The earliest finding of a circadian clock was the
change in position of plant leaves, which spread during the day and
droop at night, corresponding to an oscillation with a 24-h period
(1,2).
Subsequently, circadian clocks were also identified in the form of
clear circadian rhythms in the eclosion of insects (3–6),
hibernation of animals (7–9), and body temperature, blood pressure and
pulse in humans (10–13). The circadian clock is an inherent
rhythm developed by life on the earth's surface during the
long-term evolutionary process to adapt to ambient and external
environments (particularly, to the sunrise and sunset) (14,15).
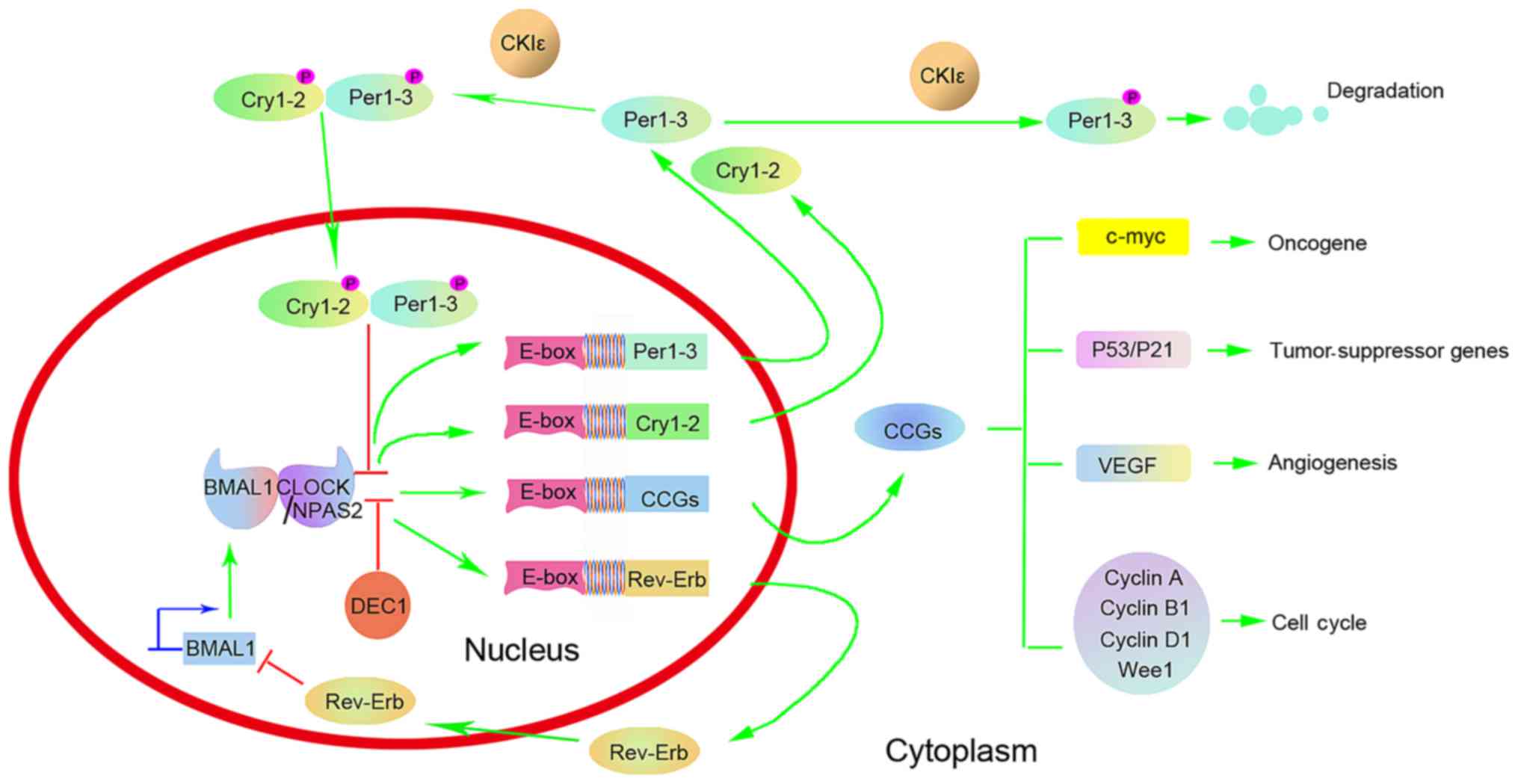
Multiple clock genes, including circadian locomotor
output cycles kaput (CLOCK), brain and muscle arylhydrocarbon
receptor nuclear translocator (ARNT)-like 1 (BMAL1), period (Per)1,
Per2, Per3, cryptochrome (Cry)1, Cry2, neuronal Per-Arnt-Sim (PAS)
domain protein 2 (NPAS2), casein kinase Iε (CKIε), timeless (Tim),
nuclear receptor subfamily 1, group D, member 1 (NR1D1, also known
as Rev-Erb-α) and differentiated embryo-chondrocyte expressed gene
(DEC), accurately regulate the human circadian clock at the
molecular level (16–18). These genes constitute two important
feedback loops. CLOCK is the core factor of the circadian clock and
combines with BMAL1 to form a heterodimer through its basic
helix-loop-helix (bHLH)-PAS structural domain. The heterodimer
combines with the E-box on the promoter of the Per1-3 and Cry1-2
genes, and activates their transcription. The coding products, the
Per1-3 and Cry1-2 proteins, are transported from the cytoplasm to
the nucleus, where they directly combine with CLOCK/BMAL1, which
inhibits their activities and further blocks the transcription of
Per1-3 and Cry1-2. In addition to activating the transcription of
Per1-3 and Cry1-2, the CLOCK/BMAL1 heterodimer also activates the
transcription of the orphan nuclear receptor Rev-Erb gene (17,19)
(Fig. 1). The protein encoded by the
Rev-Erb gene can combine with the BMAL1 promoter and block its
transcription (17). Since genetic
transcription, translation and protein transport from the cytoplasm
to the nucleus lasts a certain time, the oscillation of the
biological rhythm proceeds with a periodic length of ~24 h via
self-induction (18,19). Such a negative feedback cycle of the
clock genes forms a precise endogenous ‘molecular clock’ in the
body. Clock genes output the rhythm signal of a circadian clock
through downstream clock controlled genes (CCGs). Thereby,
molecular activity within the cell also exhibits a temporal rhythm
(18,19).
In May 1997, the Takahashi research group of
Northwestern University (Evanston, USA) successfully cloned the
murine CLOCK gene (20). This
represented a milestone in the study of the molecular mechanism of
circadian clocks in mammals. In 1999, this group reported the
cloning of the human CLOCK gene, which is located on the long arm
of chromosome 4 (4q12) and comprises a protein-coding sequence of
2,538 bp. The CLOCK gene belongs to the bHLH-PAS family of
transcriptional regulatory factors (21). The containing bHLH domain participates
in protein-protein interactions for the formation of protein dimers
(21). Two PAS structural domains
(PAS-A and PAS-B) mediate the combination of the protein with DNA.
Furthermore, the glutamine-rich C-terminus of the CLOCK protein
also participates in transcriptional activation (21). The CLOCK gene is a necessary regulator
of the circadian rhythm and serves a central role in the circadian
clock system. Homozygote mice with CLOCK mutations develop both
circadian clock rhythm and feeding rhythm disorders (22,23).
NPAS2, also known as member of PAS protein 4 (MOP4),
is located on the human chromosome 2p11.2–2q13. Similar to CLOCK,
NPAS2 also belongs to the bHLH-PAS family. NPAS2 exhibits the bHLH
structural domain at its N-terminus, and two PAS structural domains
(PAS-A and PAS-B) in addition to a nuclear receptor-joining region
at its C-terminus (39). NPAS2 can
regulate the circadian clock rhythm by forming an NPAS/BMAL1
heterodimer with BMAL1, combining with the target gene promoter
E-box, and regulating the expression of the Per and Cry genes
(40). NPAS2 is an essential gene to
maintain a normal biological rhythm. Disorders of the circadian
rhythm could be caused by mutation or deletion of NPAS2 (40). In addition, NPAS2 also regulates and
interferes with oncogenes, tumor-suppressor genes, and genes
associated with the cell cycle, cell proliferation and apoptosis
(41–43). Furthermore, NPAS2 is important in cell
cycle regulation, DNA damage repair response and tumor growth
inhibition, and may also act as a tumor-suppressor gene (41–43).
Rev-Erb-α (identified in 1989) and Rev-Erb-β
(identified in 1994) are members of the nuclear receptor
superfamily of ligand-inducible transcription factors (48,49). Both
receptors possess a DNA-binding domain with a conservative zinc
finger and a ligand-binding domain. The DNA-binding domain contains
the sequence coding the nuclear localization signal. Depending on
the circadian rhythm, these domains are expressed in the human
supraoptic nuclei, liver and heart (48,49).
Rev-Erb-α can inhibit the expression of CLOCK (50), BMAL1 (51) and NPAS2 (52). The SCN of Rev-Erb-α knockout mice do
not periodically express BMAL1 and their active phase is shortened.
This indicates that Rev-Erb-α is required for maintaining the
accuracy of the circadian clock (53). A previous study indicated that
Rev-Erb-α and Rev-Erb-β coordinated to protect against major
perturbations in circadian and metabolic physiology (54). The periodic expression of the core
circadian clock and the lipid metabolism network were observed to
be markedly dysregulated in Rev-Erb-α and Rev-Erb-β knockout mice,
which indicates that Rev-Erb-α and Rev-Erb-β are also important
components of the circadian clock core mechanism (17,55,56).
In recent years, due to the accelerated pace of life
and an increased pressure for competition, a large number of people
stay awake until late, lose sleep and miss meals, causing a
circadian clock disorder and an increase in circadian clock
disorder-related diseases (69–71).
Epidemiologic studies revealed that circadian rhythm disorders
(mainly caused by the influence of light) are correlated with
breast, ovarian and prostate cancer. Working on night or rotating
shifts is linked to a greatly increased risk for women to develop
breast and ovarian cancer, and for men to develop prostate cancer
(69–71). Clock genes contribute to the
occurrence and development of tumors by regulating and interfering
with oncogenes (c-myc), tumor-suppressor genes (P53 and P21), genes
involved in the regulation of the cell cycle (cyclins A, B1 and D1,
and WEE1 G2 checkpoint kinase) and vascular endothelial growth
factor, as well as affecting the internal secretion pathway
(72–81) (Fig. 1).
These target genes regulated by the biological clock genes are
involved in DNA damage repair, cell proliferation and apoptosis.
Thus, biological clock disorders are likely to lead to uncontrolled
cell growth and malignant transformation (73).
Although the exact association between clock genes
and common abdominal malignant tumors, including liver cancer
(82,83), colorectal cancer (84–92),
gastric cancer (93,94) and pancreatic cancer (95), is not clear yet, it has been
demonstrated that an abnormal expression of clock genes is
ubiquitous in these tumors. Abnormal expression of the CLOCK gene
may be one of the important reasons for occurrence and development
of these tumors. The relevant articles are summarized in Tables I and II.
As shown in these tables, only a few articles focus
on clock genes in abdominal tumors. The majority of them are
single-center and small-sample studies, mainly focusing on colon
cancer and genes such as CLOCK, BMAL1, Per1, Per2, Per3, Cry1,
Cry2, CKIε and Tim, whereas only a few studies focus on NPAS2,
Rev-Erb and DEC (83–91,93–95). Low
expression of Per1 and Per3 in liver, colon and pancreatic cancer
has been observed, and Per1 and Per3 are closely associated with
prognosis (83–91,93–95)
(Tables I and II).
Currently, the reason and mechanism of low
expression of clock genes in abdominal tumors are not clear.
Preliminary studies indicate that, in liver cancer, hypoxia,
hypoxia inducible factor (HIF)-1α, HIF-2α and hepatitis B virus X
protein (HBx) can disrupt the expression of circadian clock genes
(83,96). Besides HBx, hepatitis C virus can also
modulate the hepatic clock gene machinery (97). Therefore, it can be hypothesized that
the tumor microenvironment and virus infections may contribute to
circadian clock disorders in hepatocellular carcinoma cells
(83,96).
The new interdiscipline generated by the integration
of chronobiology and onco-molecular biology is expected to expand
the knowledge about tumor occurrence and development, and may
provide a new approach for tumor therapy (98–102).
Tumor chronotherapy, which is the selection of the optimum
treatment time to achieve the maximum curative effect and the
minimum toxic and side effects based on the rhythm characteristics
of tumor growth, has achieved satisfactory results in clinical
practice (98–102). However, the association between
clock genes and tumors remains to be fully understood. The
circadian clock system of Drosophila is well understood, but
this knowledge cannot be completely transferred to the human
circadian clock, as this is more complex than that of
Drosophila and large individual differences exist. Numerous
factors in the natural and social environments that can influence
the human circadian clock and the formation of tumors have not yet
been fully elucidated. However, future findings in this field will
lead to an increased knowledge in the disciplines of tumor and
circadian clock research.
This study was supported by the National Natural
Science Foundation of China (grant no. 81702885), the Fundamental
Research Funds for the Central Universities: the Independent
Innovation Fund of Huazhong University of Science and Technology
(HUST: 2016YXMS241), Chinese Foundation for Hepatitis Prevention
and Control ‘Tian Qing’ Liver Research Fund and China Postdoctoral
Science Foundation funded project (2017M613001).
|
1
|
Montenegro-Montero A, Canessa P and
Larrondo LF: Around the fungal clock: Recent advances in the
molecular study of circadian clocks in neurospora and other fungi.
Adv Genet. 92:107–184. 2015.PubMed/NCBI
|
|
2
|
Endo M: Tissue-specific circadian clocks
in plants. Curr Opin Plant Biol. 29:44–49. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Di Cara F and King-Jones K: How clocks and
hormones act in concert to control the timing of insect
development. Curr Top Dev Biol. 105:1–36. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tomioka K: Chronobiology of crickets: A
review. Zoolog Sci. 31:624–632. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Numata H, Miyazaki Y and Ikeno T: Common
features in diverse insect clocks. Zoological Lett. 1:102015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Uryu O, Ameku T and Niwa R: Recent
progress in understanding the role of ecdysteroids in adult
insects: Germline development and circadian clock in the fruit fly
Drosophila melanogaster. Zoological Lett. 1:322015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Heller HC and Ruby NF: Sleep and circadian
rhythms in mammalian torpor. Annu Rev Physiol. 66:275–289. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ruby NF: Hibernation: When good clocks go
cold. J Biol Rhythms. 18:275–286. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Coomans CP, Ramkisoensing A and Meijer JH:
The suprachiasmatic nuclei as a seasonal clock. Front
Neuroendocrinol. 37:29–42. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Leslie M: Circadian rhythms. Sleep study
suggests triggers for diabetes and obesity. Science. 336:1432012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bass J: Circadian topology of metabolism.
Nature. 491:348–356. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Richards J, Diaz AN and Gumz ML: Clock
genes in hypertension: Novel insights from rodent models. Blood
Press Monit. 19:249–254. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
McLoughlin SC, Haines P and FitzGerald GA:
Clocks and cardiovascular function. Methods Enzymol. 552:211–228.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Robinson I and Reddy AB: Molecular
mechanisms of the circadian clockwork in mammals. FEBS Lett.
588:2477–2483. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li S and Zhang L: Circadian control of
global transcription. Biomed Res Int. 2015:1878092015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang N, Chelliah Y, Shan Y, Taylor CA,
Yoo SH, Partch C, Green CB, Zhang H and Takahashi JS: Crystal
structure of the heterodimeric CLOCK:BMAL1 transcriptional
activator complex. Science. 337:189–194. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cho H, Zhao X, Hatori M, Yu RT, Barish GD,
Lam MT, Chong LW, DiTacchio L, Atkins AR, Glass CK, et al:
Regulation of circadian behaviour and metabolism by REV-ERB-α and
REV-ERB-β. Nature. 485:123–127. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bersten DC, Sullivan AE, Peet DJ and
Whitelaw ML: bHLH-PAS proteins in cancer. Nat Rev Cancer.
13:827–841. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mazzoccoli G, Pazienza V and Vinciguerra
M: Clock genes and clock-controlled genes in the regulation of
metabolic rhythms. Chronobiol Int. 29:227–251. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
King DP, Zhao Y, Sangoram AM, Wilsbacher
LD, Tanaka M, Antoch MP, Steeves TD, Vitaterna MH, Kornhauser JM,
Lowrey PL, et al: Positional cloning of the mouse circadian clock
gene. Cell. 89:641–653. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Steeves TD, King DP, Zhao Y, Sangoram AM,
Du F, Bowcock AM, Moore RY and Takahashi JS: Molecular cloning and
characterization of the human CLOCK gene: Expression in the
suprachiasmatic nuclei. Genomics. 57:189–200. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Naylor E, Bergmann BM, Krauski K, Zee PC,
Takahashi JS, Vitaterna MH and Turek FW: The circadian clock
mutation alters sleep homeostasis in the mouse. J Neurosci.
20:8138–8143. 2000.PubMed/NCBI
|
|
23
|
Lee J, Lee S, Chung S, Park N, Son GH, An
H, Jang J, Chang DJ, Suh YG and Kim K: Identification of a novel
circadian clock modulator controlling BMAL1 expression through a
ROR/REV-ERB-response element-dependent mechanism. Biochem Biophys
Res Commun. 469:580–586. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ikeda M and Nomura M: cDNA cloning and
tissue-specific expression of a novel basic helix-loop-helix/PAS
protein (BMAL1) and identification of alternatively spliced
variants with alternative translation initiation site usage.
Biochem Biophys Res Commun. 233:258–264. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bunger MK, Wilsbacher LD, Moran SM,
Clendenin C, Radcliffe LA, Hogenesch JB, Simon MC, Takahashi JS and
Bradfield CA: Mop3 is an essential component of the master
circadian pacemaker in mammals. Cell. 103:1009–1017. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Young ME, Brewer RA, Peliciari-Garcia RA,
Collins HE, He L, Birky TL, Peden BW, Thompson EG, Ammons BJ, Bray
MS, et al: Cardiomyocyte-specific BMAL1 plays critical roles in
metabolism, signaling, and maintenance of contractile function of
the heart. J Biol Rhythms. 29:257–276. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kennaway DJ, Varcoe TJ, Voultsios A and
Boden MJ: Global loss of bmal1 expression alters adipose tissue
hormones, gene expression and glucose metabolism. PLoS One.
8:e652552013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rudic RD, McNamara P, Curtis AM, Boston
RC, Panda S, Hogenesch JB and Fitzgerald GA: BMAL1 and CLOCK, two
essential components of the circadian clock, are involved in
glucose homeostasis. PLoS Biol. 2:e3772004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Khapre RV, Kondratova AA, Patel S,
Dubrovsky Y, Wrobel M, Antoch MP and Kondratov RV: BMAL1-dependent
regulation of the mTOR signaling pathway delays aging. Aging
(Albany NY). 6:48–57. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ali AA, Schwarz-Herzke B, Stahr A,
Prozorovski T, Aktas O and von Gall C: Premature aging of the
hippocampal neurogenic niche in adult Bmal1-deficient mice. Aging
(Albany NY). 7:435–449. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Konopka RJ and Benzer S: Clock mutants of
Drosophila melanogaster. Proc Natl Acad Sci USA.
68:2112–2116. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lengyel Z, Lovig C, Kommedal S, Keszthelyi
R, Szekeres G, Battyáni Z, Csernus V and Nagy AD: Altered
expression patterns of clock gene mRNAs and clock proteins in human
skin tumors. Tumour Biol. 34:811–819. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao N, Yang K, Yang G, Chen D, Tang H,
Zhao D and Zhao C: Aberrant expression of clock gene period1 and
its correlations with the growth, proliferation and metastasis of
buccal squamous cell carcinoma. PLoS One. 8:e558942013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hsu CM, Lin PM, Lai CC, Lin HC, Lin SF and
Yang MY: PER1 and CLOCK: Potential circulating biomarkers for head
and neck squamous cell carcinoma. Head Neck. 36:1018–1026. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cadenas C, van de Sandt L, Edlund K, Lohr
M, Hellwig B, Marchan R, Schmidt M, Rahnenführer J, Oster H and
Hengstler JG: Loss of circadian clock gene expression is associated
with tumor progression in breast cancer. Cell Cycle. 13:3282–3291.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu B, Xu K, Jiang Y and Li X: Aberrant
expression of Per1, Per2 and Per3 and their prognostic relevance in
non-small cell lung cancer. Int J Clin Exp Pathol. 7:7863–7871.
2014.PubMed/NCBI
|
|
37
|
Larkin JC and Woolford JL Jr: Molecular
cloning and analysis of the CRY1 gene: A yeast ribosomal protein
gene. Nucleic Acids Res. 11:403–420. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kume K, Zylka MJ, Sriram S, Shearman LP,
Weaver DR, Jin X, Maywood ES, Hastings MH and Reppert SM: mCRY1 and
mCRY2 are essential components of the negative limb of the
circadian clock feedback loop. Cell. 98:193–205. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhou YD, Barnard M, Tian H, Li X, Ring HZ,
Francke U, Shelton J, Richardson J, Russell DW and McKnight SL:
Molecular characterization of two mammalian bHLH-PAS domain
proteins selectively expressed in the central nervous system. Proc
Natl Acad Sci USA. 94:713–718. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
McNamara P, Seo SB, Rudic RD, Sehgal A,
Chakravarti D and FitzGerald GA: Regulation of CLOCK and MOP4 by
nuclear hormone receptors in the vasculature: A humoral mechanism
to reset a peripheral clock. Cell. 105:877–889. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yuan P, Wang S, Zhou F, Wan S, Yang Y,
Huang X, Zhang Z, Zhu Y, Zhang H and Xing J: Functional
polymorphisms in the NPAS2 gene are associated with overall
survival in transcatheter arterial chemoembolization-treated
hepatocellular carcinoma patients. Cancer Sci. 105:825–832. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xue X, Liu F, Han Y, Li P, Yuan B, Wang X,
Chen Y, Kuang Y, Zhi Q and Zhao H: Silencing NPAS2 promotes cell
growth and invasion in DLD-1 cells and correlated with poor
prognosis of colorectal cancer. Biochem Biophys Res Commun.
450:1058–1062. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Rana S, Shahid A, Ullah H and Mahmood S:
Lack of association of the NPAS2 gene Ala394Thr polymorphism
(rs2305160:G>A) with risk of chronic lymphocytic leukemia. Asian
Pac J Cancer Prev. 15:7169–7174. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fish KJ, Cegielska A, Getman ME, Landes GM
and Virshup DM: Isolation and characterization of human casein
kinase I epsilon (CKI), a novel member of the CKI gene family. J
Biol Chem. 270:14875–14883. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Camacho F, Cilio M, Guo Y, Virshup DM,
Patel K, Khorkova O, Styren S, Morse B, Yao Z and Keesler GA: Human
casein kinase Idelta phosphorylation of human circadian clock
proteins period 1 and 2. FEBS Lett. 489:159–165. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tsuchiya Y, Akashi M, Matsuda M, Goto K,
Miyata Y, Node K and Nishida E: Involvement of the protein kinase
CK2 in the regulation of mammalian circadian rhythms. Sci Signal.
2:ra262009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lee H, Chen R, Lee Y, Yoo S and Lee C:
Essential roles of CKIdelta and CKIepsilon in the mammalian
circadian clock. Proc Natl Acad Sci USA. 106:21359–21364. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bonnelye E, Vanacker JM, Desbiens X, Begue
A, Stehelin D and Laudet V: Rev-erb beta, a new member of the
nuclear receptor superfamily, is expressed in the nervous system
during chicken development. Cell Growth Differ. 5:1357–1365.
1994.PubMed/NCBI
|
|
49
|
Lazar MA, Hodin RA, Darling DS and Chin
WW: A novel member of the thyroid/steroid hormone receptor family
is encoded by the opposite strand of the rat c-erbA alpha
transcriptional unit. Mol Cell Biol. 9:1128–1136. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Crumbley C and Burris TP: Direct
regulation of CLOCK expression by REV-ERB. PLoS One. 6:e172902011.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Dardente H, Fustin JM and Hazlerigg DG:
Transcriptional feedback loops in the ovine circadian clock. Comp
Biochem Physiol A Mol Integr Physiol. 153:391–398. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Crumbley C, Wang Y, Kojetin DJ and Burris
TP: Characterization of the core mammalian clock component, NPAS2,
as a REV-ERBalpha/RORalpha target gene. J Biol Chem.
285:35386–35392. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Takeda Y, Kang HS, Angers M and Jetten AM:
Retinoic acid-related orphan receptor gamma directly regulates
neuronal PAS domain protein 2 transcription in vivo. Nucleic Acids
Res. 39:4769–4782. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Bugge A, Feng D, Everett LJ, Briggs ER,
Mullican SE, Wang F, Jager J and Lazar MA: Rev-erbα and Rev-erbβ
coordinately protect the circadian clock and normal metabolic
function. Genes Dev. 26:657–667. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Mazzoccoli G, Cai Y, Liu S, Francavilla M,
Giuliani F, Piepoli A, Pazienza V, Vinciguerra M, Yamamoto T and
Takumi T: REV-ERBα and the clock gene machinery in mouse peripheral
tissues: A possible role as a synchronizing hinge. J Biol Regul
Homeost Agents. 26:265–276. 2012.PubMed/NCBI
|
|
56
|
Bhargava A, Herzel H and
Ananthasubramaniam B: Mining for novel candidate clock genes in the
circadian regulatory network. BMC Syst Biol. 9:782015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Shen M, Kawamoto T, Yan W, Nakamasu K,
Tamagami M, Koyano Y, Noshiro M and Kato Y: Molecular
characterization of the novel basic helix-loop-helix protein DEC1
expressed in differentiated human embryo chondrocytes. Biochem
Biophys Res Commun. 236:294–298. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Fujimoto K, Shen M, Noshiro M, Matsubara
K, Shingu S, Honda K, Yoshida E, Suardita K, Matsuda Y and Kato Y:
Molecular cloning and characterization of DEC2, a new member of
basic helix-loop-helix proteins. Biochem Biophys Res Commun.
280:164–171. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Noshiro M, Furukawa M, Honma S, Kawamoto
T, Hamada T, Honma K and Kato Y: Tissue-specific disruption of
rhythmic expression of Dec1 and Dec2 in clock mutant mice. J Biol
Rhythms. 20:404–418. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wu T, Ni Y, Zhuge F, Sun L, Xu B, Kato H
and Fu Z: Significant dissociation of expression patterns of the
basic helix-loop-helix transcription factors Dec1 and Dec2 in rat
kidney. J Exp Biol. 214:1257–1263. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Honma S, Kawamoto T, Takagi Y, Fujimoto K,
Sato F, Noshiro M, Kato Y and Honma K: Dec1 and Dec2 are regulators
of the mammalian molecular clock. Nature. 419:841–844. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Li Y, Xie M, Song X, Gragen S, Sachdeva K,
Wan Y and Yan B: DEC1 negatively regulates the expression of DEC2
through binding to the E-box in the proximal promoter. J Biol Chem.
278:16899–16907. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Seino H, Wu Y, Morohashi S, Kawamoto T,
Fujimoto K, Kato Y, Takai Y and Kijima H: Basic helix-loop-helix
transcription factor DEC1 regulates the cisplatin-induced apoptotic
pathway of human esophageal cancer cells. Biomed Res. 36:89–96.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Jinhua H, Zhao M, Wei S, Haitao Y, Yuwen
W, Lili W, Wei L and Jian Y: Down regulation of differentiated
embryo-chondrocyte expressed gene 1 is related to the decrease of
osteogenic capacity. Curr Drug Targets. 15:432–441. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
You J, Lin L, Liu Q, Zhu T, Xia K and Su
T: The correlation between the expression of differentiated
embryo-chondrocyte expressed gene l and oral squamous cell
carcinoma. Eur J Med Res. 19:212014. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Bi H, Li S, Qu X, Wang M, Bai X, Xu Z, Ao
X, Jia Z, Jiang X, Yang Y and Wu H: DEC1 regulates breast cancer
cell proliferation by stabilizing cyclin E protein and delays the
progression of cell cycle S phase. Cell Death Dis. 6:e18912015.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Sehgal A, Price JL, Man B and Young MW:
Loss of circadian behavioral rhythms and per RNA oscillations in
the Drosophila mutant timeless. Science. 263:1603–1606.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Mazzoccoli G, Laukkanen MO, Vinciguerra M,
Colangelo T and Colantuoni V: A timeless link between circadian
patterns and disease. Trends Mol Med. 22:68–81. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Parent MÉ, El-Zein M, Rousseau MC, Pintos
J and Siemiatycki J: Night work and the risk of cancer among men.
Am J Epidemiol. 176:751–759. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Bhatti P, Cushing-Haugen KL, Wicklund KG,
Doherty JA and Rossing MA: Nightshift work and risk of ovarian
cancer. Occup Environ Med. 70:231–237. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Ijaz S, Verbeek J, Seidler A, Lindbohm ML,
Ojajärvi A, Orsini N, Costa G and Neuvonen K: Night-shift work and
breast cancer-a systematic review and meta-analysis. Scand J Work
Environ Health. 39:431–447. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhao L, Isayama K, Chen H, Yamauchi N,
Shigeyoshi Y, Hashimoto S and Hattori MA: The nuclear receptor
REV-ERBα represses the transcription of growth/differentiation
factor 10 and 15 genes in rat endometrium stromal cells. Physiol
Rep. 4:pii:e126632016. View Article : Google Scholar
|
|
73
|
Canaple L, Kakizawa T and Laudet V: The
days and nights of cancer cells. Cancer Res. 63:7545–7552.
2003.PubMed/NCBI
|
|
74
|
Schibler U: The daily timing of gene
expression and physiology in mammals. Dialogues Clin Neurosci.
9:257–272. 2007.PubMed/NCBI
|
|
75
|
Kornmann B, Schaad O, Bujard H, Takahashi
JS and Schibler U: System-driven and oscillator-dependent circadian
transcription in mice with a conditionally active liver clock. PLoS
Biol. 5:e342007. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Walker JR and Hogenesch JB: RNA profiling
in circadian biology. Methods Enzymol. 393:366–376. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Storch KF, Lipan O, Leykin I, Viswanathan
N, Davis FC, Wong WH and Weitz CJ: Extensive and divergent
circadian gene expression in liver and heart. Nature. 417:78–83.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Panda S, Antoch MP, Miller BH, Su AI,
Schook AB, Straume M, Schultz PG, Kay SA, Takahashi JS and
Hogenesch JB: Coordinated transcription of key pathways in the
mouse by the circadian clock. Cell. 109:307–320. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
McCarthy JJ, Andrews JL, McDearmon EL,
Campbell KS, Barber BK, Miller BH, Walker JR, Hogenesch JB,
Takahashi JS and Esser KA: Identification of the circadian
transcriptome in adult mouse skeletal muscle. Physiol Genomics.
31:86–95. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Kornmann B, Preitner N, Rifat D,
Fleury-Olela F and Schibler U: Analysis of circadian liver gene
expression by ADDER, a highly sensitive method for the display of
differentially expressed mRNAs. Nucleic Acids Res. 29:E51–E61.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Duffield GE, Best JD, Meurers BH, Bittner
A, Loros JJ and Dunlap JC: Circadian programs of transcriptional
activation, signaling, and protein turnover revealed by microarray
analysis of mammalian cells. Curr Biol. 12:551–557. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Lin YM, Chang JH, Yeh KT, Yang MY, Liu TC,
Lin SF, Su WW and Chang JG: Disturbance of circadian gene
expression in hepatocellular carcinoma. Mol Carcinog. 47:925–933.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Yang SL, Yu C, Jiang JX, Liu LP, Fang X
and Wu C: Hepatitis B virus X protein disrupts the balance of the
expression of circadian rhythm genes in hepatocellular carcinoma.
Oncol Lett. 8:2715–2720. 2014.PubMed/NCBI
|
|
84
|
Krugluger W, Brandstaetter A, Kállay E,
Schueller J, Krexner E, Kriwanek S, Bonner E and Cross HS:
Regulation of genes of the circadian clock in human colon cancer:
Reduced period-1 and dihydropyrimidine dehydrogenase transcription
correlates in high-grade tumors. Cancer Res. 67:7917–7922. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wang Y, Hua L, Lu C and Chen Z: Expression
of circadian clock gene human Period2 (hPer2) in human colorectal
carcinoma. World J Surg Oncol. 9:1662011. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Mazzoccoli G, Panza A, Valvano MR, Palumbo
O, Carella M, Pazienza V, Biscaglia G, Tavano F, Di Sebastiano P,
Andriulli A and Piepoli A: Clock gene expression levels and
relationship with clinical and pathological features in colorectal
cancer patients. Chronobiol Int. 28:841–851. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Oshima T, Takenoshita S, Akaike M,
Kunisaki C, Fujii S, Nozaki A, Numata K, Shiozawa M, Rino Y, Tanaka
K, et al: Expression of circadian genes correlates with liver
metastasis and outcomes in colorectal cancer. Oncol Rep.
25:1439–1446. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Wang X, Yan D, Teng M, Fan J, Zhou C, Li
D, Qiu G, Sun X, Li T, Xing T, et al: Reduced expression of PER3 is
associated with incidence and development of colon cancer. Ann Surg
Oncol. 19:3081–3088. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Karantanos T, Theodoropoulos G, Gazouli M,
Vaiopoulou A, Karantanou C, Lymberi M and Pektasides D: Expression
of clock genes in patients with colorectal cancer. Int J Biol
Markers. 28:280–285. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Wang L, Chen B, Wang Y, Sun N, Lu C, Qian
R and Hua L: hClock gene expression in human colorectal carcinoma.
Mol Med Rep. 8:1017–1022. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yu H, Meng X, Wu J, Pan C, Ying X, Zhou Y,
Liu R and Huang W: Cryptochrome 1 overexpression correlates with
tumor progression and poor prognosis in patients with colorectal
cancer. PLoS One. 8:e616792013. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Wang Y, Xing T, Huang L, Song G, Sun X,
Zhong L, Fan J, Yan D, Zhou C, Cui F, et al: Period 1 and estrogen
receptor-beta are downregulated in Chinese colon cancers. Int J
Clin Exp Pathol. 8:8178–8188. 2015.PubMed/NCBI
|
|
93
|
Zhao H, Zeng ZL, Yang J, Jin Y, Qiu MZ, Hu
XY, Han J, Liu KY, Liao JW, Xu RH and Zou QF: Prognostic relevance
of period1 (Per1) and period2 (Per2) expression in human gastric
cancer. Int J Clin Exp Pathol. 7:619–630. 2014.PubMed/NCBI
|
|
94
|
Hu ML, Yeh KT, Lin PM, Hsu CM, Hsiao HH,
Liu YC, Lin HY, Lin SF and Yang MY: Deregulated expression of
circadian clock genes in gastric cancer. BMC Gastroenterol.
14:672014. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Relles D, Sendecki J, Chipitsyna G, Hyslop
T, Yeo CJ and Arafat HA: Circadian gene expression and
clinicopathologic correlates in pancreatic cancer. J Gastrointest
Surg. 17:443–450. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Yu C, Yang SL, Fang X, Jiang JX, Sun CY
and Huang T: Hypoxia disrupts the expression levels of circadian
rhythm genes in hepatocellular carcinoma. Mol Med Rep.
11:4002–4008. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Benegiamo G, Mazzoccoli G, Cappello F,
Rappa F, Scibetta N, Oben J, Greco A, Williams R, Andriulli A,
Vinciguerra M and Pazienza V: Mutual antagonism between circadian
protein period 2 and hepatitis C virus replication in hepatocytes.
PLoS One. 8:e605272013. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Kondratov R: Circadian clock and cancer
therapy: An unexpected journey. Ann Med. 46:189–190. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Innominato PF, Roche VP, Palesh OG,
Ulusakarya A, Spiegel D and Lévi FA: The circadian timing system in
clinical oncology. Ann Med. 46:191–207. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Akgun Z, Saglam S, Yucel S, Gural Z, Balik
E, Cipe G, Yildiz S, Kilickap S, Okyar A and Kaytan-Saglam E:
Neoadjuvant chronomodulated capecitabine with radiotherapy in
rectal cancer: A phase II brunch regimen study. Cancer Chemother
Pharmacol. 74:751–756. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Zarogoulidis P, Darwiche K, Huang H,
Spyratos D, Yarmus L, Li Q, Kakolyris S, Syrigos K and Zarogoulidis
K: Time recall; future concept of chronomodulating chemotherapy for
cancer. Curr Pharm Biotechnol. 14:632–642. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Chen D, Cheng J, Yang K, Ma Y and Yang F:
Retrospective analysis of chronomodulated chemotherapy versus
conventional chemotherapy with paclitaxel, carboplatin, and
5-fluorouracil in patients with recurrent and/or metastatic head
and neck squamous cell carcinoma. Onco Targets Ther. 6:1507–1514.
2013.PubMed/NCBI
|