Introduction
Colorectal cancer (CRC) is the third most common
type of cancer affecting the gastrointestinal tract worldwide
(1). CRC has a high mortality rate
and is often associated with rapid cancer progression and advanced
tumor presentation at the time of diagnosis (2,3). Recent
changes in lifestyle, diet and an aging population have all
contributed to the increased prevalence of CRC in China (4). Patients who have been diagnosed with
early stage CRC frequently are treated with surgery in conjunction
with chemotherapy and radiotherapy to improve their survival rate
(1). However, for the majority of
patients diagnosed with CRC have already reached an advanced
pathological stage, chemotherapy is the major form of treatment
(5–8).
In particular, 5-fluorouracil (5-FU)-based regimens are used as the
international standard for late-stage CRC treatment (9,10).
Nevertheless, the increasing occurrence of multi-drug resistance
(MDR) has had negative impact on the clinical treatment of CRC
(11,12). Other currently used anti-cancer agents
often present unacceptable levels of toxicity to normal cells and
tissues (13,14), thereby limiting their
effectiveness.
The development of colorectal cancer is a complex
and multi-stage process, characterized by various interactions
between environmental carcinogens, genetic modifications and the
host immune system (15). Dense
infiltration of cytokine-producing immune/inflammatory cells
frequently result in the aberrant activation of multiple
intracellular signal transduction cascades, ultimately resulting in
the uncontrolled growth of transformed cells. In addition,
crosstalk between these signaling pathways results in a complicated
and robust cellular signal transduction network that is regulated
by compensatory mechanisms. Therefore, anti-cancer drugs that
target only a single signaling pathway might not be as effective
and is more prone to the development of MDR or adverse side effects
(16).
Hedyotis diffusa Willd (HDW) is a medicinal
herb, which belongs to the Rubiaceae family, and is widely
distributed in Northeast Asia (17).
It is a well-known traditional Chinese herbal medicine used for
‘heat-clearing’, detoxification and promoting blood circulation
(18). HDW has also been widely used
as an important component in traditional Chinese medicine (TCM)
prescription formulae for the treatment of various types of cancer,
including CRC (17–19). Our previous studies have demonstrated
that HDW is able to inhibit colon cancer growth in vivo and
in vitro by promoting apoptosis of cancer cells, inhibiting
proliferation, tumor angiogenesis and MDR reversal (20–25).
However, the precise mechanism of the in vivo anti-cancer
activity of HDW remains largely unknown. In the present study, the
present authors evaluated the efficacy of HDW in inhibiting CRC
growth in a mouse xenograft model as well as various CRC cell lines
in order to further elucidate the anti-cancer mechanism of HDW.
Materials and methods
Materials and reagents
RPMI-1640, Dulbecco's modified Eagle's medium (DMEM)
with high glucose, Fetal bovine serum (FBS),
penicillin-streptomycin (catalog no. SV30010), 0.25% trypsin-EDTA,
Pierce RIPA buffer, Pierce BCA Protein Assay kit (catalog no.
23227), and SuperSignal™ West Pico Chemiluminescent
substrate (catalog no. 34080) were all purchased from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). CellTiter 96®
AQueous Non-Radioactive Cell Proliferation Assay kit (MTS assay;
catalog no. G5430) was provided by Promega Corporation (Madison,
WI, USA). Matrigel was obtained from BD Biosciences (Franklin
Lakes, NJ, USA). TumorTACS in situ Apoptosis kit (catalog
no. 4815–30-K) was purchased from R&D Systems Inc.
(Minneapolis, MN, USA). VECTASTAIN Elite ABC kit was provided by
Vector Laboratories, Inc. (Burlingame, CA, USA). Rabbit monoclonal
antibodies against Pim-1 (catalog no. ab75776), B-cell lymphoma 2
(Bcl-2; catalog no. ab32124), cyclooxygenase-2 (COX-2; catalog no.
ab62331), inducible nitric oxide synthase (iNOS; catalog no.
ab178945), endothelial nitric oxide synthase (eNOS; catalog no.
ab66127), rabbit polyclonal antibody against Ki-67 (catalog no.
ab15580), Bcl-2-like protein 4 (Bax; catalog no. ab69643) and mouse
monoclonal antibody against hypoxia-inducible factor 1-α (HIF1-α;
catalog no. ab463) were purchased from Abcam (Cambridge, MA, USA).
Rabbit monoclonal antibodies against cytochrome C (catalog no.
4280), caspase-3 (catalog no. 9665), poly (ADP-ribose) polymerase 1
(PARP; catalog no. 9532), mouse monoclonal antibody against
caspase-9 (catalog no. 9508), and rabbit polyclonal antibody
against β-actin (catalog no. 4967) were obtained from Cell
Signaling Technology, Inc., (Danvers, MA, USA). Horseradish
peroxidase (HRP)-conjugated goat anti-rabbit immunoglobulin G (IgG;
catalog no. E030120) and goat anti-mouse IgG secondary antibodies
(catalog no. E030110) was purchased from Earthox, LLC (Millbrae,
CA, USA). Mouse interleukin (IL)-1β, IL-6, IL-4, IL-10 and tumor
necrosis factor (TNF)-α ELISA kits were obtained from Shanghai
Westang Bio-tech Co., Ltd. (Shanghai, China). Bio-Plex
Phosphoprotein Detection Reagent kit (catalog no. 171–304005) were
purchased from Bio-Rad Laboratories (Hercules, CA, USA). Culture
flask and plates were purchased from Wuxi NEST Biotechnology Co.,
Ltd. (Wuxi, Jiangsu, China). All the other chemicals used, unless
otherwise stated, were obtained from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany).
Preparation of ethanol extract from
Hedyotis diffusa Willd (EEHDW)
Authentic plant material was purchased from Guo Yi
Tang Chinese herbal medicine store (Fujian, China). Identification
of the Hedyotis diffusa Willd (HDW) herb was confirmed by Dr Wei Xu
(Department of Pharmacology, Fujian University of Traditional
Chinese Medicine, Fujian, China). Ethanol extract of HDW (EEHDW)
was prepared as previously described (20–22).
Briefly, 500 g HDW was extracted with 5,000 ml of 85% ethanol using
refluxing method and then filtered. The ethanol solvent was then
evaporated on a rotary evaporator. The resultant solution was
concentrated to a relative density of 1.05, and the dried powder of
EEHDW was obtained using spray desiccation method using a spray
dryer.
For in vitro experiments, stock solutions of
EEHDW were prepared by dissolving the EEHDW powder in 40% dimethyl
sulfoxide (DMSO) to achieve a final concentration of 500 mg/ml.
Working concentrations of EEHDW were prepared by diluting the stock
solution in culture medium (RPMI-1640 for HCT-8 and HCT-116; DMEM
for HT-29 and SW620). The final concentration of DMSO in the medium
was <0.2%. For in vivo experiments, working
concentrations of EEHDW were prepared by dissolving the EEHDW
powder in saline to a concentration of 0.1 g/ml.
Cell culture
Human CRC cell lines, HCT-8, HT-29, HCT-116 and
SW620, were purchased from Cell Bank of the Chinese Academy of
Sciences (Shanghai, China). HCT-8 and HCT-116 cells were cultured
in RPMI-1640. HT-29 and SW620 cells were cultured in DMEM. All cell
media were supplemented with 10% (v/v) FBS, 100 U/ml penicillin and
100 µg/ml streptomycin and cultured at 37°C, with 5% CO2
in a humidified incubator.
Evaluation of cell viability
Cell viability was assessed using MTS assay. The
cells (10,000/well) were incubated in 96-well plates with culture
medium at 37°C for 12 h and then treated with various
concentrations of EEHDW (0, 0.5, 1 and 2 mg/ml) for 24 and 48 h;
the control cells were treated with DMSO without EEHDW at 37°C for
24 and 48 h. Next, 10 µl MTS was added to each well, and the
samples were incubated for 1 h at 37°C. The resulting absorbance
was measured at 490 nm using an ELISA reader (BioTek Instruments,
Inc., Winooski, VT, USA).
Animals
A total of 20-week-old athymic BALB/c nu/nu male
mice (initial body weight, 20±2 g) were obtained from Shanghai SLAC
Laboratory Animal Co., Ltd. (Shanghai, China) and housed under
pathogen-free conditions (22°C; 12-h light/dark cycle). Food and
water were given ad libitum throughout the experiment. All
animal experiments were approved by the Institutional Animal Care
and Use Committee of Fujian University of Traditional Chinese
Medicine (Fujian, China).
In vivo nude mouse xenograft
study
HT-29 cells (5×106) mixed with Matrigel
(1:1) were subcutaneously injected in the right flank area of
athymic nude mice to initiate tumor growth. After 5 days of
xenograft implantation, the mice were randomly divided into two
groups (n=10) and given daily intra-gastric administration with 1
g/kg EEHDW or saline for 6 days per week for a total of 16 days.
The diameters of the tumors were measured every second day with
digital calipers and the tumor volume (V) was calculated using the
formula: V=(width)2 × length × π/6. At the end of
experiment, the mice were sacrificed using 100 mg/kg
pelltobarbitalum natricum (Sigma-Aldrich; Merck KGaA), and the
tumor tissues were removed and fixed in 4% paraformaldehyde (China
National Medicines Corporation Ltd., Beijing, China) or stored at
−80°C. Blood was collected aseptically from the orbital sinus.
Blood collection tubes were allowed to stand at room temperature
for 5 h prior to serum collection by centrifugation at 2,000 × g
for 20 min at room temperature, and then stored at −80°C.
Immunohistochemical (IHC)
staining
Tumor tissues were analyzed by IHC as previously
described (24). Briefly, following
fixation at room temperature with 4% paraformaldehyde for 24 h, the
tumor samples were paraffin-embedded using 100% liquid paraffin at
56°C until the paraffin had fully hardened, and sliced into 4
µm-thick sections. The slides were subjected to antigen retrieval
and endogenous peroxidase activity was quenched using hydrogen
peroxide. IHC staining was performed using the VECTASTAIN Elite ABC
kit according to the manufacturer's instructions. Briefly,
following blocking of non-specific proteins with normal serum in
PBS (0.1% Tween 20), the slides were incubated with Ki-67, Pim-1,
Bcl-2, Bax, COX-2, iNOS, eNOS, HIF1-α (all 1:200 diluted in PBS)
primary antibodies or PBS (negative control) overnight at 4°C.
After washing with PBS, the slides were incubated for 30 min at
room temperature with biotinylated secondary antibody (goat
anti-rabbit immunoglobulin G; cat. no. E030120) followed by
conjugated HRP-labeled streptavidin (Dako; Agilent Technologies,
Inc., Santa Clara, CA, USA) and then washed with PBS. The slides
were then incubated with 3,3′-diaminobenzidine (DAB, Sigma-Aldrich,
Merck KGaA) as the chromogen, followed by counterstaining with
diluted hematoxylin (Sigma-Aldrich, Merck KGaA) at room temperature
for 30 sec. After staining, five high-power fields (magnification,
×400) were randomly selected in each slide and the mean proportion
of positive cells in each field were counted using the true color
multi-functional cell image analysis management system (version
6.0; Image-Pro Plus; Media Cybernetics, Inc., Rockville, MD,
USA).
In situ apoptosis detection by
terminal deoxynucleotidyl-transferase-mediated dUTP nick end
labeling (TUNEL) staining
Apoptosis in tumor tissues were analyzed by TUNEL
staining using TumorTACS in situ Apoptosis kit as described
previously (22). Briefly, apoptotic
cells were examined by light microscopy (DM4000, Leica Microsystems
GmbH, Wetzlar, Germany) and counted as DAB-positive cells (brown
staining) at five arbitrarily selected microscopic fields
(magnification, ×400). TUNEL-positive cells were counted as a
percentage of the total cells.
Preparation of tumor homogenate and
western blotting
A total of three tumors were randomly selected from
the control and the EEHDW group, washed with PBS 3 times and
homogenized in Pierce RIPA buffer containing protease inhibitor and
phosphatase inhibitor cocktails. The samples were then centrifuged
at 17,000 × g for 20 min at 4°C, and the resulting protein
concentrations were determined using BCA Protein Assay reagent kit.
A total of 50 µg protein for each sample was loaded onto 10%
SDS-PAGE and resolved at 20 V for 10 min, at 80 V for 30 min and at
120 V for 1 h. The proteins were then transferred onto
nitrocellulose membranes. Following blocking with 5% non-fat milk
powder at room temperature for 2 h with the 5% non-fat milk powder
dissolved using TBS with Tween-20 (TBST, pH8.0) containing 0.1%
Tween, the membranes were incubated with cytochrome C, caspase-3,
caspase-9, PARP and β-actin (all 1:1,000) primary antibodies
overnight at 4°C, and then incubated with the aforementioned
HRP-conjugated anti-rabbit secondary antibodies to bind the
antibodies of cytochrome C, caspase-3, PARP and β-actin, or
anti-mouse secondary antibody to bind the caspase-9 antibody (all
1:5,000) for 1 h at room temperature. The membranes were then
subjected to enhanced chemiluminescence (ECL) detection using
SuperSignal™ West Pico Chemiluminescent substrate. Image
Lab™ software (version 3.0; Bio-Rad Laboratories, Inc.)
was used for densitometric analysis.
Measurement of cytokines in serum by
ELISA
Blood were collected and stored at −80°C until
further analysis. The level of IL-1β, IL-6, IL-4, IL-10 and TNF-α
in the serum was measured using IL-1β (cat. no. F10770), IL-6 (cat.
no. F10830), IL-4 (cat. no. F10810), IL-10 (cat. no. F10870) and
TNF-α (cat. no. F11630) ELISA kits (Xitang Biological Technology
Co., Ltd., Shanghai, China), respectively, according to the
manufacturer's protocol. Briefly, 100 µl diluted standard and test
samples were added in each well. Plates were covered and incubated
for 2 h at 37°C, then washed 5 times prior to incubation with 50 µl
biotinylated antibody solution for 1 h at 37°C. The Plates were
washed three times, and 100 µl streptavidin-HRP solution was
distributed to all wells prior to incubation for 1 h at 37°C. After
washing, the substrate was added and incubated for 15 min at room
temperature in a darkened room. Finally, the reactions were stopped
and the absorbance was measured at 450 nm. Wells with no
biotinylated antibody solutions or streptavidin-HRP solution were
used as the negative controls. The concentrations of the
aforementioned cytokines were determined by comparing to serial
dilutions of the purified standards.
Bio-Plex phosphoprotein assay
A total of 8 tumors were randomly selected from the
EEHDW and control groups, and homogenized. For analysis of
phosphorylation of protein kinase B (AKT), mitogen-activated
protein kinase 1/2 (Erk1/2), c-Jun N-terminal kinase (JNK), p38,
p53 and p70S6K in vitro, HT-29 cells (2.5×105)
were seeded into 25 cm2 flasks with 5 ml RPMI-1640
medium and treated with 2 mg/ml EEHDW for 24 h; the control cells
were treated with RPMI-1604 with 0.16% DMSO without EEHDW at 37°C
for 24 h. To detect signal transducer and activator of
transcription 3 (STAT3) phosphorylation in vitro, HT-29
cells were first cultured at 37°C in complete DMEM (10% FBS) until
~70% confluence, and subsequently cultured at 37°C in FBS-free
medium overnight. The medium was replaced with DMEM with 10% FBS,
and the cells were pre-treated with EEHDW (2 mg/ml) for 1 h at
37°C, followed by stimulation with 10 ng/ml IL-6 for 15 min at
37°C. Tumor tissues and treated cells were lysed using a
commercially available lysis kit (Bio-Rad Laboratories) and
centrifuged at 17,000 × g for 20 min at 4°C, and the resulting
protein concentrations were determined by BCA protein assay. The
presence of phosphorylated (p-)AKT, p-Erk1/2, p-JNK, p-p38, p-p53,
p-p70S6K and p-STAT3 were detected using a bead-based multiplex
assay for phosphoproteins (Bio-Plex Phosphoprotein assay kit;
Bio-Rad Laboratories), according to the manufacturer's
instructions. Data were collected and analyzed using the Bio-Plex
200 suspension array system (Bio-Rad Laboratories).
Statistical analysis
All data were expressed as the mean ± standard
deviation. Statistical analysis was performed using the SPSS
software (version 17.0) for Windows (SPSS, Inc. Chicago, IL, USA)
using one-way analysis of variance. Fisher's least significant
difference (for equal variances) or Dunnett's correction (for
unequal variances) were used in post-hoc tests. P<0.05 was
considered to be statistically significant.
Results
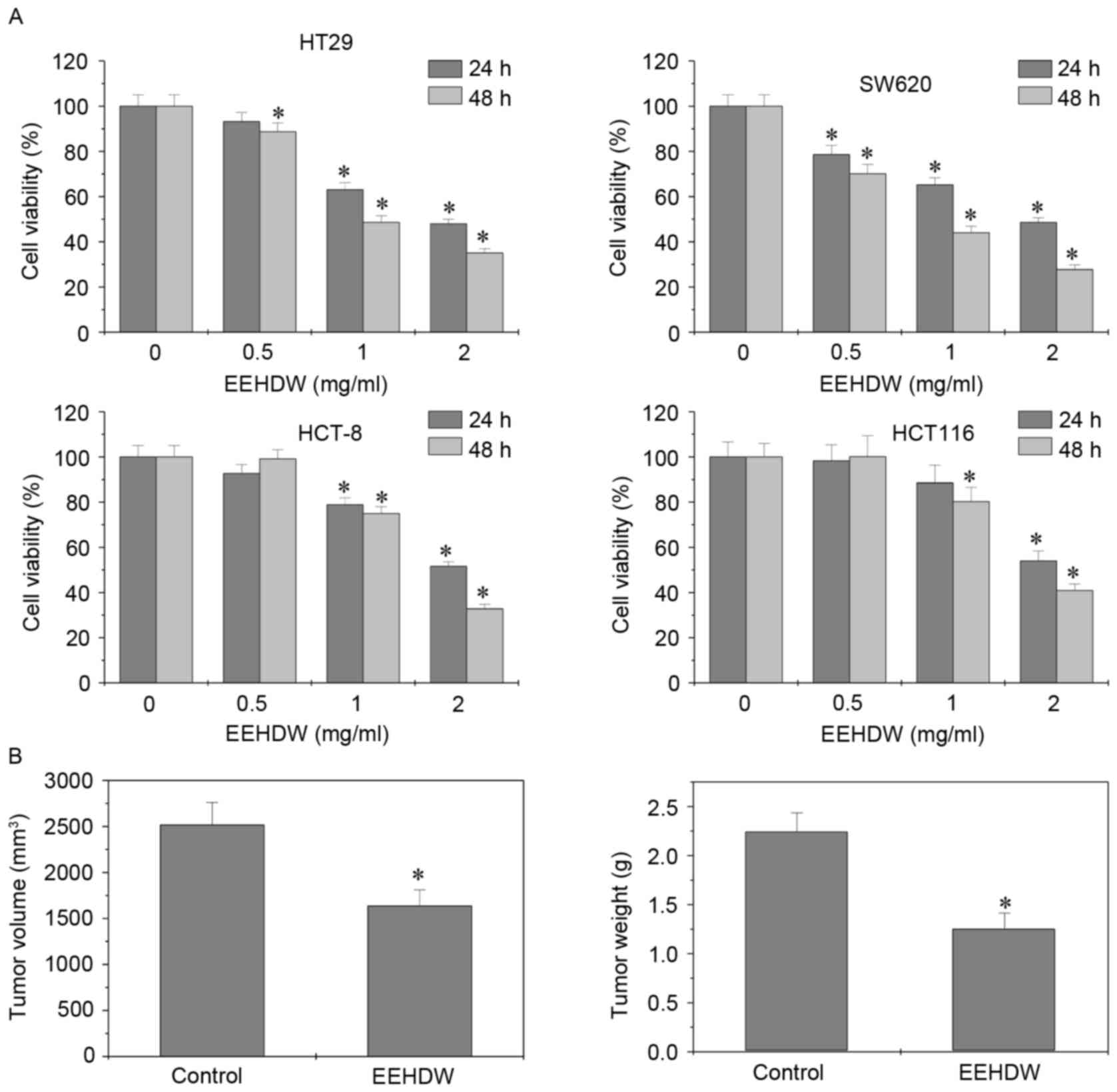
EEHDW inhibits the growth of CRC cells
and xenograft tumors in mice
The effect of EEHDW on the viability of various CRC
cell lines was determined using MTS assay. EEHDW treatment at 0.5,
1 and 2 mg/ml for 24 or 48 h was able to significantly reduce the
viability of HT-29, SW620, HCT-8 and HCT-116 cells) in a dose- and
time-dependent manner compared with the control (Fig. 1A). The in vivo anti-tumor
effect of EEHDW was subsequently determined by comparing the tumor
weight and volume between EEHDW-treated and control mice.
EEHDW-treated mice exhibited a 34.94% decrease in tumor volume
(control, 2,516±245 mm3; EEHDW-treated, 1,637±173
mm3; P<0.0037) and a 44.14% decrease in tumor weight
(control, 2.25±0.19 g; EEHDW-treated, m1.25±0.16 g; P<0.0012)
compared with the control mice (Fig.
1B). These results suggest that EEHDW was able to suppress
colorectal tumor growth in vivo and in vitro.
EEHDW inhibits cellular proliferation
and induces apoptosis in xenograft tumors
The in vivo effect of EEHDW on proliferation
and apoptosis in mice was determined using Ki-67 and TUNEL
staining, as well as western blotting for proteins in the
mitochondrial cytochrome c release-mediated caspase cascade. There
was a significant decrease in the number of Ki-67-positive cells in
the EEHDW-treated group compared with the control group
(22.20±2.63% vs. 33.50±4.22%; P<0.05), while the percentage of
TUNEL-positive cells were increased in the EEHDW-treated group
compared with the control group (35.60±5.34% vs. 21.67±3.47%;
Fig. 2A, P<0.05). Additionally,
the expression of cytochrome c, caspase-3, caspase-9 and PARP were
upregulated in the EEHDW-treated group compared with the control
group (P<0.05; Fig. 2B). Taken
together, these results demonstrated that EEHDW was able to inhibit
proliferation and promote apoptosis in vivo.
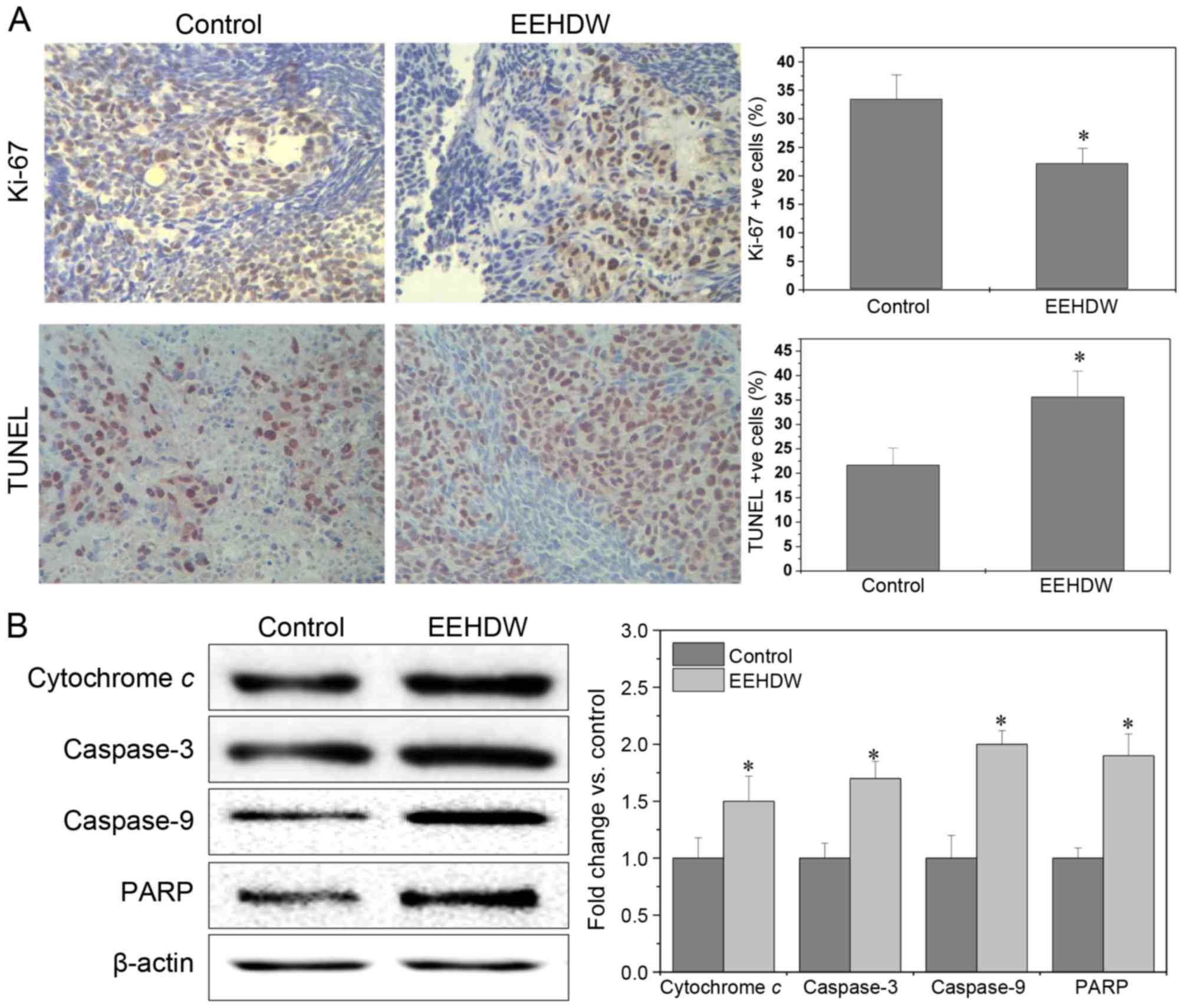 | Figure 2.EEHDW inhibits cellular proliferation
and induces apoptosis in xenograft tumors. (A) Staining for Ki-67
and TUNEL were performed to examine the in vivo effect of
EEHDW on proliferation and apoptosis. The photographs are
representative images captured at a magnification of ×400. The
percentage of positively stained cells was also quantified. To rule
out any non-specific staining, PBS was used to replace the primary
antibody as a negative control. Data are expressed as the mean ±
standard deviation from 10 individual mice in each group.
*P<0.05 vs. control mice. (B) A total of three tumors were
randomly selected from each group, and the level of cytochrome C,
caspase-3, −9 and PARP in tumor tissues was determined by western
blotting to examine the in vivo effect of EEHDW on
apoptosis. β-actin was used as the internal control. For each tumor
sample, western blotting was performed in triplicate. In
densitometric analysis, the expression of the target proteins was
normalized to the mean protein expression of control. *P<0.05,
vs. control mice. EEHDW, ethanol extract of Hedyotis Diffusa
Willd. TUNEL, terminal deoxynucleotidyl-transferase-mediated dUTP
nick end labeling; PARP, poly(ADP-ribose)polymerase 1. |
EEHDW regulates the expression of
Pim-1, Bcl-2, Bax and angiogenic factors COX-2, iNOS, eNOS and
HIF-1α
To further investigate how EEHDW is able to inhibit
tumor growth, IHC staining was performed to detect the expression
of Pim-1, Bcl-2 and Bax in mice. The percentage of Pim-1, Bcl-2 or
Bax-positive cells in the control group was, 37.71±7.31, 27.33±3.67
and 18.50±3.83% respectively, whereas in EEHDW-treated mice the
percentage was 21.75±4.57, 16.00±4.31 and 35.00±4.32%, respectively
(P<0.05; Fig. 3). These data
suggested that that EEHDW treatment was able to significantly
(P<0.02) downregulate the expression of Pim-1, a potential
oncogene, while decreasing Bcl-2 expression and increasing Bax
expression, demonstrating that EEHDW inhibits cell proliferation
and induces apoptosis in xenograft tumors. In addition, the
percentage of COX-2, iNOS, eNOS or HIF-1α-positive cells in the
control group was 33.00±5.67, 45.33±6.93, 30.33±5.87, and
28.50±4.77%, respectively, whereas in EEHDW-treated mice the
percentage was 23.75±4.12, 27.25±4.24, 20.25±3.78 and 17.00±2.86%.
These results suggested that EEHDW treatment was also able to
downregulate the expression of COX-2, iNOS, eNOS and HIF-1α
(P<0.05; Fig. 3), which are novel
tumor markers and are hypothesized to have important roles during
tumor angiogenesis (26).
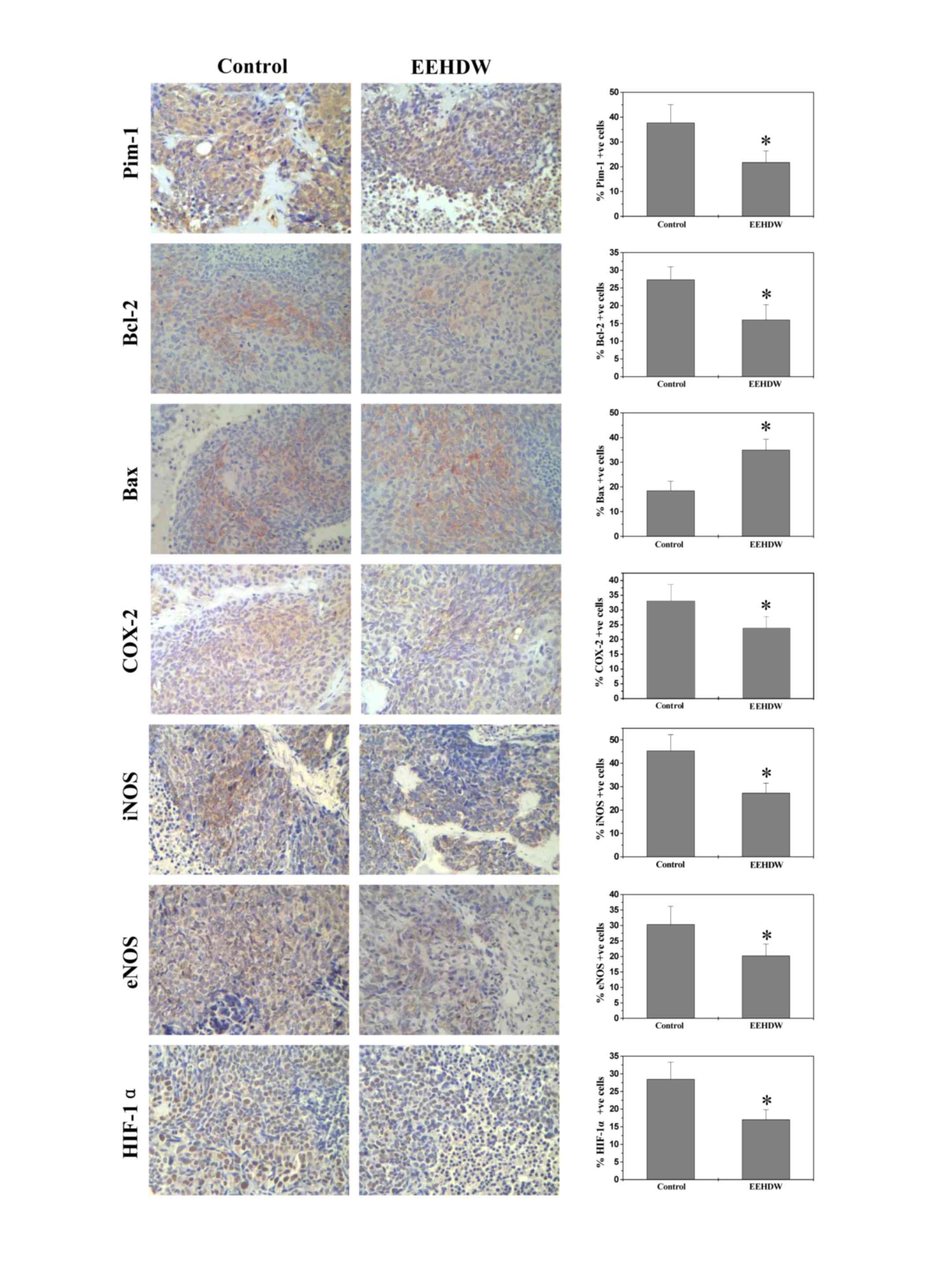 | Figure 3.EEHDW regulates the expression of
Pim-1, Bcl-2, Bax and angiogenic factors COX-2, iNOS, eNOS and
HIF-1α. Tumor tissues were processed for immunohistochemical
staining of Pim-1, Bcl-2, Bax, COX-2, iNOS, eNOS and HIF-1α. The
photographs are representative images taken at a magnification of
×400. The percentage of positively stained cells was also
quantified. Data are expressed as the mean ± standard deviation
from 10 mice in each group. *P<0.05 vs. control mice. EEHDW,
ethanol extract of Hedyotis Diffusa Willd; Bcl-2, B-cell
lymphoma 2; Bax, Bcl-2-like protein 4; COX-2, cyclooxygenase-2;
iNOS, inducible nitric oxide synthase; eNOS, endothelial nitric
oxide synthase; HIF1-α, hypoxia-inducible factor 1-α. |
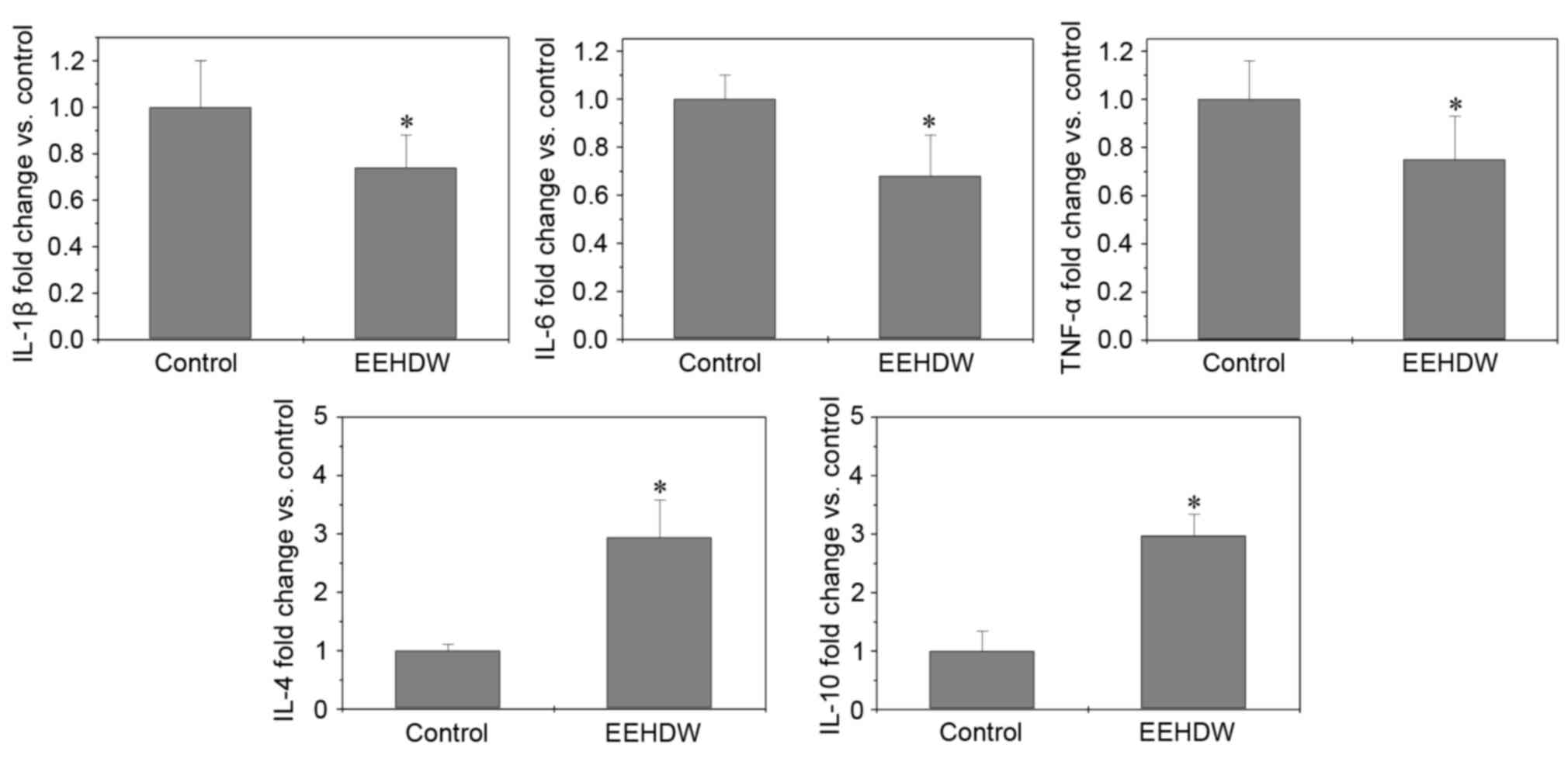
EEHDW regulates inflammatory cytokines
in serum
Dense infiltration of cytokine-producing immune
cells is frequently observed in cancer tissues: Each immune cell
subset and cytokine involved in the activation of intracellular
pathways sustains the growth of cancer cells (27). The effect of EEHDW treatment on
secreting cytokines was detected using ELISA, whereby the levels of
IL-1β, IL-6 and TNF-α were significantly (P<0.026) decreased
following EEHDW treatment, By contrast, the levels of IL-4 and
IL-10 were upregulated following EEHDW treatment compared with the
control group (P<0.05; Fig.
4).
EEHDW regulates multiple signaling
pathways in vivo and in vitro
To investigate the underlying mechanisms of EEHDW in
inhibiting tumor growth, the effect of EEHDW on the activation of
various CRC-associated signal transduction cascades was determined.
The effect of EEHDW treatment on the activation and phosphorylation
of AKT, Erk1/2, JNK, p38, p70S6K p53 and STAT3 in CRC xenograft
tumor tissues and HT-29 cells was determined using Bio-Plex
Phosphoprotein assay.
Treatment with EEHDW was able to significantly
decrease the levels of phosphorylated AKT, Erk1/2, JNK, p38, p70S6K
and STAT3 in tumor tissues (P<0.05; Fig. 5A) and HT-29 cells compared with the
control (P<0.05; Fig. 5B). By
contrast, p-p53 expression was significantly increased following
EEHDW treatment (P<0.05; Fig. 5A and
B).
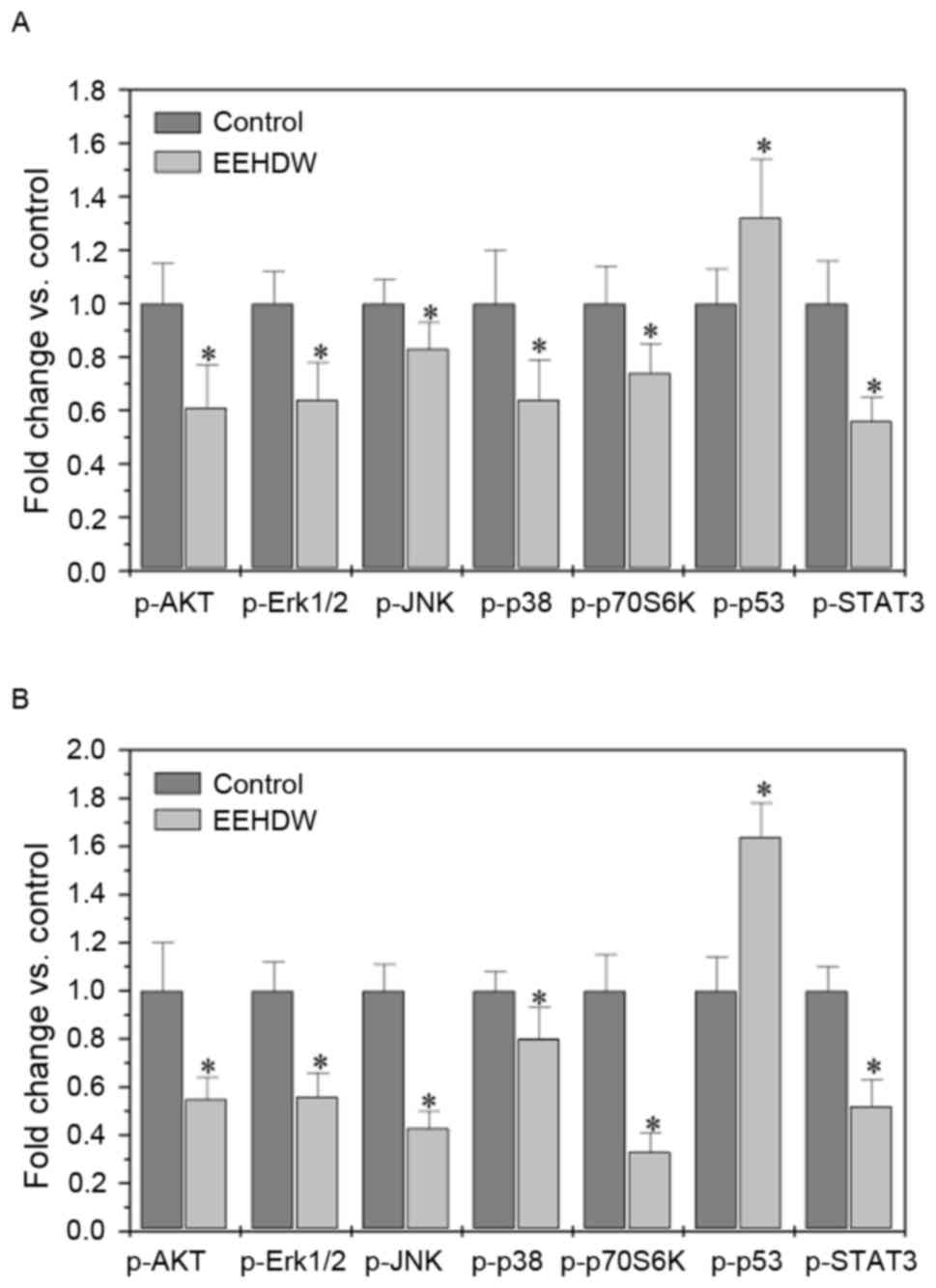 | Figure 5.EEHDW regulates multiple signaling
pathways in vivo and in vitro. The levels of
phosphorylated AKT, Erk1/2, JNK, p38, p70S6 K, STAT3 and p53 in
tumor tissues (A) or HT-29 cells (B) were determined by Bio-Plex
phosphoprotein assay. The data are normalized to the levels of
phosphorylated proteins in the controls. Data are expressed as the
mean ± standard deviation from 8 mice in each group or from 3
independent in vitro experiments. *P<0.05, vs. control.
AKT, protein kinase B; EEHDW, ethanol extract of Hedyotis
Diffusa Willd; Erk1/2, mitogen-activated protein kinase ½; JNK,
c-Jun N-terminal kinase; p-, phosphorylated; STAT3, signal
transducer and activator of transcription 3. |
Discussion
CRC is a serious global health problem with high
morbidity and mortality rates. To date, chemotherapy has been the
main therapeutic treatment for the majority of patients with CRC
(2–4).
However, clinical efficacy for CRC chemotherapy remains poor due to
increased drug resistance and high toxicity resulting in adverse
side effects (1).
Cancer development is tightly regulated by multiple
intracellular signaling pathways, including AKT, mitogen-activated
protein kinase (MAPK) and STAT3 pathways, which are usually
functionally redundant (6). The AKT
signaling pathway is activated by factors, which induce
phosphoinositide 3-kinase activation, which in turn activates the
mTOR signaling pathway. The subsequent abnormal regulation of p53
and p70S6K leads to abnormalities in cell proliferation, apoptosis
and differentiation. AKT signaling has been implicated in various
types of cancer and is frequently implicated in anti-cancer
therapeutics (28–30).
The MAPK signaling pathway is one of the main
pathways involved in cell survival and proliferation. In mammals,
there are three major subfamilies of MAPK, including ERK, JNK and
p38. Activation of MAPK signaling is regulated by a central
three-tiered kinase core, which consists of MAPK kinase kinase
(MAP3K), MAPK kinase (MAP2K) and MAPK (31,32). MAPK
signaling has an important role in intestinal epithelial
differentiation (33). However,
aberrant activation of the MAPK signaling pathway can result in
colon carcinogenesis (34–37). STAT3 is a transcription factor that
exerts an essential role in cell survival and proliferation, via
STAT3 activation, dimerization and subsequent nuclear translocation
(38,39). Constitutive activation of STAT3 is
associated with the development of various types of cancer
including CRC and represents poor disease prognosis (40–43). These
molecular pathways modulate the expression of key genes involved in
the regulation of cell proliferation, apoptosis and angiogenesis,
and are therefore important in the induction, progression and
metastasis of colorectal cancer (44).
The development of CRC is often characterized by
complex interactions within the host immune system, ultimately
resulting in the uncontrolled growth of transformed cells. For
instance, chronic inflammation is an independent risk factor for
the development of CRC, as well as other malignancies, including
gastric cancer, hepatocellular and prostate carcinoma (45). Notably, patients who suffer from
ulcerative colitis have a marked increase in the incidence of CRC
(46). Experimental models of
inflammation-associated colon carcinogenesis have suggested that
inflammatory cell-derived cytokines can either directly or
indirectly stimulate the growth of cancer cells (47–53).
Nevertheless, under certain inflammatory conditions, immune cells
can also mediate the anti-tumor response in order to eliminate
dysplastic and cancerous cells (54,55).
Notably, sporadic CRC, which represent the majority of CRC cases,
exhibits extensive inflammatory infiltration with high levels of
cytokine expression in the tumor microenvironment (27). The ability of immune cells to control
cancer cell proliferation is mostly dependent on cytokines, which
can directly target CRC cells, as well as contribute to the
abnormal activation of various signaling pathways (40,47–51). For
example, high levels of TNF-α and IL-6 expression have been
hypothesized to be associated with redundant activation of STAT3
pathway, which can lead to the progression of CRC (45).
Natural products, including those used in TCM, have
long been used to treat multiple diseases, including cancer
(17). TCM formulae consist of a
combination of many natural products, each of which contains
numerous chemical compounds. Therefore, TCM prescription is often
considered as a multi-component and multi-target method of
treatment, by exhibiting a broad range of therapeutic functions
(56–62). One such prescription is EEHDW, which
has exhibited specific anti-cancer activities (23).
In the present study, MTS was used to demonstrate
that EEHDW reduced cell viability in different CRC cell lines in a
dose- and time-dependent manner. It was also observed that EEHDW
was able to significantly reduce tumor weight and volume in
vivo. Furthermore, it was demonstrated that treatment with
EEHDW was able to inhibit proliferation and promote apoptosis as
indicated by the decrease in the percentage of Ki-67 positive cells
and increasing the percentage of TUNEL positive cells. Notably, the
upregulation of cytochrome c, caspase-3, caspase-9, PARP and Bax
and the downregulation of Pim-1, Bcl-2, COX-2, iNOS, eNOS and
HIF-1α also provided further support to suggest that EEHDW is able
to exert several effects on CRC, including the promotion of
apoptosis, and the suppression of proliferation and angiogenesis.
Although signaling pathways, including AKT, MAPK and STAT3 and
cytokines such as IL-1β, IL-6, TNF-α, IL-4 and IL-10 displayed
abnormal conditions in CRC, the present study has demonstrated that
EEHDW treatment was able to alter the regulation of the multiple
signaling pathways and cytokines.
In conclusion, EEHDW exerts its anti-cancer
activities via altering multiple targets. Therefore, EEHDW may be
developed as a promising multi-potent anti-cancer agent for the
clinical treatment of CRC.
Acknowledgements
The present study was sponsored by the Research Fund
for the Doctoral Program of Higher Education of China (grant no.
20133519110003) and the Developmental Fund of Chen Keji Integrative
Medicine (grant nos. CKJ2014013 and CKJ2015007).
Glossary
Abbreviations
Abbreviations:
|
CRC
|
colorectal cancer
|
|
EEHDW
|
ethanol extract of Hedyotis diffusa
Willd
|
|
TCM
|
traditional Chinese medicine
|
References
|
1
|
Siegel R, Miller K and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Center MM, Jemal A, Smith RA and Ward E:
World wide variations in colorectal cancer. CA Cancer J Clin.
59:366–378. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li H, Mao W, Wang X, Yu C and Du L:
Incidence and mortality of cancer in Zhejiang province in 2009.
Zhonghua Yu Fang Yi Xue Za Zhi. 47:592–596. 2013.(In Chinese).
PubMed/NCBI
|
|
5
|
Cunningham D, Atkin W, Lenz HJ, Lynch HT,
Minsky B, Nordlinger B and Starling N: Colorectal cancer. Lancet.
375:1030–1047. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jiang WQ, Fu FF, Li YX, Wang WB, Wang HH,
Jiang HP and Teng LS: Molecular biomarkers of colorectal cancer:
Prognostic and predictive tools for clinical practice. J Zhejiang
Univ Sci B. 13:663–675. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Aakif M, Balfe P, Elfaedy O, Awan FN,
Pretorius F, Silvio L, Castinera C and Mustafa H: Study on
colorectal cancer presentation, treatment and follow-up. Int J
Colorectal Dis. 31:1361–1363. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Du B and Shim JS: Targeting
epithelial-mesenchymal transition (EMT) to overcome drug resistance
in cancer. Molecules. 21:E9652016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Van Cutsem E, Nordlinger B and Cervantes
A: Advanced colorectal cancer: ESMO clinical practice guidelines
for treatment. Ann Oncol. 21:93–97. 2010. View Article : Google Scholar
|
|
10
|
Tournigand C, André T, Achille E, Lledo G,
Flesh M, Mery-Mignard D, Quinaux E, Couteau C, Buyse M, Ganem G, et
al: FOLFIRI followed by FOLFOX6 or the reverse sequence in advanced
colorectal cancer: A randomized GERCOR study. J Clin Oncol.
22:229–237. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gustin DM and Brenner DE: Chemoprevention
of colon cancer: Current status and future prospects. Cancer
Metastasis Rev. 21:323–348. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gorlick R and Bertino JR: Drug resistance
in colon cancer. Semin Oncol. 26:606–611. 1999.PubMed/NCBI
|
|
13
|
Longley DB, Allen WL and Johnston PG: Drug
resistance, predictive markers and pharmacogenomics in colorectal
cancer. Biochim Biophys Acta. 1766:184–196. 2006.PubMed/NCBI
|
|
14
|
Boose G and Stopper H: Genotoxicity of
several clinically used topoisomerase II inhibitors. Toxicol Lett.
116:7–16. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin J, Chuang CC and Zuo L: Potential
roles of microRNAs and ROS in colorectal cancer: Diagnostic
biomarkers and therapeutic targets. Oncotarget. 8:17328–17346.
2017.PubMed/NCBI
|
|
16
|
Ma X and Wang Z: Anticancer drug discovery
in the future: An evolutionary perspective. Drug Discov Today.
14:1136–1142. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang JJ, Hsu HY, Ho YH and Lin CC:
Comparative study on the immunocompetent activity of three
different kinds of Peh-Hue-Juwa-Chi-Cao, Hedyotis diffusa, H.
corymbosa and Mollugo pentaphylla after sublethal whole body
x-irradiation. Phytother Res. 11:428–432. 1997. View Article : Google Scholar
|
|
18
|
Song L: Zhonghuabencao. 61. Shanghai
Science and Technology Press; Shanghai: pp. 4331999, (In
Chinese).
|
|
19
|
Li R, Zhao H and Lin Y: Anti-tumor effect
and protective effect on chemotherapeutic damage of water soluble
extracts from Hedyotis diffusa. J Chin Pharmaceu Sci. 11:54–58.
2002.
|
|
20
|
Lin J, Chen Y, Wei L, Chen X, Xu W, Hong
Z, Sferra TJ and Peng J: Hedyotis diffusa willd extract induces
apoptosis via activation of the mitochondrion-dependent pathway in
human colon carcinoma cells. Int J Oncol. 37:1331–1338.
2010.PubMed/NCBI
|
|
21
|
Lin J, Wei L, Xu W, Hong Z, Liu X and Peng
J: Effect of hedyotis diffusa willd extract on tumor angiogenesis.
Mol Med Rep. 4:1283–1288. 2011.PubMed/NCBI
|
|
22
|
Cai Q, Lin J, Wei L, Zhang L, Wang L, Zhan
Y, Zeng J, Xu W, Shen A, Hong Z and Peng J: Hedyotis diffusa willd
inhibits colorectal cancer growth in vivo via inhibition of STAT3
signaling pathway. Int J Mol Sci. 13:6117–28. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lin M, Lin J, Wei L, Xu W, Hong Z, Cai Q,
Peng J and Zhu D: Hedyotis diffusa willd extract inhibits HT-29
cell proliferation via cell cycle arrest. Exp Ther Med. 4:307–310.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin J, Wei L, Shen A, Cai Q, Xu W, Li H,
Zhan Y, Hong Z and Peng J: Hedyotis diffusa willd extract
suppresses sonic hedgehog signaling leading to the inhibition of
colorectal cancer angiogenesis. Int J Oncol. 42:651–656. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li Q, Wang X, Shen A, Zhang Y, Chen Y,
Sferra TJ, Lin J and Peng J: Hedyotis diffusa willd overcomes
5-fluorouracil resistance in human colorectal cancer HCT-8/5-FU
cells by downregulating the expression of P-glycoprotein and
ATP-binding casette subfamily G member 2. Exp Ther Med.
10:1845–1850. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cartana ET, Gheonea DI, Cherciu IF,
Streaţa I, Uscatu CD, Nicoli ER, Ioana M, Pirici D, Georgescu CV,
Alexandru DO, et al: Assessing tumor angiogenesis in colorectal
cancer by quantitative contrast-enhanced endoscopic ultrasound and
molecular and immunohistochemical analysis. Endosc Ultrasound.
6:7–17. 2017.
|
|
27
|
Mimatsu K, Fukino N, Ogasawara Y, Saino Y
and Oida T: Utility of inflammatory marker- and nutritional
status-based prognostic factors for predicting the prognosis of
stage iv gastric cancer patients undergoing non-curative surgery.
Anticancer Res. 37:4215–4222.. 2017.PubMed/NCBI
|
|
28
|
Manning BD and Cantley LC: AKT/PKB
signaling: Navigating downstream. Cell. 129:1261–1274. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wullschleger S, Loewith R and Hall MN: TOR
signaling in growth and metabolism. Cell. 124:471–484. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shima H, Pende M, Chen Y, Fumagalli S,
Thomas G and Kozma SC: Disruption of the p70(s6k)/p85(s6k) gene
reveals a small mouse phenotype and a new functional S6 kinase.
EMBO J. 17:6649–6659. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sebolt-Leopold JS: Development of
anticancer drugs targeting the MAP kinase pathway. Oncogene.
19:6594–6599. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Seger R and Krebs EG: The MAPK signaling
cascade. FASEB J. 9:726–735. 1995.PubMed/NCBI
|
|
33
|
Taupin D and Podolski DK:
Mitogen-activated protein kinase activation regulates intestinal
epithelial differentiation. Gastroenterology. 116:1072–1080. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang X, Wang Q, Hu W and Evers BM:
Regulation of phorbol ester-mediated TRAF1 induction in human colon
cancer cells through a PKC/RAF/ERK/NF-kappaB-dependent pathway.
Oncogene. 23:1885–1895. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Roberts PJ and Der CJ: Targeting the
Raf-MEK-ERK mitogen-activated protein kinase cascade for the
treatment of cancer. Oncogene. 26:3291–3310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schwartsmann G, Di Leone LP, Dal Pizzol F
and Roesler R: MAPK pathway activation in colorectal cancer: A
therapeutic opportunity for GRP receptor antagonists. Lancet Oncol.
6:444–445. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fang JY and Richardson BC: The MAPK
signalling pathways and colorectal cancer. Lancet Oncol. 6:322–327.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Aggarwal BB, Kunnumakkara AB, Harikumar
KB, Gupta SR, Tharakan ST, Koca C, Dey S and Sung B: Signal
transducer and activator of transcription-3, inflammation and
cancer: How intimate is the relationship? Ann NY Acad Sci.
1171:59–76. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhong Z, Wen Z and Darnell J: Stat3: A
STAT family member activated by tyrosine phosphorylation in
response to epidermal growth factor and interleukin-6. Science.
264:95–98. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bromberg J and Wang TC: Inflammation and
cancer: IL-6 and STAT3 complete the link. Cancer Cell. 15:79–80.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kusaba T, Nakayama T, Yamazumi K, Yakata
Y, Yoshizaki A, Inoue K, Nagayasu T and Sekine I: Activation of
STAT3 is a marker of poor prognosis in human colorectal cancer.
Oncol Rep. 15:1445–1451. 2006.PubMed/NCBI
|
|
42
|
Lin Q, Lai R, Chirieac LR, Li C, Thomazy
VA, Grammatikakis I, Rassidakis GZ, Zhang W, Fujio Y, Kunisada K,
et al: Constitutive activation of JAK3/STAT3 in colon carcinoma
tumors and cell lines: Inhibition of JAK3/STAT3 signaling induces
apoptosis and cell cycle arrest of colon carcinoma cells. Am J
Pathol. 167:969–980. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xiong H, Zhang ZG, Tian XQ, Sun DF, Liang
QC, Zhang YJ, Lu R, Chen YX and Fang JY: Inhibition of JAK1,
2/STAT3 signaling induces apoptosis, cell cycle arrest and reduces
tumor cell invasion in colorectal cancer cells. Neoplasia.
10:287–297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
García-Caballero M, Paupert J, Blacher S,
Van de Velde M, Quesada AR, Medina MA and Noël A: Targeting
VEGFR-3/-2 signaling pathways with AD0157: A potential strategy
against tumor-associated lymphangiogenesis and lymphatic
metastases. J Hematol Oncol. 10:1222017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fearon ER and Vogelstein B: A genetic
model for colorectal tumorigenesis. Cell. 61:759–767. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Eaden JA, Abrams KR and Mayberry JF: The
risk of colorectal cancer in ulcerative colitis: A meta-analysis.
Gut. 48:526–535. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Grivennikov S, Karin E, Terzic J, Mucida
D, Yu GY, Vallabhapurapu S, Scheller J, Rose-John S, Cheroutre H,
Eckmann L and Karin M: IL-6 and Stat3 are required for survival of
intestinal epithelial cells and development of colitis-associated
cancer. Cancer Cell. 15:103–113. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huber S, Gagliani N, Zenewicz LA, Huber
FJ, Bosurgi L, Hu B, Hedl M, Zhang W, O'Connor W Jr, Murphy AJ, et
al: IL-22BP is regulated by the inflammasome and modulates
tumorigenesis in the intestine. Nature. 491:259–263. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hyun YS, Han DS, Lee AR, Eun CS, Youn J
and Kim HY: Role of IL-17A in the development of colitis-associated
cancer. Carcinogenesis. 33:931–936. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Jauch D, Martin M, Schiechl G, Kesselring
R, Schlitt HJ, Geissler EK and Fichtner-Feigl S: Interleukin 21
controls tumour growth and tumour immunosurveillance in
colitis-associatedtumorigenesis in mice. Gut. 60:1678–1686. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kirchberger S, Royston DJ, Boulard O,
Thornton E, Franchini F, Szabady RL, Harrison O and Powrie F:
Innate lymphoid cells sustain colon cancer through production of
interleukin-22 in a mouse model. J Exp Med. 210:917–931. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Popivanova BK, Kitamura K, Wu Y, Kondo T,
Kagaya T, Kaneko S, Oshima M, Fujii C and Mukaida N: Blocking
TNF-alpha in mice reduces colorectal carcinogenesis associated with
chronic colitis. J Clin Invest. 118:560–570. 2008.PubMed/NCBI
|
|
53
|
Stolfi C, Rizzo A, Franzè E, Rotondi A,
Fantini MC, Sarra M, Caruso R, Monteleone I, Sileri P,
Franceschilli L, et al: Involvement of interleukin-21 in the
regulation of colitis-associated colon cancer. J Exp Med.
208:2279–2290. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Lanca T and Silva-Santos B: The split
nature of tumor-infiltrating leukocytes: Implicationsfor cancer
surveillance and immunotherapy. Oncoimmunology. 1:717–725. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Monteleone G, Pallone F and Stolfi C: The
dual role of inflammation in colon carcinogenesis. Int J Mol Sci.
13:11071–11084. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Newman DJ, Cragg GM and Snader KM: The
influence of natural products upon drug discovery. Nat Prod Rep.
17:215–234. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Gordaliza M: Natural products as leads to
anticancer drugs. Clin Transl Oncol. 9:767–776. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Peng J, Chen Y, Lin J, Zhuang Q, Xu W,
Hong Z and Sferra TJ: Patrinia scabiosaefolia extract suppresses
proliferation and promotes apoptosis by inhibiting STAT3 pathway in
human multiple myeloma cells. Mol Med Rep. 4:313–318.
2011.PubMed/NCBI
|
|
59
|
Cai Q, Lin J, Wei L, Zhang L, Wang L, Zhan
Y, Zeng J, Xu W, Shen A, Hong Z and Peng J: Hedyotis diffusa Willd
inhibits colorectal cancer growth in vivo via inhibition of STAT3
signaling pathway. Int J Mol Sci. 13:6117–6128. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wei L, Chen Y, Lin J, Zhao Y, Chen X, Xu
W, Liu X, Sferra TJ and Peng J: Scutellaria barbata D. don induces
apoptosis of human colon carcinoma cell through activation of the
mitochondrion-dependent pathway. J Med Plants Res. 5:1962–1970.
2011.
|
|
61
|
Wei L, Lin J, Xu W, Hong Z, Liu X, Sferra
TJ and Peng J: Inhibition of tumor angiogenesis by scutellaria
barbata D. don via suppressing proliferation, migration and tube
formation of endothelial cells and downregulation of the expression
of VEGF-A in cancer cells. J Med Plants Res. 5:3260–3268. 2011.
|
|
62
|
Zheng L, Chen Y, Lin W, Zhuang Q, Chen X,
Xu W, Liu X, Peng J and Sferra TJ: Spica Prunellae extract promotes
mitochondrion-dependent apoptosis in a human colon carcinoma cell
line. Afr J Pharm Pharmacol. 5:327–335. 2011. View Article : Google Scholar
|