Introduction
Breast cancer (BC), is a type of malignancy that
poses the greatest threat to female health worldwide, despite its
relatively long 5-year survival rate (~58.75% between 1972 and 2011
in China) (1). BC is one of the most
common types of cancer in Asia, where ~39% of all worldwide cases
are diagnosed (2). The treatment and
care for patients with BC is a large economic burden in developed
and developing countries (3–5). Considering the health and economic
effects of BC, numerous studies have been conducted to investigate
the pathogenesis, and to identify therapeutic targets and potential
factors that may contribute its progression. However, the
mechanisms behind the development and progression of BC remain
poorly understood, and additional studies are required.
G9A [also known as euchromatic histone lysine
methyltransferase 2 (EHMT2)], encodes a methyltransferase
that methylates lysine residues of histone H3. The dysregulation of
G9A is associated with numerous abnormal biological
processes and the emergence of diseases, including cancer. For
example, the downregulation of G9A could trigger DNA damage
responses and inhibit the progression of colorectal cancer
(6). In head and neck squamous cell
carcinoma, G9A dysregulation has been associated with
epithelial-mesenchymal transition (EMT)-mediated metastasis and the
maintenance of cancer stem cell-like characteristics (7). Furthermore, it has been identified that
G9A performs important roles in the transition of
acute-to-chronic pain (8). In BC, the
aberrant expression of G9A could affect the stability of
SRY-box 2 (SOX2), a tumor suppressor gene, by inducing its
epigenetic silencing (9).
Furthermore, a study by Si et al (10) revealed that G9A performed
important roles in the dysfunction of the reciprocal feedback loop
between GATA binding protein 3 and zinc finger E-box binding
homeobox 2, and that it contributed to the progression of BC. The
combined analysis of its genome-wide profiles and the gene
expression data following G9A-knockdown (KO) or
overexpression in BC could improve our understanding of its
regulatory functions, and has not previously been investigated.
MicroRNAs (miRNAs/miRs) are small non-coding RNA
molecules, 22–25 nucleotides in length. miRNAs are formed from the
precursor miRNA, a double-stranded hairpin RNA or even the
intergenic region, and perform important roles in the regulation of
gene expression at the post translation level by silencing mRNA
translation or inducing protein degradation (11). Numerous miRNAs have been shown to
contribute to the progression of a number of cancer types. For
example, the downregulation of miR-711 could inhibit the
cellular proliferation of BC, and miR-711 has been shown to
be an independent prognostic factor for BC (12). miR-27a/b and
miR-494 may regulate the expression of tissue factor pathway
inhibitor α (TFPIα) in its estrogen-mediated downregulation
in BC (13).
In the present study, through the combined analysis
of genome-wide profiles of G9A obtained through chromatin
immunoprecipitation-DNA sequencing (ChIP-Seq) and the gene
expression dataset [from the Gene Expression Omnibus (GEO)]
following G9A-KO in BC, potential biomarkers were identified
for the treatment of BC. miRNAs that may regulate those biomarkers
were screened out and the core miRNAs and genes were obtained. The
findings of the present study may be valuable for the diagnosis and
treatment of BC.
Materials and methods
ChIP-Seq and gene expression
dataset
In the present study, the ChIP-Seq and gene
expression datasets were comprehensively obtained from the GEO
database (http://www.ncbi.nlm.nih.gov/geo). The ChIP-Seq dataset
(GSM1642516) was deposited from the study by Si et al
(10), which used the G9A-specific
antibody to capture the DNA fragments in MCF-7 cells and sequenced
those fragments based on the GPL11154 Illumina HiSeq 2000 (Homo
sapiens). Furthermore, an input control sample, which extracted
all DNA in the human breast adenocarcinoma MCF-7 cell line, was
adopted for the background correction in the identification of
G9A binding sites (peaks). The gene expression dataset
GSE34925 (14) contained 3
G9A-KO and 3 control BC samples, which were based on GPL6244
(HuGene-1_0-st) Affymetrix Human Gene 1.0 ST Array [transcript
(gene) version] (Affymetrix, Inc., Santa Clara, CA, USA).
Differential expression analysis
The raw CEL data (data storage format) were imported
to R based on the affy package (15).
Background correction and expression normalization was conducted
using the oligo package (16). Probe
IDs were converted to gene symbols via the annotation package of
the microarray platform and the expression values were summarized
that corresponding to multiple probe sets. A paired Student's
t-test was conducted between logarithmic transformed expression
values of G9A-KO samples and control samples.
Differentially-expressed genes (DEGs) with a fold-change expression
value of >1.41 or <0.71, and a false discovery rate corrected
P-value of <0.05 were screened out.
Functional enrichment analysis
The Database for Annotation, Visualization and
Integrated Discovery (https://david.ncifcrf.gov/) (17) was used for the functional analysis of
the DEGs. Gene Ontology (GO) terms and Kyoto Encyclopedia of Genes
and Genomes (KEGG; http://www.kegg.jp/) pathways that satisfied the
threshold of P<0.05 were screened out.
Genome-wide binding analysis of
G9A
Bowtie 2 (18), an
ultrafast genome aligning tool, was used for the mapping of the raw
reads to UCSC hg19 genome with a maximum of 2 mismatches in every
read. Model-based analysis of ChIP-Seq version 14 (macs14)
(19) was used for the identification
of the binding sites of G9A (also known as call peak) with a
P-value of <1×10−5. Peaks located in −3000 to 3000 bp
centered in the transcription start sites (TSS) were considered to
be conpeaks. Based on the ChIPseeker package (20), the nearby genes, genome features
(including promoter and 5′-untranslated region) and distances to
the nearest TSS were assigned to the peaks.
Construction of miRNA-gene
network
The overlapping genes between DEGs and genes located
in the conpeaks were screened out. miRNAs that may regulate those
overlaps were identified using the TargetScan database (http://www.targetscan.org/) (21). The miRNA-gene regulation network was
visualized through Cytoscape (http://www.cytoscape.org/).
Results
DEGs
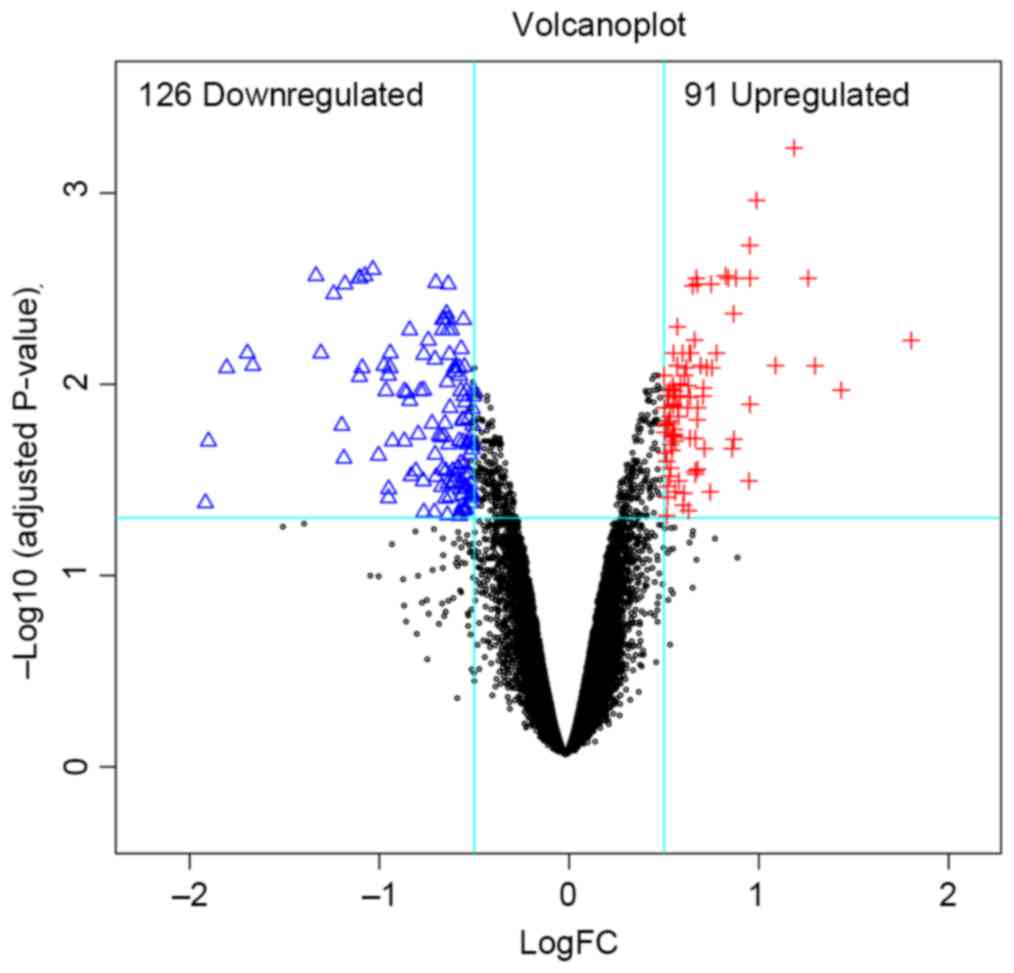
A total of 217 DEGs were identified in the G9A-KO BC
samples, and the distribution of these DEGs is demonstrated in
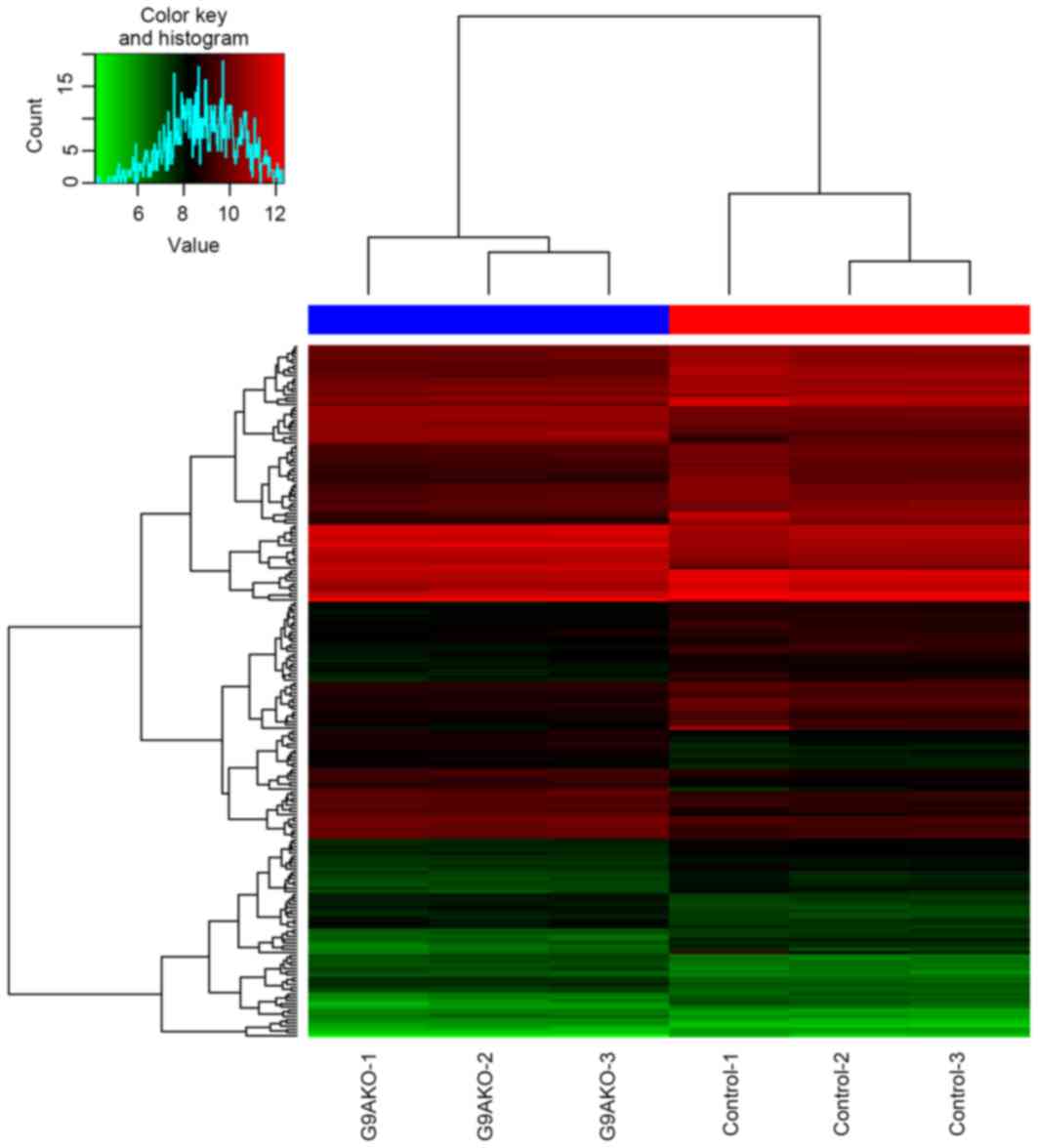
Fig. 1. The supervised clustering
(Fig. 2) based on those DEGs was able
to separate the G9A-KO and control samples well.
Enriched functions
A total of 172 GO terms and 3 KEGG pathways were
identified to be enriched in DEGs, and were mainly involved in the
cell process, the inflammatory response and cancer progression, for
example, the extracellular region part. The top 10 GO terms
according to P-value and KEGG pathways are presented in Tables I and II, respectively.
 | Table I.Top 10 GO terms according to the
P-value. |
Table I.
Top 10 GO terms according to the
P-value.
| Category | GO term | Count | P-value |
|---|
| CC | Extracellular
space | 27 |
5.22×10−8 |
| CC | Extracellular
region part | 32 |
9.50×10−8 |
| BP | Organ
development | 44 |
1.04×10−6 |
| BP | Anatomical
structure development | 53 |
1.44×10−5 |
| BP | System
development | 50 |
1.59×10−5 |
| BP | Integrin-mediated
signaling pathway | 8 |
1.70×10−5 |
| CC | Extracellular
region | 44 |
1.95×10−5 |
| BP | Inflammatory
response | 15 |
2.81×10−5 |
| BP | Blood vessel
development | 13 |
3.09×10−5 |
| BP | Defense
response | 21 |
3.32×10−5 |
| Table II.Enriched Kyoto Encyclopedia of Genes
and Genomes pathways. |
Table II.
Enriched Kyoto Encyclopedia of Genes
and Genomes pathways.
| Pathway name | Count | P-value | Genes |
|---|
| Cytokine-cytokine
receptor interaction | 13 |
2.99×10−4 | CSF2, TNFSF4,
IL24, IL7R, CXCL11, TNFSF18, CXCL10, KDR, INHBB, VEGFC, CXCL16,
IL1A, GHR |
| Focal adhesion | 11 |
5.24×10−4 | VEGFC, ITGB8,
LAMA5, ITGB4, ITGA2, RELN, BIRC3, VAV1, MYLK, KDR, SPP1 |
| ECM-receptor
interaction | 6 |
6.80×10−3 | ITGB8, LAMA5,
ITGB4, ITGA2, RELN, SPP1 |
Genome-wide binding profile of
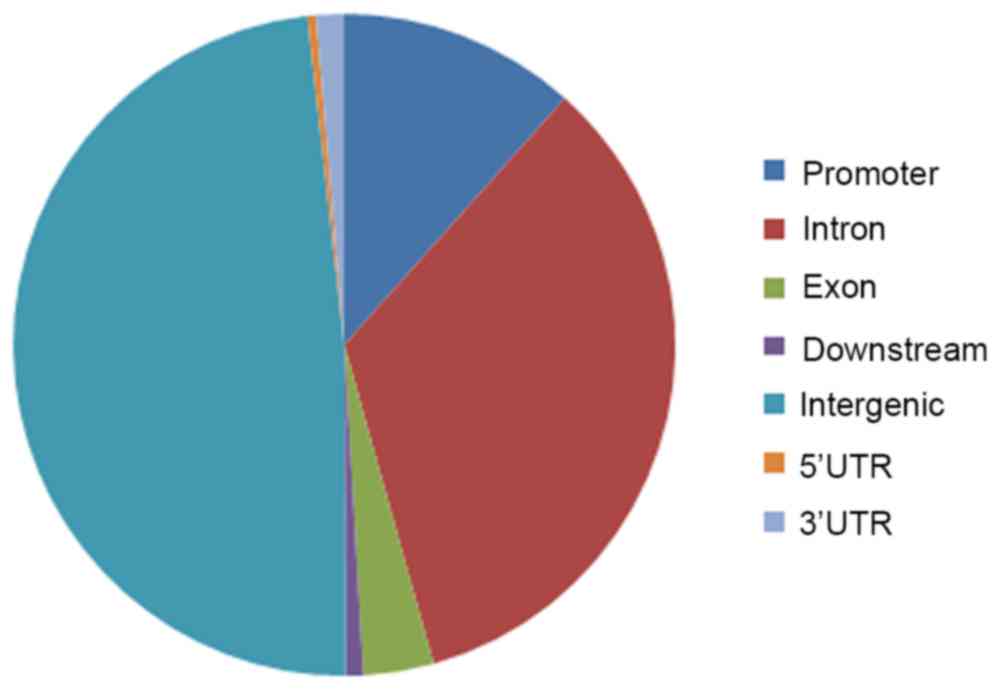
G9A
A total of 10,422 binding sites (peaks) of G9A were
identified, including 1,210 conpeaks located in −3000 to 3000 bp
centered in the TSS. The distribution of all peaks across the
genome is shown in Fig. 3. In
addition, 1,138 genes were identified to be located in the 1,210
conpeaks and 15 of these genes overlapped with DEGs (Table III).
| Table III.Overlapping genes between
differentially-expressed genes and genes located in conpeaks. |
Table III.
Overlapping genes between
differentially-expressed genes and genes located in conpeaks.
| Gene | logFC |
|---|
| SLCO4C1 | −0.65617 |
|
ARHGAP24 | −0.66706 |
| ANK2 | −0.55143 |
| TMX4 | 0.54535 |
| LARP4 | −0.50197 |
| TC2N | −0.78340 |
| TMTC1 | 0.52521 |
| SLFN5 | −0.62773 |
| CYTIP | 0.67537 |
| PLCB4 | −0.55811 |
| ATF7IP2 | −1.19761 |
| CAMK2D | −0.50487 |
| TLR4 | 0.85905 |
| INSIG1 | −1.18712 |
| GHR | −0.60367 |
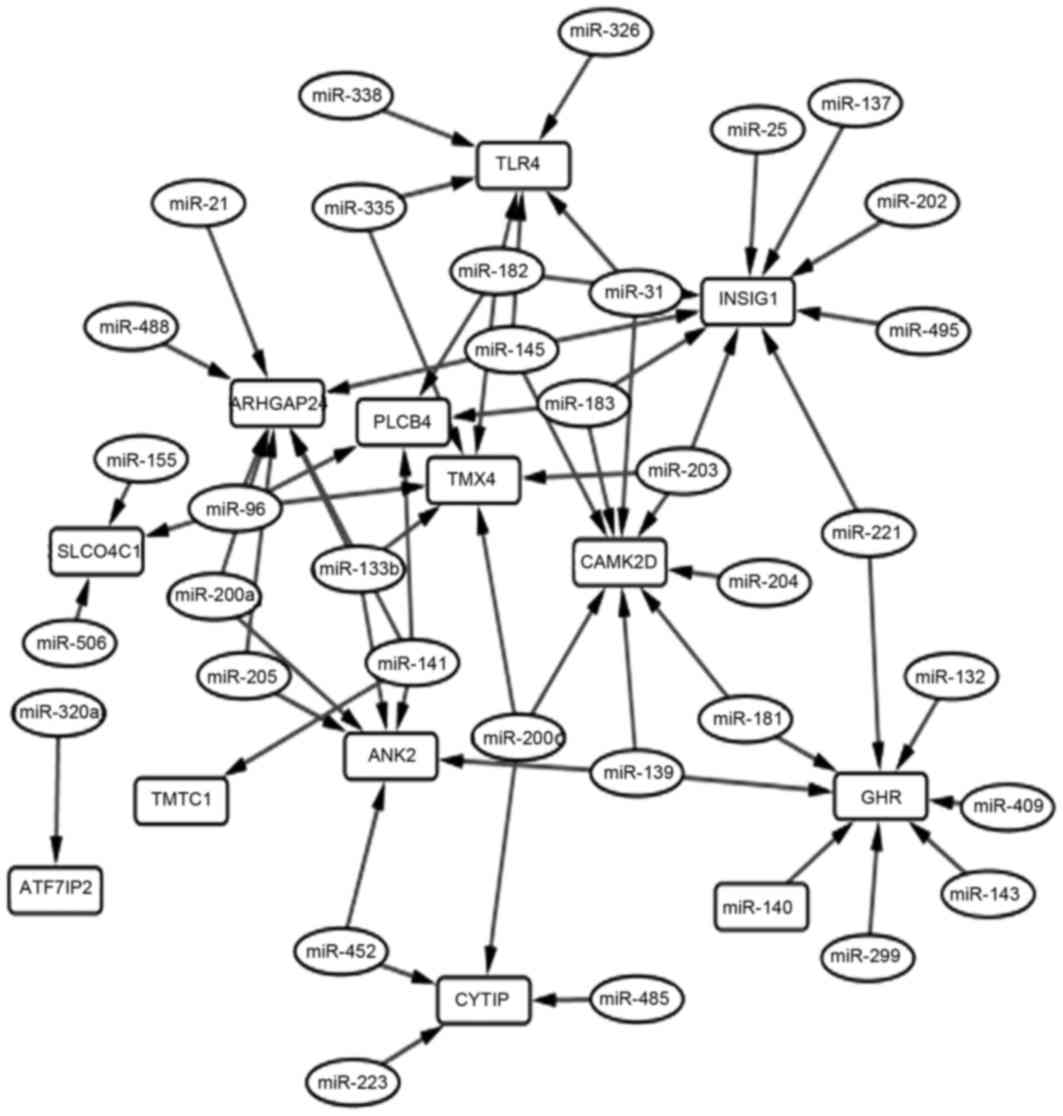
miRNA-gene regulation network
Based on TargetScan, a total of 35 miRNAs were
identified to regulate the 15 overlapping genes, and 65 miRNA-gene
pairs were obtained among them. The miRNA-gene regulation network
is shown in Fig. 4. The top 10 genes
according to their connectivity in the miRNA-gene network are
listed in Table IV. In the network,
insulin-induced gene 1 (INSIG1) was regulated by 9 miRNAs and had
the highest connectivity, which indicated that it was a hub gene.
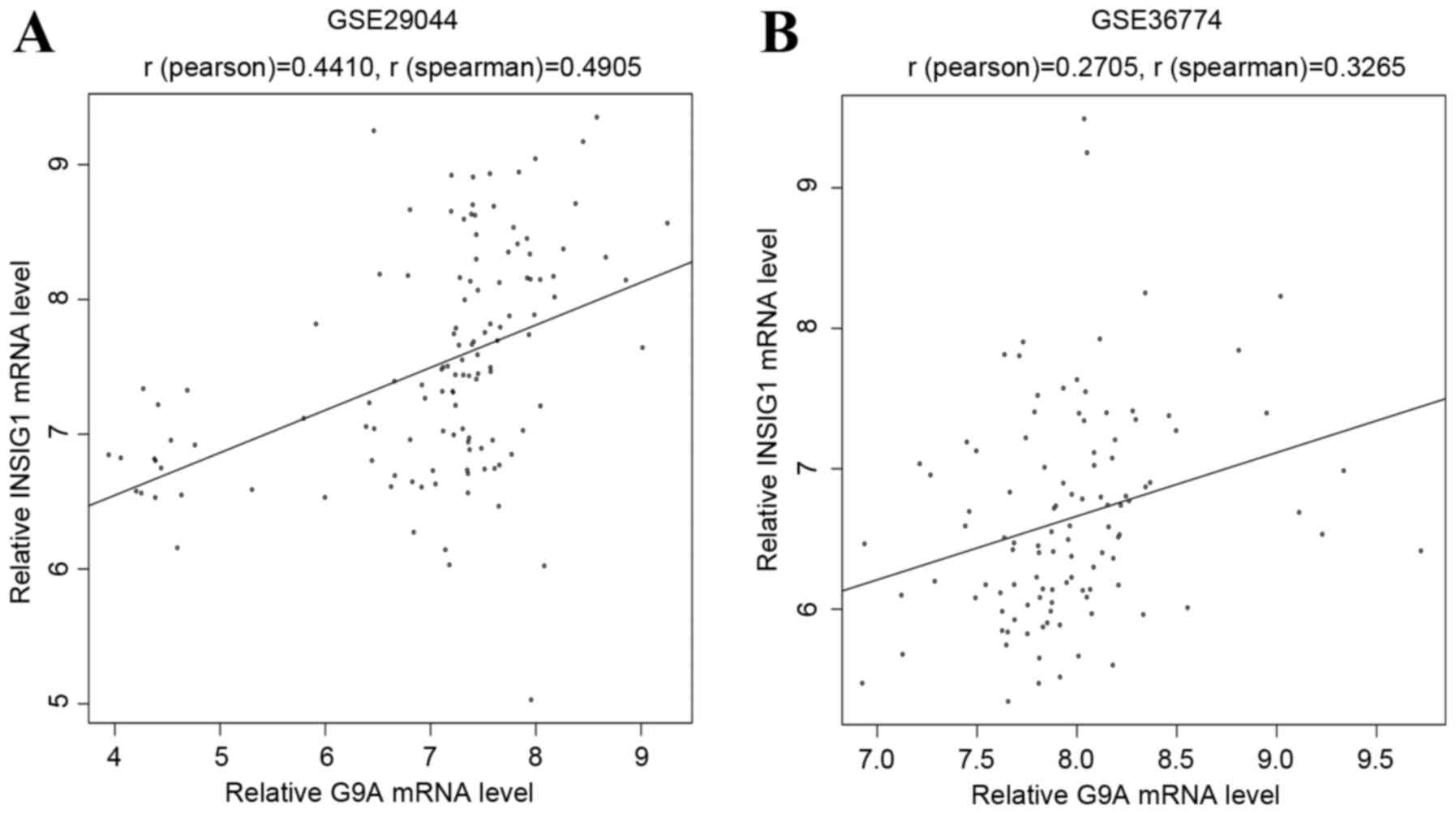
Furthermore, two other public datasets (GSE29044 and GSE36774) were
downloaded from the GEO database, which including data from breast
cancer and normal tissue samples. These two datasets were
background corrected and quantile normalized using the
preprocessCore package of R (http://www.bioconductor.org/packages/release/bioc/html/preprocessCore.html).
The Pearson and Spearman's correlation indices were respectively
calculated using R between relative INSIG1 and G9A mRNA levels. The
results indicated a significantly positive correlation (Fig. 5) between G9A and INSIG1, which may
demonstrate their important roles in the progression of BC. The top
10 genes according to their connectivity in miRNA-gene network are
presented in Fig. 4.
| Table IV.Top 10 genes according to their
connectivity in the microRNA-gene network. |
Table IV.
Top 10 genes according to their
connectivity in the microRNA-gene network.
| Gene | Connectivity |
|---|
| INSIG1 | 9 |
|
ARHGAP24 | 8 |
| CAMK2D | 8 |
| GHR | 8 |
| ANK2 | 7 |
| TLR4 | 6 |
| TMX4 | 6 |
| CYTIP | 4 |
| miR-141 | 4 |
| miR-145 | 4 |
Discussion
BC is a complicated disease, and numerous factors
may contribute to its initiation and progression, including
occupational exposure (22),
variations in expression level or mutations of specific genes
(23,24). The traditional treatment options for
BC mainly involve surgical resection, chemotherapy and
radiotherapy, which have been shown to be effective but rarely
eradicate the disease. With the development of next-generation
sequencing technology, numerous biomarkers of BC have been
identified and certain novel therapeutics have been proposed
(25). Immunotherapy is one of the
most popular therapeutic treatment options (26,27). The
identification of novel biomarkers may therefore improve the
prognosis for patients with BC.
In the present study, the genome-wide profiles of
G9A in genome wide were identified using ChIP-Seq. DEGs in
G9A-KO BC samples compared with the control samples were
screened out. The combined analysis obtained potential direct
targets of G9A, the majority of which were downregulated in
the G9A-KO BC samples, which may indicate its upregulated
functions in BC.
DEGs in G9A-KO samples are primarily involved
in biological processes or pathways associated with cellular
processes, inflammatory responses and cancer progression, including
extracellular region part, cytokine-cytokine receptor interaction
and extracellular membrane (ECM) receptor interaction, all of which
are closely associated with the development of BC. In particular,
inflammatory triple-negative BC, a type of rare and aggressive BC
with a significantly poorer 5-year survival rate compared with
other types of BC, is believed to mainly be attributed to an
abnormal inflammatory response (28,29).
Previous studies have considered that the ECM-receptor interaction
pathway may be associated with the progression of BC, which could
indicate the repeatability of the present study (30,31).
Among the 1,138 genes distributed in the 1,210 peaks
located between −3000 to 3000 bp centered in the TSS, 15 were
identified to overlapped with the DEGs in the G9A-KO BC
samples. Furthermore, 11 of those overlaps were revealed to be
downregulated in G9A-KO samples compared with the control
samples, which may indicate that G9A is mainly involved in
the upregulation of its target genes. The positive correlation
between the expression values of G9A and INSIG1 was
verified using two public datasets (Fig.
5). Furthermore, INSIG1 possessed an intimate
connectivity pattern in the miRNA-gene regulation network (Fig. 4). Thus, this suggests that G9A
and INSIG1 serve important roles in the progression of BC.
INSIG1, which is downregulated in G9A-KO BC, was
closely associated with the progression of several types of cancer
via the regulation of glucose metabolism and hypoxia-induced EMT
(32,33). BC had a strong association with
insulin resistance, which was mediated by INSIG1 (34). In previous studies, INSIG1 was
also demonstrated to be differentially expressed in BC (35,36).
Combined with the results of the present study, this indicates the
important roles of INSIG1 in BC.
miR-203 regulated 3 genes in the miRNA-gene
regulation network, including INSIG1, which has been
reported as a tumor suppressor in numerous types of cancer,
including BC (37–39). The activity of miR-203 was also
affected by a number of other factors, including kallistatin
(40). Therefore, it would be useful
to study the upstream and downstream regulation loops of
miR-203 for the diagnosis and treatment of BC, as well as
other types of cancer. Other nodes (miRNAs or genes) with high
connectivity in the miRNA-gene regulation network may also be
potential biomarkers for BC, which should be additionally verified
through molecular biological experiments.
In conclusion, the present study conducted the
combined analysis of ChIP-Seq and gene expression profiles in BC,
and identified potential biomarkers via screening. The present
results may aid the development of novel diagnostic or treatment
methods, including immunotherapy, and therefore improve the
prognosis of BC.
Acknowledgements
The present study was supported by the Municipal
Science and Technology Commission of Tianjin (grant nos.
15ZLZLZF00440 and 16ZLZXZF00120) and the Health Bureau Science and
Technology Foundation of Tianjin (grant no. 2014KZ102).
References
|
1
|
Zhu J, Chen JG, Chen YS, Zhang YH, Ding LL
and Chen TY: Female breast cancer survival in Qidong, China,
1972–2011: A population-based study. BMC Cancer. 14:3182014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fan L, Goss PE and Strasser-Weippl K:
Current status and future projections of breast cancer in Asia.
Breast Care (Basel). 10:372–378. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Onega T, Tosteson AN, Weiss J,
Alford-Teaster J, Hubbard RA, Henderson LM, Kerlikowske K, Goodrich
ME, O'Donoghue C, Wernli KJ, et al: Costs of diagnostic and
preoperative workup with and without breast MRI in older women with
a breast cancer diagnosis. BMC Health Serv Res. 16:762016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dare AJ, Anderson BO, Sullivan R, Pramesh
CS, Andre I, Adewole IF, Badwe RA and Gauvreau CL: Surgical
services for cancer care. 2015.
|
|
5
|
Daroudi R, Sari A Akbari, Nahvijou A,
Kalaghchi B, Najafi M and Zendehdel K: The economic burden of
breast cancer in Iran. Iran J Public Health. 44:1225–1233.
2015.PubMed/NCBI
|
|
6
|
Zhang J, He P, Xi Y, Geng M, Chen Y and
Ding J: Down-regulation of G9a triggers DNA damage response and
inhibits colorectal cancer cells proliferation. Oncotarget.
6:2917–2927. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu S, Ye D, Guo W, Yu W, He Y, Hu J, Wang
Y, Zhang L, Liao Y, Song H, et al: G9a is essential for
EMT-mediated metastasis and maintenance of cancer stem cell-like
characters in head and neck squamous cell carcinoma. Oncotarget.
6:6887–6901. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Laumet G, Garriga J, Chen SR, Zhang Y, Li
DP, Smith TM, Dong Y, Jelinek J, Cesaroni M, Issa JP and Pan HL:
G9a is essential for epigenetic silencing of K(+) channel genes in
acute-to-chronic pain transition. Nat Neurosci. 18:1746–1755. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee JY, Lee SH, Heo SH, Kim KS, Kim C, Kim
DK, Ko JJ and Park KS: Novel function of lysine methyltransferase
G9a in the regulation of Sox2 protein stability. PLoS One.
10:e01411182015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Si W, Huang W, Zheng Y, Yang Y, Liu X,
Shan L, Zhou X, Wang Y, Su D, Gao J, et al: Dysfunction of the
reciprocal feedback loop between GATA3- and ZEB2-nucleated
repression programs contributes to breast cancer metastasis. Cancer
Cell. 27:822–836. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu X, Liao S, Xu Z, Zhu L, Yang F and Guo
W: Identification and analysis of the porcine MicroRNA in porcine
cytomegalovirus-infected macrophages using deep sequencing. PLoS
One. 11:e01509712016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hu JY, Yi W, Zhang MY, Xu R, Zeng LS, Long
XR, Zhou XM, Zheng XS, Kang Y and Wang HY: MicroRNA-711 is a
prognostic factor for poor overall survival and has an oncogenic
role in breast cancer. Oncol Lett. 11:2155–2163. 2016.PubMed/NCBI
|
|
13
|
Ali HO, Arroyo AB, González-Conejero R,
Stavik B, Iversen N, Sandset PM, Martínez C and Skretting G: The
role of microRNA −27a/b and microRNA-494 in oestrogen mediated
downregulation of tissue factor pathway inhibitor α. J Thromb
Haemost. 14:1226–1237. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dong C, Wu Y, Yao J, Wang Y, Yu Y,
Rychahou PG, Evers BM and Zhou BP: G9a interacts with Snail and is
critical for Snail-mediated E-cadherin repression in human breast
cancer. J Clin Invest. 122:1469–1486. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: affy-analysis of Affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Stuhlmüller B: A composition of nucleic
acid sequences, specific for inflammatory disease, in particular
rheumatoid arthritis. EP 1795610 A1. December 6. 2005 June
13–2007.
|
|
17
|
Dennis G Jr, Sherman BT, Hosack DA, Yang
J, Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization, and integrated discovery. Genome Biol. 4:P32003.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Langmead B and Salzberg SL: Fast
gapped-read alignment with Bowtie 2. Nat Methods. 9:357–359. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Feng J, Liu T, Qin B, Zhang Y and Liu XS:
Identifying ChIP-seq enrichment using MACS. Nat Protoc.
7:1728–1740. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu G, Wang LG and He QY: ChIPseeker: An
R/Bioconductor package for ChIP peak annotation, comparison and
visualization. Bioinformatics. 31:2382–2383. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Song X, Cheng L, Zhou T, Guo X, Zhang X,
Chen YP, Han P and Sha J: Predicting miRNA-mediated gene silencing
mode based on miRNA-target duplex features. Comput Biol Med.
42:1–7. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fenga C: Occupational exposure and risk of
breast cancer. Biomed Rep. 4:282–292. 2016.PubMed/NCBI
|
|
23
|
Inoue K and Fry EA: Novel molecular
markers for breast cancer. Biomark Cancer. 8:25–42. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rizk SM, Shahin NN and Shaker OG:
Association between SIRT1 gene polymorphisms and breast cancer in
Egyptians. PLoS One. 11:e01519012016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shen Y and Cai T: Identifying predictive
markers for personalized treatment selection. Biometrics.
72:1017–1025. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Becht E, de Reyniès A, Giraldo NA, Pilati
C, Buttard B, Lacroix L, Selves J, Sautès-Fridman C, Laurent-Puig
Pa and Fridman WH: Immune and stromal classification of colorectal
cancer is associated with molecular subtypes and relevant for
precision immunotherapy. Clin Cancer Res. 22:4057–4066. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Abou-Shousha S, Moaz M, Sheta M and
Motawea MA: An approach to breast cancer immunotherapy: The
apoptotic activity of recombinant anti-interleukin-6 monoclonal
antibodies in intact tumor microenvironment of breast carcinoma.
Scand J Immunol. 83:427–437. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Biswas T, Efird JT, Prasad S, James SE,
Walker PR and Zagar TM: Inflammatory TNBC breast cancer: Demography
and clinical outcome in a large cohort of patients with TNBC. Clin
Breast Cancer. 16:212–216. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Suárez-Arroyo IJ, Rios-Fuller TJ,
Feliz-Mosquea YR, Lacourt-Ventura M, Leal-Alviarez DJ,
Maldonado-Martinez G, Cubano LA and Martínez-Montemayor MM:
Ganoderma lucidum combined with the EGFR tyrosine kinase inhibitor,
erlotinib synergize to reduce inflammatory breast cancer
progression. J Cancer. 7:500–511. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang X, Jia M, Li Z, Lu S, Qi X, Zhao B,
Wang X, Rong Y, Shi J, Zhang Z, et al: Bioinformatics analysis of
aggressive behavior of breast cancer via an integrated gene
regulatory network. J Cancer Res Ther. 10:1013–1018. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sun Y, Yuan K, Zhang P, Ma R, Zhang QW and
Tian XS: Crosstalk analysis of pathways in breast cancer using a
network model based on overlapping differentially expressed genes.
Exp Ther Med. 10:743–748. 2015.PubMed/NCBI
|
|
32
|
Chen HF and Wu KJ: Epigenetics, TET
proteins, and hypoxia in epithelial-mesenchymal transition and
tumorigenesis. Biomedicine (Taipei). 6:12016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tsai YP, Chen HF, Chen SY, Cheng WC, Wang
HW, Shen ZJ, Song C, Teng SC, He C and Wu KJ: TET1 regulates
hypoxia-induced epithelial-mesenchymal transition by acting as a
co-activator. Genome Biol. 15:5132014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ghose A, Kundu R, Toumeh A, Hornbeck C and
Mohamed I: A review of obesity, insulin resistance, and the role of
exercise in breast cancer patients. Nutr Cancer. 67:197–202. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hernández-Vargas H, Rodriguez-Pinilla SM,
Julián-Tendero M, Sánchez-Rovira P, Cuevas C, Antón A, Ríos MJ,
Palacios J and Moreno-Bueno G: Gene expression profiling of breast
cancer cells in response to gemcitabine: NF-kappaB pathway
activation as a potential mechanism of resistance. Breast Cancer
Res Treat. 102:157–172. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Einbond LS, Su T, Wu HA, Friedman R, Wang
X, Jiang B, Hagan T, Kennelly EJ, Kronenberg F and Weinstein IB:
Gene expression analysis of the mechanisms whereby black cohosh
inhibits human breast cancer cell growth. Anticancer Res.
27:697–712. 2007.PubMed/NCBI
|
|
37
|
Shi Y, Tan YJ, Zeng DZ, Qian F and Yu PW:
miR-203 suppression in gastric carcinoma promotes Slug-mediated
cancer metastasis. Tumour Biol. Jul 21–2015.(Epub ahead of
print).
|
|
38
|
Taipaleenmäki H, Browne G, Akech J, Zustin
J, Van Wijnen AJ, Stein JL, Hesse E, Stein GS and Lian JB:
Targeting of Runx2 by miR-135 and miR-203 impairs progression of
breast cancer and metastatic bone disease. Cancer Res.
75:1433–1444. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhou Y, Hu HY, Meng W, Jiang L, Zhang X,
Sha JJ, Lu Z and Yao Y: MEK inhibitor effective against
proliferation in breast cancer cell. Tumour Biol. 35:9269–9279.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li P, Guo Y, Bledsoe G, Yang Z, Chao L and
Chao J: Kallistatin induces breast cancer cell apoptosis and
autophagy by modulating Wnt signaling and microRNA synthesis. Exp
Cell Res. 340:305–314. 2016. View Article : Google Scholar : PubMed/NCBI
|