Introduction
Neuroglioma is a primary nervous system tumor with a
high rate of malignancy (1). Patients
that receive surgical treatment for this disease typically have a
poor prognosis, particularly those with glioblastoma multiforme
(GBM), for which the average survival time is <2 years (2). According to a previous study (3), the occurrence and progression of glioma
is associated with the abnormal expression of oncogenes and tumor
suppressor genes. However, the molecular mechanisms underlying
glioma development remain unclear (4). Investigating the association between
changes in gene function and the occurrence and malignancy of
glioma may aid in improving understanding of the molecular
mechanisms underlying glioma development, and therefore the
development of effective drugs to improve the survival of patients
with this disease (5).
Long non-coding RNA (lncRNA) is a type of RNA that
does not code for proteins and is >200 nucleotides in length
(6,7).
LncRNAs regulate gene expression on an epigenetic, transcriptional
and post-transcriptional level (8,9). The modes
by which lncRNAs regulate gene expression include chromosome
modification and transcriptional activation or interference
(10). Differential expression of
lncRNA has been demonstrated between normal and cancerous tissues,
such as lung cancer (11), colorectal
cancer (12), breast cancer (13) and glioma (14), suggesting that abnormal lncRNA
expression serves a role in tumor occurrence (15). LncRNA is a potential biomarker for
predicting the outcomes of treatment and patient prognosis, and may
be a target for gene therapy (16).
Fer-1-like family member 4 (FER1L4), a lncRNA, has
been associated with tumor occurrence and progression, particularly
in gastric and colon cancer (17,18). In
the present study, data from The Cancer Genome Atlas (TCGA) was
mined in order to investigate the association between FER1L4
expression and the prognosis of patients with glioma. Reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis was also performed to study the differential expression of
FER1L4 in glioma cell lines and normal astrocytes. The results of
the present study indicate that FER1L4 is a predictor of poor
prognosis in glioma, and serves an important role in the occurrence
and progression of glioma.
Materials and methods
Analysis of clinical data
The clinical data and gene array data for glioma in
149 patients with GBM, 158 patients with World Health Organization
(WHO) II grade glioma and 177 patients with WHO III grade glioma
was obtained from the TCGA database (19) using the platform
illuminahiseq_rnaseqv2. All glioma samples were histologically
graded in accordance with the 2007 WHO classification of tumors of
the central nervous system (2). TCGA
gene expression data for FER1L4 were extracted and merged with
clinical outcome data for further analysis. The predictive value of
FER1L4 on survival was examined, and FER1L4 expression acted as a
continuous measurement in addition to a categorical variable. A
total of 484 patients were divided into the following two groups
according to FER1L4 expression with the median as the cutoff: High
expression group (242 patients) and low expression group (242
patients).
Cell culture
Neuroglioma cell lines U373-MG, U251, LN-18, U87-MG
and SHG-44 and the normal astrocyte cell line NHA-1800 were
purchased from the Shanghai Institutes for Biological Sciences,
Chinese Academy of Sciences (Shanghai, China). Fetal bovine serum
(FBS), 0.25% trypsin and Dulbecco's modified Eagle's medium (DMEM)
were purchased from Invitrogen (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). Other consumables for cell culture were
purchased from BD Biosciences (Franklin Lakes, NJ, USA). The
neuroglioma cell lines and the normal astrocyte cell line NHA-1800
were cultured in DMEM containing 10% FBS and 1% myllicin at 37°C
with 5% CO2 in a humidified incubator. Cell passage was
performed when the cells had grown to 80% confluence.
RT-qPCR
Total RNA extraction was performed using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), and RNA
content was quantified using a UV-spectrophotometer (20). RNA was reverse transcribed to
complementary (c)DNA using a RevertAid™ First Strand cDNA synthesis
kit (Fermentas; Thermo Fisher Scientific, Inc.) and qPCR was
performed. The primers were synthesized by Shanghai Invitrogen
Biotechnology Co., Ltd. (Shanghai, China), and the sequences were
as follows: FER1L4 forward, 5′CCG TGT TGA GGT GCT GTT C-3′ and
reverse, 5′GGC AAG TCC ACT GTC AGA TG3′; GAPDH forward, 5′AAG GTG
AAG GTC GGA GTC AA-3′ and reverse, 5′AAT GAA GGG GTC ATT GAT GG-3′.
The reaction components were as follows: DreamTaq™ Green PCR Master
Mix (2X) 10 µl, 2 µl PCR Forward Primer, 2 µl PCR Reverse Primer,
cDNA 2 µl, ddH2O 4 µl. The thermocycling conditions were as
follows: Denaturation at 95°C for 2 min; 35 cycles of 95°C for 30
sec, 57.4°C for 30 sec, 72°C for 30 sec; and extension at 72°C for
10 min. The PCR products were analyzed by 2% agarose gel
electrophoresis. Gels were visualized by Gel Doc™XR gel
documentation system (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The 2−ΔΔCq method was used for the quantification
of real-time RT-PCR gene products (21).
Transfection
A day prior to transfection, cells were seeded into
a 6-well plate at a concentration of 4×105 cells/well. A
total of 2 ml DMEM containing 10% FBS was added, and the cells were
cultured 37°C with 5% CO2 in the incubator overnight.
Transfection was performed when the cells reached 70–80%
confluence. For each well, 10 µl Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) was added for every 4 µg siRNA. For
the negative control, the transfection was performed with a control
siRNA, and serum-free medium was added. Culture medium was replaced
6 h following transfection. The short interfering (si)RNAs were
provided by Shanghai Invitrogen Biotechnology Co., Ltd. (Shanghai,
China). The siRNA sequences were as follows: FER1L4,
5′-CAGGACAGCUUCGAGUUAATT-3′ for the forward, and control,
5′-UUCUCCGAACGUGUCACGUTT-3′ for the forward.
Cell counting kit (CCK)-8 assay
The CCK-8 assay kit was purchased from Dojindo
Molecular Technologies, Inc. (Kumamoto, Japan). Following
transfection, logarithmic growth phase cells were harvested and
seeded into a 96-well plate at a density of 5,000 cells/well, with
3 replicates for each group. At 24, 48, 72 and 96 h following
transfection, 10 µl CCK-8 solution was added into each well. The
cells were then cultured at 37°C for 2 h. Absorbance was measured
for each well using a microplate reader at 450 nm, and the growth
status of cells in each group was compared.
Transwell assay
Matrigel-coated transwell cell culture chambers (8
µm pore size) was purchased from EMD Millipore (Billerica, MA,
USA). The Transwell plate was coated with Matrigel® (BD
Biosciences) and placed into the incubator at 37°C to dry the
basement membrane for 24 h. Following transfection, a single-cell
suspension (5×104) was added drop wise to the upper
chamber and the cells were cultured for 24 h at 37°C, cells on the
upper surface of the filters were removed.
The filter membrane was fixed in 4%
paraformaldehyde, and stained with Coomassie blue (1 mg/ml). The
degree of invasion was quantified by counting the cells that
migrated to the lower side of the filter in ≥5 random fields using
a light microscope.
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) apoptosis assay
An Annexin V-FITC/PI apoptosis kit was purchased
from Nanjing KeyGen Biotech Co., Ltd (Nanjing, China). When the
siRNA transfection cells reached 80% confluence following
transfection they were digested with 0.25% trypsin and washed with
pre-cooled PBS three times. The cells were suspended in 400 µl
binding buffer, and 5 µl Annexin V-FITC was added into the cell
suspension. Following gentle agitation, the cells were incubated at
2–8°C in the dark for 15 min. A total of 10 µl PI was added, and
the cells were incubated at 2–8°C in the dark for a further 5 min.
The cells were subsequently detected using a FACSCalibur (BD
Biosciences, Franklin Lakes, NJ, USA) flow cytometer within 1 h and
observed under a fluorescence microscope. The apoptotic analysis
was performed using FlowJo (Tree Star, Inc., Ashland, OR, USA).
Interpretation criteria were as follows:
FITC−/PI− was defined as live cells in the
lower left quadrant; FITC+/PI− was defined as
late apoptotic cells in the upper left quadrant;
FITC+/PI+ was defined as apoptotic cells in
the upper right quadrant; FITC+/PI− was
defined as apoptotic cells in the lower right quadrant.
Statistical analysis
The results are presented as mean ± standard
deviation. Fisher's exact test and χ2 were used to
compare clinicopathological data. One-way analysis of variance and
two-sample t-tests were used to analysis the difference in lncRNA
FER1L4 expression levels of different grade glioma tissues and four
cell lines. The Kaplan-Meier estimator method was used to estimate
overall survival, and a log-rank test was used to examine the
differences between clinical characteristics and survival in
patients with glioma. Analyses were performed using GraphPad Prism
software (version 5; GraphPad Software, Inc., La Jolla, CA, USA),
P-values less than 0.05 were regarded as statistically
significant.
Results
High expression of FER1L4 predicts a
poor prognosis in patients with glioma
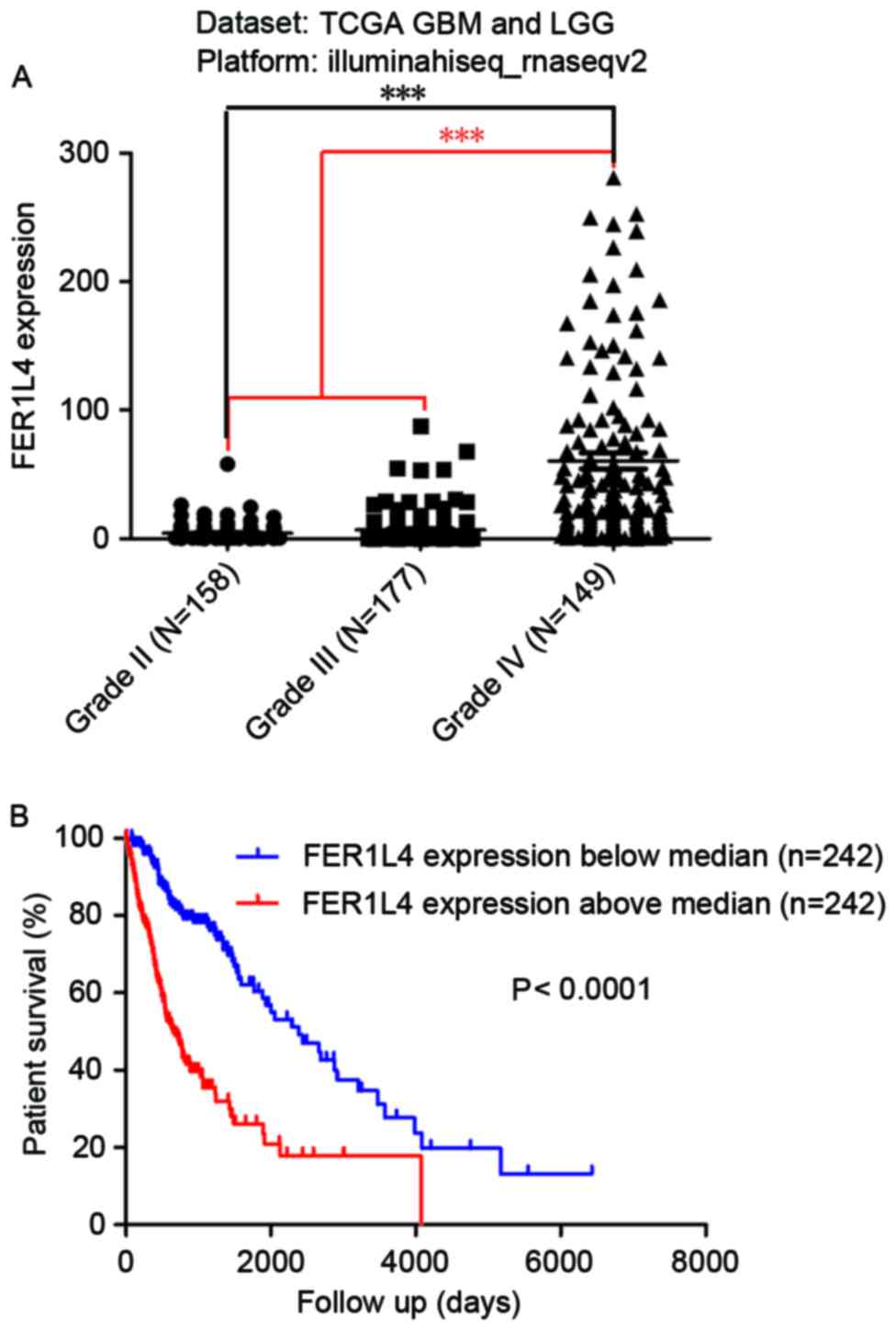
To study the association between FER1L4 and glioma,
the expression of FER1L4 in 484 patients with either GBM or LGG was
analyzed by mining the TCGA database. The expression of FER1L4 in
patients with grade IV glioma was 60.57±6.282 (N=149), which was
significantly higher compared with that in patients with LGG
(II–III; 5.754±0.5195; N=335) (***P<0.0001; Fig. 1A). In addition, the expression of
FER1L4 in patients with grade III glioma (6.896±0.8737; N=177) was
significantly higher compared with that in patients with grade II
glioma (4.475±0.489; N=158; ***P<0.0001). Survival analysis
indicated that the prognosis of patients with high FER1L4
expression was significantly worse compared with those with low
FER1L4 expression (P<0.0001; Fig.
1B).
FER1L4 is highly expressed in glioma
cell lines in vitro
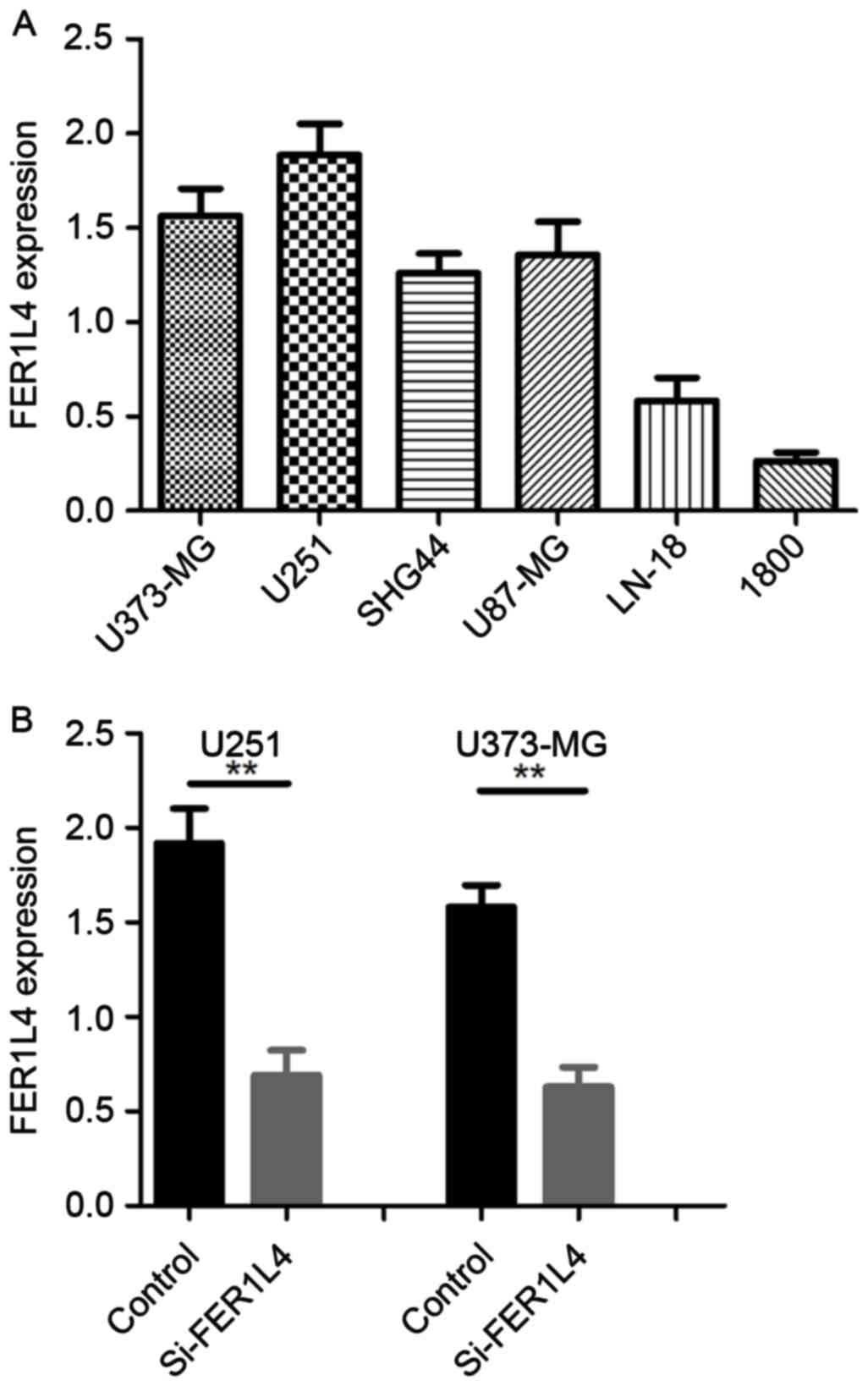
RT-qPCR was performed to characterize the expression
of FER1L4 in the glioma cell lines U373-MG, U251, U87-MG, SHG-44,
LN-18 and in the normal astrocyte cell line NHA-1800. The results
demonstrated that FER1L4 expression was higher in all the glioma
cell lines studied compared with the normal astrocyte cells
(Fig. 2A). The glioma cell lines
U373-UG and U251 exhibited the highest FER1L4 expression (Fig. 2A).
Expression of FER1L4 is decreased
following transfection with siRNA
In order to investigate the biological function of
FER1L4 in glioma cells, U373-MG and U251 cells were transfected
with siRNA directed against FER1L4. At 48 h following transfection,
FER1L4 expression was significantly downregulated in
si-FER1L4-treated cells compared with the control cells (treated
with control siRNA) (Fig. 2B).
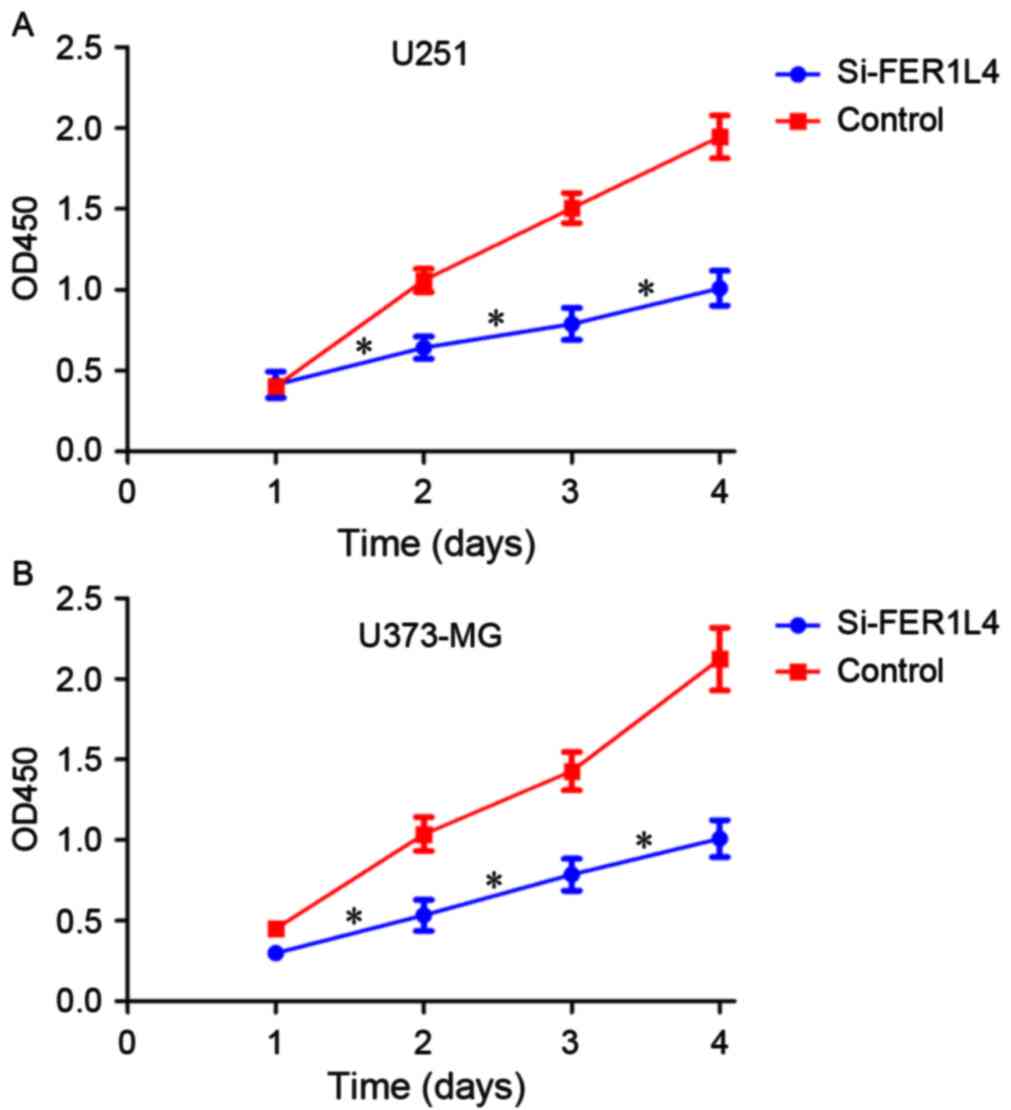
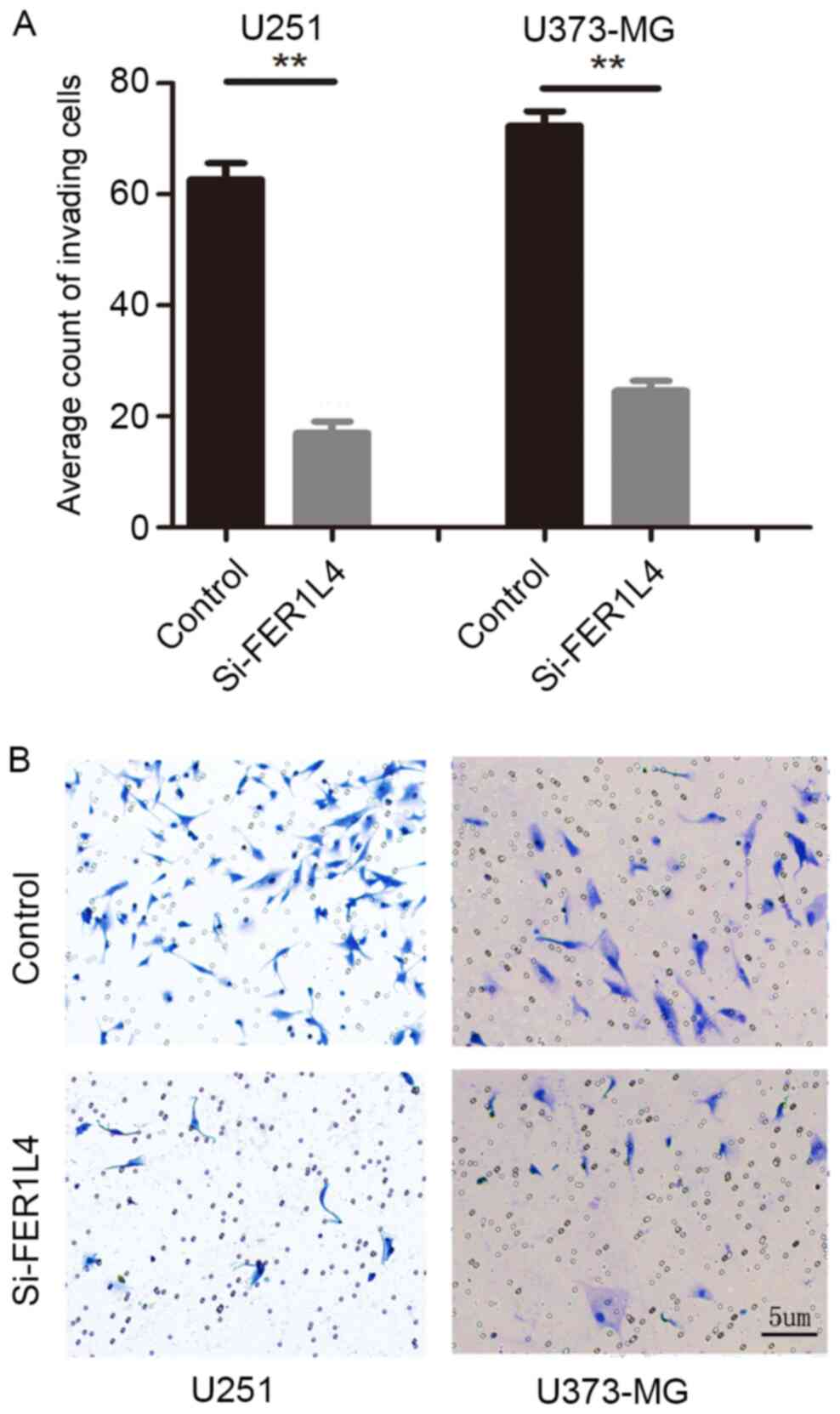
Detection of cell proliferation,
apoptosis and invasion
CCK-8 and Transwell assays were performed following
transfection with FER1L4 siRNA in order to investigate the
association between FER1L4 expression and glioma viability and
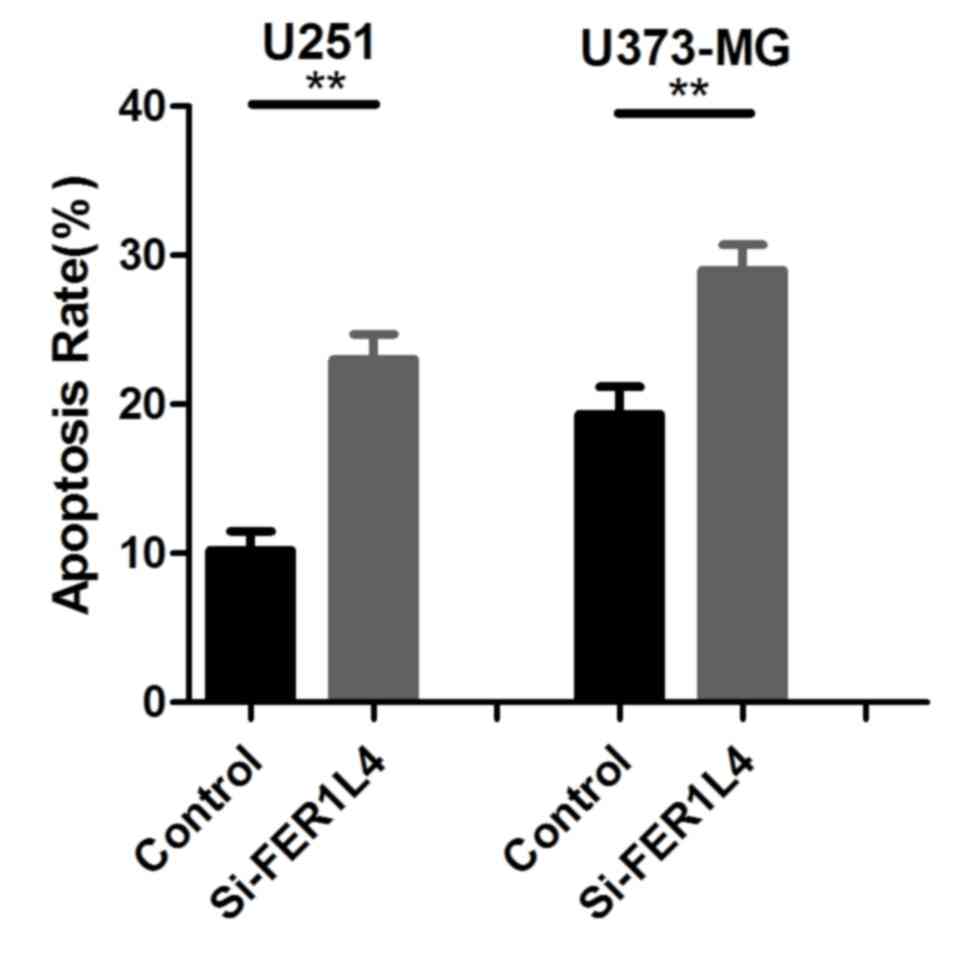
invasion, respectively. Flow cytometry was performed to detect
changes in the apoptosis of glioma cells. The survival rates of the
FER1L4 siRNA-treated glioma cells (U251 or U373-MG) were
significantly lower compared with that of the control cells
(P<0.05; Fig. 3), and the
percentage of apoptotic si-FER1L4 glioma cells was significantly
higher compared with that of the control cells (P<0.01; Fig. 4). The Transwell assay revealed that
the number of si-FER1L4-treated glioma cells penetrating the
basement membrane in each field of view was significantly lower
compared with that of the control cells (P<0.0001; Fig. 5).
Discussion
Studies that investigate the underlying molecular
mechanisms of neuroglioma pathogenesis, and identify novel
treatments and targets for gene therapy are required to improve the
prognosis of patients with neuroglioma (22). LncRNAs, including maternally expressed
3 (MEG3) and H19 imprinted maternally expressed transcript (H19)
have been associated with the occurrence and progression of
neuroglioma (23–25). MEG3 was the first lncRNA demonstrated
to have a tumor suppressor effect, and is typically upregulated in
tumor tissues compared with normal tissues (26); however, a previous study demonstrated
that MEG3 is either not expressed or markedly downregulated in 82%
of patients with glioma (24). The
activity of cellular tumor antigen p53 is regulated through the
downregulation of MEG3, which creates favorable conditions for the
in vitro growth of glioma cells (24). MEG3 may regulate the occurrence and
progression of glioma, and therefore represents a novel target for
gene therapy (27).
H19 was one of the earliest lncRNAs to be identified
and is highly expressed in the embryonic stages of development. H19
is primarily expressed in endoderm- and mesoderm-derived tissues
and belongs to the maternally-derived imprinted gene family
(28). A previous study suggested
that H19 serves a role as an oncogene and a tumor suppressor gene
(28). Shi et al (25) discussed the potential biological
functions of H19 in glioma. H19 expression was associated with
glioma stage; the expression of H19 and its derivative microRNA,
miR-675, in high-grade glioma was significantly higher compared
with that in low-grade glioma. As H19 was downregulated, miR-675
was also downregulated. H19 is the precursor of miR-675, and H19′s
oncogenic and regulatory effects on the invasiveness of glioma
cells are achieved through miR-675 and the inhibition of
cadherin-13 (25). In addition, H19
regulates the activity of transcriptional factor zinc finger
protein GLI1 (GLI1), which serves a role in the development of
astrocytoma. Abnormally high expression of GLI1 is typically
detected in brain glioma, and the upregulation of GLI1 is
associated with an increased cell proliferation index, pathology
index and the recurrence of glioma (29).
FER1L4 has primarily been studied in gastric and
colon cancer (17,18). Liu et al (30) demonstrated that FER1L4 was
downregulated in 91.80% of gastric cancer tissues studied.
Additionally, low expression of FER1L4 was associated with tumor
size, histological staging, tumor infiltration depth, lymph node
metastasis, distant metastasis, tumor-node-metastasis staging,
angioneurotic infiltration and serum CA72-4 antigen levels. The
expression of FER1L4 is a diagnostic indicator for gastric cancer
at an early stage (30). A subsequent
study demonstrated that FER1L4 is a competing endogenous RNA that
acts as a tumor suppressor in colon cancer (18), where its expression is negatively
correlated with miR-106a-5p expression. The expression levels of
FER1L4 and miR-106a-5p are correlated with tumor infiltration,
lymph node metastasis, vascular infiltration and clinical staging
in colon cancer. By increasing the expression of FER1L4
exogenously, miR-106a-5p expression can be downregulated, thus
reducing the proliferation, migration and invasion of colon cancer
(18).
In the present study, the expression of FER1L4 in
patients with different grade of glioma was investigated by mining
TCGA data. FER1L4 was significantly upregulated in patients with
high-grade glioma compared with patients with LGG, and high
expression of FER1L4 significantly predicted a poor prognosis for
patients with glioma. Therefore, FER1L4 may be a prognostic
indicator for glioma. Additionally, glioma cell invasion and
viability were significantly inhibited by transfection with a
FER1L4 siRNA, which promoted apoptosis. These results suggest that
FER1L4 has a cancer-promoting effect in glioma and is a novel
target for gene therapy. However, further studies are required to
investigate the underlying molecular mechanisms of the roles that
FER1L4 serves in glioma.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81502147), Zhejiang
Medical Science and Technology Project (2017194140) and the Youth
Scientific Innovation Foundation of Zhejiang Cancer Hospital (grant
no. QN201402).
References
|
1
|
Soffietti R, Bertero L, Pinessi L and Rudà
R: Pharmacologic therapies for malignant glioma: A guide for
clinicians. CNS Drugs. 28:1127–1137. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Morgan LL: The epidemiology of glioma in
adults: A ‘state of the science’ review. Neuro Oncol. 17:623–624.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bai H, Harmanc AS, Erson-Omay EZ, Li J,
Coşkun S, Simon M, Krischek B, Özduman K, Omay SB, Sorensen EA, et
al: Integrated genomic characterization of IDH1-mutant glioma
malignant progression. Nat Genet. 48:59–66. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Simonetti G, Gaviani P, Innocenti A,
Botturi A, Lamperti E and Silvani A: Update on treatment strategies
for anaplastic glioma: A review of literature. Neurol Sci.
35:977–981. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Y and Jiang T: Understanding high
grade glioma: Molecular mechanism, therapy and comprehensive
management. Cancer Lett. 331:139–146. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Spizzo R, Almeida MI, Colombatti A and
Calin GA: Long non-coding RNAs and cancer: A new frontier of
translational research? Oncogene. 31:4577–4587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wapinski O and Chang HY: Long noncoding
RNAs and human disease. Trends Cell Biol. 21:354–361. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
ENCODE Project Consortium, ; Birney E,
Stamatoyannopoulos JA, Dutta A, Guigó R, Gingeras TR, Margulies EH,
Weng Z, Snyder M, Dermitzakis ET, et al: Identification and
analysis of functional elements in 1% of the human genome by the
ENCODE pilot project. Nature. 447:799–816. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Derrien T, Johnson R, Bussotti G, Tanzer
A, Djebali S, Tilgner H, Guernec G, Martin D, Merkel A, Knowles DG,
et al: The GENCODE v7 catalog of human long noncoding RNAs:
Analysis of their gene structure, evolution, and expression. Genome
Res. 22:1775–1789. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tsai MC, Manor O, Wan Y, Mosammaparast N,
Wang JK, Lan F, Shi Y, Segal E and Chang HY: Long noncoding RNA as
modular scaffold of histone modification complexes. Science.
329:689–693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wan L, Zhang L, Fan K, Cheng ZX, Sun QC
and Wang JJ: Knockdown of long noncoding RNA PCAT6 inhibits
proliferation and invasion in lung cancer cells. Oncol Res.
24:161–170. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Thorenoor N, Faltejskova-Vychytilova P,
Hombach S, Mlcochova J, Kretz M, Svoboda M and Slaby O: Long
non-coding RNA ZFAS1 interacts with CDK1 and is involved in
p53-dependent cell cycle control and apoptosis in colorectal
cancer. Oncotarget. 7:622–637. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mendell JT: Targeting a long noncoding RNA
in breast cancer. N Engl J Med. 374:2287–2289. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Reon BJ, Anaya J, Zhang Y, Mandell J,
Purow B, Abounader R and Dutta A: Expression of lncRNAs in
low-grade gliomas and glioblastoma multiforme: An in silico
analysis. PLoS Med. 13:e10021922016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cabili MN, Trapnell C, Goff L, Koziol M,
Tazon-Vega B, Regev A and Rinn JL: Integrative annotation of human
large intergenic noncoding RNAs reveals global properties and
specific subclasses. Genes Dev. 25:1915–1927. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huarte M and Rinn JL: Large non-coding
RNAs: Missing links in cancer? Hum Mol Genet. 19:R152–R161. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Song H, Sun W, Ye G, Ding X, Liu Z, Zhang
S, Xia T, Xiao B, Xi Y and Guo J: Long non-coding RNA expression
profile in human gastric cancer and its clinical significances. J
Transl Med. 11:2252013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yue B, Sun B, Liu C, Zhao S, Zhang D, Yu F
and Yan D: Long non-coding RNA Fer-1-like protein 4 suppresses
oncogenesis and exhibits prognostic value by associating with
miR-106a-5p in colon cancer. Cancer Sci. 106:1323–1332. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ceccarelli M, Barthel FP, Malta TM,
Sabedot TS, Salama SR, Murray BA, Morozova O, Newton Y, Radenbaugh
A, Pagnotta SM, et al: Molecular profiling reveals biologically
discrete subsets and pathways of progression in diffuse glioma.
Cell. 164:550–563. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dehghani Mohammad, Abadi M, Ashraf N,
Chamsaz M and Shemirani F: An overview of liquid phase
microextraction approaches combined with UV-Vis spectrophotometry.
Talanta. 99:1–12. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Regier N and Frey B: Experimental
comparison of relative RT-qPCR quantification approaches for gene
expression studies in poplar. BMC Mol Biol. 11:572010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hamza MA and Gilbert M: Targeted therapy
in gliomas. Curr Oncol Rep. 16:3792014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Argyriou AA and Kalofonos HP: Molecularly
targeted therapies for malignant gliomas. Mol Med. 15:115–122.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang P, Ren Z and Sun P: Overexpression of
the long non-coding RNA MEG3 impairs in vitro glioma cell
proliferation. J Cell Biochem. 113:1868–1874. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shi Y, Wang Y, Luan W, Wang P, Tao T,
Zhang J, Qian J, Liu N and You Y: Long non-coding RNA H19 promotes
glioma cell invasion by deriving miR-675. PLoS One. 9:e862952014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou Y, Zhang X and Klibanski A: MEG3
noncoding RNA: A tumor suppressor. J Mol Endocrinol. 48:R45–R53.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li J, Bian EB, He XJ, Ma CC, Zong G, Wang
HL and Zhao B: Epigenetic repression of long non-coding RNA MEG3
mediated by DNMT1 represses the p53 pathway in gliomas. Int J
Oncol. 48:723–733. 2016.PubMed/NCBI
|
|
28
|
Ariel I, de Groot N and Hochberg A:
Imprinted H19 gene expression in embryogenesis and human cancer:
The oncofetal connection. Am J Med Genet. 91:46–50. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tchoghandjian A, Baeza-Kallee N, Beclin C,
Metellus P, Colin C, Ducray F, Adélaïde J, Rougon G and
Figarella-Branger D: Cortical and subventricular zone
glioblastoma-derived stem-like cells display different molecular
profiles and differential in vitro and in vivo properties. Ann Surg
Oncol. 19 Suppl 3:S608–S619. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu Z, Shao Y, Tan L, Shi H, Chen S and
Guo J: Clinical significance of the low expression of FER1L4 in
gastric cancer patients. Tumour Biol. 35:9613–9617. 2014.
View Article : Google Scholar : PubMed/NCBI
|