Introduction
Thyroid cancer is an endocrine malignancy with a
high morbidity rate (1). Anaplastic
thyroid cancer (ATC) is the least common, but most lethal, form of
thyroid cancer. It accounts for 1.3–9.8% of thyroid in worldwide,
with a 1-year survival rate of 20% and a disease-specific mortality
rate of nearly 100% (2). ATC has an
annual incidence of between 1 and 2 cases/million people, with a
female to male ratio of 3:2 (3).
Although the majority of well-differentiated thyroid tumors are
effectively treated by isotope therapy, relapse and poorly
differentiated thyroid cancer, such as anaplastic and medullary
thyroid cancers, show tolerance to isotope treatment (1). Therefore, finding new ways to improve
the effectiveness of isotope-based therapies for this type of
cancer is important.
All-trans-retinoic acid (ATRA) is naturally produced
by humans. ATRA is a natural derivative of vitamin A. ATRA exerts
its functions by triggering G1 phase arrest, affecting DNA
synthesis and enhancing immune cell killing efficiency (4,5). ATRA has
been shown to decrease the proliferation and invasiveness of
different types of cancer cells, including pancreatic cancer
(6), leukemia (7), breast cancer (8) and thyroid cancer cells (9). Particularly, ATRA can significantly
enhance the iodine uptake capability of thyroid tumors and other
cancers (4,5,10,11). However, the mechanisms of the
enhancement of iodine uptake in thyroid cancer induced by ATRA
remain poorly understood.
The sodium/iodine symporter (NIS) is a transmembrane
protein responsible for the transport of iodine into the thyroid
follicular cells, which is a crucial step for the synthesis of
hormones, including T3 and T4 (12). This property of NIS to concentrate
iodine ions within thyroid cells constitutes the basis for thyroid
scintigraphy and iodine isotope treatment of hyperthyroidism and
thyroid cancer (12). Loss of
thyroid-specific functions in poorly differentiated thyroid cancer
leads to the lack of efficacy of isotope therapy in these cancers
(13). Identifying methods to restore
the NIS function may be one way to improve the treatment efficacy
in anaplastic thyroid carcinoma.
Previous studies have suggested that the
transcription activity of β-catenin is closely associated with the
transcription of genes involved in thyroid cell growth and
differentiation, as well as NIS gene transcription and translation
in thyroid cancer (14,15). Therefore, it may be hypothesized that
the transcription activity of β-catenin can directly affect the
isotope sensitivity of thyroid cancer cells. The aim of the present
study was to investigate whether the transcription activity of
β-catenin plays a role in ATRA-mediated iodine uptake in thyroid
cancer.
Materials and methods
Cells
Undifferentiated human anaplastic thyroid carcinoma
SW1736 cells were provided by Professor Michael Derwahl (Institute
of Endocrinology, Hedwig Hospital, Humboldt University, Berlin,
Germany); this cell line has a tolerance to isotope treatment
(16). The cells were maintained in
DMEM (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) with
10% fetal bovine serum (FBS) (Gibco; Thermo Fisher Scientific,
Inc.) in an incubator at 37°C with 5% CO2.
ATRA (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
was dissolved in ethanol to 10 µmol/l as stock solution and stored
in the dark. SW1736 cells were seeded at a density of
5×104 cells/well for 48 h, followed by ATRA treatment (1
µmol/l) for 5 days. The negative control cells were treated with 1%
ethanol. All cells were kept at 37°C in the dark using aluminum
foil. SW1736 cells stably transfected with β-catenin shRNA were
used as positive controls.
β-catenin RNA interference and stable
cell lines
The recombinant plasmid pSUPER-β-catenin-shRNA was
previously constructed (17). It was
transfected into SW1736 cells using Lipofectamine 2000 (Thermo
Fisher Scientific, Inc.). SW1736 cells transfected with
pSUPER-β-catenin-shRNA were screened by 400 µg/ml puromycin for 4
weeks to select positive clones. Finally, cells were screened and
cultured in medium with 200 µg/ml of puromycin to establish the
stable transfected cell line.
Proliferation assay
SW1736 cells treated with 1 µmol/l of ATRA or 1%
ethanol for 5 days were seeded onto a 96-well culture plate. Each
well was loaded with 180 µl of cell suspension containing
1×104 cells. After 24, 48 and 72 h, the supernatant was
discarded and cells were treated with 150 µl DMEM and 50 µl MTT
solution (2.5 mg/ml; Sigma-Aldrich; Merck KGaA). The MTT solution
was discarded following 4 h of treatment, and the cells were mixed
with 150 µl dimethyl sulfoxide at low-speed vortex for 10 min. The
absorbance of each well was measured at 570 nm using DNM-9602 ELISA
spectrometer (Perlong Medical Equipment Co., Ltd., Beijing,
China).
In vitro invasion assay
Transwell chambers (Merck KGaA) coated with Matrigel
(Sigma-Aldrich; Merck KGaA) were placed into 6-well plates and
air-dried under a sterile laminar flux hood. DMEM medium (700 µl)
containing 10% FBS was added to the lower chamber. Cell suspension
(300 µl) containing 5.0×104 SW1736 cells treated with 1
µmol/l ATRA or 1% ethanol, but without FBS, for 5 days was added to
the upper chamber. After 24, 48 and 72 h, the Transwell chambers
were washed with phosphate-buffered saline (PBS). The upper
microporous membrane was cleaned with cotton swab. Invading cells
attached to the lower microporous membrane were fixed with 2%
paraformaldehyde for 30 min and stained with hematoxylin. Cell
counting was performed by two independent investigators blind to
grouping.
In vitro iodine uptake assay
The in vitro iodine uptake assay was
performed as previously described (18). Briefly, SW1736 cells treated with 1
µmol/l ATRA or 1% ethanol for 5 days were seeded onto 6-well plates
(5.0×104 cells/well). After the cells had attached to
the bottom surface, the supernatant was discarded. Cells were
washed with Hank's balanced salt solution (HBSS) and incubated with
3.7 kBq of 125I (Thermo Fisher Scientific, Inc.) for 20
min followed by three washes with ice-cold HBSS. Cells were then
incubated with 1 ml of ethanol for 20 min. The ethanol solution was
collected into test tubes and the radioactivity per minute was
measured using a WIZARD γ counter (Perkin-Elmer Life Sciences,
Waltham, MA, USA).
Western blot analysis
SW1736 cells treated with 1 µmol/l ATRA or 1%
ethanol for 5 days were lysed with 1% NP-40 lysis buffer
(Sigma-Aldrich; Merck KGaA) at 4°C for 30 min. Total proteins were
collected after centrifugation at 25,000 × g for 10 min at 4°C.
Proteins were quantified using Coomassie brilliant blue G-250.
Samples of 20–50 µg per well were separated on 8–12% SDS-PAGE and
transferred to nitrocellulose membranes. The membranes were blocked
with 5% non-fat milk for 2 h followed by incubation with primary
antibodies against β-catenin (dilution: 1:4,000; cat. no. ab16051;
Abcam, Cambridge, MA, USA), phosphorylated (p-)β-catenin Ser45
(dilution, 1:1,000; cat. no. PA5-17685; Invitrogen; thermo Fisher
Scientific, Inc.), p-β-catenin Y654 (dilution, 1:1,000; cat. no.
ab59430, Abcam), glycogen synthase kinase-3β (GSK-3β) (dilution,
1:10,000; cat. no. ab75814, Abcam), p-GSK-3β Ser9 (dilution,
1:10,000; cat. no. ab75814; Abcam), E-cadherin (dilution, 1:1,000;
cat. no. sc-7870; Santa Cruz Biotechnology, Inc., Dallas, TX, USA),
cytokeratin 18 (CK18) (dilution: 1:1,000; cat. no. sc-28264; Santa
Cruz Biotechnology, Inc.), vimentin (dilution: 1:1,000; cat. no.
sc-5565; Santa Cruz Biotechnology, Inc.), urokinase-type
plasminogen activator (uPA) (dilution: 1:1,000; cat. no. sc-14019;
Santa Cruz Biotechnology, Inc.) and its receptor uPAR (dilution:
1:1,000; cat. no. sc-10815; Santa Cruz Biotechnology, Inc.), and
fibronectin (dilution, 1:1,000; cat. no. sc-9068; Santa Cruz
Biotechnology, Inc.) at 4°C overnight. The next day, the membranes
were extensively washed with Tris-buffered saline and incubated
with goat anti-rabbit secondary antibody (dilution: 1:30,000; cat.
no. A9919; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 2 h
at 4°C. The bands were visualized by electrochemiluminescence
(Pierce; Thermo Fisher Scientific, Inc.). Densitometry images were
analyzed using ImageJ software (Rawak Software, Inc., the Tomancak
Laboratory, Dresden, Germany).
Heterotopic xenograft mouse model
SW1736 cells (2×106) were treated with 1
µmol/l ATRA or 1% ethanol for 5 days, and SW1736 cells stably
transfected with β-catenin shRNA were suspended in 200 µl of
culture medium and implanted subcutaneously into the left forelimb
of 4–6-week-old male SCID mice (22–25 g body weight). Intervention
was applied after the subcutaneous tumor size reached 50
mm3. The experimental mice (alcohol, β-catenin shRNA and
ATRA groups) were intraperitoneal injected with 37 MBq of
131I (Thermo Fisher Scientific, Inc.) once on the first
day of treatment. The control mice (ATRA and alcohol groups) were
intraperitoneally injected with 0.1 ml of normal saline (NS)
instead (ATRA+NS and Alcohol+NS). There were 10 mice in each group
(supplied by Beijing Huafukang Biotechnology Co., Ltd., Beijing,
China). The 131I intervention was commenced on the first
day of the experiment. Subcutaneous tumor size was measured every 5
days using a caliper (volume=long diameter × short
diameter2 + 2). Mice were sacrificed by CO2
inhalation after 35 days of observation. Tumors were collected and
weighed. All procedures and animal experiments were approved by the
Animal Care and Use Committee of Beijing Jishuitan Hospital
(Beijing, China).
Statistical analysis
All statistical analyses were conducted using SPSS
13.0 (SPSS Inc., Chicago, IL, USA). Data are expressed as the mean
± standard deviation (SD) from three independent experiments
performed in triplicate. Statistical significance was evaluated by
one-way analysis of variance, with the least significant difference
test for post-hoc analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
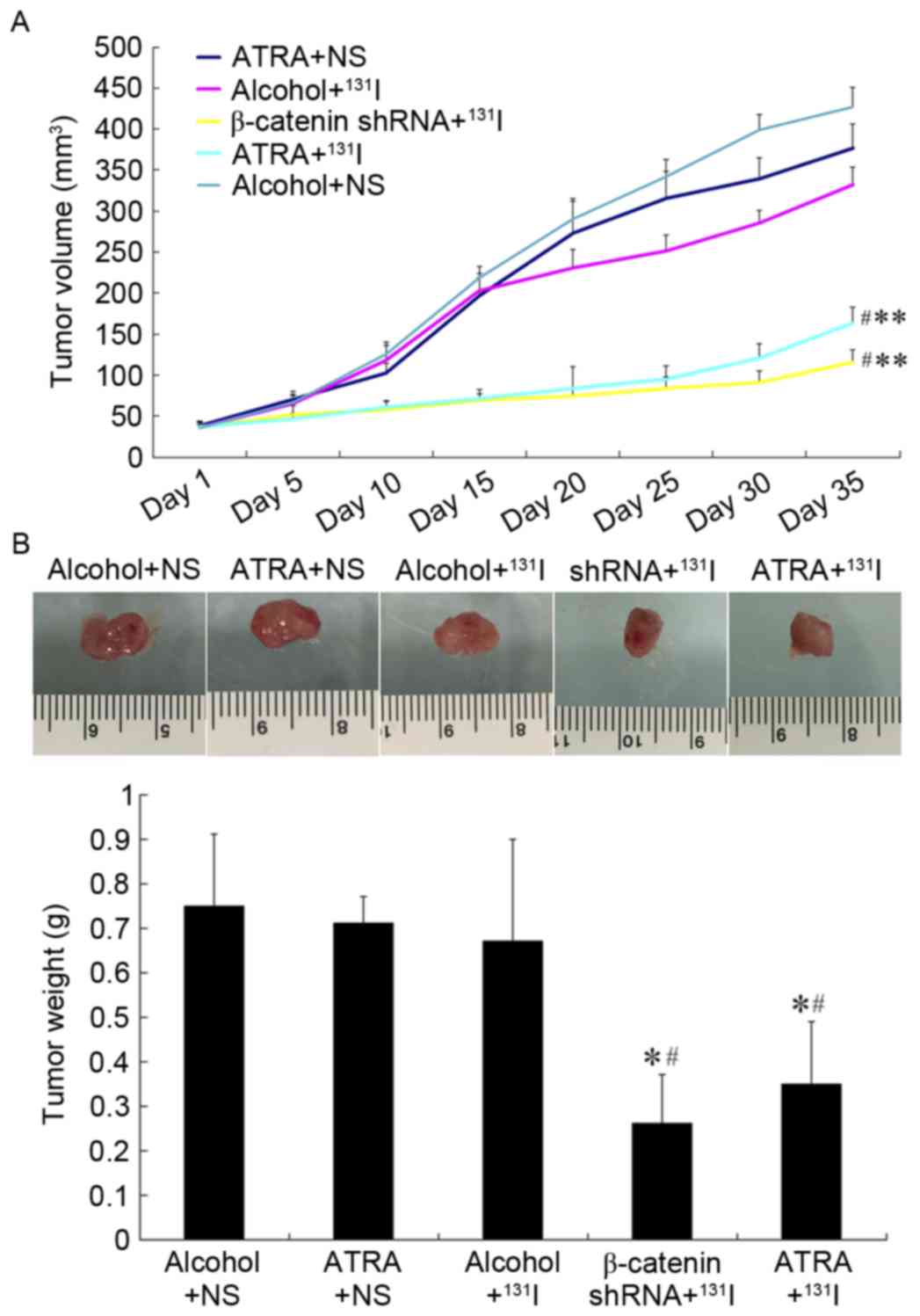
ATRA pretreatment improved
131I sensitivity and inhibited tumor growth in the
heterotopic xenograft mouse model
After 35 days of 131I treatment,
ATRA-pretreated tumor volume and weight were decreased compared
with the alcohol+131I group (163.32±19.57 vs.
332.06±21.37 mm3; 0.35±0.14 vs. 0.67±0.23 g; both
P<0.05) (Fig. 1). Similar results
were observed in the β-catenin-shRNA+131I group
(116.3±14.4 mm3 and 0.26±0.11 g, both P<0.05 vs. the
alcohol+131I group). There were no differences in tumor
volume and weight among the tumors treated with ATRA+NS,
Alcohol+NS, and Alcohol+131I groups (all P>0.05).
Furthermore, there were no differences in the tumor volume and
weight between the tumors treated with ATRA+131I and
β-catenin-shRNA+131I (both P>0.05) (Fig. 1).
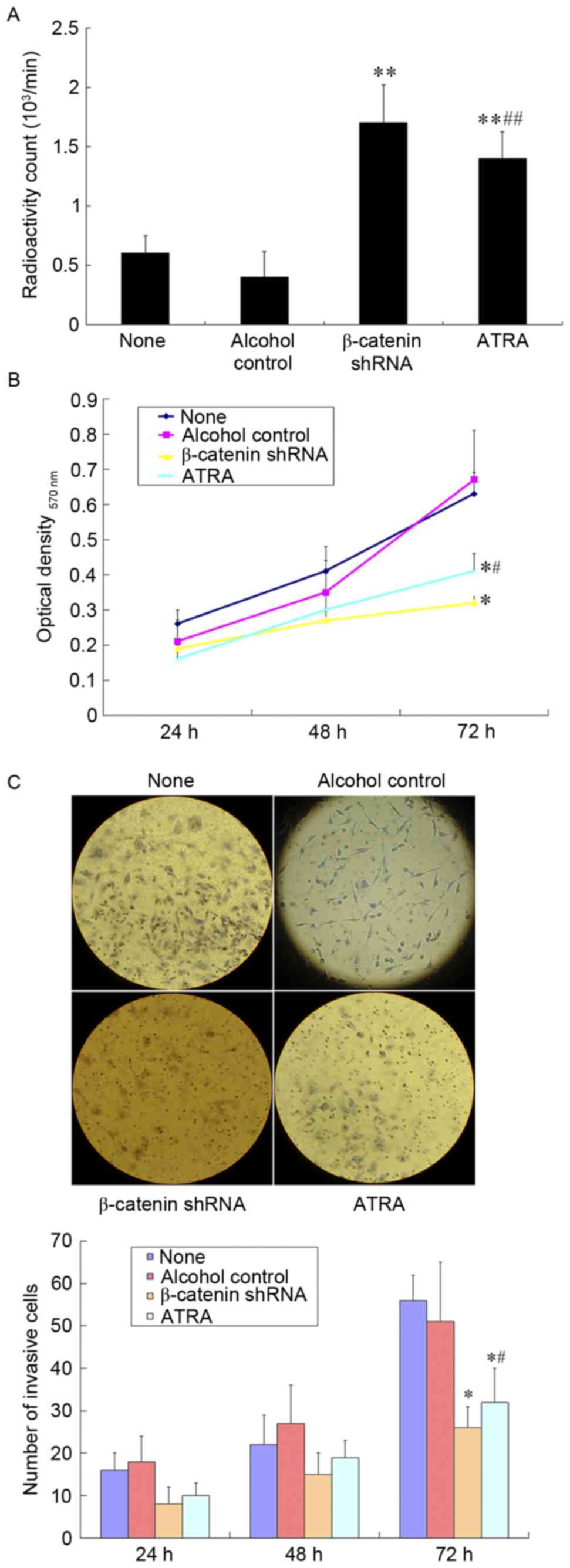
Effect of ATRA on iodine uptake in the
SW1736 cells
The iodine uptake assay revealed that ATRA increased
the uptake of iodine by SW1736 cells compared with the alcohol
control group (1.4±0.2 vs. 0.4±0.2×103/min; P<0.01)
(Fig. 2A). Similar results were
observed in the β-catenin-shRNA cells compared with the no
transfection group (1.7±0.3 vs. 0.6±0.2×103/min;
P<0.01).
ATRA inhibited the invasion and
proliferation of SW1736 cells
MTT and Transwell invasion assays indicated that
ATRA treatment decreased the cell proliferation (Fig. 2B) and invasion (Fig. 2C) compared with untreated and alcohol
control cells (P<0.05). β-catenin-shRNA cells showed results
that were similar to the ATRA-treated cells.
Effects of ATRA on phosphorylation of
β-catenin and GSK-3β in SW1736 cells
Western blot analysis revealed that ATRA treatment
decreased the expression of t-β-catenin, p-β-catenin Ser45,
p-β-catenin Y654 and p-GSK-3β, and increased the expression of
t-GSK-3β compared with untreated and alcohol control cells
(Fig. 3A). Similar results were
observed in β-catenin-shRNA cells.
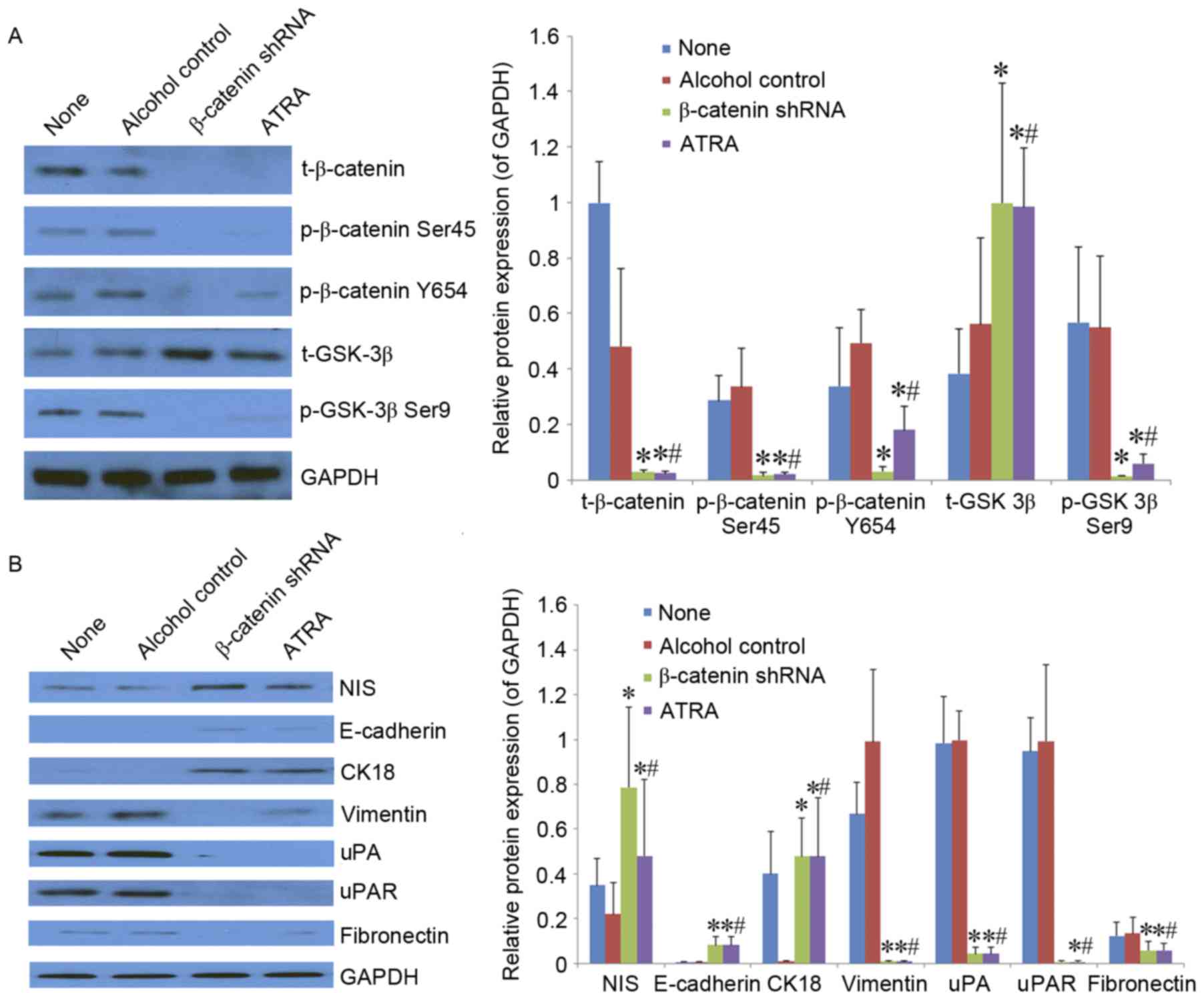 | Figure 3.Effects of ATRA on the protein
expression of GSK-3β, β-catenin, NIS and proteins involved in
epithelial-mesenchymal transition in thyroid cancer SW1736 cells.
Protein expression was determined by western blot analysis. GAPDH
was used as an internal control. (A) Total β-catenin, total GSK-3β,
phosphorylated β-catenin and phosphorylated GSK-3β. (B) NIS,
E-cadherin, CK18, vimentin, uPA and its receptor uPAR, and
fibronectin. Data are shown as mean ± standard deviation from three
independent experiments performed in triplicate. *P<0.05 vs.
None; #P<0.05 vs. alcohol control. None, SW1736 cells
without any treatment; ATRA, all-trans-retinoic acid; shRNA, short
hairpin RNA; t-, total; p-, phosphorylated; GSK-3β, glycogen
synthase kinase-3β; CK18, cytokeratin 18; NIS, sodium/iodine
symporter; uPA, urokinase-type plasminogen activator. |
Western blot analysis also revealed that ATRA
treatment increased the expression of NIS, E-cadherin and CK18, and
decreased the expression of vimentin, uPA, uPAR and fibronectin
compared with untreated and alcohol control cells (Fig. 3B). Similar results were observed in
β-catenin-shRNA cells.
Discussion
Loss of thyroid-specific functions in
undifferentiated thyroid cancers leads to a lack of efficacy of
isotope therapy (1). ATRA may restore
the NIS function to enhance iodine uptake capability in thyroid
carcinoma (4,5,10,11), but the exact mechanisms remain poorly
understood. In the present study, ATRA treatment has been shown to
increase iodine uptake by SW1736 cells, both in vitro and
in vivo. This is consistent with the study by Muhlbauer
et al (10), which revealed
that ATRA could induce iodine accumulation in rats via the NIS.
Similar results were also observed in medullary thyroid cancer stem
cells and in mice (11).
The results of the present study indicate that the
β-catenin pathway is involved in the process of ATRA-improved
iodine uptake by SW1736 cells. ATRA inhibited β-catenin
phosphorylation at Ser45 and GSK-3β phosphorylation at Ser9, which
should consequently enhance GSK-3β hydrolytic activity and decrease
β-catenin levels in the cytoplasm (19). In addition, phosphorylation of
β-catenin at Y654 was decreased, which should inhibit the
translocation of β-catenin to the nucleus (19), resulting in the restoration of NIS
expression and the recovery of cellular iodine uptake capability
and susceptibility to radioactive iodine therapy (19).
The restoration of NIS function should improve the
sensitivity of undifferentiated thyroid cancers to isotope
treatment. A recent study revealed that the Wnt/β-catenin pathway
is involved in thyroid function (14), and a previous study showed that
β-catenin nuclear translocation suppresses NIS membrane
localization in thyroid cancer cells (15). Other studies have shown that a number
of pathways could be involved in NIS expression. Ryan et al
(20) used reverse
transcription-quantitative polymerase chain reaction to measure and
compare the expression levels of NIS, estrogen receptor α (ERα) and
retinoic acid receptor in healthy human breast tissue and breast
cancer tissue; NIS mRNA levels were found to be closely associated
with ERα and retinoic acid receptor expression. Cheong et al
(21) further demonstrated that the
ERα agonist 17β-estradiol could decrease ATRA-triggered iodine
uptake by 50%. Based on these findings, it was suggested that
iodine uptake induced by ATRA was associated with ERα-mediated
transcription (21). Similarly, Lee
et al (22) found that in
thyroid cancer, ATRA reduced the phosphorylation levels of p38 MAPK
and attenuated endoplasmic reticulum stress, subsequently affecting
NIS expression and iodine uptake. Furthermore,
staurosporine-associated protease inhibitors showed evident
interference with ATRA-mediated iodine uptake (23). ATRA was reported to bind to its
specific receptor and activate the PI3K/AKT signaling pathway,
which may be one of the mechanisms involved in improvements of
iodine uptake (24). Overall,
β-catenin nuclear translocation may be one of the molecular
mechanisms by which ATRA promotes iodine uptake, but additional
studies remain necessary to confirm these results.
In the present study, ATRA decreased the
proliferation of thyroid cancer cells, which has been previously
observed in other studies of thyroid cancer cells (9,25,26), as well as in other types of cancer
cells, including pancreatic cancer (6), leukemia (7) and breast cancer (8). In addition, the present study showed
that ATRA modulates a number of proteins involved in EMT in a way
that indicates ATRA should inhibit EMT. Previous studies have shown
that ATRA increases the expression of NIS, E-cadherin and CK18, and
decreases the expression of vimentin, uPA, uPAR and fibronectin
(27–29), as was observed in the present study.
These effects should help to decrease EMT in thyroid cancers and
maintain NIS potency.
Furthermore, ATRA has been shown to inhibit the
proliferation and invasiveness of SW1736 cells by downregulating
the transcriptional activity of β-catenin (14,19).
β-catenin also participates in the proliferation and invasiveness
of cancer cells by modulating the Wnt/β-catenin pathway. Inhibiting
the Wnt/β-catenin pathway has been shown to suppress breast cancer
metastatic spread (30) and to
inhibit gastric cancer stem cells (31). Therefore, the use of ATRA in thyroid
cancer could affect three aspects, consisting of NIS upregulation,
EMT inhibition and proliferation and invasiveness inhibition.
Nevertheless, additional studies remain necessary to obtain a more
comprehensive understanding of the mechanisms involved.
In conclusion, ATRA increased iodine uptake and
inhibited proliferation and invasion of human anaplastic thyroid
carcinoma SW1736 cells. Inhibition of β-catenin phosphorylation may
be involved in this process. ATRA could be used to improve the
isotope sensitivity of anaplastic thyroid carcinoma.
Acknowledgements
This study was funded by National natural science
foundation (grant nos. 81372858, 30700968 and 30800416).
References
|
1
|
Kojic KL, Kojic SL and Wiseman SM:
Differentiated thyroid cancers: A comprehensive review of novel
targeted therapies. Expert Rev Anticancer Ther. 12:345–357. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Smallridge RC, Ain KB, Asa SL, Bible KC,
Brierley JD, Burman KD, Kebebew E, Lee NY, Nikiforov YE, Rosenthal
MS, et al: American Thyroid Association guidelines for management
of patients with anaplastic thyroid cancer. Thyroid. 22:1104–1139.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Glaser SM, Mandish SF, Gill BS,
Balasubramani GK, Clump DA and Beriwal S: Anaplastic thyroid
cancer: Prognostic factors, patterns of care, and overall survival.
Head Neck. 38 Suppl 1:E2083–E2090. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schenk T, Stengel S and Zelent A:
Unlocking the potential of retinoic acid in anticancer therapy. Br
J Cancer. 111:2039–2045. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gutierrez-Mazariegos J, Schubert M and
Laudet V: Evolution of retinoic acid receptors and retinoic acid
signaling. Subcell Biochem. 70:55–73. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Leelawat K, Ohuchida K, Mizumoto K,
Mahidol C and Tanaka M: All-trans retinoic acid inhibits the cell
proliferation but enhances the cell invasion through up-regulation
of c-met in pancreatic cancer cells. Cancer Lett. 224:303–310.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu SM, Chen W and Wang J: Distinguishing
between cancer cell differentiation and resistance induced by
all-trans retinoic acid using transcriptional profiles and
functional pathway analysis. Sci Rep. 4:55772014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tari AM, Lim SJ, Hung MC, Esteva FJ and
Lopez-Berestein G: Her2/neu induces all-trans retinoic acid (ATRA)
resistance in breast cancer cells. Oncogene. 21:5224–5232. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang M, Guo R, Xu H, Zhang M and Li B:
Retinoic acid and tributyrin induce in-vitro radioiodine uptake and
inhibition of cell proliferation in a poorly differentiated
follicular thyroid carcinoma. Nucl Med Commun. 32:605–610. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Muhlbauer M, da Silva AC, Marassi MP,
Lourenço AL, Ferreira AC and de Carvalho DP: Retinoic acid
modulation of thyroid dual oxidase activity in rats and its impact
on thyroid iodine organification. J Endocrinol. 205:271–277. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tang M, Hou YL, Kang QQ, Chen XY, Duan LQ,
Shu J, Li SL, Hu XL and Peng ZP: All-trans-retinoic acid promotes
iodine uptake via up-regulating the sodium iodide symporter in
medullary thyroid cancer stem cells. Asian Pac J Cancer Prev.
15:1859–1862. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dohan O, De la Vieja A, Paroder V, Riedel
C, Artani M, Reed M, Ginter CS and Carrasco N: The sodium/iodide
symporter (NIS): Characterization, regulation, and medical
significance. Endocr Rev. 24:48–77. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Furuya F, Shimura H, Suzuki H, Taki K,
Ohta K, Haraguchi K, Onaya T, Endo T and Kobayashi T: Histone
deacetylase inhibitors restore radioiodide uptake and retention in
poorly differentiated and anaplastic thyroid cancer cells by
expression of the sodium/iodide symporter thyroperoxidase and
thyroglobulin. Endocrinology. 145:2865–2875. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sastre-Perona A and Santisteban P:
Wnt-independent role of β-catenin in thyroid cell proliferation and
differentiation. Mol Endocrinol. 28:681–695. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lan L, Deng W, Chen HL, Huo L, Deng L,
Zhang G and Luo Y: Nuclear translocation of β-catenin represses
membrane localization of NIS in human thyroid cancer cells.
Zhonghua Yi Xue Za Zhi. 96:891–896. 2016.PubMed/NCBI
|
|
16
|
Haberkorn U, Altmann A, Jiang S, Morr I,
Mahmut M and Eisenhut M: Iodide uptake in human anaplastic thyroid
carcinoma cells after transfer of the human thyroid peroxidase
gene. Eur J Nucl Med. 28:633–638. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jiang Y, Zhao J, Wu C, Luo Y and He D:
Construction and identification of shRNA eukaryotic vector
targeting human β-catenin. Acad J Second Milit Med Uni. 30:965–967.
2009. View Article : Google Scholar
|
|
18
|
Nakamoto Y, Saga T, Misaki T, Kobayashi H,
Sato N, Ishimori T, Kosugi S, Sakahara H and Konishi J:
Establishment and characterization of a breast cancer cell line
expressing Na+/I-symporters for radioiodide concentrator gene
therapy. J Nucl Med. 41:1898–1904. 2000.PubMed/NCBI
|
|
19
|
Sastre-Perona A and Santisteban P: Role of
the wnt pathway in thyroid cancer. Front Endocrinol (Lausanne).
3:312012.PubMed/NCBI
|
|
20
|
Ryan J, Curran CE, Hennessy E, Newell J,
Morris JC, Kerin MJ and Dwyer RM: The sodium iodide symporter (NIS)
and potential regulators in normal, benign and malignant human
breast tissue. PLoS One. 6:e160232011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheong SJ, Jang D, Jeong HJ, Lim ST, Sohn
MH, Katzenellenbogen JA and Kim DW: Reduction of stimulated sodium
iodide symporter expression by estrogen receptor ligands in breast
cancer cells. Nucl Med Biol. 38:287–294. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee SJ, Kim SH, Kang JG, Kim CS, Ihm SH,
Choi MG and Yoo HJ: Effects of all-trans retinoic acid on
sodium/iodide symporter and CCAAT/enhancer-binding
protein-homologous protein under condition of endoplasmic reticulum
stress in FRTL5 thyroid cells. Horm Metab Res. 43:331–336. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Beyer S, Lakshmanan A, Liu YY, Zhang X,
Wapnir I, Smolenski A and Jhiang S: KT5823 differentially modulates
sodium iodide symporter expression, activity, and glycosylation
between thyroid and breast cancer cells. Endocrinology.
152:782–792. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Willhauck MJ, Kane ODJ, Wunderlich N, Göke
B and Spitzweg C: Stimulation of retinoic acid-induced functional
sodium iodide symporter (NIS) expression and cytotoxicity of
131I by carbamazepine in breast cancer cells. Breast
Cancer Res Treat. 125:377–386. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kang HJ, Youn YK, Hong MK and Kim LS:
Antiproliferation and redifferentiation in thyroid cancer cell
lines by polyphenol phytochemicals. J Korean Med Sci. 26:893–899.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Malehmir M, Haghpanah V, Larijani B,
Ahmadian S, Alimoghaddam K, Heshmat R, Ghavamzadeh A, Adabi K and
Ghaffari SH: Multifaceted suppression of aggressive behavior of
thyroid carcinoma by all-trans retinoic acid induced
re-differentiation. Mol Cell Endocrinol. 348:260–269.
2012.PubMed/NCBI
|
|
27
|
Lan L, Cui D, Luo Y, Shi BY, Deng LL,
Zhang GY and Wang H: Inhibitory effects of retinoic acid on
invasiveness of human thyroid carcinoma cell lines in vitro. J
Endocrinol Invest. 32:731–738. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dutta A, Sen T and Chatterjee A: All-trans
retinoic acid (ATRA) downregulates MMP-9 by modulating its
regulatory molecules. Cell Adh Migr. 4:409–418. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vasko V, Espinosa AV, Scouten W, He H,
Auer H, Liyanarachchi S, Larin A, Savchenko V, Francis GL, de la
Chapelle A, et al: Gene expression and functional evidence of
epithelial-to-mesenchymal transition in papillary thyroid carcinoma
invasion. Proc Natl Acad Sci USA. 104:pp. 2803–2808. 2007,
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jang GB, Kim JY, Cho SD, Park KS, Jung JY,
Lee HY, Hong IS and Nam JS: Blockade of Wnt/β-catenin signaling
suppresses breast cancer metastasis by inhibiting CSC-like
phenotype. Sci Rep. 5:124652015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mao J, Fan S, Ma W, Fan P, Wang B, Zhang
J, Wang H, Tang B, Zhang Q, Yu X, et al: Roles of Wnt/β-catenin
signaling in the gastric cancer stem cells proliferation and
salinomycin treatment. Cell Death Dis. 5:e10392014. View Article : Google Scholar : PubMed/NCBI
|