that TRIM24 was upregulated in GC,
which was positively correlated with the expression of β-catenin.
In addition, TRIM24 knockdown suppressed cell proliferation,
arrested cells at the G0/G1 phase, inhibited migration, invasion
and the nuclear translocation of β-catenin, and induced apoptosis.
It was confirmed that TRIM24 exerted its oncogenic functions
through activation of the Wnt/β-catenin pathway. Therefore, the
findings of the present study indicated that TRIM24 may serve as a
potential therapeutic target for GC and is important in activation
of the Wnt/β-catenin pathway during the progression of GC.
Materials and methods
Ethics statement
The present study was approved by the Ethics
Committee of the First Affiliated Hospital of Nanchang University
(Nanchang, China). Informed consent was provided by patients from
whom tissue samples were obtained for investigation.
Cell lines and culture
The human MGC803 and SGC7901 GC cell lines were
purchased from the Type Culture Collection of the Chinese Academy
of Science (Shanghai, China). GES-1, a normal human gastric mucosa
cell line, and the AGS, BGC823 and HGC-27 GC cell lines were
obtained from the Sun Yat-Sen University Cancer Center (Guangzhou,
China). All cells were cultured in Roswell Park Memorial Institute
(RPMI) 1640 medium (HyClone; GE Healthcare Life Sciences, Logan,
UT, USA) supplemented with 10% fetal bovine serum (FBS; HyClone; GE
Healthcare Life Sciences) for 24 h in a humidified chamber at 37°C
and 5% CO2.
Patients and tissue samples
A total of 4 fresh GC tissues and paired
noncancerous gastric mucosal tissues were collected by gastrectomy
from the Department of General Surgery of the First Affiliated
Hospital of Nanchang University. All the fresh samples were
immediately snap-frozen in liquid and stored at −80°.
Paraffin-embedded GC tissues (n=90) and gastric normal tissues
(n=60) were collected from the Department of Pathology of the First
Affiliated Hospital of Nanchang University between 2007 and 2009.
None of these 90 patients with GC had received neoadjuvant
chemotherapy or radiation therapy prior to surgery. The overall
survival (OS) of the patients with GC was defined as the period
between the date of surgery to the time at which the patient
succumbed to mortality from any cause, or the date of the last
follow-up if no event was documented. Tumor stage was determined
according to the 2010 American Joint Committee on Cancer criteria
(20). Histological differentiation
was based on World Health Organization criteria (21). The detailed clinical information of
the patients is summarized in Table
I.
 | Table I.Association between the expression of
TRIM24 and clinicopathological parameters of patients with gastric
cancer. |
Table I.
Association between the expression of
TRIM24 and clinicopathological parameters of patients with gastric
cancer.
|
|
| Expression of
TRIM24 |
|
|---|
|
|
|
|
|
|---|
| Variable | n | Low | High | P-value |
|---|
| Age (years) |
|
|
| 0.848 |
|
<60 | 50 | 21 | 29 |
|
|
≥60 | 40 | 16 | 24 |
|
| Gender |
|
|
| 0.294 |
|
Male | 55 | 25 | 30 |
|
|
Female | 35 | 12 | 23 |
|
| Tumor size
(cm) |
|
|
| 0.571 |
|
<5 | 47 | 18 | 29 |
|
| ≥5 | 43 | 19 | 24 |
|
| Tumor location |
|
|
| 0.605 |
|
Proximal | 32 | 12 | 20 |
|
|
Distal | 58 | 25 | 33 |
|
| Depth of
invasiona |
|
|
| 0.007c |
|
T1-T2 | 42 | 23 | 19 |
|
|
T3-T4 | 48 | 13 | 35 |
|
|
Differentiation |
|
|
| 0.176 |
| Well or
moderately | 41 | 20 | 21 |
|
|
Poor | 49 | 17 | 32 |
|
| Lauren
classification |
|
|
| 0.551 |
|
Intestinal type | 34 | 13 | 21 |
|
| Diffuse
type | 56 | 22 | 34 |
|
| TNM
stagea |
|
|
| 0.005c |
|
I–II | 43 | 27 | 16 |
|
|
III–IV | 47 | 10 | 37 |
|
| Lymph node
metastasis |
|
|
| 0.027b |
|
N0 | 39 | 21 | 18 |
|
NX | 51 | 16 | 35 |
Immunohistochemistry
The tissue samples were formalin-fixed overnight and
then paraffin-embedded at room temperature, prior to being cut into
4-µm serial sections. The sections were deparaffinized and
rehydrated by incubation in 3% hydrogen peroxide for 15 min at room
temperature. Antigen retrieval was performed by heating the
sections in citrate buffer at 95°C for 1 h. The sections were then
incubated with rabbit anti-human polyclonal TRIM24 antibody (1:200
dilution; catalog no., ab70560; Abcam, Cambridge, UK) or rabbit
anti-human monoclonal β-catenin antibody (1:100 dilution; catalog
no. 8480; Cell Signaling Technology, Inc., Danvers, MA, USA) in a
humidified chamber overnight at 4°C. Following washing three times
with phosphate-buffered saline (PBS), the sections were incubated
at room temperature with a horseradish peroxidase system and DAB
substrate (Dako, Carpinteria, CA, USA), followed by incubation with
PBS containing diaminobenzidine for 10 min at room temperature.
The tissue samples were examined by two
investigators who were blinded to the pathological information, and
immunoreactivity was scored using the German Semi-Quantitative
method (22). For TRIM24 staining,
each specimen was scored according to its staining intensity (0,
none; 1, weak; 2, moderate; 3, strong) and the percentage of
stained cells (0, 0%; 1, 1–24%; 2, 25–49%; 3, 50–74%; 4, 75–100%).
A final staining index (SI) was calculated as the product of the
intensity and percentage scores, yielding a value between 0 and 12,
which was classified as negative (−, 0–1), weak (+; 2–4), moderate
(++; 6–8) or strong (+++; 9–12). An SI ≥6 was considered a high
expression of TRIM24 and an SI<6 was considered a low expression
of TRIM24. For β-catenin, membrane or cytoplasmic staining was
considered negative, whereas nuclear expression was considered
positive. The scoring of TRIM24 and β-catenin were evaluated
independently by two board-certified clinical pathologists blinded
to the clinical parameters. Any discrepancy between the scores
assigned by the two investigators was resolved by re-evaluation and
careful discussion until a consensus was reached.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted with TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
RNA concentration was measured using spectrophotometry. cDNA was
obtained using an EasyScript First-Strand cDNA Synthesis Supermix
kit (TransGen Biotech, Inc., Beijing, China) and RT-qPCR analysis
was performed on an ABI Prism 7500 Sequence Detection system
(Applied Biosystems; Thermo Fisher Scientific, Inc.) using SYBR
Green qPCR Supermix-UDG with ROX (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
primers used for RT-qPCR analysis are listed in Table II. The target gene expression levels
were was normalized against that of the housekeeping gene,
glyceraldehyde-3-phosphate dehydrogenase, and calculated as
2−[(Cq target gene) - (Cq GAPDH)], where Cq is the
quantification cycle for each transcript (23).
| Table II.Sequences of primers for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table II.
Sequences of primers for reverse
transcription-quantitative polymerase chain reaction analysis.
| Gene | Primer |
|---|
| TRIM24 | Forward:
5′-CATATGCAGCAACAGCAACCG-3′ |
|
| Reverse: 5′-
GAAAGCCATCTGTAGGGGGT-3′ |
| β-catenin | Forward:
5′-GGATCAAACCTGACAGCCA-3′ |
|
| Reverse:
5′-GAAAACGCCATCACCACGTC-3′ |
| cyclinD1 | Forward:
5′-GATGCCAACCTCCTCAACGA-3′ |
|
| Reverse:
5′-ACTTCTGTTCCTCGCAGACC-3′ |
| c-Myc | Forward:
5′-CCCTCCACTCGGAAGGACTA-3′ |
|
| Reverse:
5′-GCGGTGCATTTTCGGTTGT-3′ |
| GAPDH | Forward:
5′-CATCACCATCTTCCAGGAGCG-3 |
|
| Reverse:
5′-TGACCTTGCCCACAGCCTTG-3′ |
Western blot analysis
The cells and tissues samples were lysed in
radioimmunoprecipitaion assay buffer (Sigma-Aldrich; Merck
Millipore, Darmstadt, Germany) supplemented with protease in
inhibitor cocktail (Roche Diagnostics GmbH, Mannheim, Germany). The
protein concentration was determined by Bradford method with bovine
serum albumin (BSA) as the standard. Equal quantities of protein
(50 µg) from each group were separated by 8% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred onto a
polyvinylidene difluoride membrane. Following incubation in
blocking buffer for 1 h at room temperature, the membrane was
incubated overnight at 4°C with the following primary antibodies:
Rabbit anti-human polyclonal TRIM24 (1:1,000 dilution; catalog no.
ab70560; Abcam), rabbit anti-human monoclonal β-catenin (1:1,000
dilution; catalog no. 8480; Cell Signaling Technology, Inc.),
rabbit anti-human monoclonal c-Myc (1:1,000 dilution; catalog no.
5605; Cell Signaling Technology, Inc.), rabbit anti-human
monoclonal cyclinD1 (1:1,000 dilution; catalog no. 2978; Cell
Signaling Technology, Inc.) and mouse anti-human monoclonal β-actin
(1:2,000; catalog no. 8H10D10; Cell Signaling Technology, Inc.).
Following three washes in Tris-buffered saline with 0.1% Tween 20,
the membrane was incubated with horseradish peroxidase-labeled
anti-rabbit or anti-mouse secondary antibody (1:3,000 dilution;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) for 1 h at room
temperature and developed using a chemiluminescence detection
system (Thermo Fisher Scientific, Inc.).
Cell transfection
Small interfering RNAs (siRNAs) targeting TRIM24 and
negative control (NC) siRNA were purchased from Shanghai GenePharma
Co., Ltd. (Shanghai, China). The TRIM24 siRNA sequence was
5′-GCUGGACUCUCUAAACAAUTT-3′. In vitro transient transfection
was performed using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
cells were divided into three groups: Blank, transfected with NC
siRNA or transfected with TRIM24 siRNA, and grown to 70–80%
confluency prior to transfection. TRIM24-knockdown was confirmed
using RT-PCR and western blot analyses. To examine the effects of
TRIM24 on Wnt/β-catenin signaling, the NC-transfected and TRIM24
siRNA-transfected cells were incubated with the Wnt/β-catenin
signaling activator, lithium chloride (LiCl; 20 mmol/l) for 24
h.
Cell proliferation assay
An MTT assay was used to assess cell proliferation,
according to the manufacturer's protocol. Briefly, the
NC-transfected and TRIM24 siRNA-transfected cells were seeded into
96-well plates at a density of 1×103 cells/well.
Following various durations (1–5 days), 20 µl MTT (Sigma-Aldrich;
Merck Millipore) was added to each well and incubated at 37°C for 4
h; dimethyl sulfoxide (150 µl; Sigma-Aldrich; Merck Millopore) was
mixed into each well for 10 min, and the absorbance at 490 nm was
measured using a microplate reader (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). Each sample had four replicates.
Colony formation assay
The NC-transfected and TRIM24 siRNA-transfected
MGC803 and HGC-27 cells (3×102 and
4×102/well, respectively) were seeded in four 6-cm
dishes and cultured for 2 weeks. Colonies (>50 cells) were
visualized using 5% crystal violet staining and counted. The
results are reported as the average of three independent
experiments.
Cell cycle and apoptosis analyses
The cells were collected 48 h following siRNA
transfection, and cell cycle phase was determined using flow
cytometry (Beckman-Coulter, Inc., Fullerton, CA, USA). The
NC-transfected and TRIM24 siRNA-transfected cells were washed in
cold PBS and fixed overnight in 1 ml of 70% ethanol. The following
day, the cells were collected, washed and stained with propidium
iodide (PI) for 30 min at 4°C. Apoptosis was quantified using an
Annexin V-Fluorescein Isothiocyanate Apoptosis Detection kit
(Sigma-Aldrich; Merck Millipore) according to the manufacturer's
instructions. Briefly, 1×106 NC-transfected and TRIM24
siRNA-transfected cells were collected 48 h following transfection,
washed twice in cold PBS, resuspended in 500 µl binding buffer,
incubated with Annexin V-PI for 15 min at room temperature and
analyzed using flow cytometry.
Transwell migration and invasion
assays
The cells were seeded into 24-well Transwell plates
with a pore size of 8 µm (Corning Incorporated, Corning, NY, USA).
The upper chamber was either left uncoated for the migration assay
or precoated with Matrigel for the invasion assay. For the
migration assay, the MGC803 (6×104) and HGC-27
(5×104) cells were seeded into the upper chamber; for
the invasion assay, 1×105 MGC803 and 1×105
HGC-27 cells were seeded into the upper chamber, respectively. The
upper chamber was filled with RPMI 1640 medium containing 10 g/l
BSA (Sigma-Aldrich; Merck Millipore) and the lower chamber was
filled with RPMI 1640 containing 25% FBS. Following incubation for
24 or 36 h at 37°C, the cells that had invaded into the lower
chamber were fixed with 4% paraformaldehyde and stained with
crystal violet for 1 h at room temperature, and counted in five
randomly-selected microscopic fields. All these experiments were
performed in triplicate.
Immunofluorescence
The cells were cultured in confocal dishes and fixed
in 4% paraformaldehyde for 20 min washed three times with PBS and
then permeabilized with 0.2% Triton X-100 for 10 min, all at room
temperature. Following blocking with 5% BSA at room temperature for
2 h, the cells were incubated with rabbit anti-human monoclonal
anti-β-catenin antibody (1:200 dilution; catalog no. 8480; Cell
Signaling Technology, Inc.) overnight at 4°C. Following washing in
PBS three times, the cells were incubated with appropriate Alexa
Fluor 488-conjugated secondary antibodies (1:500 dilution; catalog
no. ab150077; Abcam) for 1 h at room temperature, and then
counterstained with 4′,6-diamidino-2-phenylindole for 10 min.
Fluorescence images were captured using laser confocal
microscopy.
Statistical analysis
Data were analyzed using paired t-tests to compare
quantitative variables. Differences in the expression of TRIM24
between tumor and normal tissues were compared using the
Mann-Whitney U test. χ2 tests were used to assess the
correlation between the expression of TRIM24 and
clinicopathological characteristics. Survival curves were plotted
using the Kaplan-Meier method, and OS rates were compared using the
log-rank test. All analyses were performed using SPSS version 18.0
software (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
TRIM24 is overexpressed in GC cell
lines and tissues
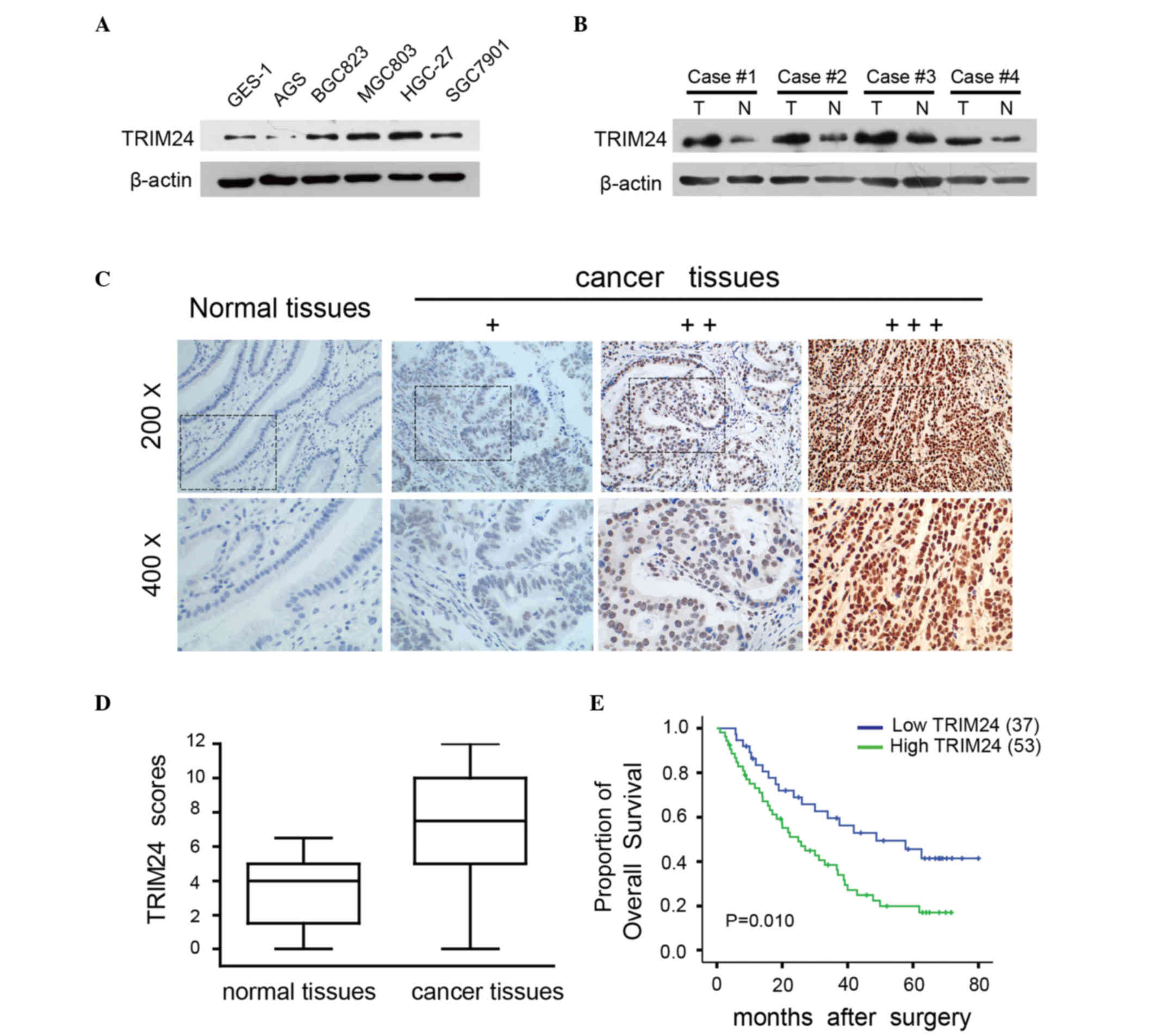
To investigate the role of TRIM24 in the progression
of GC, the present study examined the protein levels of TRIM24 in
five human GC cell lines (AGS, BGC823, MGC803, HGC-27 and SGC7901)
and one normal gastric cell line (GES-1). TRIM24 was expressed in
all cell GC lines, being particularly high in the MGC803 and HGC-27
cells, and low in the AGS cells, compared with the level in the
GES-1 cells (Fig. 1A). The expression
of TRIM24 was then randomly detected in four pairs of GC tissues
and adjacent non-tumor tissues, it was found that the expression of
TRIM24 was higher in the tumor tissues, compared with the adjacent
non-tumor tissues (Fig. 1B). The
present study subsequently performed immunohistochemistry on 60
normal gastric tissues and 90 primary GC tumor tissues to detect
the expression levels of TRIM24. It was found that TRIM24 was
expressed in the nuclei of the tumor cells, whereas the normal
tissues exhibited negative or weak nuclear staining (Fig. 1C). A significant upregulation in the
expression of TRIM24 was observed in 58.9% (53/90) of the GC
tissues, compared with the normal tissues, and this difference was
statistically significant (P<0.001; Fig. 1D).
The present study investigated the association
between the expression of TRIM24 and the clinicopathological
features of GC (Table I). A high
expression level of TRIM24 was correlated with depth of invasion
(P=0.007), tumor-node-metastasis (TNM) stage (P=0.005) and lymph
node metastasis (P=0.027). However, no correlations were found
between the expression of TRIM24 and other variables, including age
(P=0.848), gender (P=0.294), tumor size (P=0.571), location
(P=0.605), tumor differentiation (P=0.176) or Lauren classification
(P=0.551). Kaplan-Meier analysis revealed that OS rates were lower
in patients with high levels of TRIM24, compared with those with
low levels (P=0.010; Fig. 1E;
log-rank test). These data indicated that TRIM24 was overexpressed
in GC cell lines and tissues, which was associated with a more
malignant phenotype and poorer prognosis in patients with GC.
Knockdown of TRIM24 inhibits
proliferation and induces apoptosis of GC cells
To investigate the function of TRIM24 in GC, the
present study evaluated the effects of TRIM24 knockdown on the
proliferation and apoptosis of MGC803 and HGC-27 cells. The mRNA
and protein levels of TRIM24 were decreased following transfection
with TRIM24 siRNA (Fig. 2A and B).
TRIM24 knockdown inhibited proliferation, as determined using the
MTT and colony formation assays (Fig. 2C
and D). Flow cytometric analysis revealed a decrease in the
percentage of cells in the S phase and an increase in the
percentages of G1/G0 phase cells, suggesting that TRIM24 knockdown
induced G1/S arrest (Fig. 2E).
Moreover, TRIM24 knockdown increased early apoptosis in the MGC803
(10.4%) and HGC-27 (14.2%) cells, compared with the control MGC803
(4.2%) and HGC-27 (6%) cells (Fig.
2F). These results indicated that TRIM24 had an oncogenic role
in the GC cells.
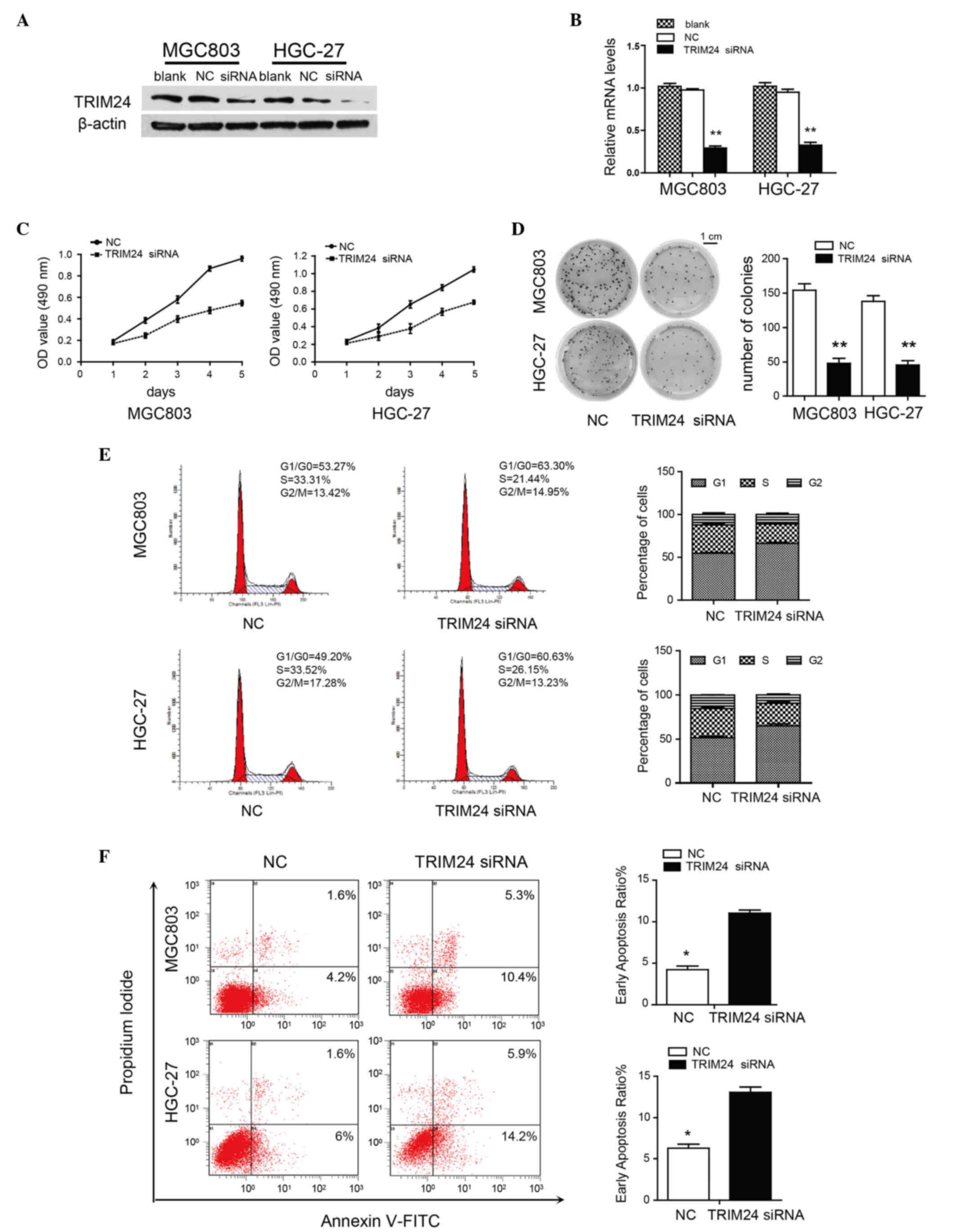 | Figure 2.Knockdown of TRIM24 inhibits GC cell
proliferation and induces apoptosis. (A) TRIM24 knockdown in MGC803
and HGC-27 cells, evaluated using western blot analysis. β-actin
was a loading control. (B) mRNA expression of TRIM24 transfected
with NC or TRIM24 siRNA, normalized against GAPDH. (C) Viability of
cells transfected with NC or TRIM24 siRNA, determined using an MTT
assay. (D) Growth of cells transfected with NC or TRIM24 siRNA,
determined with a colony formation assay. (E) Flow cytometric
analysis of cell cycle in GC cells transfected with NC or TRIM24
siRNA. (F) Apoptosis of GC cells, determined using flow cytometry.
Data are shown as the mean ± standard deviation of three
independent experiments. *P<0.05; **P<0.01. TRIM24,
tripartite motif-containing 24; GC, gastric cancer; siRNA, small
interfering RNA; NC, negative control; OD, optical density; FITC,
fluorescein isothiocyanate. |
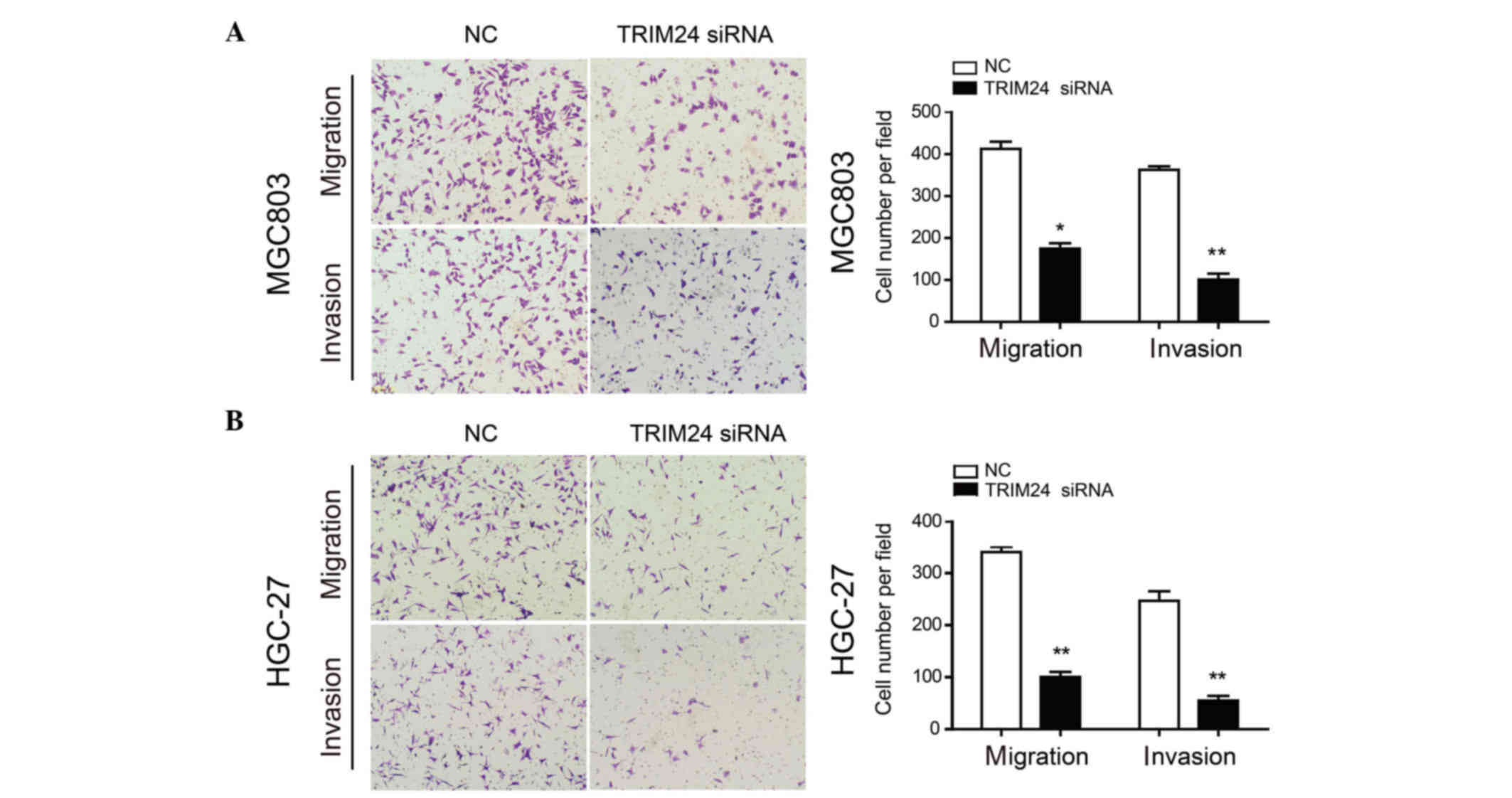
Knockdown of TRIM24 suppresses GC cell
migration and invasion
As the expression of TRIM24 was correlated with
lymph node metastasis, the present study performed Transwell
migration and invasion assays to evaluate the role of TRIM24 in
these processes. TRIM24 knockdown inhibited the migration and
invasion of MGC803 and HGC-27 cells, compared with the control
cells (Fig. 3A and B), demonstrating
that TRIM24 promoted GC cell migration and invasion.
TRIM24 regulates the Wnt/β-catenin
signaling pathway
Aberrant activation of the Wnt/β-catenin signaling
reportedly contributes to the development and progression of GC
(2,24). Therefore, the present study examined
the association between the expression of TRIM24 and β-catenin in
patients with GC. It was found that β-catenin was accumulated
predominantly in the nuclei of the GC cells, and a higher level of
TRIM24 was associated with elevated nuclear expression of β-catenin
(Fig. 4A). To further investigate the
correlation between TRIM24 and β-catenin, immunohistochemical
analysis was used to detect the expression of TRIM24 and β-catenin
in the same sample. Correlation analysis revealed that TRIM24 was
positively associated with the nuclear expression of β-catenin;
β-catenin positive staining was observed in 43 of the high
TRIM24-expressing tumors, but only in nine of the low
TRIM24-expressing tumors (P<0.001; r=0.566; Fig. 4B).
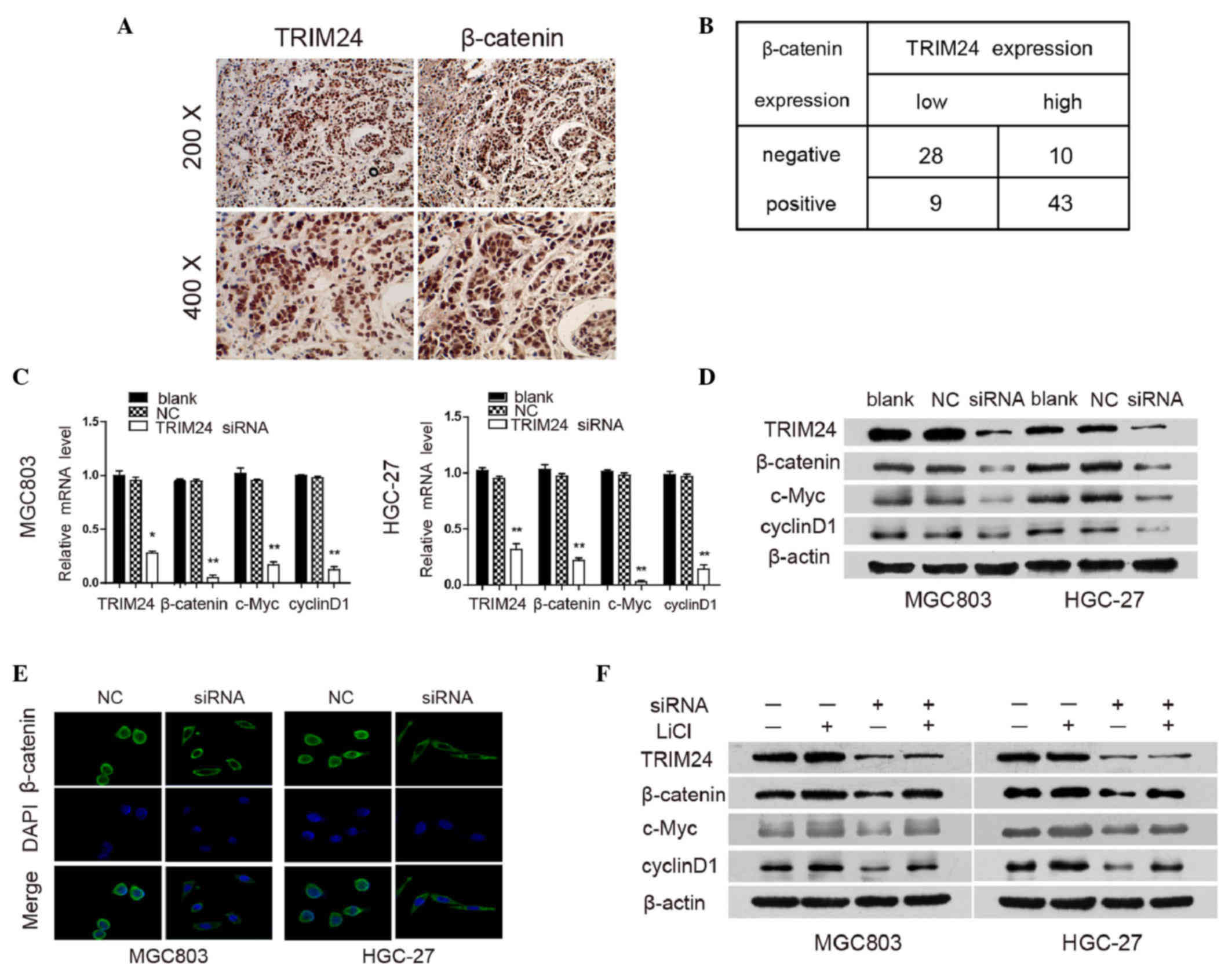 | Figure 4.TRIM24 regulates the Wnt/β-catenin
signaling pathway. (A) Representative images of GC tissues showing
concordant positive staining of TRIM24 and β-catenin in the same
sample. (B) Quantitative analysis of expression levles of TRIM24
and β-catenin in 90 GC samples. (C) mRNA expression levels of
TRIM24, β-catenin, c-Myc and cyclinD1 in GC cells transfected with
NC or TRIM24 siRNA were analyzed using reverse
transcription-quantitative polymerase chain reaction analysis, with
GAPDH used as an internal control. (D) Protein expression levels of
TRIM24, β-catenin, c-Myc and cyclinD1 in GC cells transfected with
NC or TRIM24 siRNA, as determined using western blot analysis.
β-actin was used as internal loading control. (E)
Immunofluorescence staining of subcellular localization of
β-catenin in cells. (F) Protein levels of TRIM24, β-catenin, c-Myc
and cyclinD1 in GC cells transfected with NC or TRIM24 siRNA (+/−
20 mmol/l LiCl for 24 h), using western blot analysis. Data are
presented as the mean ± standard deviation of three independent
experiments.*P<0.05; **P<0.01. TRIM24, tripartite
motif-containing 24; GC, gastric cancer; siRNA, small interfering
RNA; NC, negative control. |
To clarify the role of Wnt/β-catenin signaling in
the TRIM24-mediated progression of GC, the present study examined
the expression levels of downstream genes in the Wnt/β-catenin
signaling pathway following TRIM24 knockdown using RT-qPCR and
western blot analyses. The knockdown of TRIM24 resulted in the
downregulation of β-catenin, cyclinD1 and c-Myc at the mRNA and
protein levels (Fig. 4C and D).
Furthermore, TRIM24 knockdown significantly reduced nuclear
β-catenin accumulation in MGC803 and HGC-27 cells, compared with
the control groups, as detected using immunofluorescent staining
(Fig. 4E), indicating that the
knockdown of TRIM24 inhibited the Wnt/β-catenin pathway through
preventing the nuclear translocation of β-catenin.
To further confirm the above findings, LiCl, which
inhibits GSK-3β activity, was used to activate Wnt/β-catenin
signaling in GC cells. LiCl treatment induced the upregulation of
β-catenin, cyclinD1 and c-Myc, and also abrogated the effects of
TRIM24 knockdown on the Wnt/β-catenin pathway (Fig. 4F). These data suggested that TRIM24
functioned as a positive regulator of Wnt/β-catenin signaling,
which consequently resulted in an aggressive phenotype in GC.
Discussion
In the present study, it was demonstrated that
TRIM24 was overexpressed in GC cells and tissues, and its high
expression was correlated with tumor invasion, advanced TNM stage,
lymph node metastasis and shorter survival rates. Through
functional investigations, it was found that the downregulation of
TRIM24 suppressed cell proliferation, migration and invasion,
delayed cell cycle progression and induced apoptosis. Furthermore,
a positive correlation was found between the expression of TRIM24
and β-catenin in GC tissues, and TRIM24 exerted its oncogenic
effects by positively regulating the Wnt/β-catenin signaling
pathway.
Previous studies have shown that TRIM24 is an
oncogene, focused on promoting tumor growth, migration and
metastasis. The overexpression of TRIM24 has been reported in
several types of human tumor, and has been associated with
increased malignant behavior and poor prognosis in cancer,
including hepatocellular carcinoma (19), breast cancer (25), head and neck squamous cell carcinoma
(26), glioma (27) non-small cell lung cancer (28) and bladder cancer (29). In the present study, the
immunohistochemical analysis showed that TRIM24 was significantly
overexpressed in GC tissues. A high expression level of TRIM24 was
closely associated with the depth of invasion, advanced TNM stage
and increased lymph node metastasis, indicating that TRIM24
functioned as an oncogene to promote the invasion, metastasis and
progression of GC. However, no correlation was detected between
TRIM24 and tumor differentiation or Lauren classification, which
suggested that TRIM24 may not be involved in GC differentiation.
The finding that high expression levels of TRIM24 were associated
with poor survival rates suggested that TRIM24 was critical in the
pathogenesis and development of GC and may serve as a prognostic
biomarker.
Previous studies investigated the role of TRIM24 in
the progression of tumorigenesis and the mechanisms by which TRIM24
exerts its oncogenic functions. It has been reported that the
downregulation of TRIM24 results in the inhibition of cell
proliferation combined with a significant enhancement of apoptosis
through the modulation of B cell lymphoma-2, caspase 3 and poly
(ADP-ribose) polymerase in colorectal cancer cells (30). TRIM24 binds to the phosphoinositide
3-kinase (PI3K) promoter to activate PI3K/Akt signaling, leading to
the upregulation of downstream targets, including nuclear
factor-κB, and the induction of cell proliferation and
chemoresistance (27). In addition,
TRIM24 has been reported to interact with p53 and control the level
of phosphorylated p53 in an autoregulatory feedback loop (31). The previous studies, demonstrated that
TRIM24 can modify cell proliferation, migration, invasion and
apoptosis. These findings, in addition to the observations of the
present study, demonstrated how TRIM24 promotes the malignant
behavior of cancer cells.
The aberrant activation of Wnt/β-catenin signaling
is associated with tumorigenesis in GC. Of note, the present study
found that TRIM24 and nuclear β-catenin were often concomitantly
overexpressed in GC tissues, suggesting a regulatory role for
TRIM24 in β-catenin signaling. This was also supported by the
observation that TRIM24 knockdown decreased the expression levels
of the Wnt/β-catenin target genes, β-catenin, cyclinD1 and c-Myc.
CyclinD1 is overexpressed in several types of cancer and serves as
a regulator of cell cycle progression through the G1/S checkpoint
(32–34). c-Myc is a nuclear phosphoprotein,
which regulates the cell cycle and apoptosis (35). Thus, the inhibition of cell
proliferation and enhancement of apoptosis resulting from TRIM24
knockdown may be partly due to the downregulation of cyclinD1 and
c-Myc. The Wnt/β-catenin pathway is fundamental in the
epithelial-mesenchymal transition (EMT), an important step in
oncogenic transformation, which includes cell invasion and
metastasis (36–38). The nuclear translocation of β-catenin
leads to the downregulation of E-cadherin and subsequent induction
of EMT (39), whereas the
upregulation of β-catenin is associated with the invasion and
metastasis of several types of solid tumor (40–42). A
previous study found that TRIM24 knockdown decreased the levels of
EMT-associated factors in hepatocellular carcinoma. The findings of
the present study showed that TRIM24 promoted GC cell migration and
invasion, and was associated with translocation of β-catenin to the
nucleus. This functional analysis suggested that TRIM24 was a
crucial factor involved in multiple aspects of the progression of
GC.
A previous study reported that TRIM24 may serve as a
predictor of survival rates in patients with GC, and that TRIM24
enhances cell proliferation and chemoresistance via Akt signaling
activation (43). However, the
present study confirmed the critical role of TRIM24 in cell cycle
progression, apoptosis and invasion. In addition, the present study
was the first, to the best of our knowledge to demonstrate that
TRIM24 activated Wnt/β-catenin signaling, which also provided novel
support for activation of the Wnt/β-catenin pathway in GC. However,
the precise mechanism between TRIM24 and β-catenin remains to be
elucidated. Ubiquitination, microRNA dysregulation and DNA
methylation all contribute to dysregulation of the Wnt/β-catenin
pathway in several tumor types (44–46).
Therefore, additional studies are required to elucidate the
mechanistic basis of how TRIM24 interacts with β-catenin in GC.
In conclusion, the present study demonstrated that
TRIM24 was upregulated in human GC cells and tissues, and its
overexpression was closely associated with increased malignancy and
poor prognosis. Furthermore, the functional and mechanistic
investigations of TRIM24 performed in the present study indicated
that TRIM24 may be important in the control of GC aggressiveness,
partly by activating the Wnt/β-catenin pathway. These findings
identified TRIM24 as a potential therapeutic biomarker for patients
with GC and demonstrated its importance in activation of the
Wnt/β-catenin pathway during the progression of GC.
Acknowledgements
The study was supported by the JiangXi Province
Talent 555 Project, the National Natural Science Foundation of
China (grant nos. 81160281 and 81441083), the Major Natural Science
Foundation of Jiangxi Province (grant no. 20152ACB20024) and the
Science and Technology Project of Jiangxi Province (grant no.
20151BBG70228).
References
|
1
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Clevers H and Nusse R: Wnt/beta-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Behrens J, Jerchow BA, Würtele M, Grimm J,
Asbrand C, Wirtz R, Kühl M, Wedlich D and Birchmeier W: Functional
interaction of an axin homolog, conductin, with beta-catenin, APC,
and GSK3beta. Science. 280:596–599. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ikeda S, Kishida S, Yamamoto H, Murai H,
Koyama S and Kikuchi A: Axin, a negative regulator of the Wnt
signaling pathway, forms a complex with GSK-3beta and beta-catenin
and promotes GSK-3beta-dependent phosphorylation of beta-catenin.
EMBO J. 17:1371–1384. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ogasawara N, Tsukamoto T, Mizoshita T,
Inada K, Cao X, Takenaka Y, Joh T and Tatematsu M: Mutations and
nuclear accumulation of beta-catenin correlate with intestinal
phenotypic expression in human gastric cancer. Histopathology.
49:612–621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Klaus A and Birchmeier W: Wnt signaling
and its impact on development and cancer. Nat Rev Cancer.
8:387–398. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Clevers H: Wnt/β-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu W, Dong X, Mai M, Seelan RS, Taniquchi
K, Krishnadath KK, Halling KC, Cunningham JM, Boardman LA, Qian C,
Christensen E, Schmidt SS, Roche PC, Smith DI and Thibodeau SN:
Mutations in AXIN2 cause colorectal cancer with defective mismatch
repair by activating beta-catenin/TCF signaling. Nat Genet.
26:5012000. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hatakeyama S: TRIM proteins and cancer.
Nat Rev Cancer. 11:792–804. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Herquel B, Ouararhni K and Davidson I: The
TIF1α-related TRIM cofactors couple chromatin modifications to
transcriptional regulation, signaling and tumor suppression.
Transcription. 2:231–236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Allton K, Jain AK, Herz HM, Tsai WW, Jung
SY, Qin J, Bergmann A, Johnson RL and Barton MC: Trim24 targets
endogenous p53 for degradation. Proc Natl Acad Sci USA.
106:11612–11616. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jain AK and Barton MC: Regulation of p53:
TRIM24 enters the RING. Cell Cycle. 8:3668–3674. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsai WW, Wang Z, Yiu TT, Tsai WW, Jung SY,
Qin J, Bergmann A, Johnson RL, Barton MC, Aronow B, et al: TRIM24
links a non-canonical histone signature to breast cancer. Nature.
468:927–932. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pathiraja TN, Thakkar KN, Jiang S,
Stratton S, Liu Z, Gagea M, Shi X, Shah PK, Phan L, Lee MH, et al:
TRIM24 links glucose metabolism with transformation of human
mammary epithelial cells. Oncogene. 34:2836–2845. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhong S, Delva L, Rachez C, Cenciarelli C,
Gandini D, Zhang H, Kalantry S, Freedman LP, Pandolfi PP, et al: A
RA-dependent, tumour-growth suppressive transcription complex is
the target of the PML-RARalpha and T18 oncoproteins. Nat Genet.
23:287–295. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Belloni E, Trubia M, Gasparini P, Micucci
C, Tapinassi C, Confalonieri S, Nuciforo P, Martino B, Lo-Coco F,
Pelicci PG and Di Fiore PP: 8p11 myeloproliferative syndrome with a
novel t(7;8) translocation leading to fusion of the FGFR1 and TIF1
genes. Genes Chromosomes Cancer. 42:320–325. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Klugbauer S and Rabes HM: The
transcription coactivator HTIF1 and a related protein are fused to
the RET receptor tyrosine kinase in childhood papillary thyroid
carcinomas. Oncogene. 18:4388–4393. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang L, Heidt DG, Lee CJ, Yang H, Logsdon
CD, Zhang L, Fearon ER, Ljungman M and Simeone DM: Oncogenic
function of ATDC in pancreatic cancer through Wnt pathway
activation and beta-catenin stabilization. Cancer Cell. 15:207–219.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu X, Huang Y, Yang D, Li X, Liang J, Lin
L, Zhang M, Zhong K, Liang B and Li J: Overexpression of TRIM24 is
associated with the onset and progress of human hepatocellular
carcinoma. PloS One. 9:e854622014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Edge SB, Byrd DR, Compton CC, Fritz AG,
Greene FL and Trotti A: AJCC Cancer Staging Manual. Springer; New
York, NY: 2010
|
|
21
|
Bosman FT, Carneiro F, Hruban RH and
Theise ND: WHO Classification of Tumours of the Digestive System.
4th. IARC Press; Lyon: 2010
|
|
22
|
Pan X, Zhou T, Tai YH, Wang C, Zhao J, Cao
Y, Chen Y, Zhang PJ, Yu M, Zhen C, et al: Elevated expression of
CUEDC2 protein confers endocrine resistance in breast cancer. Nat
Med. 17:708–714. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu WK, Cho CH, Lee CW, Fan D, Wu K, Yu J
and Sung JJ: Dysregulation of cellular signaling in gastric cancer.
Cancer Lett. 295:144–153. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chambon M, Orsetti B, Berthe ML,
Bascoul-Mollevi C, Rodriguez C, Duong V, Gleizes M, Thénot S,
Bibeau F, Theillet C and Cavaillès V: Prognostic significance of
TRIM24/TIF-1α gene expression in breast cancer. Am J Pathol.
178:1461–1469. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cui Z, Cao W, Li J, Song X, Mao L and Chen
W: TRIM24 overexpression is common in locally advanced head and
neck squamous cell carcinoma and correlates with aggressive
malignant phenotypes. PloS One. 8:e638872013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang LH, Yin AA, Cheng JX, Huang HY, Li
XM, Zhang YQ, Han N and Zhang X: TRIM24 promotes glioma progression
and enhances chemoresistance through activation of the PI3K/Akt
signaling pathway. Oncogene. 34:600–610. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li H, Sun L, Tang Z, Fu L, Xu Y, Li Z, Luo
W, Qiu X and Wang E: Overexpression of TRIM24 correlates with tumor
progression in non-small cell lung cancer. PloS One. 7:e376572012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xue D, Zhang X, Liu J, Liu J, Li N, Liu C,
Liu Y and Wang P: Clinical significance and biological roles of
TRIM24 in human bladder carcinoma. Tumour Biol. 36:6849–6855. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang J, Zhu J, Dong M, Yu H, Dai X and Li
K: Knockdown of tripartite motif containing 24 by lentivirus
suppresses cell growth and induces apoptosis in human colorectal
cancer cells. Oncol Res. 22:39–45. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jain AK, Allton K, Duncan AD and Barton
MC: TRIM24 is a p53-induced E3-ubiquitin ligase that undergoes
ATM-mediated phosphorylation and autodegradation during DNA damage.
Mol Cell Biol. 34:2695–2709. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Knudsen KE, Diehl JA, Haiman CA and
Knudsen ES: Cyclin D1: Polymorphism, aberrant splicing and cancer
risk. Oncogene. 25:1620–1628. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Roy PG and Thompson AM: Cyclin D1 and
breast cancer. Breast. 15:718–727. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kishimoto I, Mitomi H, Ohkura Y, Kanazawa
H, Fukui N and Watanabe M: Abnormal expression of p16 (INK4a),
cyclin D1, cyclin-dependent kinase 4 and retinoblastoma protein in
gastric carcinomas. J Surg Oncol. 98:60–66. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Niu Z, Liu H, Zhou M, Wang H, Liu Y, Li X,
Xiong W, Ma J, Li X and Li G: Knockdown of c-Myc inhibits cell
proliferation by negatively regulating the Cdk/Rb/E2F pathway in
nasopharyngeal carcinoma cells. Acta Biochim Biophys Sin
(Shanghai). 47:183–191. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Howard S, Deroo T, Fujita Y and Itasaki N:
A positive role of cadherin in Wnt/β-catenin signalling during
epithelial-mesenchymal transition. PloS One. 6:e238992011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huber MA, Kraut N and Beug H: Molecular
requirements for epithelial-mesenchymal transition during tumor
progression. Curr Opin Cell Biol. 17:548–558. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Polette M, Mestdagt M, Bindels S,
Nawrocki-Raby B, Hunziker W, Foidart JM, Birembaut P and Gilles C:
Beta-catenin and ZO-1: Shuttle molecules involved in tumor
invasion-associated epithelial-mesenchymal transition processes.
Cells Tissues Organs. 185:61–65. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Valenta T, Hausmann G and Basler K: The
many faces and functions of β-catenin. EMBO J. 31:2714–2736. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Miyazawa K, Iwaya K, Kuroda M, Harada M,
Serizawa H, Koyanagi Y, Sato Y, Mizokami Y, Matsuoka T and Mukai K:
Nuclear accumulation of beta-catenin in intestinal-type gastric
carcinoma: Correlation with early tumor invasion. Virchows Arch.
437:508–513. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chiu CG, Chan SK, Fang ZA, Masoudi H,
Wood-Baker R, Jones SJ, Gilks B, Laskin J and Wiseman SM:
Beta-catenin expression is prognostic of improved non-small cell
lung cancer survival. Am J Surg. 203:654–659. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Miao ZF, Wang ZN, Zhao TT, Xu YY, Wu JH,
Liu XY, Xu H, You Y and Xu HM: TRIM24 is upregulated in human
gastric cancer and promotes gastric cancer cell growth and
chemoresistance. Virchows Arch. 466:525–532. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
MacDonald BT, Tamai K and He X:
Wnt/beta-catenin signaling: Components, mechanisms and diseases.
Dev Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ying Y and Tao Q: Epigenetic disruption of
the WNT/beta-catenin signaling pathway in human cancers.
Epigenetics. 4:307–312. 2009. View Article : Google Scholar
|
|
46
|
Liu Y, Huang T, Zhao X and Cheng L:
MicroRNAs modulate the Wnt signaling pathway through targeting its
inhibitors. Biochem Biophys Res Commun. 408:259–264. 2011.
View Article : Google Scholar : PubMed/NCBI
|