Introduction
Colon cancer is the third most commonly diagnosed
malignancy and the fourth leading cause of cancer mortality
worldwide (1). Its late tumor
presentation and rapid progression contribute to the high mortality
rate (1). Colorectal neoplasia
appears to be influenced by environmental factors and an
accumulation of mutations that result in uncontrolled proliferation
and angiogenesis, inhibition of apoptosis, and immune escape
(2–4).
Therefore, an improved understanding of the molecular mechanisms of
colon tumorigenesis and progression may provide more options for
treatment and improved prognosis of colorectal cancer patients.
It is known that less than 2% of the mammalian
genome represents protein-coding genes and over 90% represents
noncoding RNAs (ncRNA), which are transcribed but not translated
into proteins (5,6). MicroRNAs (miRNAs/miRs) belong to a class
of small (typically 18–25 nucleotides), single-stranded ncRNAs that
regulate post-transcriptional gene expression by base-pairing with
complementary sequences on mRNA and inhibiting protein translation
(5,6).
Recently, growing evidence has shown that miRNAs may be involved in
the pathogenesis of several cancers, providing new insight into the
biology of cancer (7–10). A previous study has indicated that
miR-370 functions as a tumor suppressor in laryngeal squamous cell
carcinoma (LSCC) through downregulation of FoxM1 (11). In ovarian cancer, miR-370 was
down-regulated by hypermethylation and overexpression of miR-370
suppressed ovarian cancer cell viability, reduced colony formation,
and enhanced ovarian cancer cell chemosensitivity to cisplatin
(cDDP) by directly targeting Endoglin (ENG) (12). However, results by Wu et al
demonstrated that miR-370 plays an important role in the
proliferation of human prostate cancer cells, by directly
suppressing the tumor suppressor FOXO1, which has been associated
with key cellular functions including cell growth, differentiation,
apoptosis, and angiogenesis (13).
The biological role of miR-370 in colon cancer and
the underlying molecular mechanism remains undefined. In the
present study, we studied the expression pattern of miR-370 and
investigated its function in human colon cancer. Moreover, the
regulation of colon cancer apoptosis by miR-370 was also examined
in order to clarify the underlying mechanism of action. Our
findings will provide a novel target in the diagnosis and treatment
of colon cancer.
Materials and methods
Clinical sample
Human normal colon tissues and colonic tissues were
obtained with informed consent under a general waiver by the
Academic Medical Center Institutional Review Board for the proper
secondary use of human material and were obtained from the People's
Hospital of Sichuan from October 2016 to December 2016. Experiments
described were approved by the Ethics Committee of Sichuan Academy
of Medical Sciences and Sichuan Provincial People's Hospital
(Chengdu, China). The tumor grade were identified according to
clinical diagnosis.
Cell culture and treatment
Human colon cancer cell HT29, DLD-1, SW480, LoVo,
HCT116 and SW620 were obtained from the American Type Culture
Collection (Manassas, VA, USA). The human colon epithelial cell
HCoEpiC was purchased from Shanghai Guandao Biotechnology. The
cells were cultured in DMEM medium containing 10% fetal bovine
serum (both from Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) and 1% penicillin and streptomycin at 37°C in 5%
CO2 incubator. The plasmid-based miR-370 expression
system and miR-NC were transfected into cells using FuGENE
HP® (Roche Diagnostics, Basel, Switzerland). The mouse
double minute 4 (MDM4) specific inhibitor NSC207895 was purchased
from Selleck Bio (MA, USA) and used to treat LoVo cells that
transfected with miR-NC or miR-370.
Cell Counting kit-8 (CCK-8) kit. For cell
viability detection, cells were seeded at 800 cells per well in
96-well plates. Then, a CCK-8 assay (Beyotime Institute of
Biotechnology, Shanghai, China) was used and absorbance was
measured at 450 nm for each well at different time points using a
micro-plate reader (Thermo Fisher Scientific, Inc.). The same
experiments were performed four times.
Quantitative real-time polymerase chain reaction
(qPCR). Total RNA was extracted from each experimental group using
TRIzol® reagent (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. qPCR for miR-370 was
performed using miRNA primers obtained from Guangzhou RiboBio Co.,
Ltd. (Guangzhou, China). U6 was used as the internal control. The
sequences were not supplied due to the rules of the company. The
qPCR conditions were as follows: Denaturation at 94°C for 2 min,
amplifcation for 30 cycles at 94°C for 0.5 min, annealing at 58°C
for 0.5 min and extension at 72°C for 1 min, followed by a terminal
elongation step at 72°C for 10 min. The RT-qPCR analysis was
performed on a Bio-Rad CFX96 thermal cycler (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). Data was analyzed with the
2−ΔΔCt method. The same experiments were performed three
times. GAPDH was used as the internal control.
Apoptosis analysis
Cells transfected with miR-NC or miR-370 were
harvested at 48 h after transfection and stained with Annexin
V-FITC/PI Apoptosis Detection kit (Keygentec, Nanjing, China) as
the the manufacturer's protocol indicated. Apoptotic cells were
assessed by flow cytometry (FACS Calibur; Becton Dickinson,
Franklin Lakes, NJ, USA). The same experiments were performed three
times.
The generation of stable clones
The SW620 cells were plated in 6-well plate
(2×105 cells/well). Then, the plasmid-based miR-370
expression system and miR-NC were transfected into cells using
FuGENE HP® (Roche Diagnostics). After transfection for
48 h, 2 mg/ml G418 (Sigma-Aldrich, St. Louis, MO, USA) was added
into the medium and lasted for 10 days. The miR-370 expression in
miR-370 and miR-NC cells were determined by qPCR. The survived
cells was named as miR-370 cells (stable expression miR-370) and
miR-NC cells.
Xenograft tumors in nude mice
Procedures involving animals conformed to the
guidelines of the Institutional Animal Care and Use Committee of
Sichuan Academy of Medical Sciences and Sichuan Provincial People's
Hospital (Chengdu, China). SW620 cells stable expression miR-NC or
miR-370 (5×106 cells in 100 µl of DMEM) were injected
subcutaneously into the flanks of nude mice (5 weeks old, male;
Vital River Laboratories, Beijing China). Tumor volumes were
measured at 10 days post cell injection and every five days. Tumor
weights were measured immediately after sacrificing and tumor
samples were harvested for RNA extraction and embedding in paraffin
for TUNEL assay analysis.
TUNEL assay
For apoptotic cell detection in tumor tissues,
serial 4-µm-thick tissue sections were prepared and dewaxed to
water. Then DeadEnd™ Fluorometric TUNEL system (Promega
Corporation, Madison, WI, USA) was performed according to the
manufacturer's and the apoptotic cells is end-labeled with
fluorophore. DAPI (Beyotime, Beijing, China) was purchased to stain
cell nuclei. Fluorescence images were captured using a fluorescence
microscopy (Olympus Corporation, Tokyo, Japan). The apoptotic index
(apoptotic cells of total cell were analyzed. The same experiments
were performed three times.
Luciferase assay
The precursor to miR-370 was synthesized and cloned
in pMiRluc (Ambion Life Technologies, Carlsbad, CA, USA) to
generate pMiRluc-370. Firefly luciferase reporter vectors with the
intact putative miR-370 recognition sequence from the 3′-UTR of
MDM4 or with random mutations cloned downstream of the firefly
luciferase gene were constructed. For the 3′UTR-luciferase assays,
293T cells were co-transfected with MDM4-WT or mut-3′-UTR
construct, pMiRluc or pMiRluc-miR-370 luciferase expression
construct using FuGENE HP® (Roche Diagnostics).
Luciferase assays were performed 24 h after transfection using the
Dual Luciferase Reporter Assay system (Promega Corporation). Values
were normalized to the activity of β-galactosidase.
Western blotting
The proteins were extracted in ice-cold RIPA lysis
buffer (Beyotime) containing 1% protease inhibitor cocktail
(Pierce, Rockford, IL, USA), then centrifuged at 12,000 rpm for 20
min at 4°C. After protein concentration measurement by BCA assay
kit (Beyotime), equivalent amounts of proteins were loaded on
SDS-PAGE and transferred to PVDF membranes (Millipore, Bedford, MA,
USA). After blocking by 5% non-fat milk in TBS/T buffer, antibodies
against MDM4 (cat. no. 04-1555, 1:2,000; Millipore), p53 (cat. no.
2527, 1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA),
γH2AX (cat. no. 9718, 1:1,000; Cell Signaling Technology, Inc.) and
caspase 3 (cat. no. 9662, 1:1,000; Cell Signaling Technology, Inc.)
were added for detection and all blots were probed with antibodies
against β-actin (cat. no. 04-116, 1:3,000; Millipore) as loading
control. ECL kit from Millipore was purchased for band exposure.
The density of each band were measured by Image J software
(National Institute of Health, Bethesda, MD, USA).
Immunohistochemical staining
SW620 xenograft tumors were instilled with 4%
formalin and immersed in the same solution before tissue processing
into paraffin-embedded blocks; 4 µm sections were then cut for
immunohistochemical staining. After antigen retrieval for 3 mins in
citrate under high pressure, tumor tissue sections were incubated
with primary antibody against MDM4 (cat. no. 04-1555, 1:300;
Millipore) for immunohistochemical staining following the
instructions of IHC kit (SP9001; ZsBio, Beijing, China) and DAB kit
(Fuzhou Maixin Biotech Co., Ltd., Fuzhou, China). The stained
tumors were visualized by a light microscope (DTX500; Nikon
Corporation, Tokyo, Japan) The MDM4 positive cells were scored by
counting the number of tumor cells expressing the proteins as
determined by MDM4 staining in 5 random selected fields.
Statistical analysis
Statistical comparisons of all results were analyzed
using analysis of variance (ANOVA) or t-test and represented
graphically as mean ± standard deviation (SD). All statistical
analyses were carried out using SPSS 20.0 statistical software
(SPSS, Inc., Chicago, IL, USA). P<0.05 was regarded as
statistically significant.
Results
miR-370 expression is low in malignant
colon tissue
To determine the expression of miR-370 in normal and
malignant colon tissue, 30 pairs of colon tissues from patients
were collected for qPCR testing (Table
I). Interestingly, miR-370 expression levels were significantly
lower in malignant colon tissues than in normal colon tissues
(relative miR-370 expression in normal colon tissues, 2.772±0.3749;
relative miR-370 expression in colon malignant tissues,
1.199±0.1948; P=0.0004; Fig. 1A).
Further analysis indicated miR-370 expression in grade I tumors was
higher than in grade II–III or grade IV tumors (Fig. 1B). These results suggest that miR-370
is downregulated in malignant colon tissues and is inversely
correlated with tumor grade.
 | Table I.Analysis of the correlation between
miR-370 expression and clinicopathological characteristics. |
Table I.
Analysis of the correlation between
miR-370 expression and clinicopathological characteristics.
|
|
| miR-370
expression |
|
|---|
|
|
|
|
|
|---|
| Characteristics | Number of
patients | Low | High | P-value |
|---|
| Sex (%) |
|
|
| >0.05 |
| Male | 13 (43.3) | 5 (38.5) | 8 (61.5) |
|
|
Female | 17 (56.7) | 9 (52.9) | 8 (47.1) |
|
| Age (%) |
|
|
| >0.05 |
| ≤60 | 10 (33.3) | 4 (40) | 6 (60) |
|
|
>60 | 20 (66.7) | 6 (50) | 6 (50) |
|
| TNM stage (%) |
|
|
|
<0.05 |
| I | 8 (26.7) | 3 (37.5) | 5 (62.5) |
|
|
II–III | 12 (40) | 4 (33.3) | 8 (66.7) |
|
| IV | 10 (33.3) | 2 (20) | 8 (80) |
|
Overexpression of miR-370 promotes
apoptosis in colon cancer cells in vitro
Since miR-370 expression was reduced in malignant
colon tissues, we were interested in its expression and role in
colon cancer cells. We determined the expression level of miR-370
in several colon cancer cell lines and in normal human colon
epithelial cells (HCoEpiC) and found miR-370 expression is
significantly downregulated in colon cancer cells (Fig. 2A). LoVo and SW620, which get lowest
miR-370 expression among the investigated colon cancer cell lines,
were selected for further research. Next, we introduced a
plasmid-based miR-370 expression system and demonstrated that the
plasmid effectively enhanced miR-370 levels in colon cancer cell
lines (LoVo and SW620) (Fig. 2B).
Overexpression of miR-370 in LoVo and SW620 cells resulted in
significantly reduced viability compared to the miR-NC (negative
control) transfected group. Viability was reduced by 52.5% in LoVo
cells and 56.6% in SW620 cells (Fig.
2C). Furthermore, an improved rate of apoptosis in LoVo and
SW620 cells was observed upon treatment with miR-370 (LoVo,
5.2±0.5% for miR-NC vs. 13.1±1.1% for miR-370, P<0.01; SW620,
2.7±0.7%for miR-NC vs. 12.7±1.6% for miR-370, P<0.01; Fig. 2D). These results indicate that
overexpression of miR-370 inhibits colon cell proliferation through
apoptotic pathways, in vitro.
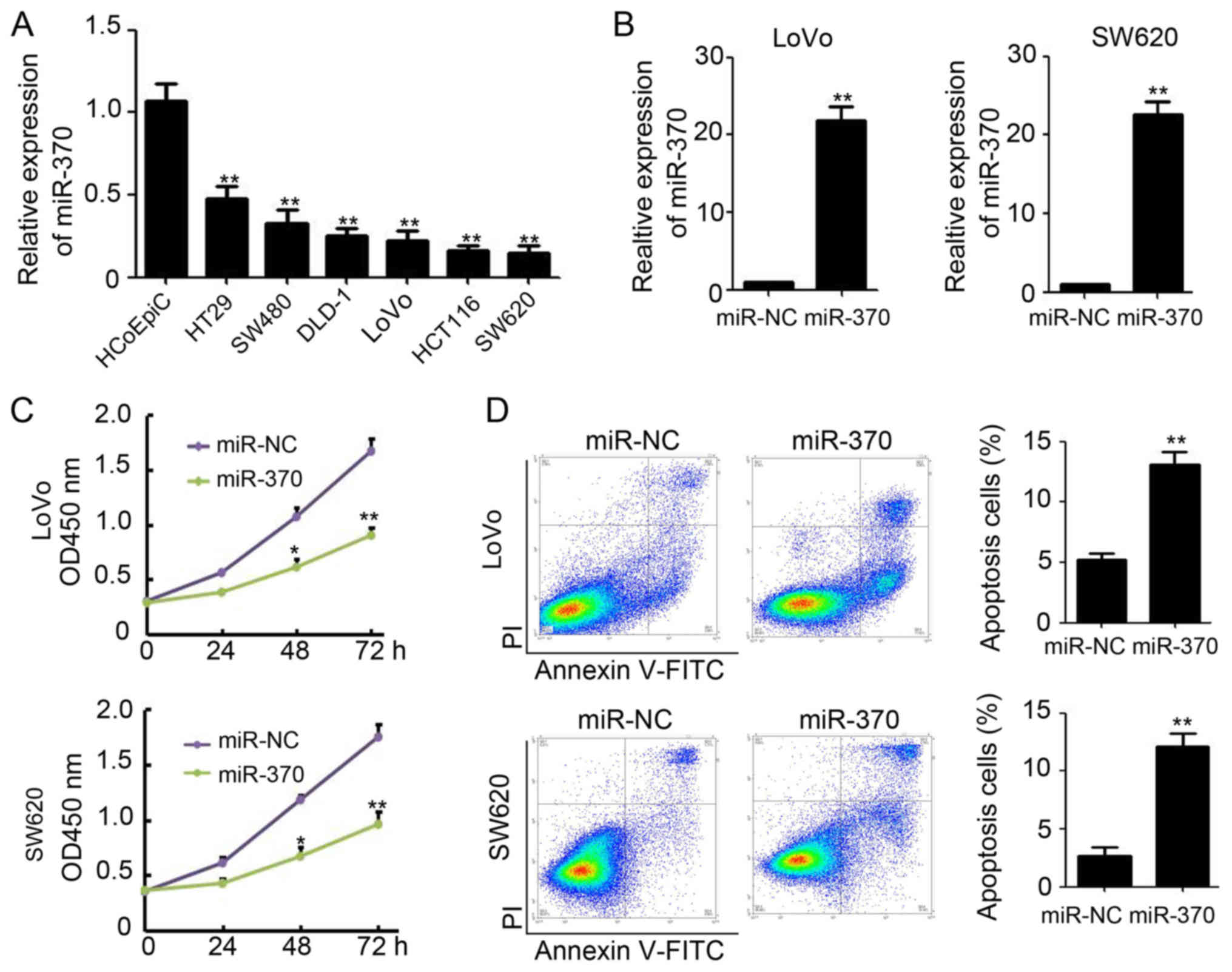 | Figure 2.Overexpression of miR-370 promotes
apoptosis in colon cancer cells in vitro. (A) qPCR analysis
of miR-370 expression in HCoEpiCs and several colon cancer cells,
n=3; **P<0.01, compared with HCoEpiCs. GAPDH was used as the
internal control. (B) qPCR analysis of miR-370 expression in miR-NC
and miR-370 transfected LoVo and SW620 cells, n=3; **P<0.01,
compared with miR-NC transfected cells. GAPDH was used as the
internal control. (C) Analysis of cell viability by CCK-8 in miR-NC
and miR-370 transfected LoVo and SW620 cells, n=3; *P<0.05;
**P<0.01, compared with miR-NC transfected cells. (D) Analysis
of cell apoptosis by Annexin V and PI in miR-NC and miR-370
transfected LoVo and SW620 cells, n=3; **P<0.01, compared with
miR-NC transfected cells. miR, microRNA; qPCR, quantitative
real-time polymerase chain reaction; CCK-8, Cell Counting
kit-8. |
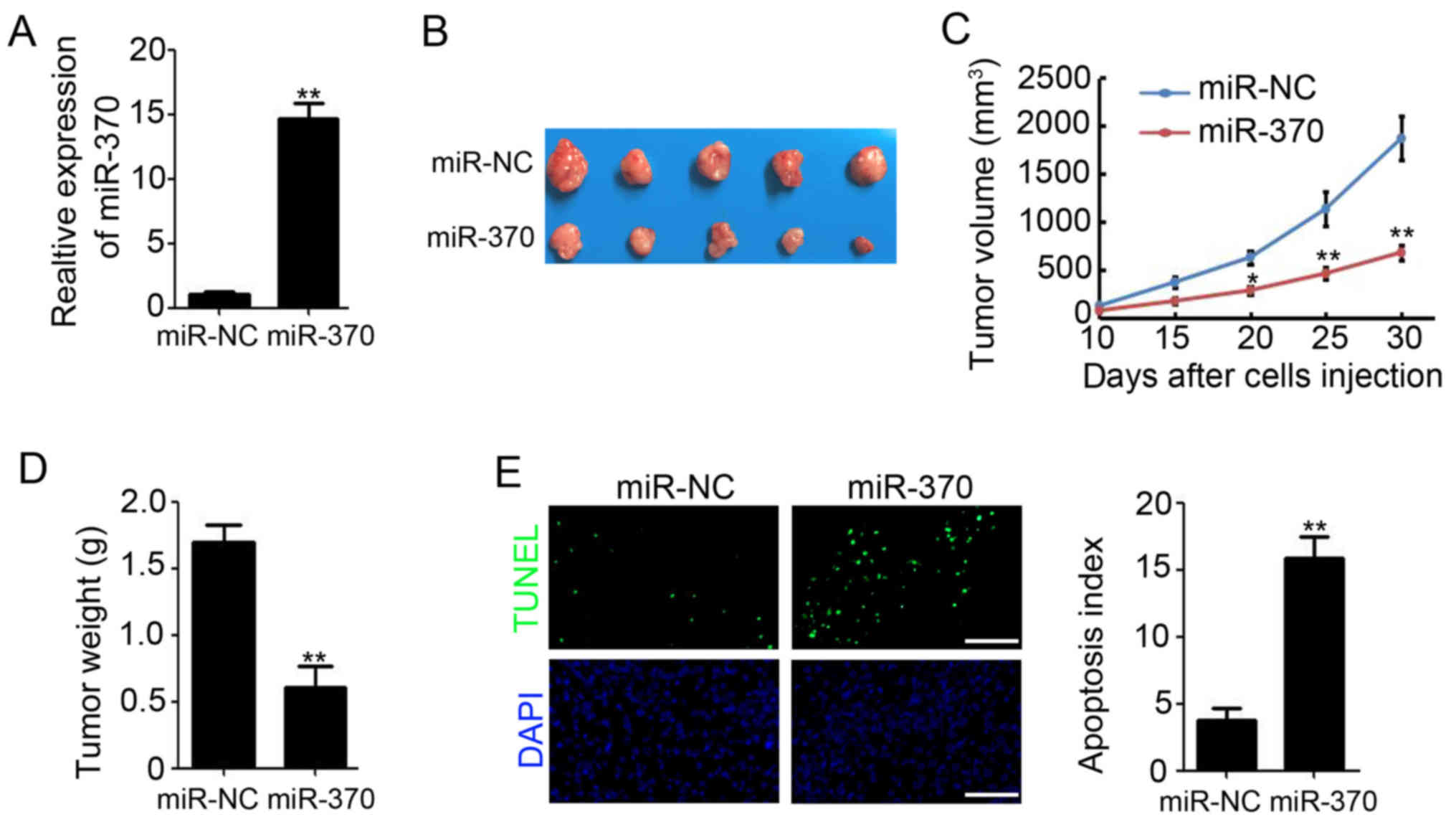
Overexpression of miR-370 reduces
colon tumor growth in vivo
To validate the observed phenomenon in vivo,
SW620 cells that stably express either miR-NC or miR-370 were used
to establish xenograft tumor models in nude mice. qPCR results
showed that the expression of miR-370 in miR-370 stably transfected
SW620 cells was higher than in miR-NC stable transfected SW620
cells (Fig. 3A). We found that
overexpression of miR-370 in colon cancer cells markedly reduced
xenograft tumor growth (Fig. 3B).
Tumor volume decreased by 63.6% (1,872.3±283.8 mm3 for
miR-NC vs. 682.3±83.4 mm3 for miR-370; P<0.01;
Fig. 3C) and tumor weight decreased
by 64.9% (1.69±0.13 g for miR-NC vs. 0.61±0.15 g for miR-370;
P<0.01; Fig. 3D). Furthermore,
TUNEL staining suggested a significantly increased apoptotic index
upon transfection of SW620 xenograft tumors with miR-370 (Fig. 3E). These results indicate that
overexpression of miR-370 retards colon tumor growth in
vivo.
miR-370 directly targets MDM4 to
promote apoptosis in colon cancer cells
To investigate the possible mechanisms by which
miR-370 promotes apoptosis in colon cancer cells, TargetScan and
miRDB were used to identify miR-370's predicted biological target,
MDM4. To verify this analysis, the MDM4 3′UTR, containing the
miR-370 binding site, was cloned downstream of the luciferase open
reading frame. Additionally, a MDM4 3′UTR mutant, which contains a
mutated miR-370 binding site, was also introduced into the
luciferase construct (Fig. 4A). Next,
293T cells, which were widely used as a tool cell in luciferase
assay (14), were performed to
investigate whether miR-370 directly targets MDM4. qPCR results
showed that the expression of miR-370 in miR-370 transfected 293T
cells was higher than in miR-NC transfected 293T cells (Fig. 4B). After the luciferase-MDM4-3′UTR
transfection, luciferase expression in miR-370 transfected 293T
cells was significantly affected (Fig.
4C). Results showed consistent reduction of luciferase
expression in miR-NC and miR-370 transfected 293T cells following
transfection with miRNA binding site mutant plasmids
(luciferase-MDM4-3′UTR mutant) (Fig.
4C). Further western blot analysis demonstrated that miR-370
reduced MDM4 expression in LoVo and SW620 cells, accompanied by
increased p53, caspase 3 and γH2AX expression, both of which are
downstream targets of MDM4 (Fig. 4D).
IHC staining also demonstrated less MDM4 positive cells were found
upon transfection of SW620 xenograft tumors with miR-370, compared
with miR-NC group (Fig. 4E). These
results suggest that miR-370 directly targets MDM4 to promote
apoptosis in colon cancer cells.
 | Figure 4.miR-370 directly targets MDM4. (A)
MDM4-3′UTR contains one predicted miR-370 binding site. The figure
shows predicted duplex formations between MDM4-3′UTR and miR-370.
The sites of target mutagenesis are indicated in blue. (B) qPCR of
miR-370 in 293T cells after enforced expression of miR-370 in 293T
cells. **P<0.01, compared to miR-NC group. (C) Relative
repression of luciferase expression was standardized to a
transfection control. **P<0.01 compared to miR-NC control. (D)
Western blot analysis of MDM4, p53, caspase 3 and γH2AX expression
in miR-NC and miR-370 transfected LoVo and SW620 cells. β-actin was
used as loading control. Relative expression analysis of MDM4, p53,
caspase 3 and γH2AX expression. n=3, **P<0.01, compared with
miR-NC transfected group. (E) IHC staining of MDM4 in miR-NC and
miR-370 xenograft tumor tissues. Scale bar, 50 µm. The percentage
of MDM4 positive cells were counted and analyzed. n=5, **P<0.01,
compared with miR-NC group. miR, microRNA; MDM4, mouse double
minute 4; qPCR, quantitative real-time polymerase chain
reaction. |
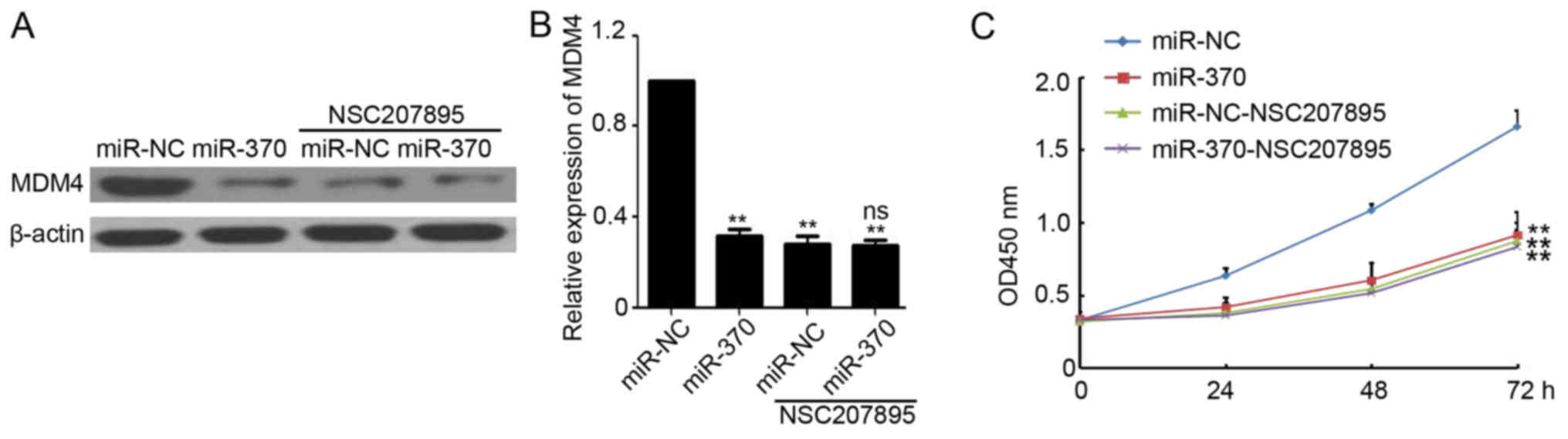
MDM4 plays a necessary role during
miR-370-mediated cancer cell growth inhibition
Next, to investigate whether MDM4 plays a necessary
role during miR-370-mediated cancer cell growth inhibition, MDM4
specific inhibitor NSC207895 was purchased and used to treat
miR-370 or miR-NC transfected LoVo cells. As shown in Fig. 5A and B, NSC207895 efficiently
inhibited the expression of MDM4 in miR-NC transfected LoVo cells.
Meanwhile, there was no significant difference on MDM4 expression
in miR-NC and miR-370 transfected cells that after NSC207895
treatment (Fig. 5A and B).
Furthermore, CCK-8 assay demonstrated that miR-370 transfected
resulted in significantly reduced viability compared to the miR-NC
transfected LoVo cells (Fig. 5C). But
NSC207895 efficiently blocked the miR-370-mediated Loco cell growth
inhibition (Fig. 5C). Collectively,
these results indicated that MDM4 plays a necessary role during
miR-370-mediated cancer cell growth inhibition.
Discussion
In the present study, we investigated the expression
patterns and role of miR-370 in colon cancer cells. We discovered
downregulation of miR-370 expression in malignant colon tissue and
miR-370 expression was inversely correlated with tumor grade.
Moreover, we determined that miR-370 functions as a tumor
suppressor in colon cancer by inhibiting tumor growth and promoting
cell apoptosis. Mechanistically, MDM4 was demonstrated to be a
potential direct target of miR-370 during colon cancer cell
apoptosis. Collectively, these findings suggest that upregulation
of miR-370 may impair colon tumor growth by targeting MDM4.
Recently, case series studies demonstrated the
expression and function of miR-370 in cancer. Previous studies
showed that miR-370 is decreased in the tissues and cell lines of
gastric cancer and overexpression of miR-370 promotes cell
apoptosis and inhibits proliferation (15,16). In
non-small cell lung cancer tissues and cell lines, miR-370
expression was also downregulated, and upregulation of miR-370
impaired tumor growth by inhibiting cell proliferation (17). Meng et al have investigated the
mechanism of miR-370 downregulation; overexpression of IL-6 induced
epigenetic-mediated miR-370 downregulation in malignant human
cholangiocytes (18). However, the
expression and function of miR-370 in colon cancer remains unclear.
In the present study, we reported that miR-370 was downregulated in
colon cancer tissues compared to the adjacent tissues of the tumor,
and confirmed miR-370 downregulation in colon cancer cell lines.
Furthermore, miR-370 expression inversely correlated with tumor
grade. Importantly, we identified the tumor suppressor role of
miR-370 in colon cancer. Overexpression of miR-370 impairs the
xenograft tumor growth and cell proliferation in vivo and
in vitro by promoting colon cancer cell apoptosis. These
results provide solid evidence for understanding the role of
miR-370 in colon cancer. Regrettably, we didn't examine the
expression of miR-370 in clinic samples after effective treatment.
And further research is required to investigate the expression of
miR-370 following treatment.
MDM4, a master regulator of p53, binds its homologue
MDM2 and this results in p53 activity repression through
MDM2-driven ubiquitination (19). As
a cytoplasmic protein, MDM4 negatively controls p53 transcriptional
activity (20). Therefore, MDM4 and
the MDM2-MDM4 complex are the key regulators of p53 protein
stability and are involved in p53 activation (21). In colon cancer, MDM4 was detected in
approximately 50% of human colon tumors and showed strong
correlation with increased extracellular signal-regulated kinase
phosphorylation (22). MDM4
expression in colon cancer was regulated by several factors. FL118
promotes degradation of MDM4, leading to p53-dependent apoptosis
(23). Furthermore, DNA
damage-induced miR-34a overexpression decreases MDM4 protein levels
in colon cancer, based on the presence of a miR-34a binding site
(24). MDM4 expression is also
regulated by mitogenic signaling pathways (22) and genotoxic stress (25). In the present study, luciferase assays
and western blot analysis revealed that MDM4 expression was
downregulated by miR-370. Through an 11 mer binding site, the MDM4
3′-UTR is a target of miR-370. Overexpression of miR-370 in colon
cancer cells inhibits MDM4 protein expression and promotes p53,
caspase 3 and γH2AX expression, resulting in cell apoptosis.
Notably, as shown in Fig. 2C and D,
only 15% apoptotic cells resulted in a 50–60% reduction in
viability/death. It may contributed to that the reduction in cell
viability was a accumulated effect of apoptosis, that was
persistent after miR-370 transfection. Furthermore, MDM4 expression
in SW620 xenograft tumor tissues was also inhibited by
overexpression of miR-370. Previous studies by Li et al
(26) and Pan et al (27) have demonstrated that miR-370 can
activated p21 expression in lung cancer cells and bladder cells and
inhibit PIM1 expression in hepatocellular carcinoma. But in our
study, inhibition of MDM4 by specific inhibitor NSC207895
efficiently blocked miR-370-mediated cell growth inhibition
(Fig. 5). It's demonstrated that MDM4
plays a necessary role during miR-370 inhibiting colon cancer cell
growth. The diversity of mechanism under miR-370 inhibiting cancer
cell growth would contributed to the different cancer cells we
investigated. But whether the miR-370-mediated induction of
apoptotic cell death in colon cancer cells is p53 dependent should
be investigated in the further study. We also need determine the
expression of MDM4 in clinic colon tumors and analyze the
correlation between miR-370 and MDM4 expression in clinic samples.
It may provide solid evidence for understanding the regulation role
of miR-370 on MDM4 expression.
In conclusion, our research demonstrated that there
is downregulation of miR-370 in malignant colon tissue and its
level is inversely correlated with tumor grade. Functional studies
found that miR-370 directly targets the oncogene MDM4 and inhibits
the growth of colon cancer in vitro and in vivo.
Taken together, our data provide clear evidence that miR-370
functions as a tumor suppressor and may serve as an effective
target for the treatment of colon cancer.
Acknowledgements
The present study was supported by Doctoral Fund of
Sichuan Provincial People's Hospital (no. 30305030584).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Calvert PM and Frucht H: The genetics of
colorectal cancer. Ann Intern Med. 137:603–612. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wei EK, Wolin KY and Colditz GA: Time
course of risk factors in cancer etiology and progression. J Clin
Oncol. 28:4052–4057. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dai L, Cui X, Zhang X, Cheng L, Liu Y,
Yang Y, Fan P, Wang Q, Lin Y, Zhang J, et al: SARI inhibits
angiogenesis and tumour growth of human colon cancer through
directly targeting ceruloplasmin. Nat Commun. 7:119962016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chawla JP, Iyer N, Soodan KS, Sharma A,
Khurana SK and Priyadarshni P: Role of miRNA in cancer diagnosis,
prognosis, therapy and regulation of its expression by Epstein-Barr
virus and human papillomaviruses: With special reference to oral
cancer. Oral Oncol. 51:731–737. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gambari R, Brognara E, Spandidos DA and
Fabbri E: Targeting oncomiRNAs and mimicking tumor suppressor
miRNAs: Nuew trends in the development of miRNA therapeutic
strategies in oncology (Review). Int J Oncol. 49:5–32. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Y, Xu Z, Li B, Zhang Z, Luo H, Wang Y,
Lu Z and Wu X: Epigenetic silencing of miRNA-9 is correlated with
promoter-proximal CpG island hypermethylation in gastric cancer in
vitro and in vivo. Int J Oncol. 45:2576–2586. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lv J, Xia K, Xu P, Sun E, Ma J, Gao S,
Zhou Q, Zhang M, Wang F, Chen F, et al: miRNA expression patterns
in chemoresistant breast cancer tissues. Biomed Pharmacother.
68:935–942. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Molina-Pinelo S, Carnero A, Rivera F,
Estevez-Garcia P, Bozada JM, Limon ML, Benavent M, Gomez J, Pastor
MD, Chaves M, et al: MiR-107 and miR-99a-3p predict chemotherapy
response in patients with advanced colorectal cancer. BMC Cancer.
14:6562014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nadal E, Zhong J, Lin J, Reddy RM, Ramnath
N, Orringer MB, Chang AC, Beer DG and Chen G: A MicroRNA cluster at
14q32 drives aggressive lung adenocarcinoma. Clin Cancer Res.
20:3107–3117. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yungang W, Xiaoyu L, Pang T, Wenming L and
Pan X: miR-370 targeted FoxM1 functions as a tumor suppressor in
laryngeal squamous cell carcinoma (LSCC). Biomed Pharmacother.
68:149–154. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen XP, Chen YG, Lan JY and Shen ZJ:
MicroRNA-370 suppresses proliferation and promotes endometrioid
ovarian cancer chemosensitivity to cDDP by negatively regulating
ENG. Cancer Lett. 353:201–210. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu Z, Sun H, Zeng W, He J and Mao X:
Upregulation of MircoRNA-370 induces proliferation in human
prostate cancer cells by downregulating the transcription factor
FOXO1. PLoS One. 7:e458252012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luo P, He T, Jiang R and Li G:
MicroRNA-423-5p targets O-GlcNAc transferase to induce apoptosis in
cardiomyocytes. Mol Med Rep. 12:1163–1168. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zeng Y, Fu M, Wu GW, Zhang AZ, Chen JP,
Lin HY, Fu YA, Jia J, Cai ZD, Wu XJ and Lan P: Upregulation of
microRNA-370 promotes cell apoptosis and inhibits proliferation by
targeting PTEN in human gastric cancer. Int J Oncol. 49:1589–1599.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lo SS, Hung PS, Chen JH, Tu HF, Fang WL,
Chen CY, Chen WT, Gong NR and Wu CW: Overexpression of miR-370 and
downregulation of its novel target TGFbeta-RII contribute to the
progression of gastric carcinoma. Oncogene. 31:226–237. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen T, Gao F, Feng S, Yang T and Chen M:
MicroRNA-370 inhibits the progression of non-small cell lung cancer
by downregulating oncogene TRAF4. Oncol Rep. 34:461–468. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Meng F, Wehbe-Janek H, Henson R, Smith H
and Patel T: Epigenetic regulation of microRNA-370 by interleukin-6
in malignant human cholangiocytes. Oncogene. 27:378–386. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hirose M, Yamato K, Endo S, Saito R, Ueno
T, Hirai S, Suzuki H, Abei M, Natori Y and Hyodo I: MDM4 expression
as an indicator of TP53 reactivation by combined targeting of MDM2
and MDM4 in cancer cells without TP53 mutation. Oncoscience.
1:830–843. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tan BX, Khoo KH, Lim TM and Lane DP: High
Mdm4 levels suppress p53 activity and enhance its half-life in
acute myeloid leukaemia. Oncotarget. 5:933–943. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang MJ, Luo YJ, Shi ZY, Xu XL, Yao GL,
Liu RP and Zhao H: The associations between MDM4 gene polymorphisms
and cancer risk. Oncotarget. 7:55611–55623. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gilkes DM, Pan Y, Coppola D, Yeatman T,
Reuther GW and Chen J: Regulation of MDMX expression by mitogenic
signaling. Mol Cell Biol. 28:1999–2010. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ling X, Xu C, Fan C, Zhong K, Li F and
Wang X: FL118 induces p53-dependent senescence in colorectal cancer
cells by promoting degradation of MdmX. Cancer Res. 74:7487–7497.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mandke P, Wyatt N, Fraser J, Bates B,
Berberich SJ and Markey MP: MicroRNA-34a modulates MDM4 expression
via a target site in the open reading frame. PLoS One.
7:e420342012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Markey M and Berberich SJ: Full-length
hdmX transcripts decrease following genotoxic stress. Oncogene.
27:6657–6666. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li C, Ge Q, Liu J, Zhang Q, Wang C, Cui K
and Chen Z: Effects of miR-1236-3p and miR-370-5p on activation of
p21 in various tumors and its inhibition on the growth of lung
cancer cells. Tumour Biol. 39:10104283177108242017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pan XP, Wang HX, Tong DM, Li Y, Huang LH
and Wang C: miRNA-370 acts as a tumor suppressor via the
downregulation of PIM1 in hepatocellular carcinoma. Eur Rev Med
Pharmacol Sci. 21:1254–1263. 2017.PubMed/NCBI
|