Introduction
SET and MYND domain containing 3 (SMYD3) is a novel
histone methyltransferase gene identified in hepatoma and colon
carcinoma cells by Hamamoto et al (1). SMYD3 is located on human chromosome 1
and encodes two protein isoforms that are composed of 428 and 369
amino acids. Previous studies have demonstrated that SMYD3 is
frequently overexpressed in numerous types of cancer cells,
including hepatic, colon, gastric and cervical carcinoma, and
breast cancer (2–4), whilst the expression levels were lower
in the corresponding normal tissue. A number of previous studies
have demonstrated that SMYD3 has vital roles in the process of
tumor development via its functions as a histone methylation enzyme
and a transcription factor (5,6). SMYD3
modifies chromatin structure by catalyzing the methylation of
histone H3 at lysine 4 (H3K4), H4K20 and H4K5 (5,6). Also,
SMYD3 regulates the transcription of target genes via associating
with RNA polymerase II or HELZ RNA helicase and binding at the
motif CCCTCC or GGAGGG in the promoter (1).
MicroRNAs (miRNAs) are small, non-coding, endogenous
RNA molecules of 18–22 nucleotides that were first identified in
Caenorhabditis elegans. miRNAs suppress gene expression by
binding the targeted mRNA transcripts, which causes translational
repression or mRNA degradation. Previous studies demonstrated that
miRNAs serve important roles in tumorigenesis through the
regulation of genes involved in cancer development and maintenance
(7,8).
A number of studies have identified that histone
methylation and miRNAs are essential in the initiation and
progression of cancer (7–9). However, the association between SMYD3
and miRNAs is yet to be elucidated. To investigate this further,
the current study analyzed the global regulatory effects of SMYD3
on miRNAs in breast cancer cells using miRNA microarrays and
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR).
Materials and methods
Cell lines and plasmids
The MCF-7 human breast cancer cell line was obtained
from the American Type Culture Collection (Manassas, VA, USA). The
plasmid pcDNA5-TO/TAP-DEST-SMYD3 was a gift from Professor Philip
Tucker from the Institute for Cellular and Molecular Biology,
University of Texas (Austin, TX, USA). The CON049 (GeneChem, Co.,
Ltd., Shanghai, China) plasmid was used as a negative control for
short hairpin (sh)SMYD3 (GeneChem, Co., Ltd.). The SMYD3-ΔNHSC
plasmid was constructed by deleting the 205–208th amino acids
(Asn-His-Ser-Cys) of pcDNA5-TO/TAP-DEST-SMYD3. The miR-200c-3p
mimic and its negative control were obtained from Qiagen, Inc.,
(Valencia, CA, USA).
Cell culture and transfection
The MCF-7 cells were cultured in Dulbecco's modified
Eagle's medium (DMEM/F-12; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (Tianjin
Kangyuan Biotechnology Co., Ltd., Beijing, China) at 37°C in an
atmosphere containing 5% CO2. For plasmid transfection
experiments, MCF-7 cells were cultured in DMEM/F-12 medium without
added hormones at 60% confluence for 12 h, and then transfected
using TurboFect™ in vitro transfection reagent (Roche
Diagnostics, Indianapolis, IN, USA) according to the manufacturer's
protocol. Following incubation for 6 h, the medium was removed and
replaced with normal culture medium (DMEM/F-12 medium without
hormones) for 24 h. Then, the RT-qPCR was performed as described
below. For miR-200c-3p mimic transfection experiments,
Lipofectamine® 2000 reagents (Invitrogen; Thermo Fisher Scientific,
Inc.) were used following the manufacturer's protocol.
RNA extraction and complementary DNA
(cDNA) synthesis
Briefly, total cellular RNA was extracted from
cultured cells using TRIzol reagent according to the manufacturer's
protocol (Invitrogen; Thermo Fisher Scientific, Inc.) and 2 µg
total RNA was reverse-transcribed using M-MLV reverse transcriptase
(Promega Corporation, Madison, WI, USA) according to the
manufacturer's protocols. miRNA was isolated using the miRcute
miRNA isolation kit (#DP501; Tiangen Biotech, Co., Ltd., Beijing,
China), followed by cDNA synthesis using SuperScript First-Strand
Synthesis system (Invitrogen; Thermo Fisher Scientific, Inc.), but
with the specific stem loop primer primers. The miRNA RT-PCR primer
sequences were as follows: JH6-miR200c-3p,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTCCATC-3′; JH7-miR200c-3p,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTCCATCA-3′; JH7-miR149-3p,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGGGAGTG-3′. The total RNA
and dNTPs were incubated at 65°C for 5 min, cooled on ice briefly.
Subsequently, the miRNA specific primer, RNase inhibitor and
reverse transcriptase were added. After being incubated at 25°C for
10 min, 37°C for 50 min and 70°C for 15 min, the cDNA was
synthesized and used as a template of qPCR.
miRNA microarray chip analysis
miRNA expression profiling microarray was completed
using Agilent human miRNA (8×60K) V18.0 miRNA array (LC Sciences,
LLC., Houston, TX, USA). The microarray probe sequence was derived
from Sanger MiRBase version 15.0 (http://microrna.sanger.org). Each chip contained
multiple quality control probes and employed dual-color chip to
examine miRNA expression profiling in MCF-7 cells with
overexpressed or normal endogenous expression of SMYD3. Probes were
synthesized in situ with photosensitive photogenerated
reagents. The sequence consisted of two fragments: A chemically
modified oligonucleotide encoding fragment complementary to target
miRNA; and an extension arm at the distance specific to the
connected encoding sequence that reduced the hybridization spatial
impairment. Slides were scanned on an Agilent microarray scanner
(model G2565A; Agilent Technologies, Inc., Santa Clara, CA, USA) at
100 and 5% sensitivity settings. Agilent Feature Extraction
software version 8.1 (Agilent Technologies, Inc.) was used for
image analysis (10–12).
RT-qPCR of mRNA and miRNA
RT-qPCR was performed using a StepOne™ Real-Time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.).
Bestar® SYBR-Green qPCR Mastermix was obtained from DBI Bioscience
(http://www.xinghanbio.com/cpzs;
Shanghai, China). The thermal profiles were 95°C for 10 sec and
60°C for 1 min. Melting curve analysis was performed for each PCR
to confirm the specificity of amplification. At the end of each
phase, fluorescence was measured and quantified. Data is presented
as the relative expression level of mRNA following normalization to
GAPDH or the relative expression levels of miRNA following
normalization to U6 following calculations using the
2−ΔΔCq method (13). The
PCR primer sequences were as follows: GAPDH forward,
5′-ATTCAACGGCACAGTCAAGG-3′ and reverse, 5′-GCAGAAGGGGCGGAGATGA-3′;
zinc finger E-box binding homeobox (ZEB)1 forward,
5′-AAGGGCAAGAAATCCTGGGG-3′ and reverse, 5′-CTCTGGTCCTCTTCAGGTGC-3′;
ZEB2 forward, 5′-AAATGCACAGAGTGTGGCAAGG-3′ and reverse,
5′-CTGCTGATGTGCGAACTGTAGGA-3′; SMYD3 forward,
5′-AAGTTCGAACCGCCAAGAG-3′ and reverse, 5′-AAGGCAGCGGTCGCAGACGA-3′;
myosin light chain 9 (MYL9) forward, 5′-GAGCCCAAGCGCCTTCT-3′ and
reverse, 5′-GTCAATGAAGCCATCACGGT-3′; cysteine rich angiogenic
induced 61 (CYR61) forward, 5′-AAGGGGCTGGAATGCAACTT-3′ and reverse,
5′-TTGGGGACACAGAGGAATGC-3′; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′;
and reverse, 5′-AACGCTTCACGAATTTGCGT-3′; miR200c-3p forward,
5′-ACACTCCAGCTGGGTAATACTGCCGGGTAAT-3′; miR149-3p forward,
5′-ACACTCCAGCTGGGTCTGGCTCCGTGTCTTG-3′; and general reverse,
5′-TGGTGTCGTGGAGTCG-3′. All experiments were repeated three
times.
miRNA target gene prediction
In total 5 online prediction software programs,
including miRDB (http://mirdb.org/), TargetScan
(http://www.targetscan.org/),
DIANA-microT (http://diana.cslab.ece.ntua.gr/microT/), microRNA.org (http://www.microrna.org/microrna/home.do) and RNA22
(https://cm.jefferson.edu/rna22), were
used to predict the target genes of miRNAs. The intersection of 3/5
of these software programs were selected as the potential target
genes.
Statistical analysis
Statistical evaluations were performed using
GraphPad Prism (version 5.0; GraphPad Software, Inc., La Jolla, CA,
USA), using 3 independent experiments and were analyzed using
Student's t-test. P<0.05 was considered to indicate a
statistically significant difference. The homologous alignment
analysis was performed using DNAMAN (version 6.0; Lynnon LLC., San
Ramon, CA, USA) and Primer Premier (version 5.0; Premier Biosoft
International, Palo Alto, CA, USA).
Results
Effects of SMYD3 overexpression on the
miRNA expression profile of MCF-7 cells
To investigate the regulatory effects of SMYD3 on
global miRNA in breast cancer cells, MCF-7 cells were transfected
with SMYD3 and microarray analysis was performed using the Agilent
human miRNA (8×60K) V18.0 microarrays chip that contains 1,871
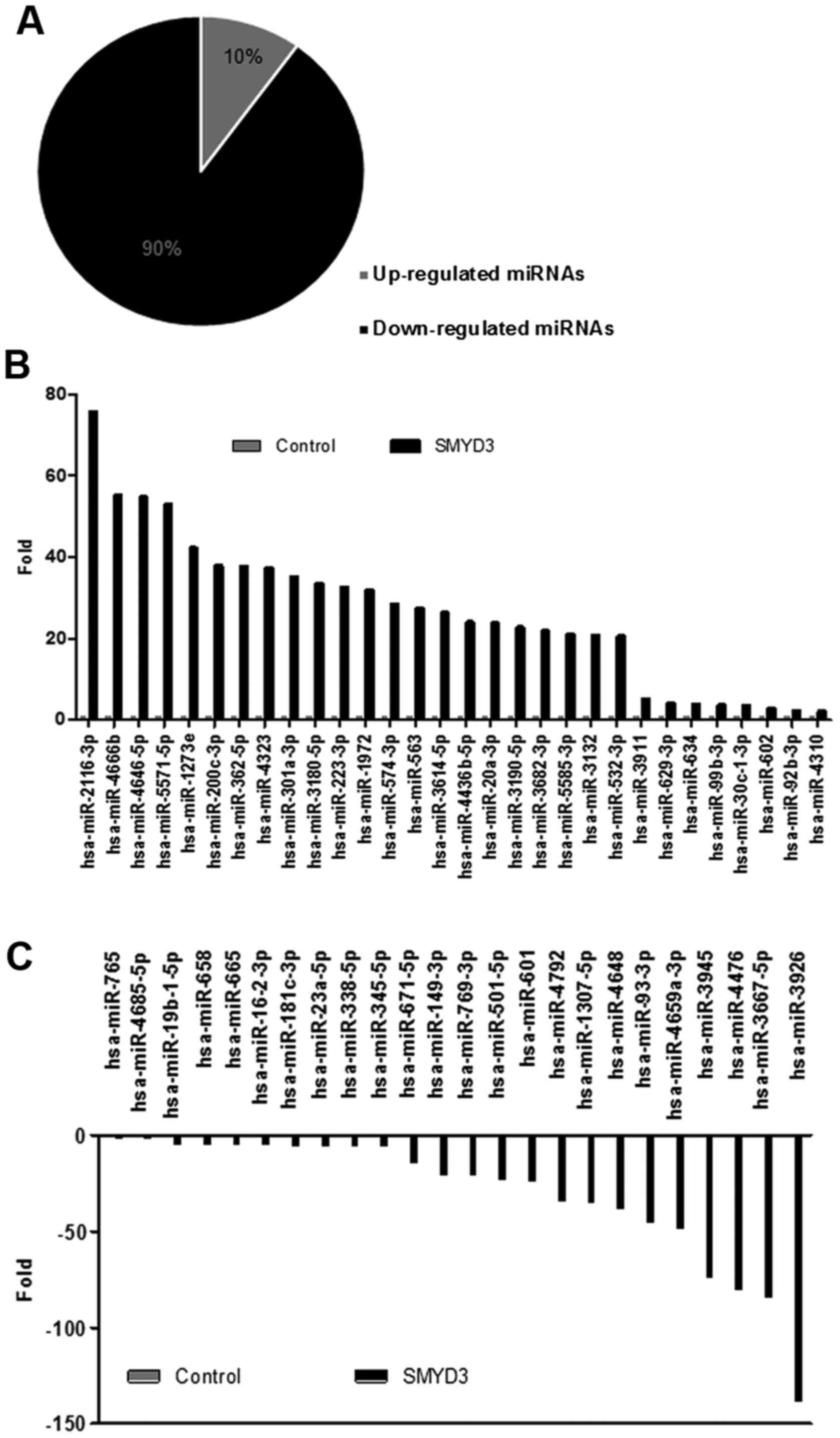
miRNAs. As presented in Fig. 1A, the
expression profile analysis identified 1,871 microRNAs with altered
expression, including 191 miRNAs that were upregulated in
SMYD3-transfected cells compared with mock transfected cells,
whilst 1,680 miRNAs were downregulated. Amongst these, 30 miRNAs
were upregulated >2.0-fold and 24 miRNAs were downregulated
<2.0-fold (Fig. 1B and C).
Furthermore, to investigate the function of these miRNAs in more
depth, the potential target genes of these miRNAs and their
functions were summarized using bioinformatic analysis (Table I) (14–57).
 | Table I.Functional classification of miRNAs
and their potential target genes altered by SMYD3 in MCF-7
cells. |
Table I.
Functional classification of miRNAs
and their potential target genes altered by SMYD3 in MCF-7
cells.
| miRNA | Fold change | Functions | Predicted target
genes |
|---|
| Upregulated
miRNAs |
| 1
hsa-miR-2116-3p | 75.68 | Carcinogenesis of
HCC | PEA15; ZDHHC11;
TNRC6B; CFLAR; MTCH2 |
| 2
hsa-miR-4666b | 55.23 | Prostate cancer
metastasis | VEZF1; EP300;
SLC5A12; USP46 |
| 3
hsa-miR-4646-5p | 54.83 | Oncogenic stress,
heat shock response and TLR pathway | CLIP3; SLC5A3;
TGFBR1 |
| 4
hsa-miR-5571-5p | 52.98 | Sjögren's
syndrome | PGM2L1; ATRN;
RAD23B; ZNF10; NFAT5 |
| 5
hsa-miR-1273e | 42.31 | Fas apoptotic
inhibitory, cell cycle, lysosomal/endosomal systems | ZC3H6; GAB1; GABPA;
PDK3; AAK1; KLF12 |
| 6
hsa-miR-200c-3p | 37.92 | Migration,
invasion, EMT and radiosensitivity in cancer cells | ZEB2; ZEB1; VASH2;
RECK; ERRFI1; CCNJ; LHFP; LOX; CDK17; CYP1B1; SESN1 |
| 7
hsa-miR-362-5p | 37.80 | Cell proliferation
and apoptosis resistance in gastric cancer and acral melanoma | LUC7L3; ACE2;
STRN3; CHSY1; SYS1 |
| 8
hsa-miR-4323 | 37.41 |
Dedifferentiation | CAMK2G; HDGF; AFF4;
ATXN7L3; PPP1R9B |
| 9
hsa-miR-301a-3p | 35.23 | Placenta growth
factor, NF-κB activator in pancreatic cancer cells and a T cells
activator in EAE | TSC1; DDX6; MIER1;
EIF2C4; FMR1; SLAIN1; MYBL1; ZFPM2; ENPP5 |
| 10
hsa-miR-3180-5p | 33.62 | Triple negative
breast cancer, ulcerative colitis | CDR1; KCNJ3;
AKIRIN1; ACTR2; SAMD5 |
| 11
hsa-miR-223-3p | 32.69 | Gastric cancer,
recurrent ovarian cancer, eosinophil maturation, IGF-1R signaling
and tumor suppression | FBXW7; SP3; FBXO8;
CRIM1; IL6ST; LMO2 |
| 12
hsa-miR-1972 | 31.95 | Lung cancer's early
diagnosis, enterovirus 71 (EV71) infection | SLC41A1; DOPEY1;
CYP27B1 |
| 13
hsa-miR-574-3p | 28.42 | Chondrogenesis | EP300; CUL2;
SAMD4A; MESDC1 |
| 14 hsa-miR-563 | 27.50 | Oxidative stress
response | COL1A2; RNF111;
SNX24; COL3A1 |
| 15
hsa-miR-3614-5p | 26.42 | Breast cancer
susceptibility | CPEB3; BBX; PTPRD;
NF2 |
| 16
hsa-miR-4436b-5p | 24.11 | Prion diseases and
transcriptional misregulation in cancer | RBM26; FAM53C;
NAV3 |
| 17
hsa-miR-20a-3p | 23.86 | Carcinogenesis of
gastric cancer and ovarian cancer | SLC38A2; RNF19A;
CADPS2; WDR47; CDH13 |
| 18
hsa-miR-3190-5p | 22.82 | Breast cancer and
irritable bowel syndrome susceptibility | ARID3B; ZNF704;
LIMD1; ZC3H7B |
| 19
hsa-miR-3682-3p | 21.96 | TLR pathway;
gastrointestinal cancer | ARMCX3; FXR1; MKL2;
SHISA9; EPC2; TRDMT1 |
| 20
hsa-miR-5585-3p | 21.16 | Cellular cycle,
apoptosis, stress responses | NRG3; IGF1;
ARHGAP5; XRCC4; MEX3B; RBM24 |
| 21
hsa-miR-3132 | 20.75 | Differential immune
responses following hantavirus infection | ICAM5; TMEM132E;
MTUS2; BLID |
| 22
hsa-miR-532-3p | 20.65 | Apoptosis
process | CYTH1; OTUD7B;
GSG1L; HECTD1; DTX4 |
| 23
hsa-miR-3911 | 5.13 | Oral
carcinogenesis | SSR1; BCOR; RAB6B;
SNTB2 |
| 24
hsa-miR-629-3p | 4.13 | Systemic lupus
erythematosus and lung cancer | TP53INP2; ADIPOR2;
MYOCD; ZNF322 |
| 25 hsa-miR-634 | 3.87 | Regulating
glioblastoma cell proliferation | PDIK1L; CERK;
TBC1D8; CRISPLD2 |
| 26
hsa-miR-99b-3p | 3.73 | AKT/mtor signaling
pathway | TMEM87A; RANBP6;
ZNF664 |
| 27
hsa-miR-30c-1-3p | 3.62 |
Doxorubicin-resistant in human breast
cancer cell | PCDHAC2; PCDHA6;
PCDHA4; PCDHA11 |
| 28 hsa-miR-602 | 2.91 | Acute rejection and
an early stage of HBV-mediated hepatocarcinogenesis | NOG; HTT; ORMDL1;
HABP4; CSK |
| 29
hsa-miR-92b-3p | 2.35 |
Hepatocarcinogenesis and G1/S phase of
cell cycle | CD69; SLC12A5;
FBXW7; MYO1B; DCAF6; RAB23; FNIP1; MAP2K4; RNF38; ACTC1 |
| 30
hsa-miR-4310 | 2.18 | Nitrate
transport | EPHA7; PLS3; ANO3;
ANKIB1; RWDD4; CAB39; KITLG; NOVA1 |
| Downregulated
miRNAs |
| 1
hsa-miR-765 | −2.24 | Squamous cell
carcinoma and traumatic brain injury and neural tube defects | PLP2; TIMP3;
ARID1A; CHRDL1; MYO1A |
| 2
hsa-miR-4685-5p | −2.31 | NF-κB signaling
pathway and TLR pathway | NHSL1; THPO;
DIAPH1; CHAC1; USP42; PPP3CB |
| 3
hsa-miR-19b-1-5p | −4.91 | 5-FU-resistant and
tumorigenicity | EDNRB; RBMS3;
PHF20 |
| 4
hsa-miR-658 | −4.95 | Acute rejection,
lupus nephritis and gastric cancer | RASD2; CNTNAP1;
DPYSL5; PPP1R16B |
| 5
hsa-miR-665 | −5.59 | DNA
hypermethylation and immune response; breast cancer, sporadic
amyotrophic lateral sclerosis and mesial temporal lobe
epilepsy | TGFBR1; GFI1B;
ALKBH5; PFKFB2; COPS7B |
| 6
hsa-miR-16-2-3p | −5.61 | P53 activation,
doxorubicin resistant, tumor cell proliferation and apoptosis,
polycythemia vera | RAB6A; INTU; SGIP1;
SP8 |
| 7
hsa-miR-181c-3p | −5.81 | Wnt/b-catenin
signaling, neuronal apoptosis, methylation | FAM122B; NHEJ1 |
| 8
hsa-miR-23a-5p | −5.87 | Glutamine
metabolism, EMT | PPA2; VSIG1; SF3B1;
FOXC1 |
| 9
hsa-miR-338-5p | −5.93 | HCC, androgen | PHC3; STAG2; SP3;
TNPO1; RAB28; BMI1 |
| 10
hsa-miR-345-5p | −6.14 | Multidrug
resistance, cell proliferation and invasion | SUV420H1;
CTTNBP2NL; RPA1; RFC1 |
| 11
hsa-miR-671-5p | −14.66 | Non-coding
antisense transcripts and gene silencing | LIN9; SLC30A6;
TBL2 |
| 12
hsa-miR-149-3p | −20.94 | EMT, proliferation,
metastasis and cell cycle | SMARCD; YBX2;
MS4A4A; UBE2Q1; SRRM2; CNGA2 |
| 13
hsa-miR-769-3p | −21.45 | Adenosine
deamination; Nervous system- related and tumor-related biological
processes and signaling pathways | PSMD14; GPC4;
TMEM139; GAP43 |
| 14
hsa-miR-501-5p | −23.13 | Apoptosis,
tumorigenesis | LPAR1; DNAJC6;
KIAA202; RNF165; PEX12; JMY |
| 15 hsa-miR-601 | −24.18 | The negative
regulation of translational initiation, ESCC and CRC | LHFPL2; SNN;
ZBTB38; SIRT1 |
| 16
hsa-miR-4792 | −34.45 | HIV infection and
infectious pancreatic necrosis virus | PCDH19; RNF214;
MAPK9; FRMPD4 |
| 17
hsa-miR-1307-5p | −35.72 | Metastasis | COL22A1; DPP4 |
| 18
hsa-miR-4648 | −38.77 | Brain specific | YWHAZ; SLC9A2;
BOLL; TNPO3 |
| 19
hsa-miR-93-3p | −46.09 | Tumor proliferation
and differentiation angiogenesis and metastasis | SYNJ1; AEBP2;
TARDBP; GLCCI1 |
| 20
hsa-miR-4659a-3p | −49.33 | Immune
response | ASCL1; CSNK1G1;
ZBTB39; KCND2; PRRX1 |
| 21
hsa-miR-3945 | −74.33 | Traumatic
pathogenesis, cerebrovascular disease | FNDC3B; RIC8B;
KBTBD2; BIRC6 |
| 22
hsa-miR-4476 | −80.76 | Myocardial
damage | POU2AF1; ATP6AP1;
C9ORF152; DPYSL2; LENG8; RANBP10 |
| 23
hsa-miR-3667-5p | −84.87 | Inflammation | RAP1A; CPEB3;
CMTM6; MYCBP; HOXD10 |
| 24
hsa-miR-3926 | −139.10 | TLR pathway and
prominin-1/CD133 | KCNG3; PIGA;
RBMXL1 |
Verifying the transcriptional
regulatory effect of SMYD3 on the target miRNAs, miR-200c-3p and
miR149-3p
To confirm the results of miRNA microarray,
miR-200c-3p, a predicted target that was upregulated by 37.9-fold,
and miR149-3p, a predicted target that was downregulated by
20.9-fold by SMYD3 in the chip assay, were selected for further
investigation. The pcDNA5-TO/TAP-DEST-SMYD3 plasmid and the shSMYD3
plasmid were transfected into MCF-7 cells for the overexpression
and knockdown of SMYD3, respectively. In accordance with the
results of miRNA microarray, the results of RT-qPCR identified that
miR-200c-3p was significantly upregulated following SMYD3
overexpression (P<0.001) and downregulated following RNA
interference (RNAi)-mediated suppression of SMYD3 (P<0.05;
Fig. 2A and B), whereas the level of
miR149-3p exhibited a significant negative association with the
expression of SMYD3 (P<0.001; Fig. 2C
and D).
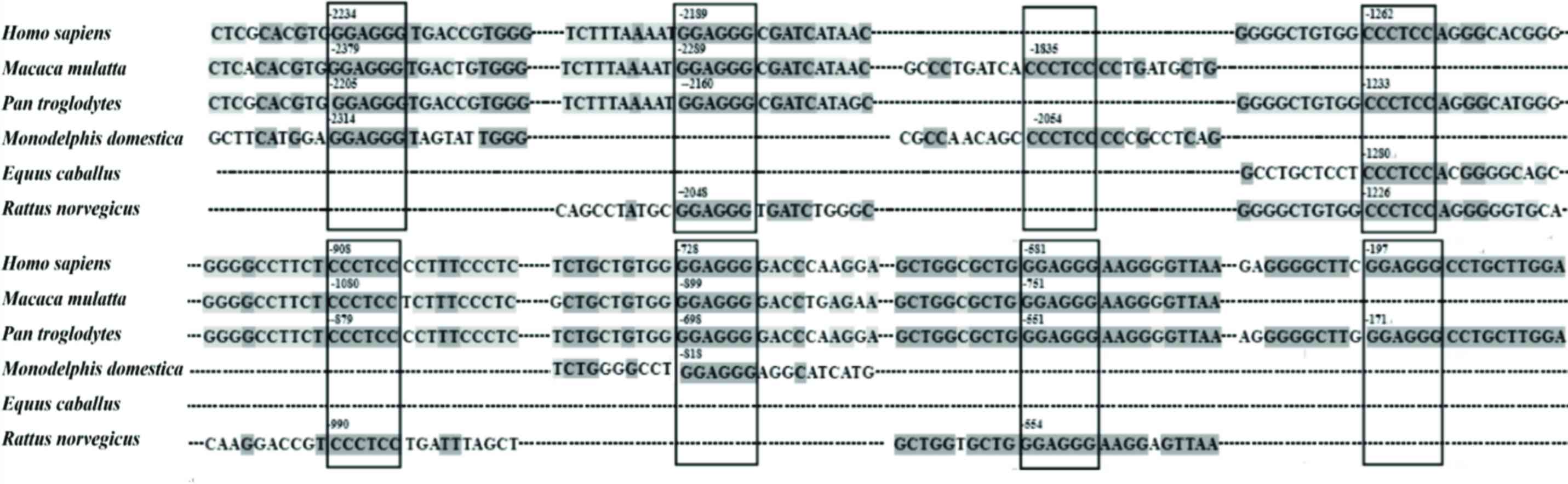
The promoter sequence of miR-200c-3p from a number
of species, including of Homo sapiens, Macaca
mulatta, Pan troglodytes, Monodelphis domestica,
Equus caballus and Rattus norvegicus, were analyzed
using bioinformatic methods. The results identified that 8
conserved SMYD3 binding sites exist in the promoter region (−2500
to +1) of miR-200c-3p from numerous species, indicating that
miR-200c-3p may be a target for the transcriptional regulation of
SMYD3 (Fig. 3).
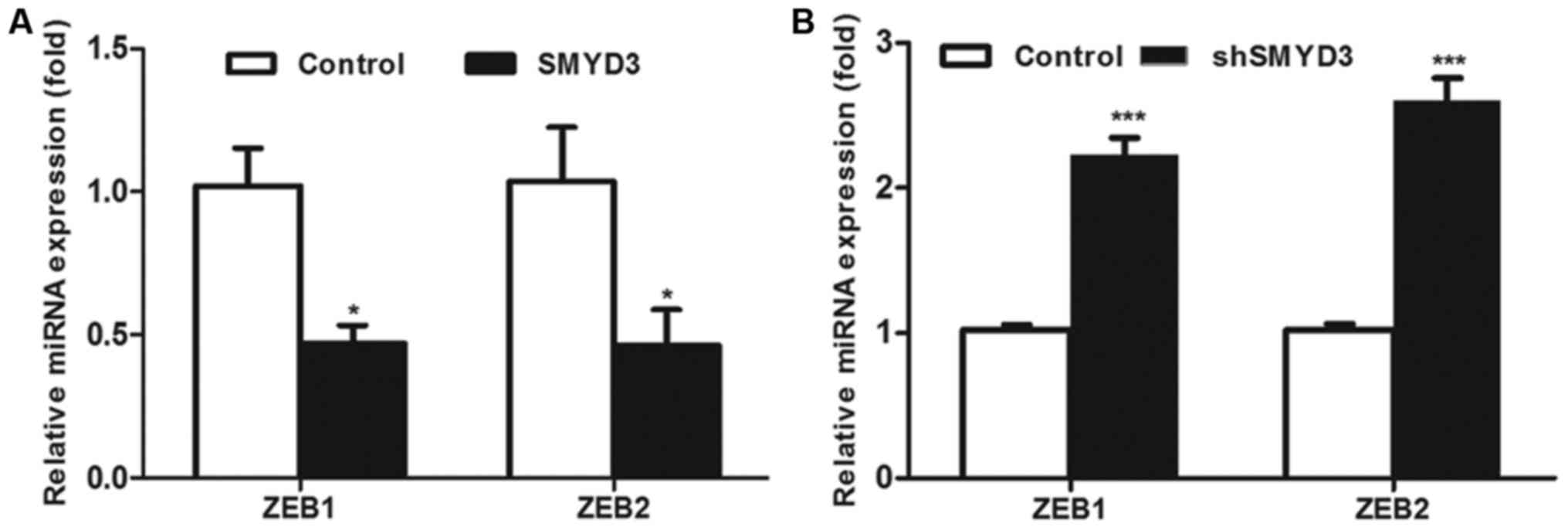
Furthermore, the results of RT-qPCR assay
demonstrated that the mRNA level of ZEB1 and ZEB2, two predicted
target genes of has-miR-200c-3p presented in Table I, was decreased by
SMYD3-overexpression, whereas they were increased by RNAi-induced
suppression of endogenous SMYD3 (Fig. 4A
and B). Taken together, these results demonstrate that SMYD3
activates the transcription of miR-200c-3p and therefore indirectly
decreases the mRNA levels of ZEB1/2.
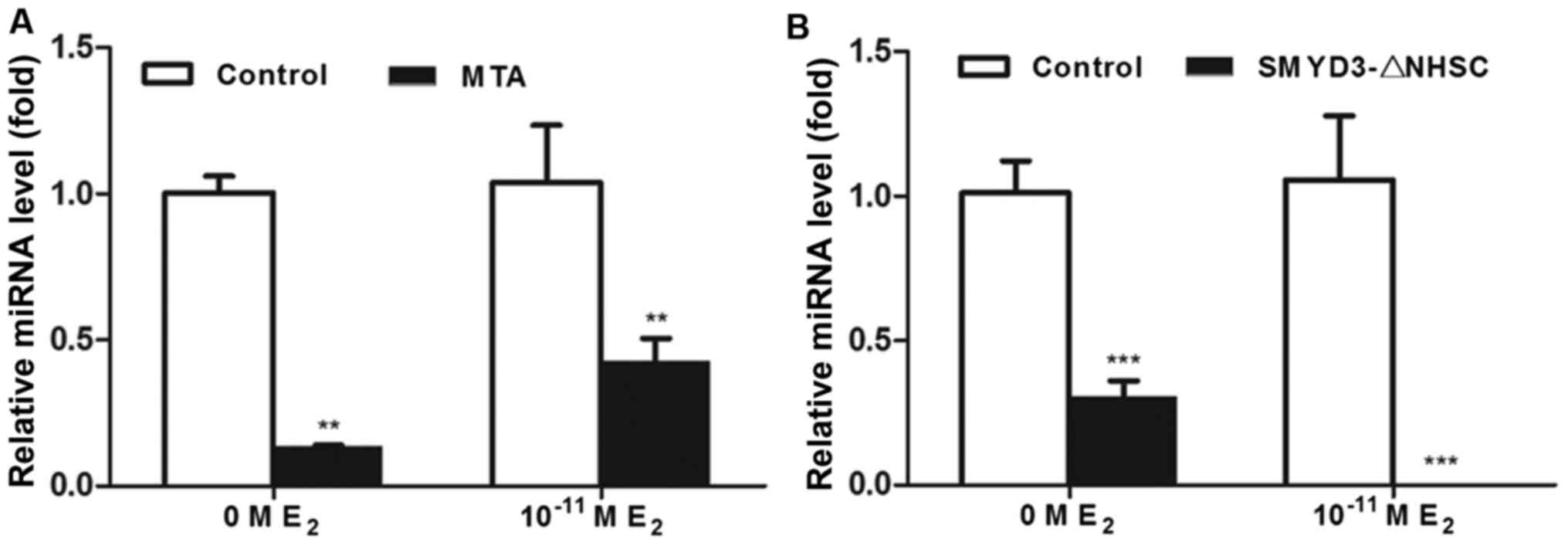
Histone methylation activity is
essential in the SMYD3-mediated transactivation of miR-200c-3p
Considering the results of the present study and the
previous studies that focusing on the histone methylation activity
of SMYD3, it was hypothesized that SMYD3 may function as a
transactivator of miR-200c-3p via its histone methylation function.
To investigate this, MCF-7 cells were treated with 100 µM
methylthioadenosine, a histone methylation inhibitor, with or
without physiological concentration of estradiol (10–11 M) and
subsequently the transcription level of miR-200c-3p was detected
using RT-qPCR. As presented in Fig.
5, the transcription level of miR-200c-3p was significantly
downregulated by the histone methylation inhibitor (Fig. 5A). Consistently, SMYD3-ΔNHSC, a
histone methyltransferase-activity-depleted mutant of SMYD3
(58,59), also decreases the transcription level
of miR-200c-3p (Fig. 5B). These
results suggested that the histone methylation activity may have
vital roles in SMYD3-mediated transactivation of miR-200c-3p.
Regulatory effects of SMYD3 and
miR-200c-3p on target genes of the RhoA-myocardin-related
transcription factor A (MRTF-A) signaling pathway
The authors' previous study demonstrated that SMYD3
promotes MRTF-A-mediated transactivation of MYL9 and migration of
MCF-7 breast cancer cells (59).
However, other previous studies have identified that miR-200c
represses migration and invasion of breast cancer cells via
suppression of the RhoA-MRTF-A signaling pathway (60,61). As
the results of the present study demonstrated the overexpression of
SMYD3 directly transactivates miR-200c-3p, whether SMYD3 and
miR-200c-3p supports or opposes each others' effects on the
transcription of MYL9 and CYR61, two MRTF-A-dependent
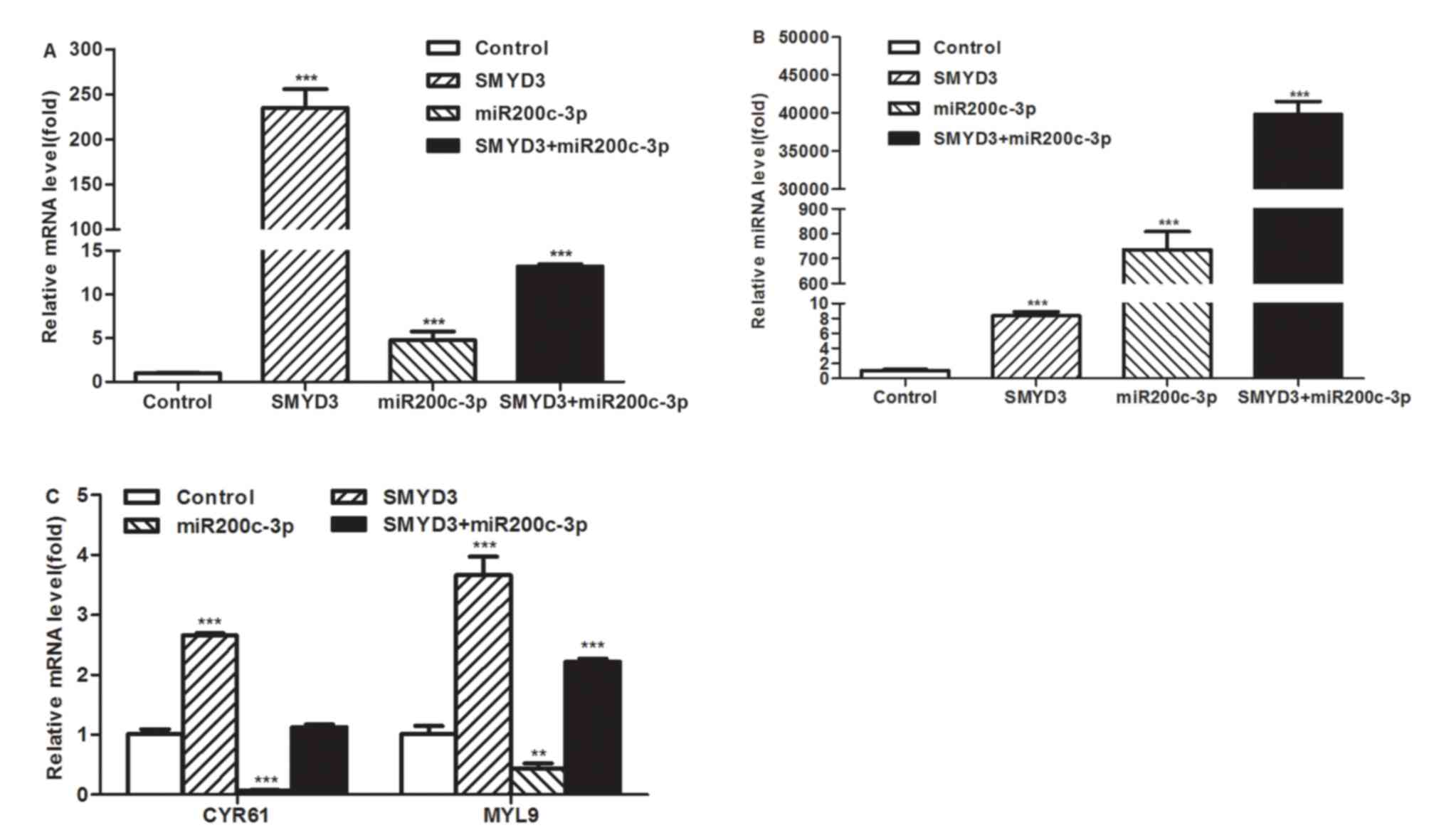
migration-related genes, was investigated. The miR-200c-3p mimic
and/or SMYD3-overexpression plasmid were transfected into MCF7
cells and the transcription level of MYL9 and CYR61 were detected
using RT-qPCR. The results of RT-qPCR demonstrated that the
transcriptional level of SMYD3 and the miR-200c-3p increased
significantly following the transfection (Fig. 6A and B). As presented in Fig. 6C, the mRNA levels of MYL9 and CYR61
were suppressed by miR-200c-3p and increased by the overexpression
of SMYD3. Furthermore, the upregulation of MYL9 and CYR61 remained
after SMYD3 was co-transfected with miR-200c-3p mimics, but the
effect in combination group was reduced compared with that of the
SMYD3-transfected group. These results demonstrated that
miR-200c-3p may be a downstream negative regulator of the
SMYD3-mediated pathway in the migration of breast cancer cells.
Discussion
The primary focus of epigenetics is to elucidate the
heritable changes in gene expression and regulation, which occur
without mutation to the DNA sequence, in the process of gene
transcription in eukaryotic cells. The heritable alterations
include DNA methylation, histone modification and regulation by
noncoding RNAs. A number of studies have established that the
majority of malignant tumor types exhibit cancer-associated
epigenetics alterations (62).
However, the interdisciplinary investigation of miRNA and histone
modification has received increasing interest.
A previous study has analyzed downstream coding
genes of SMYD3 using cDNA microarray and the results identified
that there were 61 genes upregulated >3-fold and genes
downregulated <3-fold by SMYD3 (1). However, to date, the downstream miRNAs
regulated by SMYD3 are yet to be elucidated. Therefore, the present
study focused on the effect of SMYD3 on the miRNA expression
profile in MCF-7 breast cancer cells by analyzing microarray and
RT-qPCR data. These results demonstrated that 30 miRNAs were
upregulated >2.0-fold, whilst 24 miRNAs were downregulated
<2.0-fold following the overexpression of SMYD3. The analysis of
bioinformatic and previously published data identified that the
majority of these potential downstream miRNAs of SMYD3 were
associated with the proliferation, migration and therapy resistance
of tumor.
To further validate the results of microarray,
RT-qPCR was performed to detect the effects of SMYD3 on the
expression of miR-200c-3p and miR-149, which are 2 representative
target miRNAs of SMYD3 with opposing alterations, and the results
were in accordance with those of the microarray. Previous studies
have demonstrated that miR-149 and miR-200c are associated with the
progression of cancer (63–65). miR-149 has been established as a tumor
suppressor by inhibiting the spreading, migration and invasion of
basal-like breast cancer cells (63).
However, the roles of miR-200c in cancer remain to be fully
understood. A number of previous studies have reported that
miR-200c may inhibit the epithelial-mesenchymal transition and
enhance the chemosensitivity and radiosensitivity of cancer cells
(64,65), whereas another previous study reported
a metastasis-promoting role of miR-200 in breast cancer (64). In addition, a separate study
demonstrated that the expression level of miR-200c different
between the subtypes of breast cancer (65). Compared with the normal breast
epithelial cells (MCF-10A), the basal cancer cells (MDA-MB-231 and
BT549) exhibited a reduced expression of miR-200c (65). However, the expression of miR-200c in
luminal cancer cells (MCF-7 and BT474) was increased compared with
that in the normal breast epithelial cells (65). Additionally, previous studies have
suggested that miR-200c may repress migration and invasion of
breast cancer cells via the suppression of RhoA-MRTF-A signaling
pathways (60,61), whereas the current study suggests that
SMYD3 promotes MRTF-A-mediated transactivation of MYL9 and
migration of MCF-7 breast cancer cells (59). The results in the present study have
demonstrated that the expression of miR-200c in MCF-7 breast cancer
cells may be upregulated by SMYD3, and the overexpression of
miR-200c partially inhibits the transactivation effects of SMYD3 on
the MRTF-A-dependent migration-associated genes. Taken together,
these results indicated that miR-200c may be a downstream negative
regulator of the SMYD3-mediated pathway in the migration of breast
cancer cells, and may promote a negative feedback loop to prevent
excessive induction of migration of breast cancer cells. However,
the underlying mechanism of miR-200c in breast cancer remains to be
elucidated by future investigation.
Additionally, previous studies have identified that
SMYD3 alters chromatin structure by catalyzing the
di-/tri-methylation of histone H3 at lysine 4 (H3K4me2/3), H4K20me3
and H4K5me (5,6). As part of the established methylation
marks at H3K4 or H3K36, mono-methylations of H3K9, H3K27, H3K79,
H4K20 and H2BK5 are associated with transcriptional activation,
whereas trimethylations of H3K9, H3K27, H3K79 and H4K20 are
associated with transcriptional repression (5,6). The
current study demonstrates that the histone methylation activity is
essential for SMYD3-mediated transactivation of miR-200c-3p.
Therefore, SMYD3 antagonistic regulation of the downstream miRNAs
may also depend on the distinct modification on the histone
adjacent to the promoter.
In conclusion, the effect of SMYD3 on the miRNA
expression profile in MCF-7 breast cancer cells was analyzed using
microarray and RT-qPCR in the present study. To the best of our
knowledge, this is the first study focused on the transcriptional
regulation of SMYD3 on miRNAs. These results may provide a novel
theoretical basis to elucidate the mechanism underlying the
initiation, progression, diagnose, prevention and therapy of breast
cancer.
Acknowledgements
The current study was supported by grants from the
National Natural Science Foundation of China (grant no. 31470816,
31300642 and 31000343), the College Students' Innovation and
Entrepreneurship Training Program of Tianjin (grant no.
201510057057) and the Program for Changjiang Scholars and
Innovative Research Team in University of Ministry of Education of
China (grant no. IRT1166).
References
|
1
|
Hamamoto R, Furukawa Y, Morita M, Iimura
Y, Silva FP, Li M, Yagyu R and Nakamura Y: SMYD3 encodes a histone
methyltransferase involved in the proliferation of cancer cells.
Nat Cell Biol. 6:731–740. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hamamoto R, Silva FP, Tsuge M, Nishidate
T, Katagiri T, Nakamura Y and Furukawa Y: Enhanced SMYD3 expression
is essential for the growth of breast cancer cells. Cancer Sci.
97:113–118. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang SZ, Luo XG, Shen J, Zou JN, Lu YH and
Xi T: Knockdown of SMYD3 by RNA interference inhibits cervical
carcinoma cell growth and invasion in vitro. BMB Rep. 41:294–299.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu Y, Luo X, Deng J, Pan Y, Zhang L and
Liang H: SMYD3 overexpression was a risk factor in the biological
behavior and prognosis of gastric carcinoma. Tumour Biol.
36:2685–2694. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Van Aller GS, Reynoird N, Barbash O,
Huddleston M, Liu S, Zmoos AF, McDevitt P, Sinnamon R, Le B, Mas G,
et al: Smyd3 regulates cancer cell phenotypes and catalyzes histone
H4 lysine 5 methylation. Epigenetics. 7:340–343. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Foreman KW, Brown M, Park F, Emtage S,
Harriss J, Das C, Zhu L, Crew A, Arnold L, Shaaban S and Tucker P:
Structural and functional profiling of the human histone
methyltransferase SMYD3. PloS One. 6:e222902011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yahya SM and Elsayed GH: A summary for
molecular regulations of miRNAs in breast cancer. Clin Biochem.
48:388–396. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang L, Deng L, Chen F, Yao Y, Wu B, Wei
L, Mo Q and Song Y: Inhibition of histone H3K79 methylation
selectively inhibits proliferation, self-renewal and metastatic
potential of breast cancer. Oncotarget. 5:10665–10677. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang H, Ach RA and Curry B: Direct and
sensitive miRNA profiling from low-input total RNA. RNA.
13:151–159. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hughes TR, Mao M, Jones AR, Burchard J,
Marton MJ, Shannon KW, Lefkowitz SM, Ziman M, Schelter JM, Meyer
MR, et al: Expression profiling using microarrays fabricated by an
ink-jet oligonucleotide synthesizer. Nature Biotechnol. 19:342–347.
2001. View Article : Google Scholar
|
|
12
|
Dennis G Jr, Sherman BT, Hosack DA, Yang
J, Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization and integrated discovery. Genome Biol. 4:P32003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Katayama Y, Maeda M, Miyaguchi K, Nemoto
S, Yasen M, Tanaka S, Mizushima H, Fukuoka Y, Arii S and Tanaka H:
Identification of pathogenesis-related microRNAs in hepatocellular
carcinoma by expression profiling. Oncology Lett. 4:817–823.
2012.
|
|
15
|
Watahiki A and Wang Y, Morris J, Dennis K,
O'Dwyer HM, Gleave M, Gout PW and Wang Y: MicroRNAs associated with
metastatic prostate cancer. PloS One. 6:e249502011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Moossavi S and Rezaei N: Toll-like
receptor signalling and their therapeutic targeting in colorectal
cancer. Int Immunopharmacol. 16:199–209. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Place RF and Noonan EJ: Non-coding RNAs
turn up the heat: An emerging layer of novel regulators in the
mammalian heat shock response. Cell Stress Chaperones. 19:159–172.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li J, Tan Q, Yan M, Liu L, Lin H, Zhao F,
Bao G, Kong H, Ge C, Zhang F, et al: miRNA-200c inhibits invasion
and metastasis of human non-small cell lung cancer by directly
targeting ubiquitin specific peptidase 25. Mol Cancer. 13:1662014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shi L, Fei X, Sun G, Wang Z, Wan Y, Zeng Y
and Guo J: Hypothermia stimulates glioma stem spheres to
spontaneously dedifferentiate adjacent non-stem glioma cells. Cell
Mol Neurobiol. 35:217–230. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mycko MP, Cichalewska M, Machlanska A,
Cwiklinska H, Mariasiewicz M and Selmaj KW: MicroRNA-301a
regulation of a T-helper 17 immune response controls autoimmune
demyelination. Proc Natl Acad Sci USA. 109:pp. E1248–E1257. 2012;
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Archanioti P, Gazouli M, Theodoropoulos G,
Vaiopoulou A and Nikiteas N: Micro-RNAs as regulators and possible
diagnostic bio-markers in inflammatory bowel disease. J Crohns
Colitis. 5:520–524. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li BS, Zhao YL, Guo G, Li W, Zhu ED, Luo
X, Mao XH, Zou QM, Yu PW, Zuo QF, et al: Plasma microRNAs, miR-223,
miR-21 and miR-218, as novel potential biomarkers for gastric
cancer detection. PLoS One. 7:e416292012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Madhavan D, Cuk K, Burwinkel B and Yang R:
Cancer diagnosis and prognosis decoded by blood-based circulating
microRNA signatures. Front Genet. 4:1162013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Guerit D, Philipot D, Chuchana P, Toupet
K, Brondello JM, Mathieu M, Jorgensen C and Noël D: Sox9-regulated
miRNA-574-3p inhibits chondrogenic differentiation of mesenchymal
stem cells. PLoS One. 8:e625822013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cheung O, Puri P, Eicken C, Contos MJ,
Mirshahi F, Maher JW, Kellum JM, Min H, Luketic VA and Sanyal AJ:
Nonalcoholic steatohepatitis is associated with altered hepatic
MicroRNA expression. Hepatology. 48:1810–1820. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marrale M, Albanese NN, Cali F and Romano
V: Assessing the impact of copy number variants on miRNA genes in
autism by Monte Carlo simulation. PLoS One. 9:e909472014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li X, Zhang Z, Yu M, Li L, Du G, Xiao W
and Yang H: Involvement of miR-20a in promoting gastric cancer
progression by targeting early growth response 2 (EGR2). Int J Mol
Sci. 14:16226–16239. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fan X, Liu Y, Jiang J, Ma Z, Wu H, Liu T,
Liu M, Li X and Tang H: miR-20a promotes proliferation and invasion
by targeting APP in human ovarian cancer cells. Acta Biochim
Biophys Sin (Shanghai). 42:318–324. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ivashchenko A, Berillo O, Pyrkova A,
Niyazova R and Atambayeva S: The properties of binding sites of
miR-619-5p, miR-5095, miR-5096, and miR-5585-3p in the mRNAs of
human genes. Biomed Res Int. 2014:7207152014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shin OS, Kumar M, Yanagihara R and Song
JW: Hantaviruses induce cell type- and viral species-specific host
microRNA expression signatures. Virology. 446:217–224. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cheng Y, Kuang W, Hao Y, Zhang D, Lei M,
Du L, Jiao H, Zhang X and Wang F: Downregulation of miR-27a* and
miR-532-5p and upregulation of miR-146a and miR-155 in LPS-induced
RAW264.7 macrophage cells. Inflammation. 35:1308–1313. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu MC, Lai NS, Chen HC, Yu HC, Huang KY,
Tung CH, Huang HB and Yu CL: Decreased microRNA(miR)-145 and
increased miR-224 expression in T cells from patients with systemic
lupus erythematosus involved in lupus immunopathogenesis. Clin Exp
Immunol. 171:91–99. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jeansonne D, Pacifici M, Lassak A, Reiss
K, Russo G, Zabaleta J and Peruzzi F: Differential effects of
microRNAs on glioblastoma growth and migration. Genes (Basel).
4:46–64. 2013.PubMed/NCBI
|
|
34
|
Jin Y, Tymen SD, Chen D, Fang ZJ, Zhao Y,
Dragas D, Dai Y, Marucha PT and Zhou X: MicroRNA-99 family targets
AKT/mTOR signaling pathway in dermal wound healing. PLoS One.
8:e644342013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fang Y, Shen H, Cao Y, Li H, Qin R, Chen
Q, Long L, Zhu XL, Xie CJ and Xu WL: Involvement of miR-30c in
resistance to doxorubicin by regulating YWHAZ in breast cancer
cells. Braz J Med Biol Res. 47:60–69. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sengupta S, Nie J, Wagner RJ, Yang C,
Stewart R and Thomson JA: MicroRNA 92b controls the G1/S checkpoint
gene p57 in human embryonic stem cells. Stem Cells. 27:1524–1528.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Balakathiresan N, Bhomia M, Chandran R,
Chavko M, McCarron RM and Maheshwari RK: MicroRNA let-7i is a
promising serum biomarker for blast-induced traumatic brain injury.
J Neurotrauma. 29:1379–1387. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kurokawa K, Tanahashi T, Iima T, Yamamoto
Y, Akaike Y, Nishida K, Masuda K, Kuwano Y, Murakami Y, Fukushima M
and Rokutan K: Role of miR-19b and its target mRNAs in
5-fluorouracil resistance in colon cancer cells. J Gastroenterol.
47:883–895. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hackl M, Brunner S, Fortschegger K,
Schreiner C, Micutkova L, Mück C, Laschober GT, Lepperdinger G,
Sampson N, Berger P, et al: miR-17, miR-19b, miR-20a, and miR-106a
are down-regulated in human aging. Aging Cell. 9:291–296. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sui W, Dai Y, Huang Y, Lan H, Yan Q and
Huang H: Microarray analysis of MicroRNA expression in acute
rejection after renal transplantation. Transpl Immunol. 19:81–85.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tsuruta T, Kozaki K, Uesugi A, Furuta M,
Hirasawa A, Imoto I, Susumu N, Aoki D and Inazawa J: miR-152 is a
tumor suppressor microRNA that is silenced by DNA hypermethylation
in endometrial cancer. Cancer Res. 71:6450–6462. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lezina L, Purmessur N, Antonov AV, Ivanova
T, Karpova E, Krishan K, Ivan M, Aksenova V, Tentler D, Garabadgiu
AV, et al: miR-16 and miR-26a target checkpoint kinases Wee1 and
Chk1 in response to p53 activation by genotoxic stress. Cell Death
Dis. 4:e9532013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ciafre SA, Galardi S, Mangiola A, Ferracin
M, Liu CG, Sabatino G, Negrini M, Maira G, Croce CM and Farace MG:
Extensive modulation of a set of microRNAs in primary glioblastoma.
Biochem Biophys Res Commun. 334:1351–1358. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rathore MG, Saumet A, Rossi JF, de
Bettignies C, Tempé D, Lecellier CH and Villalba M: The NF-κB
member p65 controls glutamine metabolism through miR-23a. Int J
Biochem Cell Biol. 44:1448–1456. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Huang XH, Wang Q, Chen JS, Fu XH, Chen XL,
Chen LZ, Li W, Bi J, Zhang LJ, Fu Q, et al: Bead-based microarray
analysis of microRNA expression in hepatocellular carcinoma:
miR-338 is downregulated. Hepatol Res. 39:786–794. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Schou JV, Rossi S, Jensen BV, Nielsen DL,
Pfeiffer P, Høgdall E, Yilmaz M, Tejpar S, Delorenzi M, Kruhøffer M
and Johansen JS: miR-345 in metastatic colorectal cancer: A
non-invasive biomarker for clinical outcome in non-KRAS mutant
patients treated with 3rd line cetuximab and irinotecan. PloS One.
9:e998862014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rossi JJ: A novel nuclear miRNA mediated
modulation of a non-coding antisense RNA and its cognate sense
coding mRNA. EMBO J. 30:4340–4341. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ke Y, Zhao W, Xiong J and Cao R: miR-149
inhibits non-small-cell lung cancer cells EMT by targeting FOXM1.
Biochem Res Int. 2013:5067312013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Liu F, Xiong Y, Zhao Y, Tao L, Zhang Z,
Zhang H, Liu Y, Feng G, Li B, He L, et al: Identification of
aberrant microRNA expression pattern in pediatric gliomas by
microarray. Diagn Pathol. 8:1582013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sun Y, Wu J, Wu SH, Thakur A, Bollig A,
Huang Y and Liao DJ: Expression profile of microRNAs in c-Myc
induced mouse mammary tumors. Breast Cancer Res Treat. 118:185–196.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ohdaira H, Nakagawa H and Yoshida K:
Profiling of molecular pathways regulated by microRNA 601. Comput
Biol Chem. 33:429–433. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Chang ST, Thomas MJ, Sova P, Green RR,
Palermo RE and Katze MG: Next-generation sequencing of small RNAs
from HIV-infected cells identifies phased microrna expression
patterns and candidate novel microRNAs differentially expressed
upon infection. MBio. 4:e00549–e00512. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Du L, Schageman JJ, Subauste MC, Saber B,
Hammond SM, Prudkin L, Wistuba II, Ji L, Roth JA, Minna JD and
Pertsemlidis A: miR-93, miR-98, and miR-197 regulate expression of
tumor suppressor gene FUS1. Mol Cancer Res. 7:1234–1243. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Alashti F Asghari and Minuchehr Z: MiRNAs
which target CD3 subunits could be potential biomarkers for
cancers. PLoS One. 8:e787902013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yang J, Liang Y, Han H and Qin H:
Identification of a miRNA signature in neutrophils after traumatic
injury. Acta Biochim Biophys Sin (Shanghai). 45:938–945. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Rustagi Y and Rani V: Screening of
microRNA as potential CardiomiRs in Rattus noveregicus heart
related dataset. Bioinformation. 9:919–922. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Rappa G, Mercapide J, Anzanello F, Pope RM
and Lorico A: Biochemical and biological characterization of
exosomes containing prominin-1/CD133. Mol Cancer. 12:622013.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kim H, Heo K, Kim JH, Kim K, Choi J and An
W: Requirement of histone methyltransferase SMYD3 for estrogen
receptor-mediated transcription. J Biol Chem. 284:19867–19877.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Luo XG, Zhang CL, Zhao WW, Liu ZP, Liu L,
Mu A, Guo S, Wang N, Zhou H and Zhang TC: Histone methyltransferase
SMYD3 promotes MRTF-A-mediated transactivation of MYL9 and
migration of MCF-7 breast cancer cells. Cancer Lett. 344:129–137.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Jurmeister S, Baumann M, Balwierz A,
Keklikoglou I, Ward A, Uhlmann S, Zhang JD, Wiemann S and Sahin Ö:
MicroRNA-200c represses migration and invasion of breast cancer
cells by targeting actin-regulatory proteins FHOD1 and PPM1F. Mol
Cell Biol. 32:633–651. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Wu XD, Liu WL, Zeng K, Lei HY, Zhang QG,
Zhou SQ and Xu SY: Advanced glycation end products activate the
miRNA/RhoA/ROCK2 pathway in endothelial cells. Microcirculation.
21:178–186. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Cava C, Bertoli G and Castiglioni I:
Integrating genetics and epigenetics in breast cancer: Biological
insights, experimental, computational methods and therapeutic
potential. BMC Syst Biol. 9:622015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Bischoff A, Huck B, Keller B, Strotbek M,
Schmid S, Boerries M, Busch H, Müller D and Olayioye MA: miR149
functions as a tumor suppressor by controlling breast epithelial
cell migration and invasion. Cancer Res. 74:5256–5265. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zhang HF, Xu LY and Li EM: A family of
pleiotropically acting microRNAs in cancer progression, miR-200:
Potential cancer therapeutic targets. Curr Pharm Des. 20:1896–1903.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Sun Q, Liu T, Yuan Y, Guo Z, Xie G, Du S,
Lin X, Xu Z, Liu M, Wang W, et al: MiR-200c inhibits autophagy and
enhances radiosensitivity in breast cancer cells by targeting
UBQLN1. Int J Cancer. 136:1003–1012. 2015. View Article : Google Scholar : PubMed/NCBI
|