Introduction
Breast cancer is the most common malignant tumor
among women and is a significant global health problem. It is
estimated that ~1.3 million women are diagnosed with breast cancer
and >450,000 breast cancer-associated mortalities are recorded
every year (1). Although the
morbidity of breast cancer and mortality rates have decreased, due
to prevention, screening, early detection and treatment, and the
survival time of breast cancer patients has been extended, there
are a significant amount of individuals diagnosed with breast
cancer, which transforms into malignant tumors (2,3).
MicroRNAs (miRs) are a large family of single
stranded non-coding RNAs composed of 22 nucleotides. miRs regulate
gene expression, which affects various biological behaviors,
including the formation, growth and development of embryos,
hematopoiesis, proliferation, apoptosis and differentiation of
cells, and occurrence and development of tumors (4). Increasing studies suggest that miRs are
important in the occurrence and development of tumors, and certain
miRs act as oncogenes or anti-oncogenes (5). One such miR is miR-21, which has been
demonstrated to be overexpressed in pulmonary, mammary gland and
prostate and colon malignant tumors, gastrointestinal, esophageal
and pancreatic cancer and head and neck neoplasms (6).
Curcumin is a polyphenol with a low molecular weight
and is extracted from Curcuma longa L, which belongs to the
Zingiberaceae family (7). Generally,
curcumin is regarded as the most effective constituent in turmeric
and accounts for 2–8% in the majority of turmeric preparations.
According to various studies, curcumin has numerous pharmacological
effects, including anti-oxidative (8), anti-inflammatory (9), anticancer (10), free radical clearance (1) and antimicrobial effects (2). In addition, curcumin has numerous
pharmacological functions on the cardiovascular and digestive
systems (11). The present study
investigated whether the anticancer effect of curcumin inhibits
breast cancer cell growth through the miR-21/phosphatase and tensin
homolog (PTEN)/protein kinase B (Akt) signaling pathway.
Materials and methods
Materials
Dulbecco's modified Eagle's medium (DMEM) and fetal
bovine serum (FBS) was obtained from HyClone™ (GE Healthcare,
Logan, UT, USA). Curcumin (purity >98%) was supplied by
Sigma-Aldrich (St. Louis, MO, USA). 3-(4,5-dimethyl-2-thiazolyl)-2,
5-diphenyl-2H-tetrazolium bromide (MTT) and lactate dehydrogenase
(LDH) were supplied by Tianjin Chemical Reagent No. 1 Plant
(Tianjin, China). Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) Apoptosis Detection kit was obtained
from Beijing Biosea Biotechnology, Co., Ltd. (Beijing, China). A
BCA Protein Assay kit was supplied by Wuhan Boster Bioengineering
Co., Ltd. (Wuhan, China). Caspase-3 (C1116) and caspase-9 (C1158)
activity kits were obtained from Beyotime Institute of
Biotechnology (Beijing, China). TRIzol reagent was supplied by
Invitrogen™ (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Primescript™ RT Master mix kit was obtained from Takara
Biotechnology Co., Ltd. (Dalian, China). ABI Prism 7900HT Real-Time
PCR system was supplied by Applied Biosystems® (Thermo
Fisher Scientific, Inc.).
Cell culture
Human breast cancer MCF-7 cell line was purchased
from Shanghai Institute of Cell Biology, Chinese Academy of
Sciences (Shanghai, China). The MCF-7 cells were cultured in DMEM
containing 10% FBS with 100 U/ml penicillin and 100 U/ml
streptomycin (Sigma-Aldrich) in a humidified atmosphere of 95% air
with 5% CO2 at 37°C.
MTT assay
MCF-7 cells were seeded at a density of
2×104 cells/well (0.2 ml/well) in 96-well plates
(Corning, Inc., Corning, NY, USA) for 24 h. Following exposure to
various concentrations of curcumin [0 (with DMSO vehicle), 0.5,
1.0, 2.0, 5.0 and 10.0 µM] for 24, 48 and 72 h, respectively
(12), 20 µl MTT solution (Tianjin
Chemical Reagent No. 1 Plant, Tianjin, China) was added to each
well and the cells were incubated for an additional 4 h. In total,
200 µl dimethyl sulfoxide (Sinopharm Chemical Reagent Co., Ltd.,
Shanghai, China) was added to each well and the plates were
agitated for 20 min at room temperature. The absorbance of the
samples was measured at 490 nm using a microplate reader (model
3550; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
LDH assay
MCF-7 cells were seeded in 96-well plates at a
density of 2×104 cells/well (0.2 ml/well) for 24 h.
Following exposure to various concentrations of curcumin [0 (with
DMSO vehicle), 0.5, 1.0, 2.0, 5.0 and 10.0 µM] for 24, 48 and 72 h,
100 µl LDH solution was added to each well and the plates was
incubated at room temperature for 30 min. The absorbance of the
samples was measured at 490 nm using a microplate reader (Bio-Rad
Laboratories, Inc.).
Cell apoptosis evaluation using flow
cytometry
MCF-7 cells were seeded in 6-well plates (Corning,
Inc.) at a density of 1×106 cells/well (2 ml/well) for
24 h. The cells were centrifuged at 2,000 × g for 10 min and
collected following treatment with curcumin at 0 (with DMSO
vehicle), 1, 2 and 5 µM for 48 h. The cells were then washed twice
with cold phosphate-buffered saline (PBS). The MCF-7 cells were
resuspended with 500 µl binding buffer (Beijing Biosea
Biotechnology, Co., Ltd., Beijing, China). Subsequently, miscible
liquids were mixed with 10 µl Annexin V for 30 min in the dark at
room temperature, followed by the addition of 5 µl PI (Beijing
Biosea Biotechnology, Co., Ltd.) for 30 min in the dark at room
temperature. Subsequently, the samples were analyzed by FACSAria
flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
4′,6-diamidino-2-phenylindole (DAPI)
staining assay
MCF-7 cells were seeded in 6-well plates at a
density of 1×106 cells/well (2 ml/well) for 24 h. The
cells were collected following treatment with curcumin at 0 (with
DMSO vehicle), 1, 2 and 5 µM for 48 h, and subsequently washed
twice with cold PBS. The MCF-7 cells were fixed in 4%
paraformaldehyde (Sinopharm Chemical Reagent Co., Ltd.),
permeabilized with 0.1% Triton X-100 (Sinopharm Chemical Reagent
Co., Ltd.) and stained with 2 µg/ml DAPI (Beyotime Institute of
Biotechnology) for 10 min. Nucleic morphology of the MCF-7 cells
was observed under a fluorescence microscope (Nikon Corporation,
Tokyo, Japan).
Caspase-3 and caspase-9
activities
MCF-7 cells were seeded in 6-well plates at a
density of 1×106 cells/well (2 ml/well) for 24 h. MCF-7
cells were collected following treatment with curcumin at 0 (with
DMSO vehicle), 1, 2 and 5 µM for 48 h, and the cells were subjected
to lysis by a cell lysis buffer (Beyotime Institute of
Biotechnology) for 0.5–1 h on ice. The cell lysates were
centrifuged at 12,000 × g for 10 min at 4°C. The concentration of
the proteins was determined by BCA Protein Assay kit. The proteins
were mixed with reaction buffers from the caspase-3/9 activity kits
(caspase-3, Ac-DEVD-pNA; caspase-9, Ac-LEHD-pNA) and incubated at
37°C for 2 h in the dark. The activities of caspase-3 and caspase-9
were measured at an absorbance of 405 nm using the microplate
reader from Bio-Rad Laboratories, Inc.
Reverse transcription-quantitative
polymerase chain reaction (qPCR) of miR-21 expression
MCF-7 cells were seeded in 6-well plates at a
density of 1×106 cells/well (2 ml/well) for 24 h. MCF-7
cells were collected following treatment with curcumin at 0 (with
DMSO vehicle), 1, 2 and 5 µM for 48 h, and total RNA was extracted
using TRIzol reagent (Invitrogen™; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. PrimeScript™ RT Master
mix was used to transcribe cDNA according to the manufacturer's
protocol (Takara Biotechnology Co., Ltd.). qPCR was performed with
an ABI Prism 7900HT Real-Time PCR system and the SYBR Green I
florescent dye method (Applied Biosystems; Thermo Fisher
Scientific, Inc.) was used to quantify miR-21 expression (3). PCR was initiated at 95°C for 3 min,
followed by 40 cycles at 95°C for 20 sec and 60°C for 30 sec. The
primers used are as follows: miR-21, forward
5′-GGGGTAGCTTATCAGACTGATG-3′ and reverse 5′-TGTCGTGGAGCGGCAATTG-3′;
U6, forward 5′-CGCTTCGGCACATATACTA-3′ and reverse
5′-CGCTTCACGAATTTGCGTGTCA-3′. The relative level of miR-21 was
calculated with the comparative 2−ΔΔCq method using
small RNA U6 (13).
Western blot analysis for PTEN/Akt
protein expression
MCF-7 cells were seeded in 6-well plates at a
density of 1×106 cells/well (2 ml/well) for 24 h. The
cells were collected following treatment with curcumin at 0 (with
DMSO vehicle), 1, 2 and 5 µM for 48 h, and MCF-7 cells were
subjected to cell lysis with a cell lysis buffer for 30 min to 1 h
on ice. The cell lysates were centrifuged at 12,000 × g for 10 min
at 4°C and the concentration of proteins was determined by BCA
Protein Assay kit. The proteins were subjected to 12% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis. Following
electrophoresis, the gels were transferred to polyvinylidene
difluoride membrane (0.22 mm). The membrane was blocked in 5%
non-fat milk in PBS at room temperature for 2 h. The transferred
membrane was incubated with rabbit polyclonal anti-PTEN (dilution,
1:1,500; sc-6817-R; Santa Cruz Biotechnologies, Inc., Dallas, TX,
USA), anti-phospho-Akt (p-Akt; dilution, 1:1,500; sc-135650; Santa
Cruz Biotechnologies, Inc.) and anti-β-actin (dilution, 1:500;
D110007; Sangon Biotech, Co., Ltd., Shanghai, China) antibodies
overnight at 4°C. After washing with PBS three times, the membrane
was incubated with a secondary polyclonal peroxidase-labeled
antibody (6403–05; Wuhan Amyjet Scientific, Inc., Wuhan, China) for
2 h. The membrane was detected using the Immun-Star™ WesternC™
Chemiluminescent kit (Bio-Rad Laboratories, Inc.) and
autoradiography films (Amersham HyperFilm ECL; GE Healthcare).
Quantification was performed using Quantity One software version
3.0 (Bio-Rad Laboratories, Inc.).
Transfection of MCF-7 cells with
miR-21 plasmids
Negative control plasmids and miR-21 plasmids were
designed and purchased from Sangon Biotech, Co., Ltd. The MCF-7
cells were seeded onto a 6-well plate at a density of
1×106 cells/well for 24 h, and were transiently
transfected using Lipofectamine® 2000 transfection
reagent (Invitrogen™; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol.
Statistical analysis
Data are expressed as the mean ± standard error of
the mean. Comparisons of two groups were performed using Student's
t-test and multiple comparisons were made by performing one-way
analysis of variance followed by Bonferroni's test. Statistical
analysis was conducted using SPSS software (version 15; SPSS, Inc.,
Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Anticancer effect of curcumin inhibits
cell growth
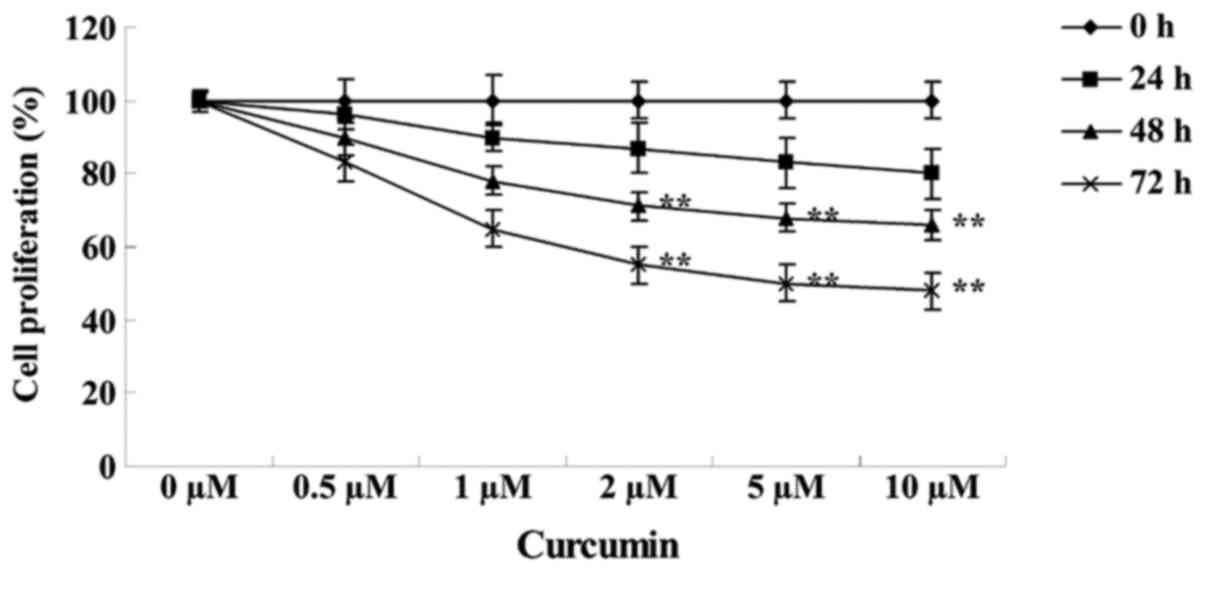
The structure of curcumin is shown in Fig. 1. MTT assay revealed that the growth of
MCF-7 cells was reduced following treatment with various
concentrations of curcumin (0, 0.5, 1.0, 2.0, 5.0 and 10.0 µM) for
24, 48 and 72 h. Subsequent to treatment with 2, 5 and 10 µM
curcumin for 48 (P=0.0097, 0.0041 and 0.0022, respectively) or 72 h
(P=0.0072, 0.0033 and 0.0009, respectively), the cell growth of
MCF-7 cells was significantly inhibited in a concentration- and
time-dependent manner compared with cells treated with 0 µM
curcumin (Fig. 2).
Anticancer effect of curcumin induces
cytotoxicity
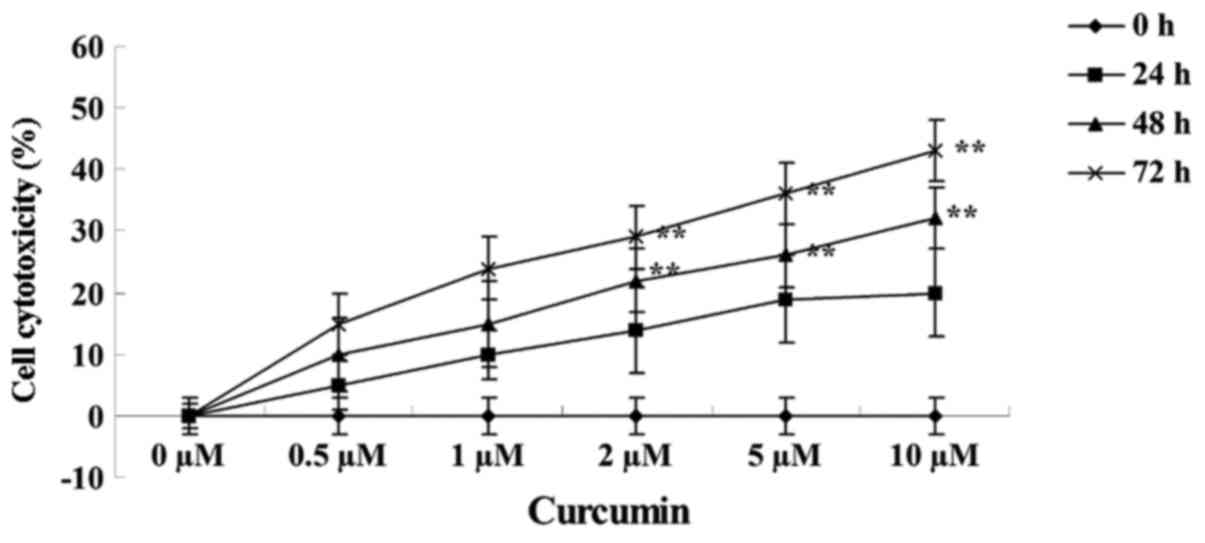
The effects of treating MCF-7 cells with various
concentrations of curcumin (0, 0.5, 1.0, 2.0, 5.0 and 10.0 µM) for
24, 48 and 72 h on cell cytotoxicity was analyzed using a LDH
assay. Curcumin treatment at 2, 5 and 10 µM for 48 (P=0.0088,
0.0038 and 0.0017, respectively) and 72 h (P=0.0079, 0.0027 and
0.0003, respectively) increased the cytotoxicity of MCF-7 cells in
a concentration- and time-dependent manner (Fig. 3).
Anticancer effect of curcumin induces
cell apoptosis
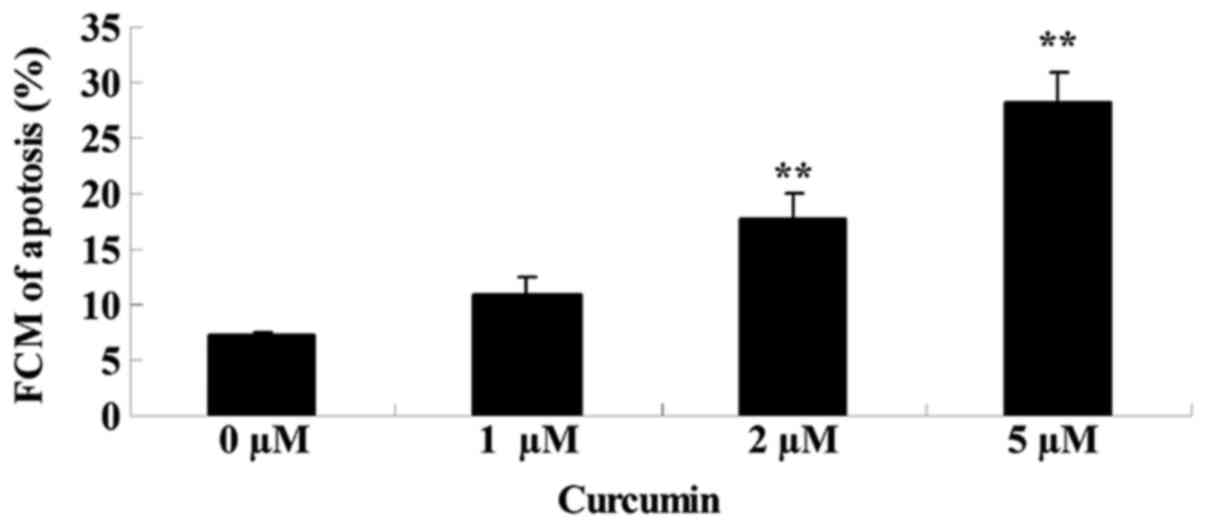
The effects of treating MCF-7 cells with various
concentrations of curcumin (0, 1, 2 and 5 µM) for 48 h on cell
apoptosis was analyzed using flow cytometry. As shown in Fig. 4, curcumin induced significant
apoptotic cell death of MCF-7 cells in a concentration-dependent
manner, when treated with 2 (P=0.0021) and 5 µM (P=0.0004) at 48
h.
Anticancer effect of curcumin induces
apoptotic morphology in cells
The effects of treating MCF-7 cells with various
concentrations of curcumin (0, 1, 2 and 5 µM) for 48 h on nucleic
morphology was analyzed using DAPI staining. As shown in Fig. 5, nucleic apoptosis of MCF-7 cells was
observed to be significantly promoted by treatment with 2 and 5 µM
curcumin at 48 h in a concentration-dependent manner.
Anticancer effect of curcumin induces
caspase-3/9 activities
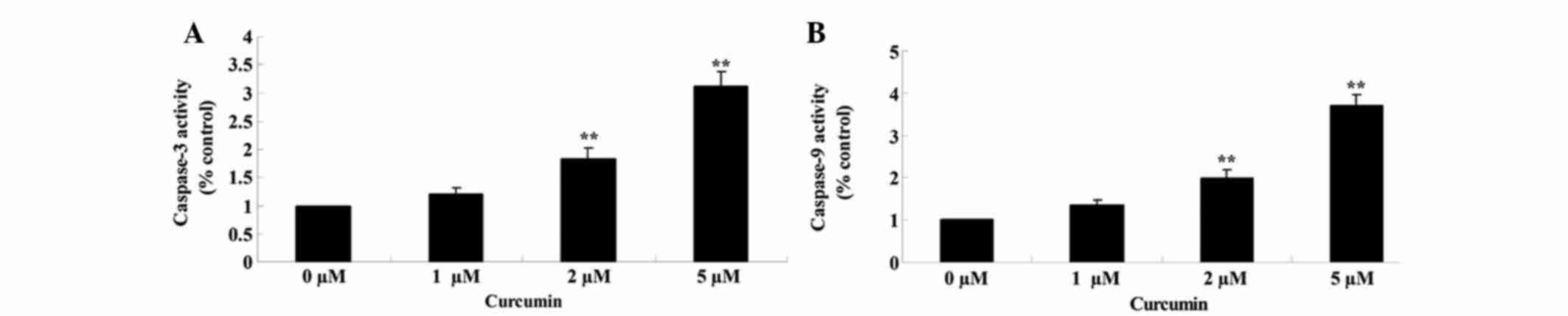
Effects of curcumin on caspase-3/9 activities of
MCF-7 cells was examined using commercial caspase-3/9 activity
kits, after the cells had been treated with curcumin at 0, 1, 2 and
5 µM for 48 h. When treated with curcumin at 2 and 5 µM, caspase-3
(P=0.0057 and 0.0012) and −9 (P=0.0042 and 0.0008) activities in
MCF-7 cells were significantly increased in a
concentration-dependent manner (Fig. 6A
and B).
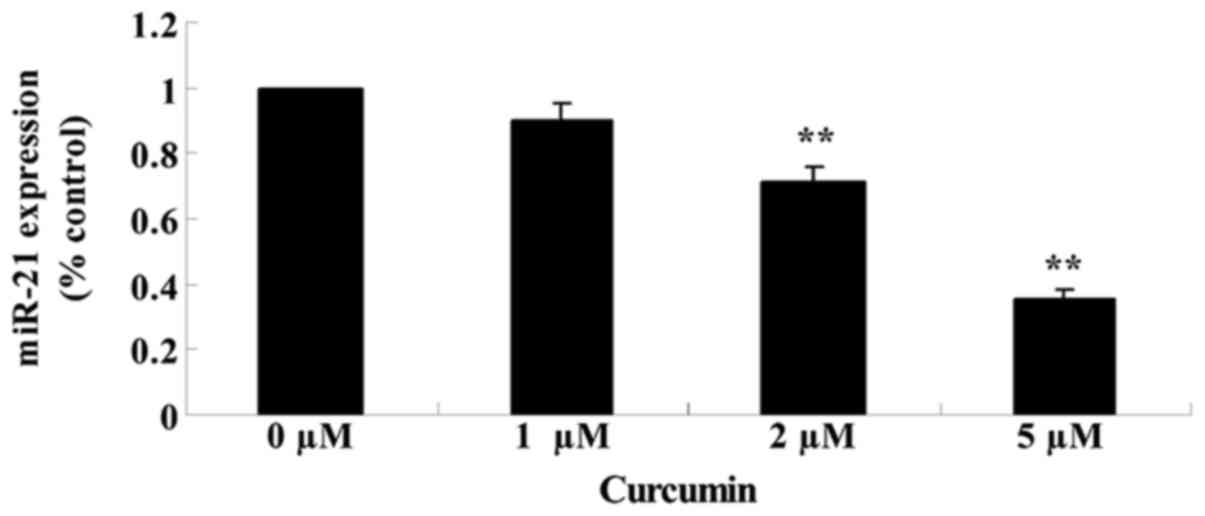
Anticancer effect of curcumin
regulates miR-21 expression
The effects of treating MCF-7 cells with various
concentrations of curcumin (0, 1, 2 and 5 µM) for 48 h on the
expression of miR-21 was analyzed using qPCR. Fig. 7 reveals that the expression of miR-21
was significantly suppressed in cells treated with curcumin in a
concentration-dependent manner (P=0.0079 and 0.0021).
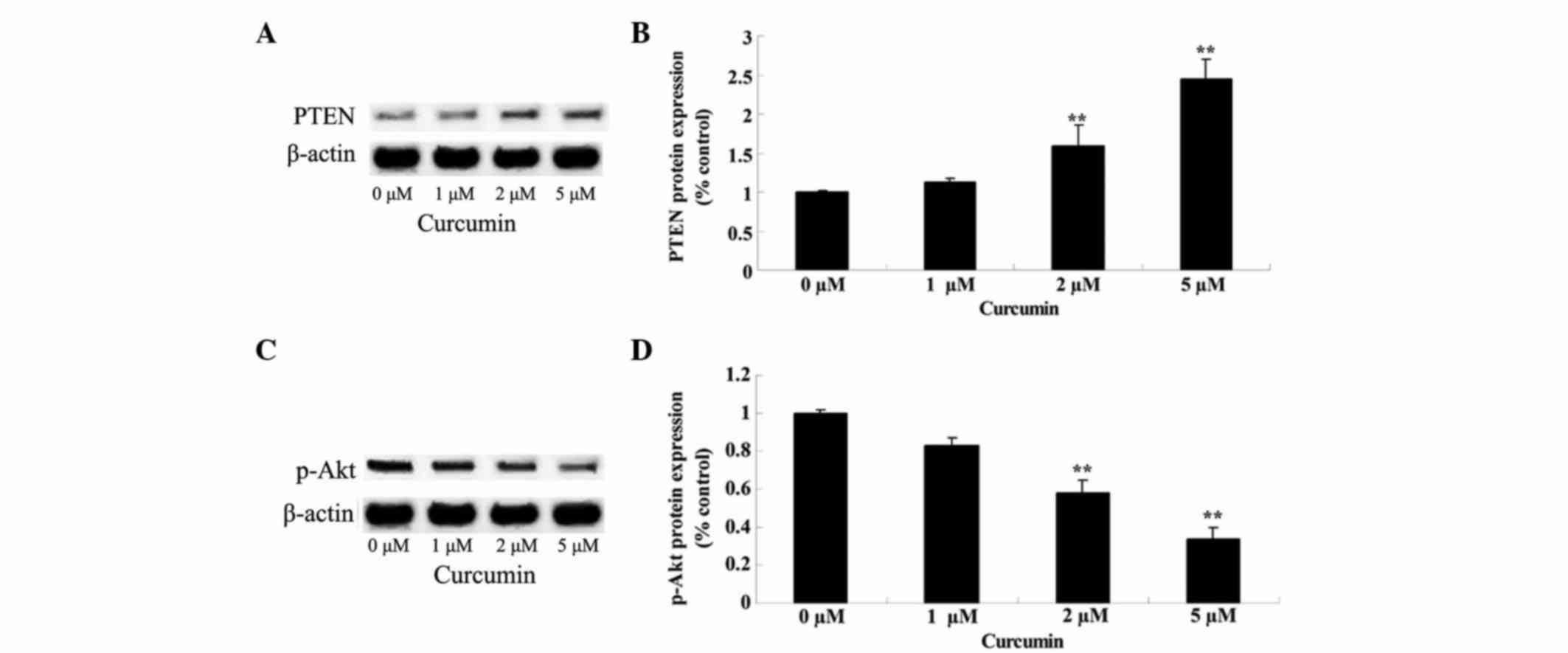
Anticancer effect of curcumin
regulates PTEN/Akt
Effects of curcumin on the PTEN/Akt signaling
pathway in MCF-7 cells was verified by western blot analysis,
subsequent to the cells being treated with curcumin at 0, 1, 2 and
5 µM for 48 h. Following treatment, PTEN protein expression was
significantly increased (P=0.0081 and 0.0009) and p-Akt protein
expression was significantly decreased (P=0.0092 and 0.0033) in a
concentration-dependent manner (Fig.
8).
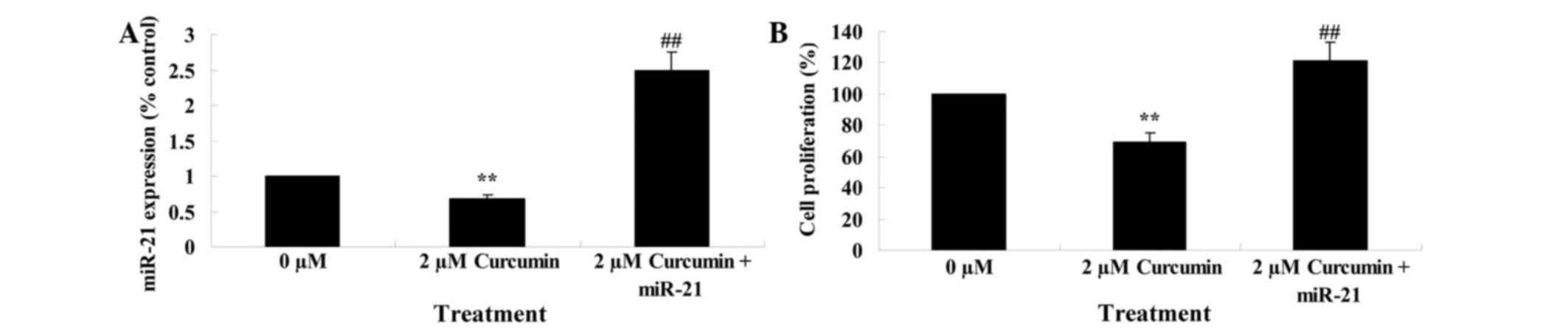
Upregulation of miR-21 regulates the
anticancer effect of curcumin on cell growth
The previous results had demonstrated that miR-21
plays a role in the anticancer effect of curcumin on cell growth.
Therefore, miR-21 was upregulated in MCF-7 cells by transfection of
MCF-7 cells with miR-21 plasmids, and the anticancer effect of
curcumin on the cell growth of these transfected cells was
evaluated. miR-21 plasmids significantly upregulated miR-21
expression (Fig. 9A). Overexpression
of miR-21 significantly reduced the anticancer effect of curcumin,
as demonstrated by an increase in cell proliferation of transfected
cells compared with parental MCF-7 cells (P=0.0037; Fig. 9B).
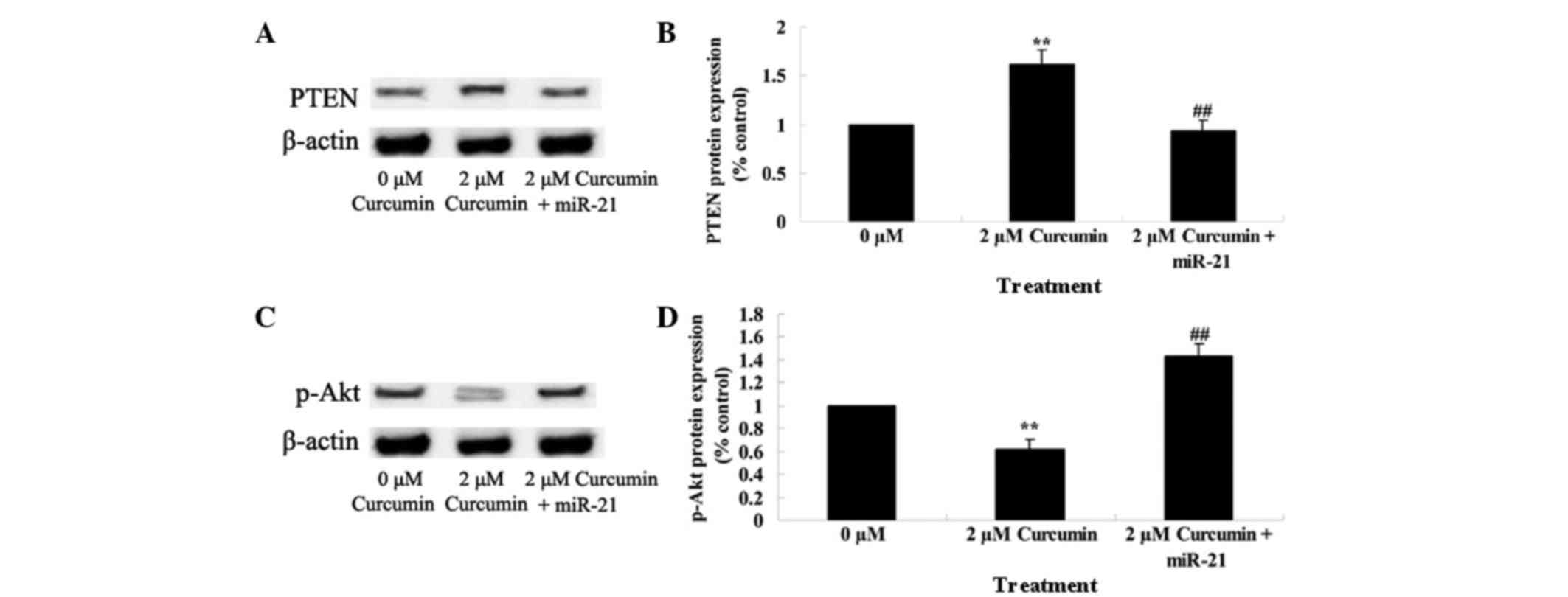
Upregulation of miR-21 regulates the
anticancer effect of curcumin through the PTEN/Akt pathway
The present study additionally investigated whether
miR-21 regulates the anticancer effect of curcumin through the
PTEN/Akt pathway in MCF-7 cells. As shown in Fig. 10, miR-21 plasmids significantly
decreased the anticancer effect of curcumin by decreasing the
expression of PTEN and increasing the expression of p-Akt
(P=0.0039).
Discussion
Breast cancer is one of the most common malignant
tumors worldwide. According to the latest global estimates, in 2002
morbidity from breast cancer was 37.5 per 100,000 individuals, and
the incidence was 1.15 million patients (3). For nearly 30 years, the morbidity caused
by breast cancer in the majority of countries has increased
(14,15). The present study demonstrates that the
anticancer effect of curcumin significantly inhibits cell growth,
increases cytotoxicity and induces apoptosis and caspase-3/9
activities in MCF-7 cells in a concentration- and time-dependent
manner. In addition, the anticancer activity of curcumin has been
demonstrated to inhibit cell proliferation and induce apoptosis in
lung adenocarcinoma A549 cells (16)
non-small cell lung cancer (17),
gastric cancer (12) and colorectal
cancer (18). Therefore, curcumin may
be a potential drug for breast cancer treatment.
For numerous types of tumors, inhibiting the
expression of miR-21 leads to an inhibition in cell growth, induces
apoptosis and inhibits cell migration and invasion (19). miR-21 is regarded as a proto-oncogene,
and has an increased expression in various human cancer tissues and
cells, including glioma, multiple myeloma and cervical, ovarian,
bladder, prostate, lung and breast cancer (20,21).
miR-21 not only participates in the differentiation, proliferation
and apoptosis of cells, but is closely associated with the
occurrence, growth, invasion and metastasis of tumors (22). The present study demonstrates that the
anticancer effect of curcumin significantly reduced the expression
of miR-21 in MCF-7 cells in a concentration-dependent manner.
Similarly, Mudduluru et al (23) revealed that curcumin inhibits the
invasion and metastasis of colorectal cancer cells through the
regulation of miR-21 expression. Gaining insight into the molecular
characteristics of miR-21 may aid in understanding the antitumor
function of curcumin in various human cancers.
Previous studies have suggested that PTEN is a
target gene with certain biological functions. As a
tumor-inhibiting factor, PTEN controls the growth, invasion and
metastasis of tumors (24).
Inhibition of PTEN may affect downstream molecules of the
phosphoinositide 3-kinase (PI3K) signaling pathway, including Akt,
which leads to a stimulation of the growth and invasion and
metastatic abilities of tumors (25).
An overexpression of miR-21 in human hepatoma cells leads to a
decrease in the expression of PTEN; therefore, it may be
hypothesized that there is a negative association between PTEN and
miR-21 (26). In addition, miR-21
possibly promotes the occurrence and growth of tumors by inhibiting
the expressions of PTEN (27). The
present study demonstrated that the anticancer effects of curcumin
significantly promoted the protein expression of PTEN and
suppressed the protein expression of p-Akt in MCF-7 cells.
Furthermore, Khaw et al (28)
reported that curcumin inhibits telomerase and induces cell
apoptosis of brain tumour cells by upregulating the expression of
PTEN. Additionally, Wong et al (29) revealed that curcumin inhibits uterine
leiomyosarcoma cells via Akt. However, the detailed mechanisms
concerning the effects of curcumin treatment on the PTEN/Akt
pathway remains unclear, and further clarification is required.
The PTEN/PI3K/Akt signaling pathway exists
extensively in all types of tumor cells (30). Akt affects the apoptosis of cancer
cells, myocardial cells and other cells (31). This pathway affects cell cycle
progress by controlling the activation of genes associated with
apoptosis; therefore, control of the growth and proliferation of
cells is via this pathway. This pathway participates in the
formation of tumor vasculature and consequently is important in the
formation of tumors (32). In
addition, the PTEN/PI3K/Akt pathway participates in the invasion
and metastasis of tumors. PTEN inhibits the formation of tumors
through negative regulation of the PI3K/Akt signaling pathway, and
an inactivation or mutation of PTEN genes decreases the inhibition
of the pathway causing cell canceration (33). In the process of PI3K/AKT signal
transduction, PI3K is activated, which in turn activates PIP3 as a
secondary messenger that activates signal molecules downstream. The
PI3K signaling pathway is important in a series of
pathophysiological procedures, including cell differentiation,
apoptosis, proliferation and migration and the neoplastic
transformation of cells (34).
Certain studies have demonstrated that PTEN/PI3K/Akt signaling
pathways are closely associated with the cell growth,
proliferation, survival and apoptosis, metabolism of tumors, tumor
angiogenesis, invasion and metastasis, drug resistance in cancer
and tumor immune response escape (35).
Overall, an upregulation of miR-21 significantly
inhibited the anticancer effect of curcumin on cell growth in MCF-7
cells, suppressed the promotion of PTEN protein expression and
increased p-Akt protein expression. In conclusion, the present
results of this initial study indicate that curcumin reduces the
malignancy of breast cancer cells, and this anticancer effect is
controlled by the miR-21/PTEN/Akt signaling pathway. These results
provide a basis for the future management of breast cancer. In
conclusion, the results of the present study indicate that curcumin
reduces the malignancy of breast cancer cells, and this anticancer
effect is controllled by a miR-21/PTEN/ Akt signaling pathway,
which may present a novel method of treating breast cancer.
References
|
1
|
Pu Z, Yuan X, Zhang X, Chen Q and Xie H:
Meta-analysis on the association between CYP2D6*10 gene
polymorphism and disease free survival of breast cancer patients
receiving tamoxifen treatment in Asia. Bangladesh J Pharmacol.
9:652–662. 2014. View Article : Google Scholar
|
|
2
|
Rao X, Evans J, Chae H, Pilrose J, Kim S,
Yan P, Huang RL, Lai HC, Lin H, Liu Y, et al: CpG island shore
methylation regulates caveolin-1 expression in breast cancer.
Oncogene. 32:4519–4528. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang J, Wang Y, Yin Q, Zhang W, Zhang T
and Niu Y: An associated classification of triple negative breast
cancer: The risk of relapse and the response to chemotherapy. Int J
Clin Exp Pathol. 6:1380–1391. 2013.PubMed/NCBI
|
|
4
|
Moshammer MI, Kalipciyan M, Bartsch R,
Steger GG, Sedivy R and Mader RM: Exosomal microRNA transfer varies
with specific microRNAs functional in colorectal cancer and
cellular differentiation. Int J Clin Pharmacol Ther. 52:87–88.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Catania A, Maira F, Skarmoutsou E, D'Amico
F, Abounader R and Mazzarino MC: Insight into the role of microRNAs
in brain tumors (review). Int J Oncol. 40:605–624. 2012.PubMed/NCBI
|
|
6
|
Osawa S, Shimada Y, Sekine S, Okumura T,
Nagata T, Fukuoka J and Tsukada K: MicroRNA profiling of gastric
cancer patients from formalin-fixed paraffin-embedded samples.
Oncol Lett. 2:613–619. 2011.PubMed/NCBI
|
|
7
|
Chiu J, Khan ZA, Farhangkhoee H and
Chakrabarti S: Curcumin prevents diabetes-associated abnormalities
in the kidneys by inhibiting p300 and nuclear factor-kappaB.
Nutrition. 25:964–972. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang J, Du XX, Jiang H and Xie JX:
Curcumin attenuates 6-hydroxydopamine-induced cytotoxicity by
anti-oxidation and nuclear factor-kappa B modulation in MES23.5
cells. Biochem Pharmacol. 78:178–183. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu SJ, Tam KW, Tsai YH, Chang CC and Chao
JC: Curcumin and saikosaponin a inhibit chemical-induced liver
inflammation and fibrosis in rats. Am J Chin Med. 38:99–111. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhen L, Fan D, Yi X, Cao X, Chen D and
Wang L: Curcumin inhibits oral squamous cell carcinoma
proliferation and invasion via EGFR signaling pathways. Int J Clin
Exp Pathol. 7:6438–6446. 2014.PubMed/NCBI
|
|
11
|
Ye M and Zhang J and Zhang J, Miao Q, Yao
L and Zhang J: Curcumin promotes apoptosis by activating the
p53-miR-192-5p/215-XIAP pathway in non-small cell lung cancer.
Cancer Lett. 357:196–205. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu X, Sun K, Song A, Zhang X, Zhang X and
He X: Curcumin inhibits proliferation of gastric cancer cells by
impairing ATP-sensitive potassium channel opening. World J Surg
Oncol. 12:3892014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak and Schmittgen: Analysis of relative
gene expression data using real-time quantitative PCR and the
2-ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jiang H, Bai X, Zhang C and Zhang X:
Evaluation of HER2 gene amplification in breast cancer using nuclei
microarray in situ hybridization. Int J Mol Sci. 13:5519–5527.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Menendez JA and Lupu R: RNA
interference-mediated silencing of the p53 tumor-suppressor protein
drastically increases apoptosis after inhibition of endogenous
fatty acid metabolism in breast cancer cells. Int J Mol Med.
15:33–40. 2005.PubMed/NCBI
|
|
16
|
Teong B, Lin CY, Chang SJ, Niu GC, Yao CH,
Chen IF and Kuo SM: Enhanced anti-cancer activity by
curcumin-loaded hydrogel nanoparticle derived aggregates on A549
lung adenocarcinoma cells. J Mater Sci Mater Med. 26:53572015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen QY, Jiao DM, Wang LF, Wang L, Hu HZ,
Song J, Yan J, Wu LJ and Shi JG: Curcumin inhibits
proliferation-migration of NSCLC by steering crosstalk between a
Wnt signaling pathway and an adherens junction via EGR-1. Mol
Biosyst. 11:859–868. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fan L, Li A, Li W, Cai P, Yang B, Zhang M,
Gu Y, Shu Y, Sun Y, Shen Y, et al: Novel role of Sarco/endoplasmic
reticulum calcium ATPase 2 in development of colorectal cancer and
its regulation by F36, a curcumin analog. Biomed Pharmacother.
68:1141–1148. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shi J: Considering exosomal miR-21 as a
biomarker for cancer. J Clin Med. 5:422016. View Article : Google Scholar
|
|
20
|
Frey MR, Carraro G, Batra RK, Polk DB and
Warburton D: Sprouty keeps bowel kinases regular in colon cancer,
while miR-21 targets Sprouty. Cancer Biol Ther. 11:122–124. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Han M, Liu M, Wang Y, Chen X, Xu J, Sun Y,
Zhao L, Qu H, Fan Y and Wu C: Antagonism of miR-21 reverses
epithelial-mesenchymal transition and cancer stem cell phenotype
through AKT/ERK1/2 inactivation by targeting PTEN. PLoS One.
7:e395202012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yin C, Zhou X, Dang Y, Yan J and Zhang G:
Potential role of circulating miR-21 in the diagnosis and prognosis
of digestive System Cancer: A systematic review and meta-analysis.
Medicine (Baltimore). 94:e21232015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mudduluru G, George-William JN, Muppala S,
Asangani IA, Kumarswamy R, Nelson LD and Allgayer H: Curcumin
regulates miR-21 expression and inhibits invasion and metastasis in
colorectal cancer. Biosci Rep. 31:185–197. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Leslie NR, Kriplani N, Hermida MA,
Alvarez-Garcia V and Wise HM: The PTEN protein: Cellular
localization and post-translational regulation. Biochem Soc Trans.
44:273–278. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yi XP, Han T, Li YX, Long XY and Li WZ:
Simultaneous silencing of XIAP and survivin causes partial
mesenchymal-epithelial transition of human pancreatic cancer cells
via the PTEN/PI3K/Akt pathway. Mol Med Rep. 12:601–608.
2015.PubMed/NCBI
|
|
26
|
He C, Dong X, Zhai B, Jiang X, Dong D, Li
B, Jiang H, Xu S and Sun X: MiR-21 mediates sorafenib resistance of
hepatocellular carcinoma cells by inhibiting autophagy via the
PTEN/Akt pathway. Oncotarget. 6:28867–28881. 2015.PubMed/NCBI
|
|
27
|
Schwarzenbach H, Milde-Langosch K,
Steinbach B, Müller V and Pantel K: Diagnostic potential of
PTEN-targeting miR-214 in the blood of breast cancer patients.
Breast Cancer Res Treat. 134:933–941. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Khaw AK and Hande MP, Kalthur G and Hande
MP: Curcumin inhibits telomerase and induces telomere shortening
and apoptosis in brain tumour cells. J Cell Biochem. 114:1257–1270.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wong TF, Takeda T, Li B, Tsuiji K,
Kitamura M, Kondo A and Yaegashi N: Curcumin disrupts uterine
leiomyosarcoma cells through AKT-mTOR pathway inhibition. Gynecol
Oncol. 122:141–148. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang J, Yang Y, Zhang Z, He Y, Liu Z, Yu
Y, Wu S, Cai B and Feng Y: Gankyrin plays an essential role in
estrogen-driven and GPR30-mediated endometrial carcinoma cell
proliferation via the PTEN/PI3K/AKT signaling pathway. Cancer Lett.
339:279–287. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xue G, Zippelius A, Wicki A, Mandalà M,
Tang F, Massi D and Hemmings BA: Integrated Akt/PKB signaling in
immunomodulation and its potential role in cancer immunotherapy. J
Natl Cancer Inst. 107:2015
|
|
32
|
Park JH, Lee JY, Shin DH, Jang KS, Kim HJ
and Kong G: Loss of Mel-18 induces tumor angiogenesis through
enhancing the activity and expression of HIF-1alpha mediated by the
PTEN/PI3K/Akt pathway. Oncogene. 30:4578–4589. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao M, Zhou A, Xu L and Zhang X: The role
of TLR4-mediated PTEN/PI3K/AKT/NF-κB signaling pathway in
neuroinflammation in hippocampal neurons. Neuroscience. 269:93–101.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li P, Xiao LY and Tan H: Muc-1 promotes
migration and invasion of oral squamous cell carcinoma cells via
PI3K-Akt signaling. Int J Clin Exp Pathol. 8:10365–10374.
2015.PubMed/NCBI
|
|
35
|
Yang X, Cheng Y, Li P, Tao J, Deng X,
Zhang X, Gu M, Lu Q and Yin C: A lentiviral sponge for miRNA-21
diminishes aerobic glycolysis in bladder cancer T24 cells via the
PTEN/PI3K/AKT/mTOR axis. Tumour Biol. 36:383–391. 2015. View Article : Google Scholar : PubMed/NCBI
|