Introduction
Prostate cancer is one of the most common malignant
tumors in males, the second most frequently diagnosed cancer, and
the sixth leading cause of cancer-associated mortality in the male
population in the USA (1). Typically,
prostate cancer is widely considered to be a tumor of
mild-to-moderate malignancy, with a 5-year survival rate of ~100%
for localized tumors (1). However,
the survival rate decreases to ~27.9% in the event of distant
metastasis (1). Despite various
advancements in therapies, the majority of patients with metastatic
cancer eventually succumb to the disease. These aspects demonstrate
the importance of elucidating the genetic mechanisms underlying
prostate cancer, and of developing novel therapeutic targets.
MicroRNAs (miRNAs) are small (~22 nucleotides)
non-coding RNA molecules that suppress gene expression by
interacting with the 3′-untranslated regions (3′-UTRs) of specific
target mRNAs (2). Distinct miRNA
expression profiles have been identified in human prostate cancer
tissues and cell lines (3–8). One of those miRNAs, miR-193a-3p, was
initially identified by Lagos-Quintana et al (9) in 2003, and was revealed to be a
tumor-suppressor agent (10) as well
as a regulator for transdifferentiation in parietal cells (11). Lin et al (12) conducted a set of microarray assays
(GEO accession: GSE36802), which identified lower expression levels
of miR-193a-3p in prostate cancer tissues compared with that in
benign hyperplasia prostate tissue. However, to the best of our
knowledge, no previous studies have focused on the function or
underlying mechanisms of miR-193a-3p in prostate cancer.
In the present study, the expression of miR-193a-3p
was investigated in the DU-145 and PC3 prostate cancer cell lines.
In addition, a gain-of-function study was performed to assess the
effects of the forced expression of miR-193a-3p in prostate cancer
cells, revealing their ability inhibit cell proliferation by
regulating cyclin D1 (CCND1). These results improve the
current understanding of the underlying mechanisms by which
miR-193a-3p inhibits prostate cancer.
Materials and methods
Reagents, cell culture and
transfection
All oligonucleotide segments were synthesized by
GenePharma Co., Ltd. (Shanghai, China), including an miR-193a-3p
mimic, negative control (NC) duplex and small interfering RNA
(siRNA) against CCND1 (siCCND1). The sequences are
listed in Table I.
 | Table I.Oligonucleotide and primer
sequences. |
Table I.
Oligonucleotide and primer
sequences.
| Name | Sequence
(5′-3′)a |
|---|
| miR-193a-3p mimic
(sense) |
AACUGGCCUACAAAGUCCCAGU |
| NC |
ACUACUGAGUGACAGUAGA |
| siCCND1
(sense) |
GGAGAACAAACAGAUCAUC |
| U6-F |
TGCGGGTGCTCGCTTCGGCAGC |
| miR-193-3p-F |
ACTGGCCTACAAAGTCCCAGT |
| CCND1-F |
GCTGCGAAGTGGAAACCATC |
| CCND1-R |
CCTCCTTCTGCACACATTTGAA |
| GAPDH-F |
ACAACTTTGGTATCGTGGAAGG |
| GAPDH-R |
GCCATCACGCCACAGTTTC |
|
CCND1-UTR-F |
ATTGCAGAGGATGTTCATAAGGCCAGTATGATTTATAAATGCAAT |
|
CCND1-UTR-R |
ATTGCATTTATAAATCATACTGGCCTTATGAACATCCTCTGCAAT |
|
CCND1-Mut-F |
ATTGCAGAGGATGTTCATAACCGGTCAATGATTTATAAATGCAAT |
|
CCND1-Mut-R |
ATTGCATTTATAAATCATTGACCGGTTATGAACATCCTCTGCAAT |
All cell lines used in this study were purchased
from the Shanghai Institute of Biochemistry and Cell Biology of the
Chinese Academy of Sciences (Shanghai, China). The cell lines
included the DU-145 and PC3 human prostate cancer cell lines, as
well as the RWPE normal prostate epithelium cell line. All cell
lines were cultured in RPMI-1640 medium (Takara Biotechnology Co.,
Ltd., Dalian, China) supplemented with 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 50 µg/ml
streptomycin and 50 U/ml penicillin. Bovine pituitary extract (0.05
mg/ml; Thermo Fisher Scientific, Inc.) and human recombinant
epidermal growth factor (5 ng/ml; Thermo Fisher Scientific, Inc.)
were added to the culture medium of the RWPE cells. The cell
culture environment was thermostatic at 37°C with constant humidity
and 5% CO2. The cells were mainly seeded into 6-well
plates at a density of 4×105 cells, lower density was
used depending on certain experiments. Once the cells reached
60–70% confluency, all transfections were performed with Invitrogen
Lipofectamine® 2000 Reagent (Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions, with the
synthesized RNA mimics and NC. Transfected cells were cultured for
48 or 72 h at the same conditions described above.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from prostate cancer cells
and normal cells with standard TRIzol® solution
(Invitrogen; Thermo Fisher Scientific, Inc.). For miRNA expression,
RT reactions were performed with a One Step PrimeScript miRNA cDNA
Synthesis kit (Takara Biotechnology Co., Ltd.), followed by PCR
with SYBR® Premix Ex Taq (Takara Biotechnology Co.,
Ltd). For mRNA, cDNA was synthesized from the total RNA using a
PrimeScript RT Reagent Kit (Takara Biotechnology Co., Ltd.). qPCR
amplification reactions for CCND1 expression were performed
with SYBR® Premix Ex Taq II with ROX (Takara
Biotechnology Co., Ltd.,). For miRNA and mRNA amplifications,
analysis was performed with the ABI 7500 Fast Real-Time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). According to
supplier's instructions, the PCR conditions consisted of 95°C for
30 sec, followed by 40 cycles of amplification (95°C for 3 sec and
60°C for 30 sec). All fold changes were calculated using the
comparative Cq (ΔΔCq) method using GAPDH for normalization
(13). All relevant primer sequences
are listed in Table I, and details of
the reaction mixtures for RT and qPCR are listed in Tables II and III.
| Table II.Reaction mixture for reverse
transcription. |
Table II.
Reaction mixture for reverse
transcription.
| Component | Volume | Final
concentration |
|---|
| 5X PrimeScript RT
Master Mix | 2 µl | 1X |
| Total RNA | 500 ng | – |
| RNase Free
dH2O | ≤10 µl | – |
| Table III.Reaction mixture for quantitative
polymerase chain reaction (total volume, 10 µl). |
Table III.
Reaction mixture for quantitative
polymerase chain reaction (total volume, 10 µl).
| Component | Volume | Final
concentration |
|---|
| SYBR Premix Ex Taq
II | 5 µl | 1X |
| PCR Forward
Primer | 0.4 µl | 0.4 µM |
| PCR Reverse
Primer | 0.4 µl | 0.4 µM |
| ROX Reference
Dye | 0.2 µl | 1X |
| cDNA solution (from
reverse transcription) | 1 µl | – |
|
dH2O | 3 µl | – |
Cell growth and viability assays
DU-145 and PC3 cells (6×103) were seeded
into each well of a 96-well plate. Following an overnight
incubation (37°C with 5% CO2), the cells were
transfected with RNA duplexes (NC, miR-193a-3p or siCCND1)
for 48–72 h as described in the previous section; the concentration
of miR-193a-3p ranged from 0–75 nM. The medium was removed at
various time points, then Cell Counting Kit-8 solution (WST-8;
Dojindo Molecular Technologies, Inc., Kumamoto, Japan) was added to
each well and the cells were incubated at 37°C for 1 h. The
absorbance of the solution was determined spectrophotometrically at
450 nm with an MRX II absorbance reader (Dynex Technologies, Inc.,
Chantilly, VA, USA).
Colony formation assay
The cells were harvested 24 h following RNA
treatment (50 nM NC or 50 nM miR-193a-3p), resuspended in RPMI-1640
supplemented with 10% FBS, and seeded into 6-well plates at a
density of 500 cells/well. The cultures were maintained under
standard conditions (37°C and 5% CO2) for 14 days. The
colonies were then fixed with 50% methanol for 15 min and stained
with 5% crystal violet solution for 20 min. All colonies were
scanned into a computer, then colonies visible to unaided eyes were
counted and plotted for both cell lines.
Cell cycle analysis
Cells were harvested at 48 h following transfection
and fixed in 75% ethanol at −20°C. Following fixing overnight, the
cells were washed twice with phosphate-buffered saline (PBS) and
stained with DNA Prep Stain (Beckman Coulter, Inc., Brea, CA, USA)
for 30 min. Cell cycle analysis was performed using the BD LSRII
Flow Cytometry system with FACSDiva software version 6.0 (BD
Biosciences, Franklin Lakes, NJ, USA). The data were analyzed with
the ModFit LT 5.0 software package (Verity Software House, Topsham,
ME, USA).
Western blot analysis
PC3 and DU-145 cells were lysed with
radioimmunoprecipitation assay buffer solution (Beyotime Institute
of Biotechnology, Nantong, China) following 48 h of transfection on
ice. The supernatants were collected following centrifugation at
13,000 × g at 4°C for 20 min and the protein concentration was
determined using a bicinchoninic acid protein assay kit
(Sigma-Aldrich; Merck Millipore, Darmstadt, Germany). Supernatant
samples (2 µl) were heated at 99°C for 5 min prior to loading and
were separated by 10% sodium dodecyl sulfate (SDS)-polyacrylamide
gels (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The proteins
were transferred to a polyvinylidene difluoride membrane in
transfer buffer containing 48 mmol/l Tris-HCl, 39 mmol/l glycine,
0.037% SDS and 20% methanol at 4°C for 45 min. The membranes were
blocked with 5% non-fat milk for 1 h prior to incubation overnight
with primary antibodies against CCND1 and GAPDH (catalog nos.
EPR2241 and EPR16891; Epitomics; Abcam, Cambridge, MA, USA) at a
dilution of 1:1,000. Following three washes in TBS, the membranes
were then incubated with an horseradish peroxidase-conjugated goat
anti-rabbit secondary antibody (catalog no. 7074; Cell Signaling
Technology, Inc., Boston, MA, USA) at room temperature for 1 h.
Following three further washes in TBS, the proteins were detected
and visualized using an electrochemiluminescence system (Pierce;
Thermo Fisher Scientific, Inc.).
Plasmid construction and luciferase
reporter assay
Bioinformatic screening was performed in order to
initially identify the binding target of miR-193a-3p. TargetScan
(http://www.targetscan.org) analysis
revealed CCND1 as a potential binding site of miR-193a-3p.
Two pairs of oligonucleotide fragments containing the miRNA target
region and mutant miRNA target region of interest were designed and
purchased from Sangon Biotech Co., Ltd. (Shanghai, China; Table I). The oligonucleotide pair was
annealed at 90°C for 3 min and cooled to 37°C for 15 min prior to
being inserted into the pmirGLO Dual-Luciferase miRNA Target
Expression Vector (Promega Corporation, Madison, WI, USA) between
the SacI and SalI sites. All the insertions were confirmed using
sequencing to identify any mutations.
PC3 and DU-145 cells (7×104 cells/well)
were cultured in a 24-well plate for 24 h prior to transfection.
The pmirGLO vectors containing the wild type (Wt) 3′-UTR regions
were co-transfected with miR-193a-3p-expressing or NC vectors. The
co-transfection process was duplicated with vectors including a
mutant (Mut) 3′-UTR region. The cells were harvested following a
24-h incubation at 37°C and luciferase activity was assayed using a
Dual-Luciferase Reporter Assay System (Promega Corporation).
Renilla luciferase (with the cloned 3′-UTR) activity was normalized
to Firefly luciferase activity. Multiple (n≥3) independent
experiments were conducted and the means and standard deviations
were calculated from duplicate wells. The expression of luciferase
was analyzed as described below.
CCND1 rescue experiments
The pIRES-enhanced green fluorescent protein
(EGFP)-CCND1 plasmid (GeneChem Co., Ltd., Shanghai, China)
was constructed following the insertion of the human CCND1
coding sequence into a pIRES-EGFP vector and confirmed by
sequencing. The cells were co-transfected with either miR-193-3p
mimics or NC with pIRES-EGFP-CCND1 or an empty pIRES-EGPF
vector. The cells were harvested at 48 h post-transfection and flow
cytometry was used to analyze the cell cycle. In addition,
CCND1 expression levels were determined by western
blotting.
Statistical analysis
The data are presented as the mean ± standard
deviation of three independent experiments. The variations between
the cell samples were analyzed using a Student's t-test. All data
analysis was performed using Prism version 6 (GraphPad Software,
Inc., La Jolla, CA, USA) for Windows. P<0.05 was considered to
indicate a statistically significant result.
Results
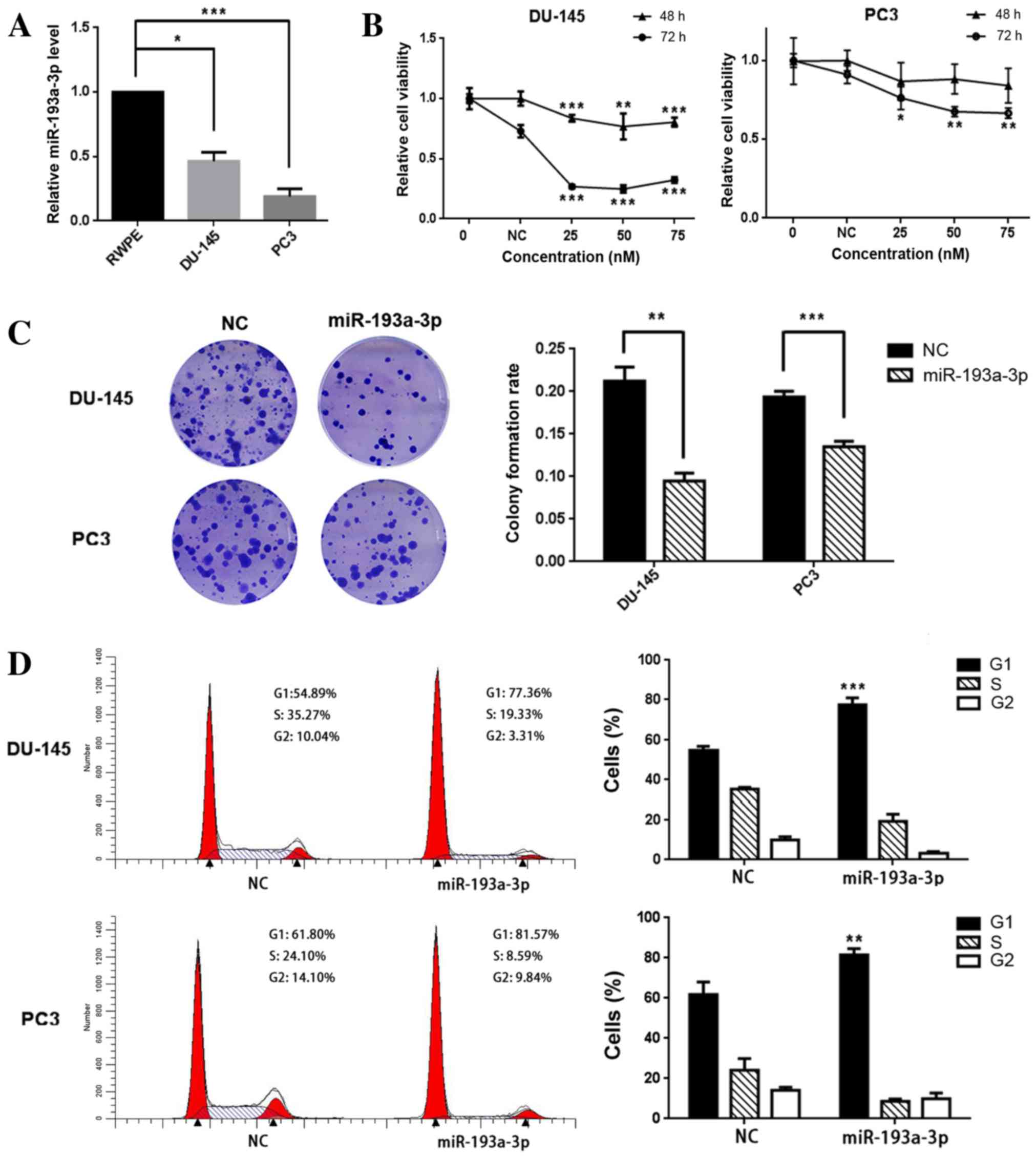
miR-193-3p is downregulated in
prostate cancer cell lines
To investigate the role of miR-193a-3p in human
prostate cancer, RT-qPCR was used to evaluate the gene expression
levels in PC3 and DU-145 cell lines and the RWPE non-malignant cell
line. The results demonstrated that the expression levels of
miR-193a-3p in PC3 and DU-145 cells were significantly reduced,
compared with those in RWPE cells; the relative expression levels
of DU-145 and PC3 were 46.3% (P=0.018) and 19.0% (P<0.001),
respectively, relative to that in RWPE cells (Fig. 1A).
miR-193-3p suppresses prostate cancer
cell proliferation and colony formation, and induces
G1-phase arrest
In order to investigate whether miR-193a-3p
functions as a tumor-suppressor or enhancer, PC3 and DU-145 cells
were transfected with miR-193a-3p mimics. miR-193a-3p transfection
was found to suppress the growth and viability of the prostate
cancer cells (Fig. 1B); compared with
the NC, 25 nM miR-193a-3p was able to reduce PC3 cell viability by
~13% (P=0.083) and ~24% (P=0.021) at 48 and 72 h post-transfection,
respectively, and the corresponding results for DU-145 were ~17%
(P<0.001) and ~63% (P<0.001), respectively. Similarly, the
colony formation ability was inhibited in the two prostate cancer
lines; the colony formation rates of miR-193a-3p-transfected cells
were lower, compared with those in the NC-transfected cells, in the
two cancer cell lines [59.1% (P=0.007) and 31.6% (P<0.001),
respectively; Fig. 1C].
In order to investigate the mechanisms underlying
miR-193a-3p-mediated cell growth suppression, flow cytometry was
used to analyze the distribution of the cell cycle in PC3 and
DU-145 cells following their transfection with miR-193a-3p. The
cell cycle distribution demonstrated a significant
G1-phase arrest in the DU-145 and PC3 cells
[54.89–77.36% (P<0.001) and 61.80–81.57% (P=0.004),
respectively; Fig. 1D]. The results
indicate that miR-193a-3p is able to suppress the growth of
prostate cancer cells by arresting the cells in the G1
phase.
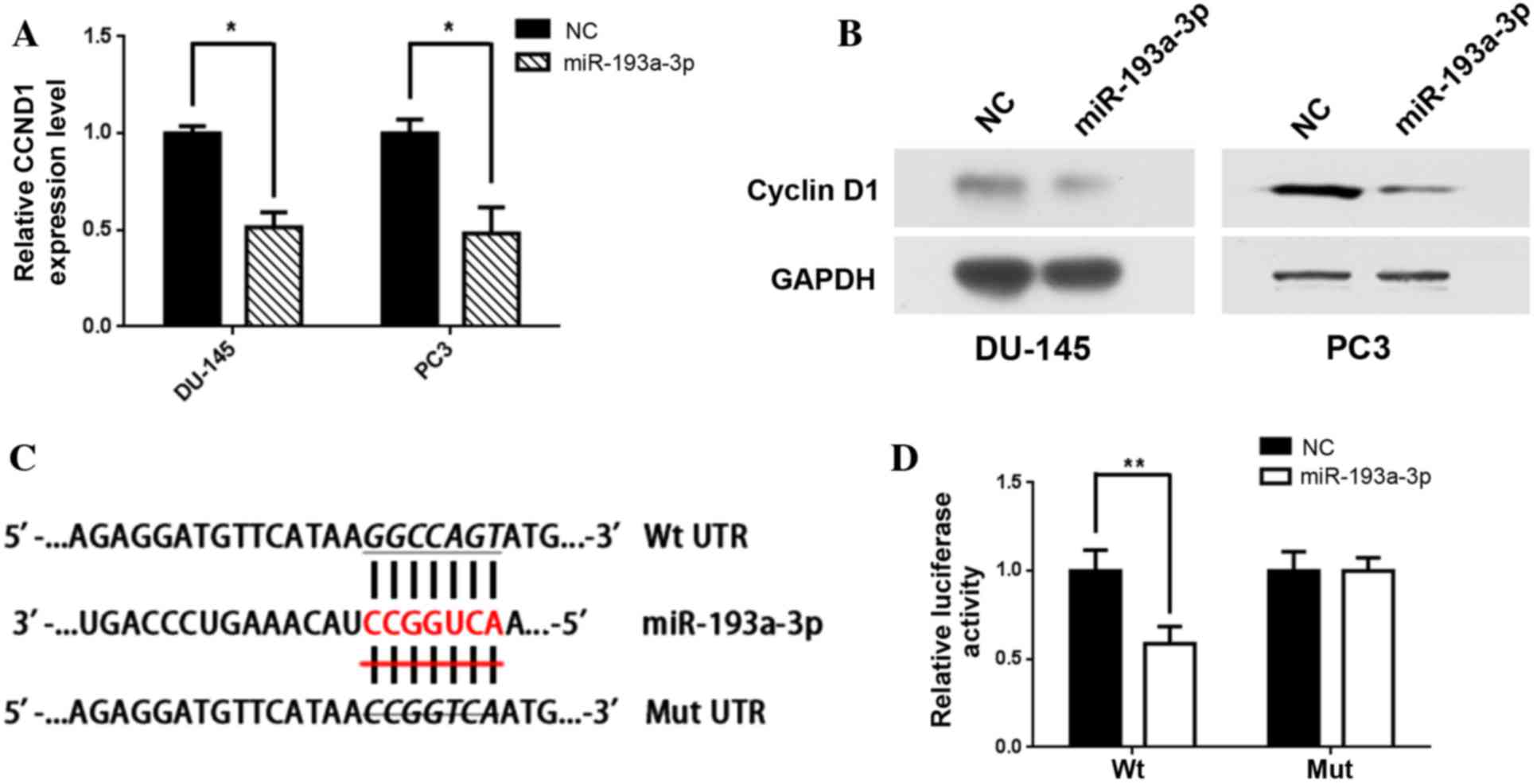
miR-193-3p inhibits the expression of
CCND1
TargetScan provided 221 potential downstream targets
with conserved sites for miR-193a-3p. Among the genes predicted,
CCND1 exhibited a significant decline in mRNA expression
levels in the PC3 and DU-145 cells. Furthermore, RT-qPCR revealed
that the relative mRNA expression levels of CCND1 were
significantly inhibited following transfection with miR-193a-3p as
compared with the NC [relative expression, 0.52 (P=0.03) and 0.49
(P=0.04), respectively; Fig. 2A], and
western blot analysis also indicated that CCND1 protein levels were
markedly suppressed in the cell lines transfected with miR-193a-3p
(Fig. 2B).
In order to determine whether CCND1 is a
direct target of miR-193a-3p, the 3′-UTR of CCND1 was
inserted into a pmirGLO Dual-Luciferase miRNA Target Expression
Vector downstream of the firefly luciferase. A second vector in
which the putative binding site of miR-193a-3p in the 3′-UTR was
mutated was also constructed (Fig.
2C). The results indicated that co-transfection of the Wt
3′-UTR and miR-193a-3p in PC3 cells significantly suppressed
luciferase activity compared with cells co-transfected with the Wt
3′-UTR and NC (P=0.004). However, co-transfection with Mut 3′-UTR
and miR-193a-3p was not observed to alter luciferase activity
(P=0.22) in PC3 cells (Fig. 2D).
Furthermore, the knockdown of CCND1 by
siCCND1 suppressed cell viability in the two cancer cell lines
[reduced by 18.6% in DU-145 (P=0.005) and 18.5% (P=0.013) in PC3;
shown in Fig. 3A]. In addition, cell
colony counts were significantly decreased by siCCND1 transfection
[reduced by 60.6% (P<0.001) in DU-145 and 36.3% (P<0.001) in
PC3, respectively; Fig. 3B] The
results also indicate that siCCND1 may induce G1/S phase arrest,
leading to a significant increase in the proportion of G1 phase
cells (P<0.001 in both cell lines; Fig. 3C). Western blot analysis demonstrated
that siCCND1 had a similar effect to transfection with miR-193a-3p,
inhibiting CCND1 expression (Fig.
3D). The results suggest that the knockdown of CCND1 may
mimic the biological effect of miR-193a-3p.
Restoration of CCND1 expression
partially rescues miR-193-3p-induced cell cycle arrest
To investigate whether forced CCND1
expression is able to counter the cell cycle arrest induced by
miR-193a-3p, the human CCND1 coding sequence was inserted
into a pIRES-EGFP vector. Western blot analysis revealed that CCND1
expression was restored following treatment with the
pIRES-EGFP-CCND1 vector in miR-193a-3p-transfected cells,
compared to treatment with the empty vector (Fig. 4A). Furthermore, the cell cycle was
evaluated by flow cytometry following the co-transfection of PC3
cells with miR-193a-3p mimics or NC, and pIRES-EGFP-CCND1 or
empty pIRES-EGPF vectors. The results demonstrated that restoration
of CCND1 expression is able to partially reverse the
miR-193a-3p-induced cell cycle arrest (Fig. 4B and C).
Discussion
miRNAs have previously been indicated to be
promising diagnostic and prognostic factors in tumor studies
(14), and dysregulated miRNA
expression profiles have been observed in several previous studies
of prostate cancer (15–17). miRNAs have been reported to have vital
roles in prostate cancer cell proliferation, progression and
metastasis (18). Microarray analysis
has indicated that miR-193a-3p is a specific miRNA that has
decreased expression in prostate cancer tissues (GEO accession:
GSE36802). Similar results were obtained in the current study from
quantification experiments in the PC3 and DU-145 prostate cancer
cell lines. However, the specific function of miR-193a-3p in
prostate cancer has yet to be elucidated. Therefore, a series of
experiments were conducted to study the function of miR-193a-3p in
prostate cancer.
In order to identify how prostate cancer cell
function is altered by miR-193a-3p, gain-of-function studies were
conducted in PC3 and DU-145 cells. The results of the cell
viability and colony formation assays demonstrated that
transfection with miR-193a-3p markedly suppressed cell
proliferation in PC3 and DU-145 cells. Furthermore, flow cytometry
revealed both PC3 and DU-145 exhibited significant G1/S
phase arrest following the transfection of miR-193a-3p, which may
be one of the mechanisms underlying miR-193a-3p-mediated growth
inhibition. The results suggested that miR-193a-3p had a profound
effect on the cell cycle, proliferation and survival in prostate
cancer.
Subsequently, the aim of the current study was to
identify the molecular mechanisms underlying the alterations in
cell function induced by miR-193a-3p. RT-qPCR revealed reduced
expression levels of CCND1 and the luciferase assay further
demonstrated that CCND1 is a target of miR-193a-3p.
Furthermore, the knockdown of CCND1 exhibited a similar
effect to the transfection of miR-193a-3p, inducing the inhibition
of cell growth and initiating G1 phase arrest. Restored
CCND1 expression was able to partially reverse the
miR-193a-3p-induced cell cycle arrest. In addition, co-transfection
with pIRES-EGFP-CCND1 partially rescued the
miR-193a-3p-induced G1 phase arrest. Therefore, the
results of the present study indicated that miR-193a-3p is capable
of suppressing the proliferation of prostate cancer cells by
targeting CCND1.
Cyclins are a set of regulatory subunits of
holoenzymes that regulate the progression of cells through the cell
cycle. When it was first discovered in 1991 (19), CCND1 was considered to be a
regulator of the cell cycle. In more recent studies, CCND1
has also been identified as an oncogene that is frequently
dysregulated in breast cancer (20)
and non-Hodgkin's lymphoma (21), as
well as other types of cancer. During normal physiological
functioning, CCND1 serves as a key sensor and integrator of
extracellular signals, mediating cell function through the binding
of cyclin-dependent kinases. CCND1 also induces the
sequential inactivation of the cell cycle-inhibitory function of
retinoblastoma protein (Rb) (22),
which serves as a gatekeeper of the G1 phase in the cell
cycle; passage through this restriction point leads to DNA
synthesis and G1/S phase transition (23). Therefore, inhibiting CCND1
expression by miR-193a-3p transfection may increase the proportion
of prostate cancer cells in the G1 phase, which is
concordant with the results of the present study. However,
CCND1 may have additional oncogenic effects beyond its role
in the cell cycle. Studies in solid tumor models have demonstrated
that CCND1 is able to function as a transcription regulator by
interacting with certain transcription factors, and
chromatin-remodeling and histone-modifying enzymes (22,24,25). CCND1
may also promote chromosome instability by binding to specific
genes that regulate chromosome segregation and chromatin
reorganization (26). Notably, CCND1
has previously been implicated in promoting DNA repair by binding
to recombination-activating gene 1 and by homologous DNA
recombination (27), which is also
considered an anti-apoptotic effect. A previous study reported that
cytoplasmic CCND1 was involved in the estradiol-induced DNA damage
response; CCND1 binds estrogen receptor α at the cytoplasmic
membrane and augments protein kinase B (Akt) phosphorylation
(Ser473) and histone γ H2AX foci formation (28). In the present study, decreased
expression levels of CCND1 were observed following
transfection with miR-193a-3p and the luciferase assay also
identified that miR-193a-3p inhibits CCND1 expression by
directly binding to the 3′-UTR at a post-transcriptional level.
Tian et al (29) suggested
that miR-19b, miR-23b, miR-26a and miR-92a may promote prostate
cancer cell proliferation by co-regulating the expression of
phosphatase and tensin homolog, phosphoinositide 3-kinase/Akt and
CCND1 in vitro. Therefore, the suppression of cell
proliferation by inhibiting CCND1 expression in prostate
cancer is a promising therapeutic target.
In conclusion, the current study identified the
downregulation of miR-193a-3p in certain prostate cancer cell lines
and revealed that miR-193a-3p may function as a potential
tumor-suppressor in prostate cancer, which is able to induce
G1-phase arrest by targeting CCND1. Although
further studies are required to identify other targets of
miR-193a-3p, the results demonstrated that the restoration of
miR-193-3p expression may be a novel therapeutic strategy for
preventing the progression of prostate cancer.
Acknowledgements
This study was supported by grants from the
following institutions: The Qianjiang Talent Project of Technology
Office in Zhejiang Province (grant no. 2013R10045); the Scientific
Research Foundation of the Ministry of Public Health of China
(grant no. WKJ2012-2-009); the Zhejiang Province Key Project of
Science and Technology (grant no. 2014C4008-2); the National
Natural Science Foundation of China (grant nos. 81372773 and
81101717); and the Scientific Research Fund of Zhejiang Provincial
Education Department (grant no. Y201120149).
Glossary
Abbreviations
Abbreviations:
|
miRNA
|
microRNA
|
|
miR-193a-3p
|
microRNA-193a-3p
|
|
CCND1
|
cyclin D1
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
NC
|
negative control
|
|
Wt
|
wild type
|
|
Mut
|
mutant type
|
References
|
1
|
NCI: SEER stat fact sheets: Prostate
cancer. In: Surveillance, epidemiology and end results program.
National Institute of Health. 2013 https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE36802Accessed.
September 4–2013.
|
|
2
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fendler A, Jung M, Stephan C, Erbersdobler
A, Jung K and Yousef GM: The antiapoptotic function of miR-96 in
prostate cancer by inhibition of FOXO1. PLoS One. 8:e808072013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kojima S, Enokida H, Yoshino H, Itesako T,
Chiyomaru T, Kinoshita T, Fuse M, Nishikawa R, Goto Y, Naya Y, et
al: The tumor-suppressive microRNA-143/145 cluster inhibits cell
migration and invasion by targeting GOLM1 in prostate cancer. J Hum
Genet. 59:78–87. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Arora S, Saini S, Fukuhara S, Majid S,
Shahryari V, Yamamura S, Chiyomaru T, Deng G, Tanaka Y and Dahiya
R: MicroRNA-4723 inhibits prostate cancer growth through
inactivation of the abelson family of nonreceptor protein tyrosine
kinases. PLoS One. 8:e780232013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang L, Li B, Li L and Wang T:
MicroRNA-497 suppresses proliferation and induces apoptosis in
prostate cancer cells. Asian Pac J Cancer Prev. 14:3499–3502. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
He L, Yao H, Fan LH, Liu L, Qiu S, Li X,
Gao JP and Hao CQ: MicroRNA-181b expression in prostate cancer
tissues and its influence on the biological behavior of the
prostate cancer cell line PC-3. Genet Mol Res. 12:1012–1021. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mao Y, Chen H, Lin Y, Xu X, Hu Z, Zhu Y,
Wu J, Xu X, Zheng X and Xie L: MicroRNA-330 inhibits cell motility
by downregulating Sp1 in prostate cancer cells. Oncol Rep.
30:327–333. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lagos-Quintana M, Rauhut R, Meyer J,
Borkhardt A and Tuschl T: New microRNAs from mouse and human. RNA.
9:175–179. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kozaki K, Imoto I, Mogi S, Omura K and
Inazawa J: Exploration of tumor-suppressive microRNAs silenced by
DNA hypermethylation in oral cancer. Cancer Res. 68:2094–2105.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kietzmann L, Guhr SS, Meyer TN, Ni L,
Sachs M, Panzer U, Stahl RA, Saleem MA, Kerjaschki D, Gebeshuber CA
and Meyer-Schwesinger C: MicroRNA-193a regulates the
transdifferentiation of human parietal epithelial cells toward a
podocyte phenotype. J Am Soc Nephrol. 26:1389–1401. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin PC, Chiu YL, Banerjee S, Park K,
Mosquera JM, Giannopoulou E, Alves P, Tewari AK, Gerstein MB,
Beltran H, et al: Epigenetic repression of miR-31 disrupts androgen
receptor homeostasis and contributes to prostate cancer
progression. Cancer Res. 73:1232–1244. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Allegra A, Alonci A, Campo S, Penna G,
Petrungaro A, Gerace D and Musolino C: Circulating microRNAs: New
biomarkers in diagnosis, prognosis and treatment of cancer
(review). Int J Oncol. 41:1897–1912. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tay FC, Lim JK, Zhu H, Hin LC and Wang S:
Using artificial microRNA sponges to achieve microRNA
loss-of-function in cancer cells. Adv Drug Deliv Rev. 81:117–127.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wojcicka A, de la Chapelle A and
Jazdzewski K: MicroRNA-related sequence variations in human
cancers. Hum Genet. 133:463–469. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim WT and Kim WJ: MicroRNAs in prostate
cancer. Prostate Int. 1:3–9. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qiang XF, Zhang ZW, Liu Q, Sun N, Pan LL,
Shen J, Li T, Yun C, Li H and Shi LH: MiR-20a promotes prostate
cancer invasion and migration through targeting ABL2. J Cell
Biochem. 115:1269–1276. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Peters G: The D-type cyclins and their
role in tumorigenesis. J Cell Sci Suppl. 18:89–96. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee EY and Muller WJ: Oncogenes and tumor
suppressor genes. Cold Spring Harb Perspect Biol. 2:a0032362010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jares P, Colomer D and Campo E: Molecular
pathogenesis of mantle cell lymphoma. J Clin Invest. 122:3416–3423.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fu M, Wang C, Li Z, Sakamaki T and Pestell
RG: Minireview: Cyclin D1: Normal and abnormal functions.
Endocrinology. 145:5439–5447. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Baldin V, Lukas J, Marcote MJ, Pagano M
and Draetta G: Cyclin D1 is a nuclear protein required for cell
cycle progression in G1. Genes Dev. 7:812–821. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bienvenu F, Jirawatnotai S, Elias JE,
Meyer CA, Mizeracka K, Marson A, Frampton GM, Cole MF, Odom DT,
Odajima J, et al: Transcriptional role of cyclin D1 in development
revealed by a genetic-proteomic screen. Nature. 463:374–378. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aggarwal P, Vaites LP, Kim JK, Mellert H,
Gurung B, Nakagawa H, Herlyn M, Hua X, Rustgi AK, McMahon SB and
Diehl JA: Nuclear cyclin D1/CDK4 kinase regulates CUL4 expression
and triggers neoplastic growth via activation of the PRMT5
methyltransferase. Cancer Cell. 18:329–340. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Casimiro MC, Crosariol M, Loro E, Ertel A,
Yu Z, Dampier W, Saria EA, Papanikolaou A, Stanek TJ, Li Z, et al:
ChIP sequencing of cyclin D1 reveals a transcriptional role in
chromosomal instability in mice. J Clin Invest. 122:833–843. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jirawatnotai S, Hu Y, Michowski W, Elias
JE, Becks L, Bienvenu F, Zagozdzon A, Goswami T, Wang YE, Clark AB,
et al: A function for cyclin D1 in DNA repair uncovered by protein
interactome analyses in human cancers. Nature. 474:230–234. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li Z, Chen K, Jiao X, Wang C, Willmarth
NE, Casimiro MC, Li W, Ju X, Kim SH, Lisanti MP, et al: Cyclin D1
integrates estrogen-mediated DNA damage repair signaling. Cancer
Res. 74:3959–3970. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tian L, Fang YX, Xue JL and Chen JZ: Four
microRNAs promote prostate cell proliferation with regulation of
PTEN and its downstream signals in vitro. PLoS One. 8:e758852013.
View Article : Google Scholar : PubMed/NCBI
|