Introduction
Ovarian cancer is a gynecological malignancy with
one of the highest mortality rates worldwide (1). Even though the incidence of cervical
cancer and endometrial cancer are higher at present (2), ovarian cancer ranks fifth as a cause of
cancer-associated mortality among women (3,4). Ovarian
cancer is not typically diagnosed at an early stage, with the
majority of women already at advanced stages (International
Federation of Gynecology and Obstetrics stage) when diagnosed,
which presents a severe challenge for resection and recovery
(5). Based on this situation, a
combination of surgery and chemotherapy are the conventional
methods of treatment. However, despite the efficacy of
chemotherapy, its side effects are inevitable. Thus, a novel
therapeutic agent against ovarian cancer is required.
At present, the use of natural agents for cancer
prevention and therapy is attracting increased attention. Grifolin,
which originates from the edible fruiting bodies of the mushroom
AIbatrellus confluence, has been reported to possess
antioxidant and antitumor activities (6). However, the effect of grifolin on
ovarian cancer cells remains to be investigated. Therefore, the aim
of the present study was to examine the effect of grifolin on the
human ovarian cancer cell line, A2780, in vitro.
The present study revealed that grifolin effectively
suppressed viability and induced apoptosis of human ovarian cancer
A2780 cells. The present study also researched the exact targets
and molecular mechanisms underlying grifolin-induced anticancer
activity in A2780 cells, and revealed that inactivation of protein
kinase B (Akt) and extracellular signal-related kinase (ERK1/2) is
necessary for grifolin-induced cell cycle arrest and cell
apoptosis. Grifolin may therefore be a promising anti-tumor agent
for the treatment of ovarian cancer.
Materials and methods
Cell culture
The A2780 cell line was cultured in RPMI-1640 medium
with 10% fetal bovine serum. Cells were maintained in a humidified
incubator with 5% CO2 at 37°C. The cell culture media
was obtained from HyClone (GE Healthcare Life Sciences, Logan, UT,
USA), the fetal bovine serum was provided by Cell Signaling
Technology, Inc., (Danvers, MA, USA).
Drugs and chemicals
Grifolin was provided by the Kunming Institute of
Botany; Chinese Academy of Sciences (Kunming, China) with the
following structure: 2-trans, trans-farnesyl-5-methylresorcinol
(purity, >99%; high performance liquid chromatography analysis).
Grifolin was prepared at a concentration of 100 mmol/l stock
solution in dimethyl sulfoxide (DMSO) and stored under −20°C so
that the final concentration of DMSO was <0.1% in all assays it
was used in. DMSO was provided by (Shanghai Biyuntian
Bio-Technology Company, Ltd., Shanghai, China).
Cell proliferation assay
The effect of grifolin on A2780 cell viability was
evaluated by MTT assay. Cells were seeded at a density of
3,000–3,500 cells/well in 96-well plates, which permitted
logarithmic growth during the 72 h assay, and cultured overnight at
37°C with 5% CO2. Adherent cells were subsequently
exposed to grifolin at varying concentrations (0, 20, 40, 60, 80
and 100 µM) for 24, 48 and 72 h. The cells were then washed with
PBS twice and incubated in 20 µl MTT (HyClone; GE Healthcare Life
Sciences) for 4 h. The cell suspension was discarded and the cells
were supplemented with 100 µl DMSO. At the MTT endpoint, an
infinite M200PRO microplate reader (Bio-Rad Laboratories, Hercules,
CA, USA) was used to measure cell viability at 490 or 550 nm
absorbance. The cell viability trend and the half maximal
inhibitory concentrations (IC50) were analyzed using SPSS
Statistics 18.0 software (SPSS, Inc., Chicago, IL, USA). Finally,
GraphPad Prism 5 software (GraphPad Software, Inc., La Jolla, CA,
USA) was used to draw up the bar graph.
Clonogenic assay
A2780 cells (5×102) were seeded on 6-well
plates and exposed to grifolin (0, 25, 50 or 75 µM) for 48 h with
5% CO2 at 37°C. The cells were then washed with PBS
twice and RPMI-1640 culture medium was added into the plates for
~10 days. The cells were washed with PBS twice, fixed with 4%
paraformaldehyde solution for ~15 min, and then stained with 1 ml
0.2% crystal violet (HK Jimei Biology Science and Technology
Company, Ltd., Beijing, China) for 30 min. Finally, the numbers of
clones were counted and analyzed by using the DP71 fluorescence
microscope (Olympus Corporation, Tokyo, Japan) with ×200
magnification 400–600 cells per field of view assessed.
Flow cytometry assay
Flow cytometry was used to detect the cell cycle and
apoptosis distribution. Cells (1×106) were treated with
0, 25, 50 or 75 µM grifolin for 24 h, collected and washed with
ice-cold PBS twice, fixed in 75% ethanol overnight at 4°C and
stained with 20 µl Rnase A (Cell Signaling Technology, Inc.,
Danvers, MA, USA) and 400 µl propidium iodide (PI) at room
temperature for 30 min. Apoptosis analysis was also performed using
annexin V-fluorescein isothiocyanate and PI, following the
manufacturers protocol. The apoptosis kit used in flow cytometry
assay was provided by the (#50T; Jiehui Biology Science and
Biotechnology Company, Ltd., Beijing, China,). The number of cells
was calculated by FACScan flow cytometry (Bio-Rad Laboratories).
The data was analyzed by WinMDI v2.9 software (The Scripps Research
Institute, San Diego, CA, USA).
Western blot analysis
A2780 cells at a density of 4×105 were
seeded per well and were incubated with grifolin at concentrations
of 0, 25, 50 and 75 uM for 24 h, then harvested and homogenized in
100 µl RIPA lysis buffer for ~30 min on ice. The cells were
centrifuged for 15 min 4°C at the speed 15,000 × g. The protein
samples (20 µg/lane) were separated on 10 or 12% SDS-PAGE,
transferred to polyvinylidene difluoride membranes. Following
blocking using 5% non-fat milk at room temperature for ~2 h, the
membranes were incubated with primary antibodies against
phosphorylated (p-) ERK1/2 (#4370; 1:1,000; Cell Signaling
Technology, Inc.), ERK1/2 (#4060; 1:1,000; Cell Signaling
Technology, Inc., MA, USA), p-Akt (#4691; 1:1,000; Cell Signaling
Technology, Inc.), Akt (#4691; 1:1,000; Cell Signaling Technology,
Inc.), cleaved-poly (ADP-ribose) polymerase (PARP; #5625; 1:1,000;
Cell Signaling Technology, Inc.), BCL2 associated X, apoptosis
regulator (Bax; #ab32503; 1:1,000; Abcam, Cambridge, UK), B cell
lymphoma-2 (Bcl-2; #ab32124; 1:1,000; Abcam), cleaved-Caspase-3
(#ab2302; 1:1,000; Abcam), CyclinD1 (#2922; 1:1000; Cell Signaling
Technology, Inc.), cyclin dependent kinase 4 (CDK4; #12790;
1:1,000; Cell Signaling Technology, Inc.) and β-actin (#8457;
1:10,000; Cell Signaling Technology, Inc.) at 4°C overnight. The
blots were washed with TBST (1,000 ml TBST with 1 ml Tween-20)
three times and incubated with respective secondary antibodies
(Goat anti-rabbit IgG Alexa Fluor® 488; #ab150077;
1:10,000; Cell Signaling Technology, Inc.) at 4°C for 1–2 h.
Following three washes in TBST, the ImageQuant LAS 4000
(Immobilon®; EMD Millipore, Billerica, MA, USA) was used
to visualize the blots and the expression levels of the proteins
were analyzed using the NIH Image J system 1.48v (National
Institutes of Health, Bethesda, MD, USA). Statistical significance
was calculated using SPSS 18.0 software (SPSS, Inc.) and the
histograms were obtained using the Graph Pad system v5.0 (GraphPad
Software, Inc.).
Statistical analysis
A student t-test and one-way analysis of variance
was used to compare the means of different treatments, using SPSS
18.0 software (SPSS, Inc.). Date were expressed as the mean ±
standard deviation, and P<0.05 was considered to indicate a
statistically significant difference.
Results
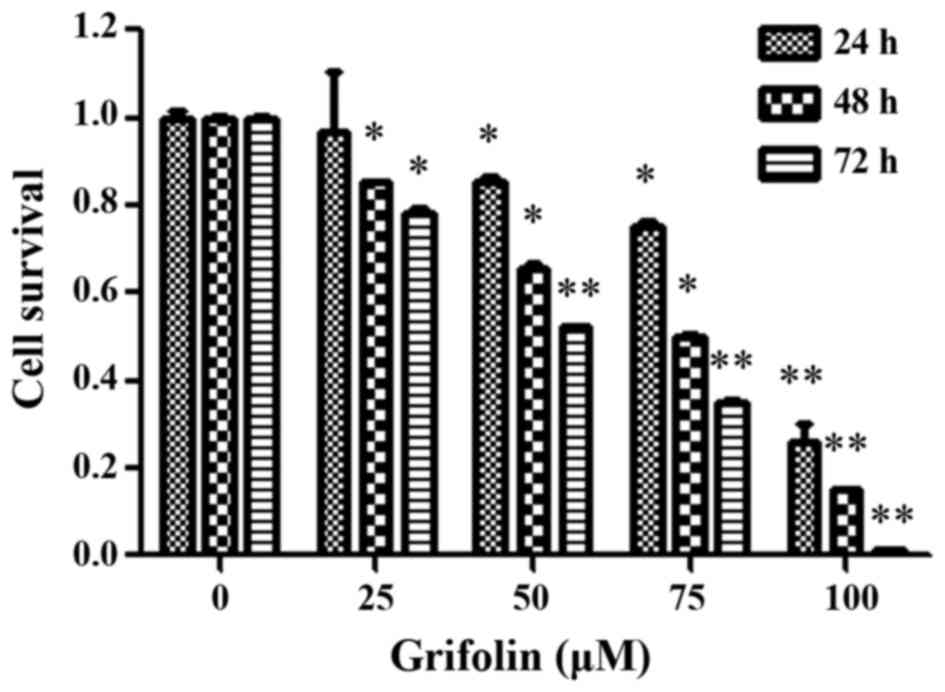
Grifolin decreases cell viability in
the human ovarian cancer cell line, A2780
The effect of grifolin on human ovarian cancer cell
viability was analyzed using the MTT assay, clonogenic assay and
flow cytometry. For this purpose, A2780 cells were incubated with
grifolin (0, 25, 50, 75 and 100 µM) for 24, 48 and 72 h. The
results revealed a significant decrease in the viability of A2780
cells, in a dose-dependent and time-dependent manner (P<0.05;
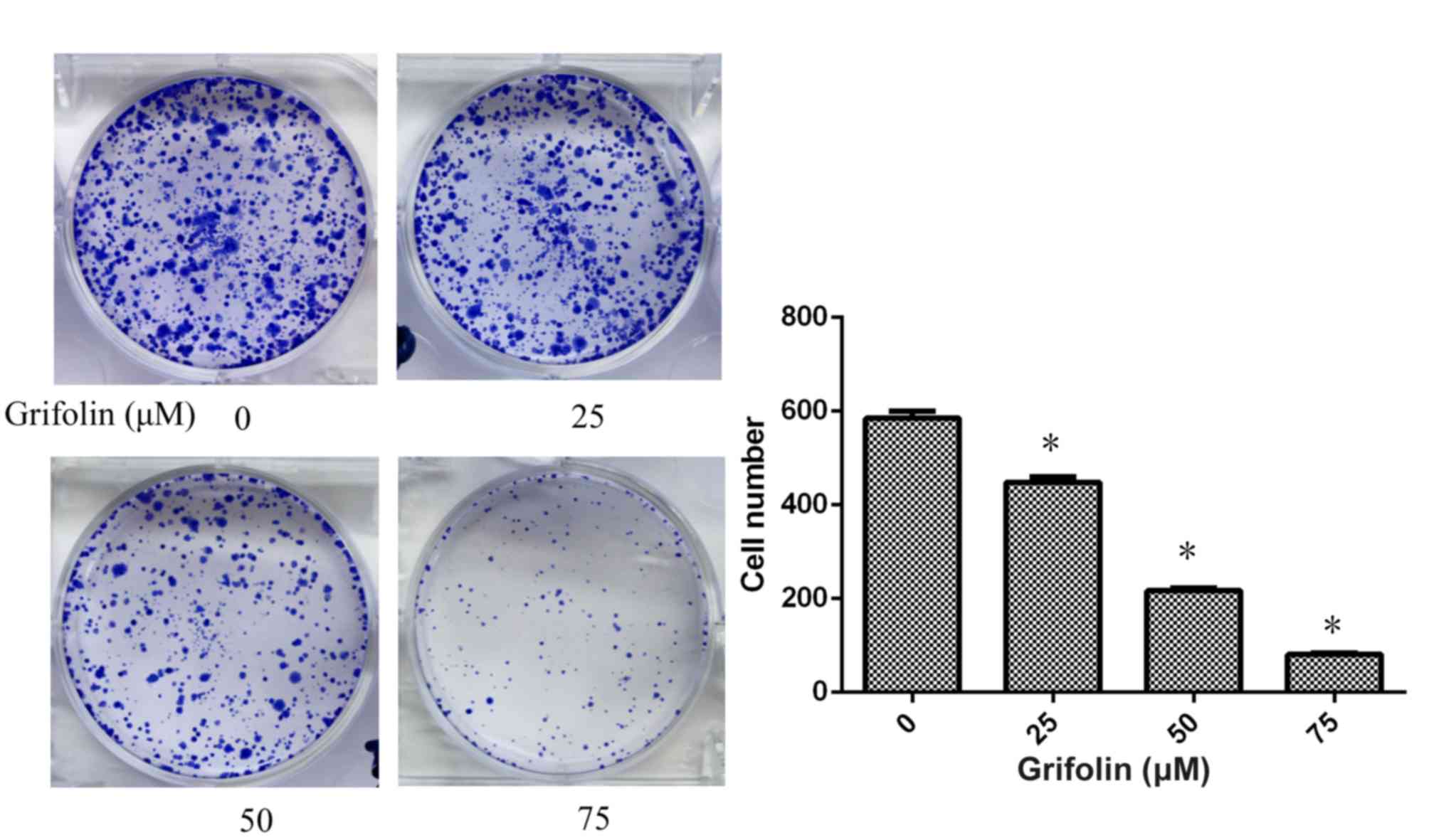
Fig. 1). A2780 cells
(5×102) were seeded on 6-well culture plates and treated
with grifolin at concentrations of 0, 25, 50 and 75 µM. The results
revealed that grifolin suppressed A2780 cell viability
significantly (P<0.05; Fig. 2).
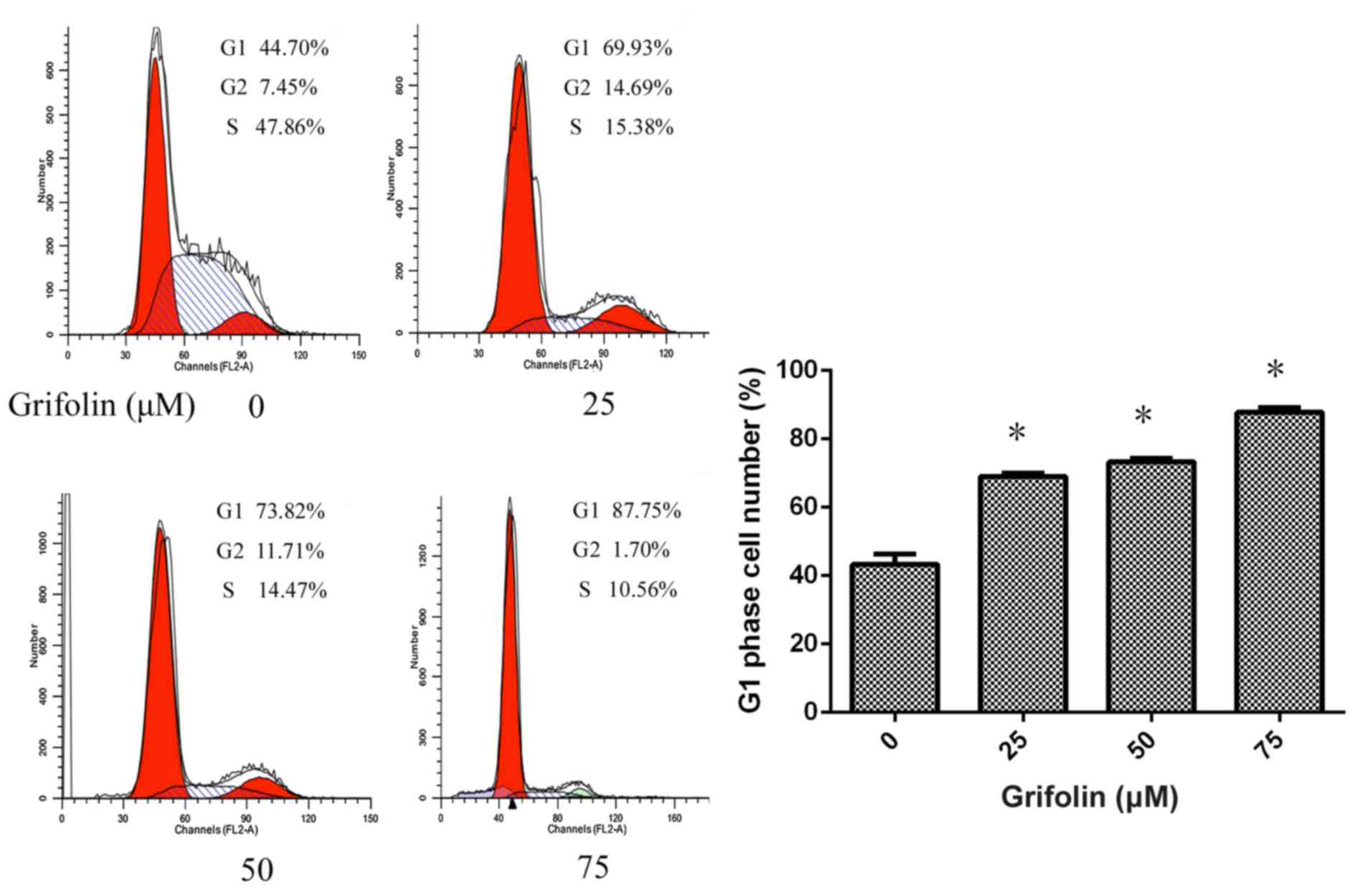
Flow cytometry analysis was used to determine the stage that A2780
cells were suspended in, and grifolin was demonstrated to result in
a cell cycle block at the G1 phase (P<0.05; Fig. 3).
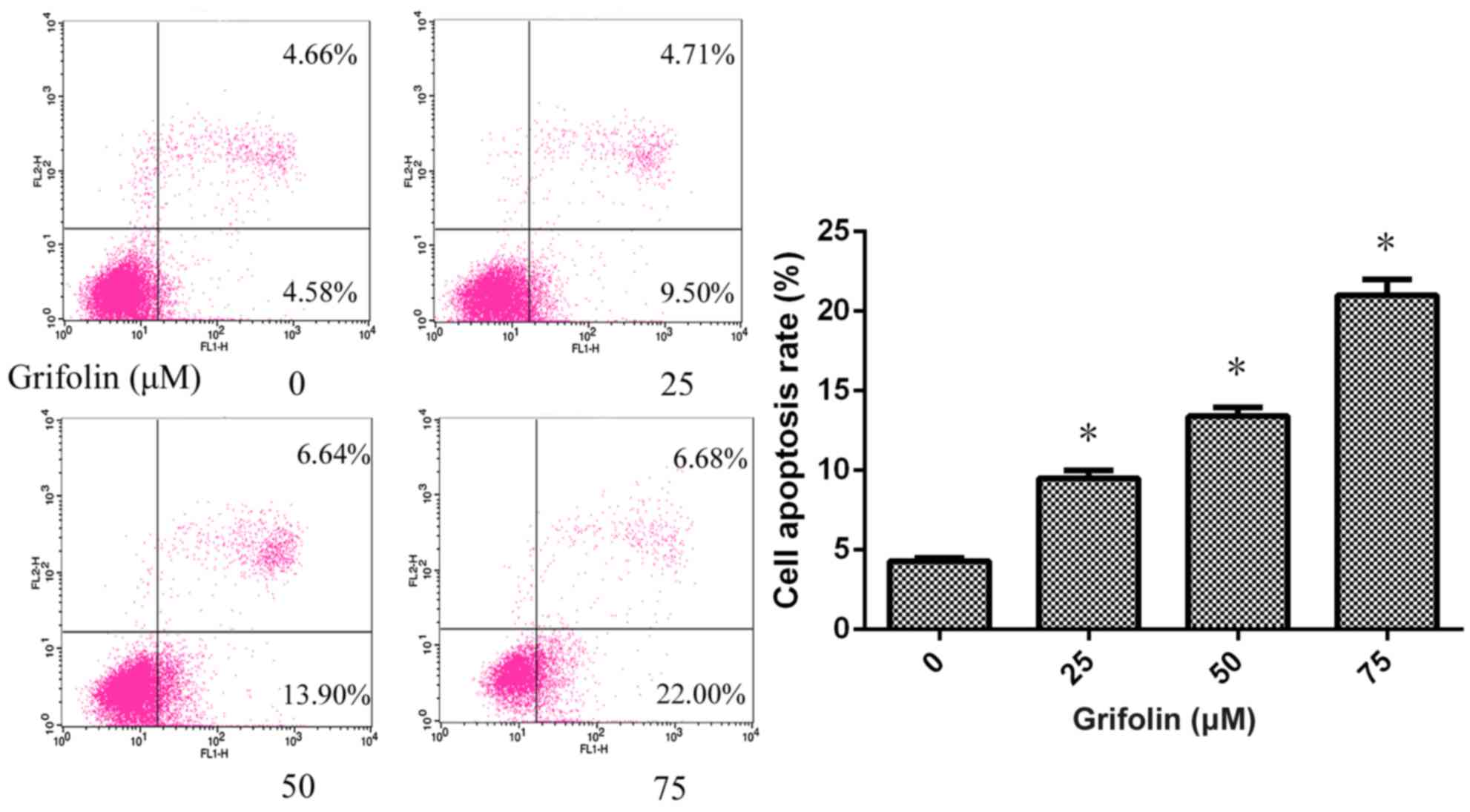
Grifolin induced apoptosis in human
A2780 ovarian cancer cells
Flow cytometry analysis was used to determine
whether grifolin induced apoptosis in A2780 cells. Grifolin-treated
A2780 cells were stained with annexin V-FITC and PI following
treatment with grifolin (0, 25, 50, and 75 uM) for 24 h.
Grifolin-treated A2780 cells were demonstrated to have undergone
apoptosis in a dose-dependent manner, and these results
corroborated the data from the MTT assay (P<0.05; Fig. 4).
Grifolin treatment affected the
expression of cell cycle proteins and apoptosis family
proteins
Next, the protein expressions of cell cycle proteins
(cyclinD1 and CDK4) and apoptosis associated proteins (Bcl-2, Bax,
cleaved-caspase-3 and cleaved-PARP) were analyzed to further assess
the potential mechanisms underlying grifolin-induced apoptosis and
inhibition of cell viability. Western blot analysis was used to
assess the expression of cyclinD1 and CDK4, which were visibly
decreased in cells treated with grifolin compared with the control
(P<0.05; Fig. 5) Western blot
analysis also revealed that Bcl-2 expression was visibly decreased,
while the expression of Bax was visibly increased compared with
untreated cells (Fig. 5). In
addition, the expression of cleaved caspase-3 and cleaved-PARP was
significantly increased compared with untreated cells (P<0.05;
Fig. 5). These results demonstrated
that grifolin treatment downregulated cell cycle proteins and
upregulated the Bax/Bcl-2 ratio, cleaved caspase-3 and cleaved-PARP
expression.
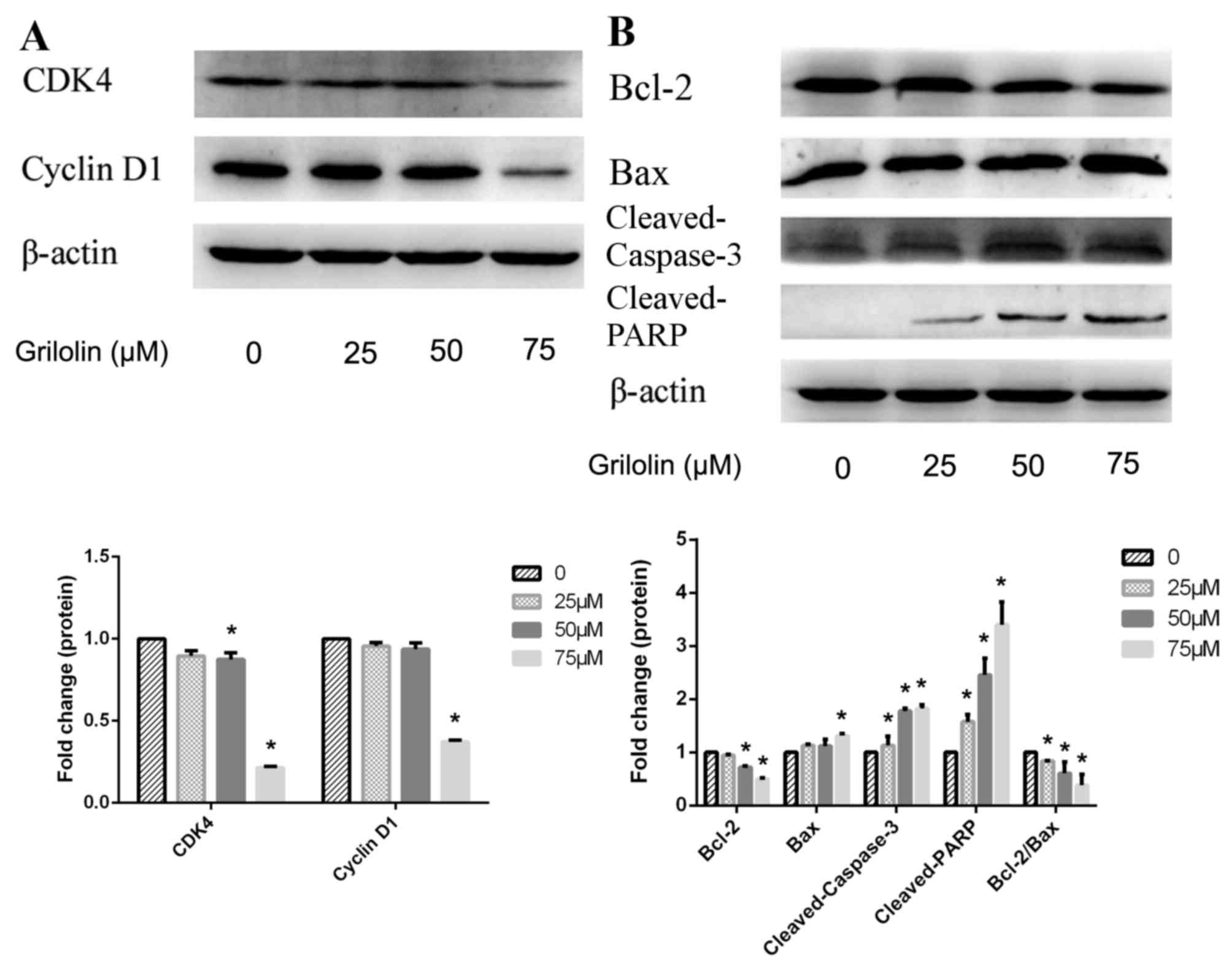 | Figure 5.(A) Western blot analysis of cell
cycle proteins of A2780 human ovarian cancer cells, following 24 h
treatment with 0, 25, 50 or 75 µM grifolin. (B) Western blot
analysis of apoptosis-associated proteins following 24 h treatment
with 0, 25, 50 or 75 µM grifolin. Equal loading and transfer were
demonstrated by repeat probing with β-actin. CDK4, cyclin dependent
kinase 4; Bcl-2, B cell lymphoma-2; Bax, Bcl-2 associated X,
apoptosis regulator; PARP, poly (ADP-ribose) polymerase. Data are
representative of 3 independent experiments. *P<0.05 vs. 0 µM
control. |
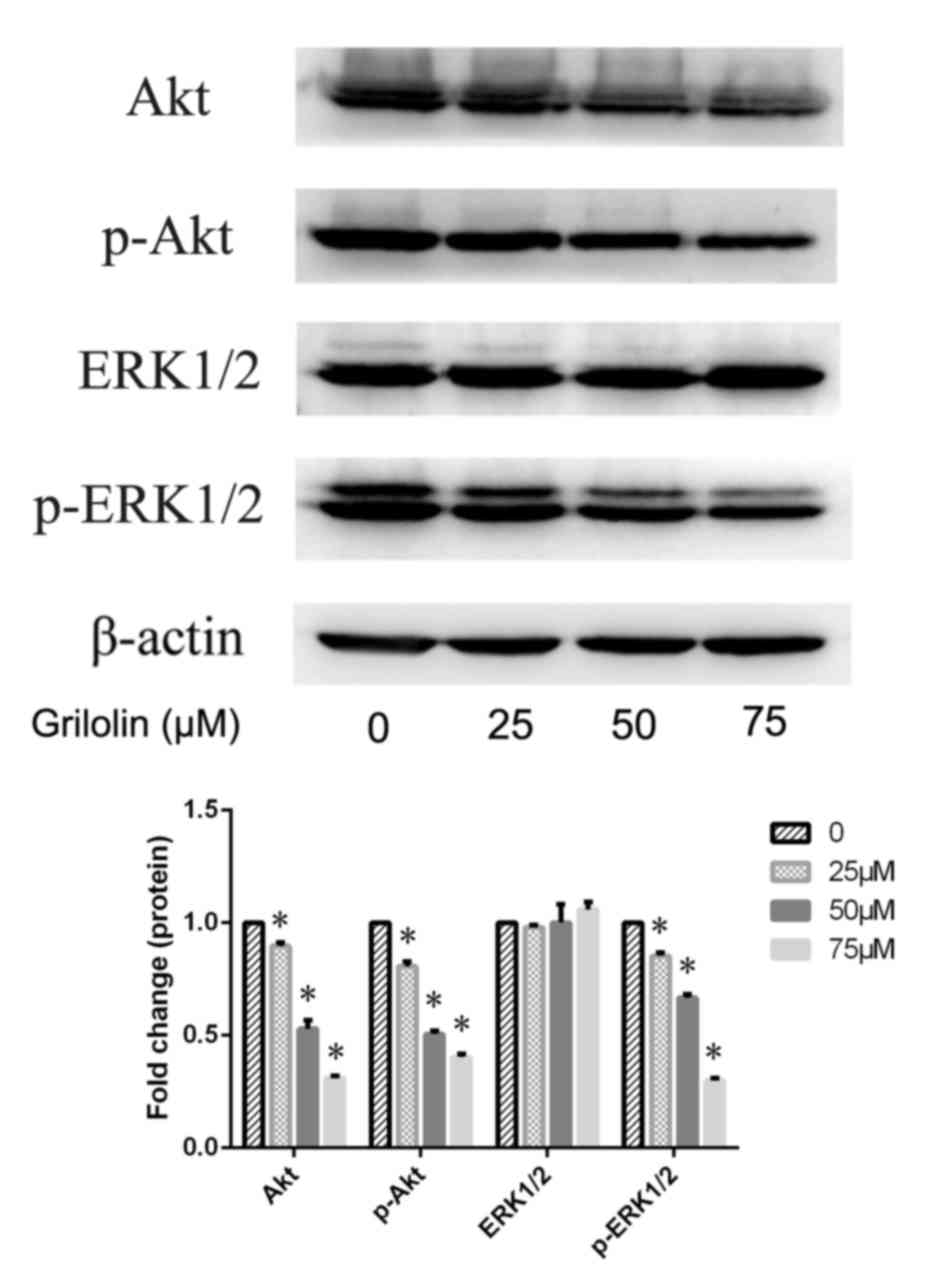
Grifolin inactivated Akt and ERK1/2 in
A2780 cells
Activation of phosphatidylinositol 3-kinase
(PI3K)/Akt and mitogen-activated protein kinase (MAPK) pathways has
been demonstrated to be associated with the expression of
apoptosis-associated proteins. Thus, western blotting was used to
evaluate the expression of Akt and ERK1/2 in grifolin-treated A2780
cells, and to determine whether the Akt and ERK1/2 pathways are
potential signaling mechanisms underlying grifolin-induced
apoptosis. The expression levels of p-Akt and p-ERK1/2 were
significantly decreased in a dose-dependent manner in
grifolin-treated cells compared with the untreated control
(P<0.05; Fig. 6). These results
suggested that inactivation of Akt and ERK1/2 may be the key
mechanism underlying grifolin-induced apoptosis.
Discussion
At present, there has been growing interest in the
function of grifolin, in particular concerning its anticancer
effects. Grifolin, a secondary metabolite product isolated from the
mushroom A. confluence, exhibits anti-cancer and therapeutic
properties. Multiple studies have revealed that grifolin inhibits
cell growth in certain types of cancer (7,8). Grifolin
has been demonstrated to induce cell-cycle arrest via the ERK1/2
pathway in human nasopharyngeal carcinoma cells (7) and to induce apoptosis through inhibition
of the Akt pathway in human osteosarcoma cells (8). However, there is little research
concerning the effect of grifolin on ovarian cancer cells to date.
In the present study, A2780 ovarian cancer cells were treated with
different concentrations of grifolin in order to elucidate the
underlying signaling pathways and molecular mechanisms. The results
revealed that grifolin triggered apoptosis and cell cycle arrest in
a dose-and time-dependent fashion, and decreased the
phosphorylation of Akt and ERK1/2 in A2780 ovarian cancer cells
in vitro. The results indicated that grifolin may inhibit
cell viability and induce apoptosis in human ovarian cancer cells
via two pathways; the Akt and the ERK1/2 pathways.
Multiple studies have demonstrated that mammalian
cell cycle progression is closely associated with cyclins and CDKs
(9,10). Uncontrolled cell cycle progression is
an identified characteristic of multiple types of cancer (9). A previous study demonstrated that
grifolin significantly induced cell cycle arrest at the G1 phase in
human nasopharyngeal carcinoma cell line CNE1 (7). Verifying this finding, the data obtained
from flow cytometry analysis in the present study indicated that
grifolin induced G1 cell cycle arrest in A2780 cells. Thus,
attention was directed towards the underlying molecular mechanisms
of cell cycle arrest in A2780 cells treated with grifolin. CyclinD1
has been demonstrated to be a mitogenic sensor that amplifies
extracellular growth signals (10).
Overexpression of different cyclins and CDKs provides cancer cells
with the advantage of rapid cyclical growth (11). The cyclinD1/CDK4 complex promotes DNA
synthesis and cell cycle progression from the G1 to the S phase
(12). In accordance, downregulation
of cyclinD1 and CDK4 may be a promising treatment for cancer. As
expected, the present study demonstrated that grifolin treatment
resulted in a dose-dependent decrease in the expression of cyclinD1
and CDK4. Thus, the data obtained by flow cytometry were further
verified by the results of the western blot assay.
Members of the caspase family participate in
physiological activity through pathways including the mitochondrial
and death receptor pathway (13).
Activation of distinct caspase cascades is known to be involved in
activating and cleaving certain proteins that are crucial to
cellular activities and functions (14). In general, caspase-3 is recognized as
the key apoptotic mediator (15). A
previous study demonstrated that one of the caspase family
substrates, PARP, which possesses a wide range of physiological
functions, the most prominent of which is recovering damaged DNA
(16), is cleaved by caspase-3 into
two fragments (24 and 89 kDa, respectively), with different
functions. The 24 kDa PARP fragment weakens DNA repair by
inhibiting DNA repair enzymes, thereby contributing to cell death
(17). In the present study,
treatment with grifolin resulted in the upregulation of cleaved
caspase-3 and cleaved PARP in A2780 cells, further demonstrating
that the anti-cancer activity is regulated by caspase-dependent
cell apoptosis. As well as investigating the activation of
caspase-3 and PARP, the expression of Bcl-2 family members, which
have been identified as upstream regulators of the caspase cascade
(18,19), were assessed. It is possible to
subdivide the Bcl-2 protein family into pro-apoptotic components,
including Bax and Bcl-2 associated agonist of cell death, and
anti-apoptotic components, including Bcl-2 and B cell
lymphoma-extra large (20). The
present study demonstrated that Bcl-2 protein levels were
decreased, as demonstrated by western blotting, while the
expression of Bax was slightly altered. However, the Bcl-2/Bax
ratio was significantly lowered. It has been confirmed that the
Bcl-2/Bax ratio is crucial for regulating cellular apoptosis
(21). Alteration of this ratio may
increase the rate of apoptosis of A2780 cells exposed to
grifolin.
Cellular signaling pathways are known to include
diverse proteins, and are complex communication networks that
control basic biological functions. Several cellular signaling
pathways are known to regulate cell proliferation and apoptosis,
including nuclear factor-κB, PI3K/Akt, MAPK and p53 signaling
pathways. The aim of the present study was to define the pathways
participating in grifolin-induced anti-cancer effects. The majority
of previous studies have confirmed that activation of the ERK1/2 or
Akt pathway is associated with the anticancer effect of grifolin
(7,8)
Nevertheless, to date, knowledge concerning the effect of grifolin
in human ovarian cancer cells and underlying mechanisms is lacking.
Thus, the present study aimed to explore the signaling pathways
underlying the pro-apoptotic action of grifolin on A2780 cells.
Accumulating evidence indicates that the Akt pathway is a complex
signal transduction pathway participating in rigorous regulation of
cell differentiation, proliferation and apoptosis (22,23). Akt
phosphorylation promotes cell survival and proliferation, and
activation of Akt may contribute to tumorigenesis and tumor
progression (24). The results of the
present study demonstrated that the activation of Akt in A2780
cells was decreased by grifolin in a concentration-dependent
manner. MAPK members are classified into three major subfamilies:
p38MAP, ERK1/2 and c-Jun N-terminal Kinase, and phosphorylation of
ERK1/2 lead to progressing cell cycle and inhibiting cell apoptosis
(25). Furthermore, in the present
study, the phosphorylation level of ERK1/2 was evaluated and
demonstrated to be markedly decreased by grifolin. Hence, it is
possible to conclude that grifolin induced apoptosis and arrested
cell cycle progression at the G1 phase by downregulating
phosphorylated ERK1/2 and Akt levels, which resulted in altered
expression of caspase-3, Bax, CDK4 and cyclinD1.
In conclusion, grifolin, which has been known for
several decades to demonstrate various pharmacological and
physiological effects and to inhibit the growth of multiple cancer
cell lines, also had an anticancer effect against the human ovarian
cancer cell line, A2780. To the best of our knowledge, the present
study is the first report to explore the activity of grifolin on
human ovarian cancer cells. The results demonstrated that
inactivation of Akt and ERK1/2 was the mechanism underlying
grifolin-induced cell cycle arrest and cell apoptosis. Grifolin may
therefore be a promising antitumor agent for the treatment of
ovarian cancer.
Acknowledgements
The present study was funded by the National Natural
Science Foundation of China (grant nos. 81072121, 81372808 and
81173614), the Science and Technology Development Planning of
Shandong (grant nos. 2012G0021823 and 2011GSF12122) and the Science
and Technology Development Planning of Jinan (grant no.
201303035).
References
|
1
|
Gloss BS and Samimi G: Epigenetic
biomarkers in epithelial ovarian cancer. Cancer Lett. 342:257–263.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen C, Chang YC, Lan MS and Breslin M:
Leptin stimulates ovarian cancer cell growth and inhibits apoptosis
by increasing cyclin D1 and Mcl-1 expression via the activation of
the MEK/ERK1/2 and PI3K/Akt signaling pathways. Int J Oncol.
42:1113–1119. 2013.PubMed/NCBI
|
|
3
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jayson GC, Kohn EC, Kitchener HC and
Ledermann JA: Ovarian cancer. Lancet. 384:1376–1388. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luo X, Yu X, Liu S, Deng Q, Liu X, Peng S,
Li H, Liu J and Cao Y: The role of targeting kinase activity by
natural products in cancer chemoprevention and chemotherapy
(review). Oncol Rep. 34:547–554. 2015.PubMed/NCBI
|
|
7
|
Ye M, Luo X, Li L, Shi Y, Tan M, Weng X,
Li W, Liu J and Cao Y: Grifolin, a potential antitumor natural
product from the mushroom Albatrellus confluens, induces cell-cycle
arrest in G1 phase via the ERK1/2 pathway. Cancer Lett.
258:199–207. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jin S, Pang RP, Shen JN, Huang G, Wang J
and Zhou JG: Grifolin induces apoptosis via inhibition of PI3K/AKT
signalling pathway in human osteosarcoma cells. Apoptosis.
12:1317–1326. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hilakivi-Clarke L, Wang C, Kalil M,
Riggins R and Pestell RG: Nutritional modulation of the cell cycle
and breast cancer. Endocr Relat Cancer. 11:603–622. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hall M and Peters G: Genetic alterations
of cyclins, cyclin-dependent kinases, and Cdk inhibitors in human
cancer. Adv Cancer Res. 68:67–108. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kaukoniemi KM, Rauhala HE, Scaravilli M,
Latonen L, Annala M, Vessella RL, Nykter M, Tammela TL and
Visakorpi T: Epigenetically altered miR-193b targets cyclin D1 in
prostate cancer. Cancer Med. 4:1417–1425. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Denault JB and Boatright K: Apoptosis in
biochemistry and structural biology. 3–8 February 2004, Keystone,
CO, USA. IDrugs. 7:315–317. 2004.PubMed/NCBI
|
|
14
|
Gupta S, Kass GE, Szegezdi E and Joseph B:
The mitochondrial death pathway: A promising therapeutic target in
diseases. J Cell Mol Med. 13:1004–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lakhani SA, Masud A, Kuida K, Porter GA
Jr, Booth CJ, Mehal WZ, Inayat I and Flavell RA: Caspases 3 and 7:
Key mediators of mitochondrial events of apoptosis. Science.
311:847–851. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Langelier MF and Pascal JM: PARP-1
mechanism for coupling DNA damage detection to poly(ADP-ribose)
synthesis. Curr Opin Struct Biol. 23:134–143. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sukhanova MV, Khodyreva SN and Lavrik OI:
Influence of poly(ADP-ribose) polymerase-1 and its apoptotic 24-kD
fragment on repair of DNA duplexes in bovine testis nuclear
extract. Biochemistry (Mosc). 71:736–748. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Monian P and Jiang X: Clearing the final
hurdles to mitochondrial apoptosis: Regulation post cytochrome C
release. Exp Oncol. 34:185–191. 2012.PubMed/NCBI
|
|
19
|
Martinou JC and Youle RJ: Mitochondria in
apoptosis: Bcl-2 family members and mitochondrial dynamics. Dev
Cell. 21:92–101. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cotter TG: Apoptosis and cancer: The
genesis of a research field. Nat Rev Cancer. 9:501–507. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Antonsson B: Bax and other pro-apoptotic
Bcl-2 family ‘killer-proteins’ and their victim the mitochondrion.
Cell Tissue Res. 306:347–361. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Blanco-Aparicio C, Renner O, Leal JF and
Carnero A: PTEN, more than the AKT pathway. Carcinogenesis.
28:1379–1386. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fingar DC, Salama S, Tsou C, Harlow E and
Blenis J: Mammalian cell size is controlled by mTOR and its
downstream targets S6K1 and 4EBP1/elF4E. Genes Dev. 16:1472–1487.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Serrano ML, Sánchez-Gómez M, Bravo MM,
Yakar S and LeRoith D: Differential expression of IGF-I and insulin
receptor isoforms in HPV positive and negative human cervical
cancer cell lines. Horm Metab Res. 40:661–667. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Diab S, Kumarasiri M, Yu M, Teo T, Proud
C, Milne R and Wang S: MAP kinase-interacting kinases-emerging
targets against cancer. Chem Biol. 21:441–452. 2014. View Article : Google Scholar : PubMed/NCBI
|