Introduction
Hypopharyngeal squamous cell carcinoma (HSCC) is one
of the most common types of aggressive head and neck malignancy,
with an incidence of ~10 cases/million people annually (1). A total of >75% of patients with HSCC
have an advanced stage of the disease at the time of diagnosis
(2), and the prognosis of these
patients is poor with a 5-year overall survival rate of 30–35%
(3–5).
The poor prognosis and frequently advanced stage at diagnosis are
typically attributed to the local aggressiveness of the tumor,
which exhibits a propensity for submucosal spread, invasion into
adjacent structure and metastasis due to abundant lymphatic
drainage (6,7). These events are regulated by numerous
key molecular oncogenic pathways. Therefore, understanding these
pathways in HSCC may assist in understanding the biological
characteristics of the tumor, and lead to novel insights to the
search for improved disease therapy and preventative
strategies.
Long noncoding RNAs (lncRNAs) are a novel class of
RNA molecules defined as transcripts of >200 nucleotides (nt)
that lack protein coding potential. They are typically transcribed
by RNA polymerase II, but possess no open reading frame and map to
intronic and intergenic regions. Additionally, lncRNAs demonstrate
epigenetic characteristics that are similar to protein-coding genes
(8). It has been estimated that
~15,000 lncRNAs are present in the human genome (9). Previous studies have demonstrated that
lncRNAs may function as oncogenes or tumor suppressors in the
cancer initiatome (10), serve
important roles in carcinogenesis and cancer proliferation,
invasion and metastasis, and correlate with cancer prognosis
(11,12). Therefore, lncRNAs are a potential
novel class of cancer biomarkers (13).
The present study focuses on lncRNA paternally
expressed 10 (PEG10; NONCODE Gene ID NONHSAG048235), which is
located on human chromosome 7 between the 94285681 and 94298949
base sites and is 763 bp in length (14). It was demonstrated that aberrant
expression of PEG10 is associated with a number of malignancies,
including hepatocellular carcinoma (15), B-cell lymphocytic leukemia (16), lymphoma (17) and esophageal cancer (18). With respect to esophageal cancer, it
was identified that PEG10 regulates proliferation and invasion of
esophageal cancer cells (18). In
addition, studies have revealed that PEG10 is highly conserved
across mammalian species, and serves an important role in cell
proliferation, differentiation and metastasis (10,19).
However, the potential role of PEG10 in human hypopharyngeal
carcinoma remains to be elucidated. Therefore, the aim of the
present study was to investigate the role of PEG10 in HSCC.
Materials and methods
Patient selection and tissue sample
collection
Patients who underwent curable surgery for
hypopharyngeal carcinoma between January 2010 and October 2014 were
screened, and cases, in accordance with the following criteria,
were selected for the study: i) Diagnosis of HSCC was confirmed by
postoperative pathological results; ii) patient did not receive
chemotherapy or radiotherapy prior to surgery; iii) patient
presented with local disease without any distant metastases at the
time of diagnosis, and the tumor was resectable; iv) complete tumor
resection was described in the surgical record with expected
disease-free survival of ≥3 months; v) patient received standard
surgery for hypopharyngeal carcinoma, namely, primary tumor
resection with lymph node dissection, and surgical margins were
microscopically negative with no residual tumor; and vi)
clinicopathological information (sex, age, tumor location, lymph
node metastasis, differentiation and classification) was
available.
For every eligible patient, primary tumor tissue and
para-carcinoma tissue samples were immediately collected and frozen
in liquid nitrogen following resection and stored at −80°C until
RNA extraction. Para-carcinoma tissue was defined as normal tissue
adjacent to the primary tumor without any microscopic invasion.
All patients were from the Second Hospital of
Shandong University and the Provincial Hospital Affiliated to
Shandong University (Jinan, China). Written informed consent was
obtained from all patients prior to participation in the study. The
medical ethics committee of the Second Hospital of Shandong
University and the Shandong Provincial Hospital approved the
present study.
RNA extraction and reverse
transcription quantitative polymerase chain reaction (RT-qPCR)
assay
For all eligible patients, total RNA was extracted
from frozen primary tumor and para-carcinoma tissue samples. Total
RNA was extracted and purified with the RecoverAll™ Total Nucleic
Acid Isolation kit (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). To ensure that total RNA fulfilled the requirements for the
qPCR, the purity and quantity of RNA for each sample were analyzed
(a minimum A260/A280 ratio of >1.8 was applied for all samples)
and RNA integrity was assessed by formaldehyde-modified gel
electrophoresis. Reverse transcription reactions were performed
using 2 µg total RNA, lncRNA PEG10 reverse transcription primer
(Invitrogen; Thermo Fisher Scientific, Inc.), 5X first-strand
buffer (Thermo Fisher Scientific, Inc.), 0.1 M DTT (Thermo Fisher
Scientific, Inc.), a dNTP mixture (Takara, Bio, Inc., Otsu, Japan),
Moloney Murine Leukemia Virus reverse transcriptase (Thermo Fisher
Scientific, Inc.) and recombinant RNasin® RNase inhibitor (Promega
Corporation, Madison, WI, USA). Following reverse transcription,
qPCR reactions were performed using a 7900 HT Fast RealTime PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Dissociation curve
analysis was performed to assess the specificity of the amplified
product. GAPDH was used as an internal control by comparing its
expression levels in each of the specimens. Finally, 1.5%
non-denaturing agarose gel electrophoresis using 4 µl qPCR products
was used to evaluate their quantity and specificity.
The thermocycler conditions for the RT-qPCR were as
follows: 10 min denaturation at 95°C, followed by 40 cycles of 95°C
for 15 sec, and 60°C for 1 min. SYBR-Green was used as the
fluorophore. The method of quantification used for the qPCR was
2−ΔΔCq method (20) and
the results were replicated three times.
The following primer sequences were used: lncRNA
PEG10 primer sequence: Forward, 5′-CATCCTTCCTGTCTTCGC-3′; reverse,
5′-CCCTCTTCCACTCCTTCTTT-3′; probe,
5′-Fam-CCGCTTATTTCACGCGAGGA-Tamra-3′; GAPDH primer sequence:
Forward, 5′-TGGTATCGTGGAAGGACTCA-3′; reverse,
5′-CCAGTAGAGGCAGGGATGAT-3′; Probe,
5′-Fam-CGCCACAGTTTCCCGGAGG-Tamra-3′. All primers were synthesized
by Invitrogen (Thermo Fisher Scientific, Inc.).
Cell culture and transfection
The HSCC FaDu cell line was obtained from Shanghai
Institutes for Biological Science (Shanghai, China). FaDu cells
were cultured in RPMI-1640 medium (Nanjing KeyGen Biotech Co.,
Ltd., Nanjing, China) supplemented with 10% fetal bovine serum
(FBS; Invitrogen; Thermo Fisher Scientific, Inc.) All cells were
cultured at 37°C in a humidified atmosphere containing 5%
CO2.
In order to obtain high expression of lncRNA PEG10,
FaDu cells were transfected with lncRNA PEG10. Small interfering
RNA (siRNA) against lncRNA PEG10 were also designed and transfected
to inhibit lncRNA PEG10 expression. Transfection was performed
using Lipofectamine 2000® (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. The experimental
groups included FaDu cells transfected with lncRNA PEG10 (PEG10+),
cells transfected with silencer (si) lncRNA (si-lnc) and the
negative control cells (NC) transfected with a nonsense siRNA
control sequence. Transfection efficiency was measured in each
experimental group 5 times using RT-qPCR analysis.
Cell proliferation assay
The Cell Counting kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was used to evaluate the
proliferative activity of FaDu cells according to the
manufacturer's protocol. Following transfection, cells from each
experimental group were seeded into 96-well plates at a density of
1×104 cells/well, with 5 replicate wells/group. The
absorbance value for each well was determined using CCK-8. The
optical density (OD) value represented the proliferation of FaDu
cells and was measured daily over 4 consecutive days at a
wavelength of 450 nm (OD450) to estimate the number of viable
cells.
Transwell invasion assay
Transwell invasion assays were used to evaluate
invasive activity of FaDu cells and were performed using a 24-well
chamber containing Matrigel (3.9 µg/µl, 60–80 µl)-coated membranes
with a pore size of 8 µm (Costar; Corning Incorporated, Corning,
NY, USA). Experimental group cells and the control cells were
collected and resuspended in serum-free medium at a concentration
of 2×105 cells/ml. Cell suspensions (200 µl) were added
to the upper chamber, the bottom chamber was filled with 500 µl of
culture medium containing 10% FBS and cells were incubated for 48 h
at 37°C with 5% CO2. A total of 5 wells were used for
each group. Following incubation, the medium and stationary cells
on the upper membrane surface were removed with a cotton tip and
the cells that had passed through the filter were fixed in
methanol, stained with 0.1% crystal violet for 10 min, mounted and
dried at 80°C for 30 min. The number of cells invading the Matrigel
was counted in 3 randomly selected fields using an inverted
microscope at magnification, ×200. The experiment was repeated 5
times.
Wound healing assay
Wound healing assays were performed to evaluate the
migration activity of FaDu cells. Then, 5×105 cells/ml
cells from each experimental group were seeded on 6-well plates and
cultured in RPMI-1640 medium supplemented with 10% FBS for 24 h to
form a confluent monolayer cell. An artificial scratch wound on
each confluent monolayer was created with a 10-µl pipette tip.
Serum-free medium was added for an additional 24 h incubation, and
images of the cells were captured at 4 different time points (0,
24, 36 and 48 h) using an inverted microscope at magnification,
×200. The distance between the borders of the wounded region
lacking cells was measured, and the relative migration ratio was
calculated at each time point with 5 replicates/group.
Statistical analysis
The expression level of lncRNA PEG10 in each
experimental group was calculated as mean ± standard deviation. The
differences between expression levels of lncRNA PEG10 were analyzed
with paired sample t-tests, independent sample t-tests or one-way
analysis of variance (ANOVA) with the Bonferroni test used as the
post-hoc test. Statistical analyses were performed using the SPSS
statistical software program (SPSS v17.0, SPSS Inc., Chicago, IL,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Eligible patient clinicopathological
characteristics and association with the expression of lncRNA
PEG10
Following the inclusion criteria, 56 patients were
eligible for the present study. The eligible patients included 40
males and 16 females, with a median age of 59 years (range, 47–76).
The post-operation stage was defined according to the 8th edition
American Joint Committee on Cancer (AJCC) classification system
(21). All patient characteristics
are summarized in Table I.
 | Table I.Association of lncRNA PEG10 expression
with clinicopathological characteristics. |
Table I.
Association of lncRNA PEG10 expression
with clinicopathological characteristics.
| Clinicopathological
characteristics | n | lncRNA PEG10
expression (2−ΔΔCq values)a | P-value |
|---|
| Patient number | 56 |
|
|
| Age, years |
|
| 0.3580 |
| ≤60 | 32 | 3.2725±1.2293 |
|
|
>60 | 24 | 3.5850±1.2734 |
|
| Sex |
|
| 0.1219 |
|
Male | 40 | 3.2430±1.2221 |
|
|
Female | 16 | 3.8150±1.2514 |
|
| Primary tumor
site |
|
| 0.9938 |
|
Pyriform sinuses | 39 | 3.3967±1.3083 |
|
|
Posterior and lateral
hypo-pharyngeal walls | 13 | 3.4169±1.1604 |
|
|
Postcricoid area | 4 | 3.4675±1.1915 |
|
| Pathological
differentiation |
|
| 0.4564 |
|
Well-differentiated | 12 | 3.0208±1.1527 |
|
|
Moderately differentiated | 19 | 3.5931±1.1285 |
|
| Poorly
differentiated | 25 | 3.4496±1.3761 |
|
| Primary tumor
size |
|
| 0.0174 |
| T1 | 13 | 2.8131±1.0142 |
|
| T2 | 23 | 3.2330±1.1519 |
|
| T3 | 20 | 3.9915±1.2943 |
|
| Lymph node
status |
|
| 0.0002 |
| N0 | 19 | 2.5774±0.9506 |
|
| N+ | 37 | 3.8322±1.1720 |
|
| Clinical staging
(Tumor node metastasis) |
|
| <0.0000 |
|
I/II | 13 | 2.2100±0.7106 |
|
|
III | 15 | 3.2920±0.7290 |
|
| IV | 28 | 4.0232±1.2550 |
|
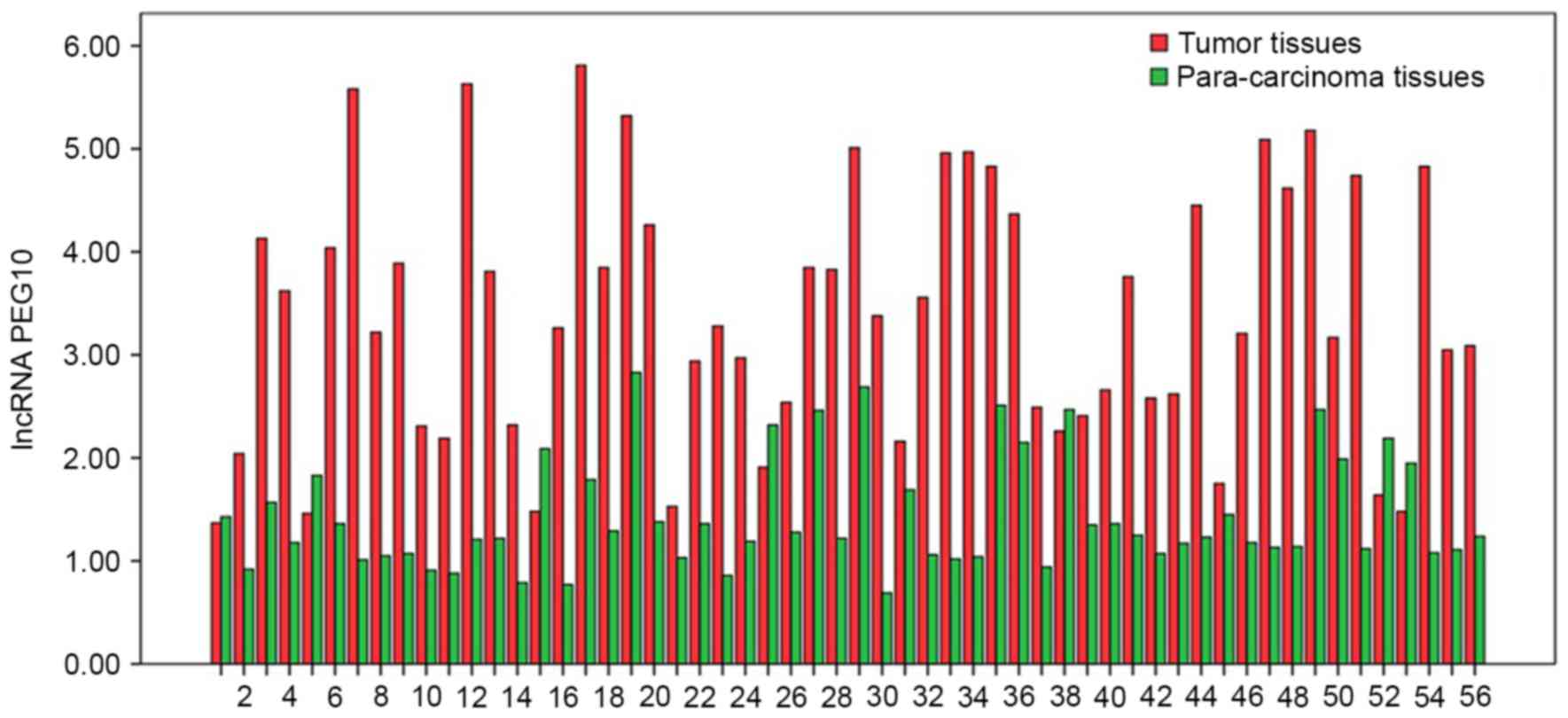
The expression levels of lncRNA PEG10 in tumor
tissues and para-carcinoma tissues, presented as 2−ΔΔCq
values, were 3.406±1.247 and 1.429±0.544, respectively (P<0.05;
Fig. 1). The associations between the
expression of lncRNA PEG10 in tumor tissues and clinicopathological
characteristics of patients with HSCC were also analyzed. The
results demonstrated that primary tumor size, lymph node status and
Tumor node metastasis (TNM) stage were significantly associated
with an increased expression of lncRNA PEG10 (P<0.05; Table I). By contrast, there were no
significant associations observed between lncRNA PEG10 expression
and age, sex, primary tumor site or pathological differentiation
(Table I). Based on these results,
additional experimental studies with lncRNA PEG10 were performed to
determine its role in HSCC pathogenesis.
Expression profile of lncRNA PEG10 in
transfected FaDu cells
Expression levels of lncRNA PEG10 were examined
using RT-qPCR in FaDu cells transfected with lncRNA PEG10 (PEG10+),
negative control RNA (NC) or silencer lncRNA PEG10 (si-lnc).
Expression levels quantified as 2−ΔΔCq values were
2.448±0.440, 1.762±0.280 and 0.552±0.209 in PEG10+, NC and si-lnc
groups, respectively (Fig. 2).
Differences among groups were identified to be significantly
different using one-way ANOVA analysis (P<0.05). Post-hoc
testing demonstrated that lncRNA PEG10 expression was significantly
reduced in cells transfected with silencer si-lnc RNA compared with
PEG10+ and NC cells (P<0.05), while lncRNA PEG10 expression was
enhanced in PEG10+ cells compared with NC cells (P<0.05).
lncRNA PEG10 promotes cell
proliferation
The alterations in lncRNA PEG10 expression in
transfected FaDu cells were associated with alterations in
proliferative capacity. Mean OD450 values from replicate
cell proliferation assays of PEG10+, NC and si-lnc cells,
respectively, were as follows: 24 h: 0.496±0.075, 0.384±0.035 and
0.260±0.052; 48 h: 0.892±0.033, 0.672±0.046 and 0.428±0.080; 72 h:
1.124±0.101, 0.900±0.073 and 0.560±0.037; 96 h: 1.308±0.101,
1.054±0.102 and 0.752±0.056. The cell growth curves are
demonstrated in Fig. 3. Proliferation
values for the PEG10+ group were significantly increased compared
with those of the NC group at all time points (P<0.05), while
values for the si-lnc group were significantly decreased at the
identical time points compared with the PEG10+ and NC groups
(P<0.05). These results indicate that overexpression of lncRNA
PEG10 promoted in vitro proliferation of FaDu cells.
lncRNA PEG10 promotes cell
invasion
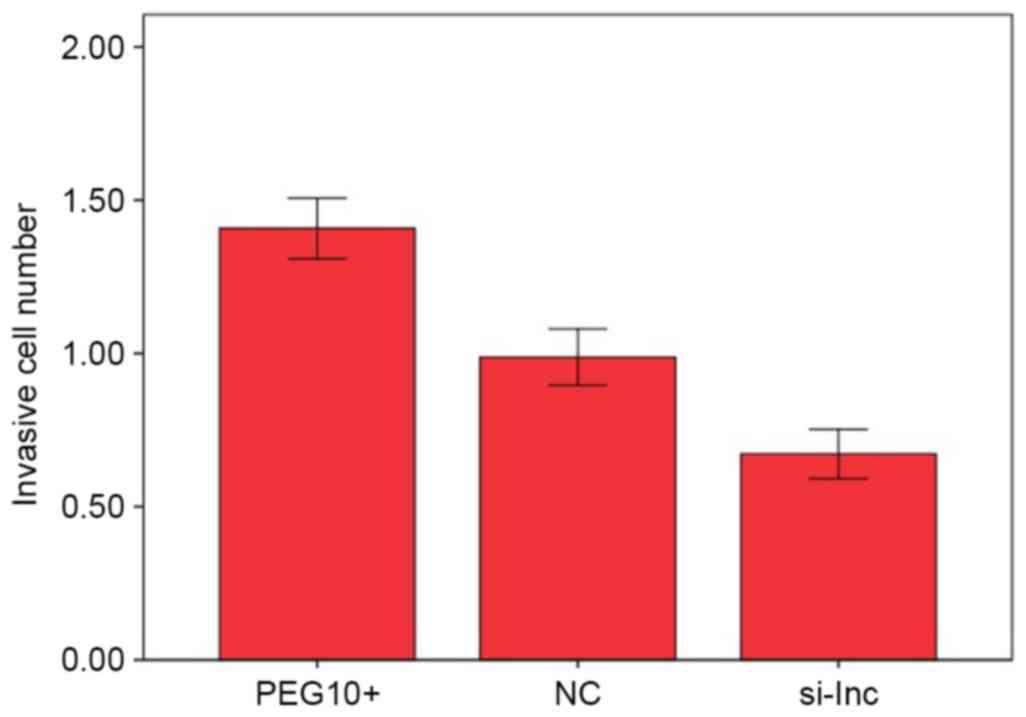
In Transwell assays, the means of cell numbers
penetrating the membranes in the PEG10+, NC and si-lnc groups were
1.408±0.099×105, 0.988±0.092×105 and
0.672±0.081×105, respectively. The number of invading
cells was significantly higher in the PEG10+ group compared with
the NC group (P<0.05; Fig. 4). By
contrast, the number of invading cells in the si-lnc group was
significantly lower compared with the that in the PEG10+ and NC
groups (P<0.05; Fig. 4). These
results indicate that the overexpression of lncRNA PEG10 increased
the invasive capacity of FaDu cells in vitro.
lncRNA PEG10 promotes cell
migration
Finally, cell migration in transfected FaDu cells
was examined using wound healing assays. The mean relative
migration ratios for cells in the PEG10+, NC and si-lnc groups,
respectively, were as follows: 24 h: 0.514±0.045, 0.356±0.036 and
0.320±0.047; 36 h: 0.808±0.041, 0.652±0.051 and 0.552±0.054; 48 h:
0.952±0.031, 0.830±0.036 and 0.698±0.038. The relative migration
ratio of FaDu cells in the PEG10+ group was significantly increased
compared with the ratios in the NC and si-lnc groups at each time
point (P<0.05; Fig. 5). Similarly,
the relative migration ratio of FaDu cells in the NC group was
significantly increased compared with the ratio in the si-lnc group
at 36 and 48 h (P<0.05; Fig. 5),
but the difference was not significant at 24 h. These results
suggest that downregulation of lncRNA PEG10 dramatically decreased
the migration ability of FaDu cells.
Discussion
In humans, ~70% of the genome is transcribed to
generate a range of noncoding RNAs (22). Based on transcript size, noncoding
RNAs are classified into small noncoding RNAs (<200 nt) and
lncRNAs (>200 nt). Small noncoding RNAs, particularly miRNAs,
are well characterized as post-transcriptional regulators of mRNAs
and have established roles in cancer (23,24).
lncRNAs remain poorly characterized, but evidence for their
importance and functionality has increased in previous years. Due
to their numerous structural and biochemical characteristics,
lncRNAs are implicated in a diverse range of functions, including
nuclear architecture, immune surveillance, imprinting, epigenetic
regulation, cellular trafficking, splicing and pluripotency of
embryonic stem cells. lncRNAs may regulate gene expression at a
number of levels, including chromatin modification, transcription,
splicing, translation, post-transcriptional regulation, processing
of small RNAS and other important functions (25,26). They
may affect and regulate the cell cycle, proliferation,
differentiation and apoptosis. Previous studies provide evidence
that lncRNAs are also involved in cancer development and the
maintenance of tumorigenesis (25),
and serve important roles in modulating the proliferation, invasion
and metastasis of tumors (27).
Previously, dysregulation of lncRNAs has been detected in numerous
types of cancer, including breast cancer, hepatocellular carcinoma,
melanoma, bladder and prostate cancer (12). Furthermore, the role of certain
lncRNAs in cancer, including HOX transcript antisense RNA,
metastasis associated lung adenocarcinoma transcript 1 and taurine
up-regulated 1 (28–30), has been previously elucidated.
HSCC is one of the most prevalent types of
aggressive head and neck malignancy. The most important clinical
characteristic of this cancer is its tendency to invade adjacent
structures and lymph node metastasis (31). However, despite previous
investigation, the underlying mechanisms controlling invasion and
metastasis of HSCC remain to be elucidated. The modulatory
functions of lncRNAs in HSCC remain to be fully understood.
Therefore, the present study investigated the effect of lncRNA
PEG10 on proliferation, invasion and metastasis in HSCC.
In a previous esophageal cancer study (18), PEG10 was indicated to regulate
proliferation, invasion and metastasis of esophageal cancer cells.
In the present study, the expression levels of PEG10 in tumor
tissues and normal para-carcinoma tissues in HSCC were measured and
demonstrated that there were increased levels in the cancerous
tissue. This indicates that PEG10 is associated with HSCC and may
possibly participate in the development and carcinogenesis of HSCC.
In the present study, the association between PEG10 expression and
clinicopathological characteristics in 56 eligible patients
exhibited a marked association between PEG10 expression in HSCC
tumor tissues and primary tumor size, local lymph node status and
TNM stage. These results suggest that overexpression of lncRNA
PEG10 in tumor tissues promotes the proliferation, invasion and
local lymph node metastasis of HSCC, supporting the hypothesis of
the present study regarding the role of PEG10 in cancer, and
providing a basis for further in vitro experimental studies
using the HSCC FaDu cell line.
In a series of in vitro experiments, PEG10
levels were manipulated in transfected FaDu cells and it was
identified that increased expression of PEG10 enhanced the
proliferation, invasion and migration of FaDu cells. These results
suggest that an overexpression of lncRNA PEG10 may promote
proliferation, invasion and metastasis of HSCC cells, in accordance
with the association between PEG10 expression and
clinicopathological characteristics. Therefore, results from
clinical and in vitro approaches support the conclusion that
lncRNA PEG10 is involved in HSCC.
There have been a small number of studies regarding
the potential mechanisms underlying the ability of PEG10 to promote
proliferation, invasion and metastasis of HSCC cells. Zang et
al (18) suggested that lncRNA
PEG10 may modulate these functions by controlling cell cycle
distribution. As for other lncRNAs, certain studies have indicated
that lncRNAs may regulate cell proliferation, invasion and
metastasis by modulating downstream protein expression, cell cycle
arrest and apoptosis (18,32). As lncRNAs may regulate gene expression
at transcriptional and post-transcriptional levels (12,24,25), it
has been difficult to demonstrate the exact underlying mechanism by
which PEG10 promotes proliferation, invasion and metastasis of HSCC
cells without systematic studies.
In conclusion, the present study demonstrated that
lncRNA PEG10 was upregulated in HSCC and that an overexpression of
lncRNA PEG10 promoted proliferation, invasion and migration of the
FaDu HSCC cell line. These data suggest that lncRNA PEG10 may be a
effective therapeutic target in hypopharyngeal carcinoma therapy
and that mechanisms underlying its effect should be further
investigated.
Acknowledgements
The present study was supported by the National
Science Foundation of China (grant no. 81102019), the Natural
Science Foundation of Shandong Province (grant no. 2012BSE27043)
and the Medicine and Health Science Technology Development Project
of Shandong Province (grant no. 2014WS0422).
References
|
1
|
Davies L and Welch HG: Epidemiology of
head and neck cancer in the United States. Otolaryngol Head Neck
Surg. 135:451–457. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Smith RB, Apostolakis LW, Karnell LH, Koch
BB, Robinson RA, Zhen W, Menck HR and Hoffman HT: National cancer
data base report on osteosarcoma of the head and neck. Cancer.
98:1670–1680. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hoffman HT, Karnell LH, Shah JP, Ariyan S,
Brown GS, Fee WE, Glass AG, Goepfert H, Ossoff RH and Fremgen AM:
Hypopharyngeal cancer patient care evaluation. Laryngoscope.
107:1005–1017. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bova R, Goh R, Poulson M and Coman WB:
Total pharyngolaryngectomy for squamous cell carcinoma of
hypopharynx: A review. Laryngoscope. 115:864–869. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Edge SB and Compton CC: The American Joint
Committee on Cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Elias MM, Hilgers FJ, Keus RB, Gregor RT,
Hart AA and Balm AJ: Carcinoma of the pyriform sinus: A
retrospective analysis of treatment results over a 20-year period.
Clin Otolaryngol Allied Sci. 20:249–253. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wahlberg PC, Andersson KE, Biörklund AT
and Möller TR: Carcinoma of the hypopharynx: Analysis of incidence
and survival in Sweden over a 30-year period. Head Neck.
20:714–719. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kurokawa R: Long noncoding RNA as a
regulator for transcription. Prog Mol Subcell Biol. 51:29–41. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Walsh AL, Tuzova AV, Bolton EM, Lynch TH
and Perry AS: Long noncoding RNAs and prostate carcinogenesis: The
missing ‘linc’? Trends Mol Med. 20:428–436. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wapinski O and Chang HY: Long noncoding
RNAs and human disease. Trends Cell Biol. 21:354–361. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Prensner JR and Chinnaiyan AM: The
emergence of lncRNAs in cancer biology. Cancer Discov. 1:391–407.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gutschner T and Diederichs S: The
hallmarks of cancer: A long non-coding RNA point of view. RNA Biol.
9:703–719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yarmishyn AA and Kurochkin IV: Long
noncoding RNAs: A potential novel class of cancer biomarkers. Front
Genet. 6:1452015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ono R, Kobayashi S, Wagatsuma H, Aisaka K,
Kohda T, Kaneko-Ishino T and Ishino F: A retrotransposonderived
gene, PEG10, is a novel imprinted gene located on human chromosome
7q21. Genomics. 73:232–237. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Okabe H, Satoh S, Furukawa Y, Kato T,
Hasegawa S, Nakajima Y, Yamaoka Y and Nakamura Y: Involvement of
PEG10 in human hepatocellular carcinogenesis through interaction
with SIAH1. Cancer Res. 63:3043–3048. 2003.PubMed/NCBI
|
|
16
|
Kainz B, Shehata M, Bilban M, Kienle D,
Heintel D, Krömer-Holzinger E, Le T, Kröber A, Heller G,
Schwarzinger I, et al: Overexpression of the paternally expressed
gene 10 (PEG10) from the imprinted locus on chromosome 7q21 in
high-risk B-cell chronic lymphocytic leukemia. Int J Cancer.
121:1984–1993. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Peng W, Fan H, Wu G, Wu J and Feng J:
Upregulation of long noncoding RNA PEG10 associates with poor
prognosis in diffuse large B cell lymphoma with facilitating
tumorigenicity. Clin Exp Med. 16:177–182. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zang W, Wang T, Huang J, Li M, Wang Y, Du
Y, Chen X and Zhao G: Long noncoding RNA PEG10 regulates
proliferation and invasion of esophageal cancer cells. Cancer Gene
Ther. 22:138–144. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hu C, Xiong J, Zhang L, Huang B, Zhang Q,
Li Q, Yang M, Wu Y, Wu Q, Shen Q, et al: PEG10 activation by
co-stimulation of CXCR5 and CCR7 essentially contributes to
resistance to apoptosis in CD19+CD34+ B cells from patients with B
cell lineage acute and chronic lymphocytic leukemia. Cell Mol
Immunol. 1:280–294. 2004.PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
The 8th edition American Joint Committee
on Cancer (AJCC) classification system. http://cancerstaging.org/About/Pages/8th-Edition.aspx2010
|
|
22
|
Djebali S, Davis CA, Merkel A, Dobin A,
Lassmann T, Mortazavi A, Tanzer A, Lagarde J, Lin W, Schlesinger F,
et al: Landscape of transcription in human cells. Nature.
489:101–108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fabian MR and Sonenberg N: The mechanics
of miRNA-mediated gene silencing: A look under the hood of miRISC.
Nat Struct Mol Biol. 19:586–593. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kong YW, Ferland-McCollough D, Jackson TJ
and Bushell M: microRNAs in cancer management. Lancet Oncol.
13:e249–e258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gibb EA, Vucic EA, Enfield KS, Stewart GL,
Lonergan KM, Kennett JY, Becker-Santos DD, MacAulay CE, Lam S,
Brown CJ and Lam WL: Human cancer long non-coding RNA
transcriptomes. PLoS One. 6:e259152011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ma L, Bajic VB and Zhang Z: On the
classification of long non-coding RNAs. RNA Biol. 10:924–933. 2013.
View Article : Google Scholar
|
|
27
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen FJ, Sun M, Li SQ, Wu QQ, Ji L, Liu
ZL, Liu ZL, Zhou GZ, Cao G, Jin L, et al: Upregulation of the long
non-coding RNA HOTAIR promotes esophageal squamous cell carcinoma
metastasis and poor prognosis. Mol Carcinog. 52:908–915. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu JH, Chen G, Dang YW, Li CJ and Luo DZ:
Expression and prognostic significance of lncRNA MALAT-1 in
pancreatic cancer tissues. Asian Pac J Cancer Prev. 15:2971–2977.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang Q, Geng PL, Yin P, Wang XL, Jia JP
and Yao J: Down-regulation of long non-coding RNA TUG1 inhibits
osteosarcoma cell proliferation and promotes apoptosis. Asian Pac J
Cancer Prev. 14:2311–2315. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kleist B, Bankau A, Lorenz G, Jng KA and
Poetsch M: Different risk factors in basaloid and common squamous
head and neck cancer. Laryngoscope. 114:1063–1068. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hu L, Wu Y, Tan D, Meng H, Wang K and Bai
Y: Up-regulation of long noncoding RNA MALAT-1 contributes to
proliferation and metastasis in esophageal squamous cell carcinoma.
J Exp Clin Cancer Res. 34:72015. View Article : Google Scholar : PubMed/NCBI
|