Introduction
The treatment of breast cancer depends on its
pathological type and estrogen receptor α (ERα) expression.
Although the incidence of breast cancer has increased year by year
among women in United States from 2004 to 2014, and has begun
affecting younger age groups, the increased attention to (in terms
of scientific studies) and subsequent improvements in the
comprehensive treatment of breast cancer have resulted in decreases
in breast cancer mortality in the United States over the past two
decades (1). Among these
improvements, endocrine therapy in breast cancer has received
attention and has become the focus of numerous studies. A previous
study demonstrated that >70% of patients with breast cancer are
ERα-positive (ERα+), affording clinicians the
opportunity to provide adjuvant endocrine therapy (2). The selective estrogen receptor modulator
tamoxifen (TAM) and aromatase inhibitors, including anastrozole,
exemestane and letrozole, may inhibit disease recurrence in
patients with ERα+ breast cancer, providing evidence of
the involvement of ERα in the pathogenesis of breast cancer
(3,4).
However, a number of patients with breast cancer treated using
endocrine therapy will develop resistance to these drugs, resulting
in disease recurrence and leading to tumor progression. A previous
study indicated that ~40% of patients that received TAM or other
endocrine treatment would experience disease recurrence (5). Prior studies have proposed a variety of
possible mechanisms of TAM resistance in breast cancer, including
ER and growth factor receptor interaction (6), and ER loss and mutation (7). However, the mechanism of endocrine
resistance is complicated, and certain studies have identified that
specific cases of secondary drug-resistant breast cancer remain
ER+ (8,9). ER post-transcriptional modification is
also considered to be one of the important mechanisms of TAM
resistance (10). In addition, the
abnormal expression of microRNAs (miRNAs/miRs) associated with drug
resistance may lead to tumor cell drug resistance and secondary
metastasis.
miRNAs are short, non-coding RNA molecules that
regulate gene expression at the post-transcriptional level by
directly binding to the 3′-untranslated region (3′-UTR) of the
target mRNAs, causing translation inhibition, which is often
accompanied by mRNA decay (11).
Microarray technology is used to detect abnormal expression
profiles of miRNAs in a range of types of cancer, such as breast
cancer. A number of studies have demonstrated that miRNA expression
profiles are associated with tumor grades and patient prognoses
(12,13). In cancer cells, miRNAs have been
divided into oncogenic miRNAs and tumor suppressor miRNAs. Iorio
et al (14) used microarrays
containing all known miRNAs to quantify the expression levels of
miRNAs in 76 breast tumors, and compared the results with those
obtained from 34 normal breast tissue samples. The results
indicated that the expression levels of 29 miRNAs changed
significantly in breast cancer. Additional analysis indicated that
miR-10b, miR-125b, miR-145, miR-21 and miR-155 were necessary to
distinguish between cancerous and normal tissues, with miR-10b,
miR-125b and miR-145 being significantly downregulated, and miR-21
and miR-155 being significantly upregulated (14). Iorio et al (14) demonstrated that these 5 miRNAs
exhibited significant effects on the formation and growth of
tumors, likely because they served a role in promoting or
inhibiting the expression of estrogen and progesterone receptors in
breast cancer. At present, the principle mechanism of TAM
resistance in breast cancer remains unclear, and the mechanism of
miRNA regulation has not been fully elucidated. Therefore, the
present study initially established TAM-resistant breast cancer
cell lines by two different methods, and identified novel avenues
for reversing TAM resistance by using HiSeq 2500 sequencing
technology to screen resistance-associated miRNAs.
The ERα expression level is an important indicator
of prognosis and is associated with breast cancer drug resistance.
It has been demonstrated that miRNA-206 expression is significantly
reduced in ERα+ breast cancer (15); miR-206 may inhibit the expression of
ERα on mRNA and protein levels, indicating that miRNA-206 could be
used as a breast cancer endocrine therapy target (16). In addition, the study of TAM-resistant
breast cancer cell lines has indicated that miR-221/miR-222 was
significantly elevated in the TAM-resistant cell line (17). miR-221/miR-222 inhibited the
expression of tumor suppressor cyclin-dependent kinase inhibitor 1B
(p27kip1), which resulted in p27kip1 decline
(17). With the reduction of
p27kip1, the migratory and invasive abilities of cells
were increased, promoting the metastasis of breast cancer (18). The expression of miR-21 is associated
with ERα. A decrease in miR-21 expression led to the activation of
expression of its target genes, programmed cell death 4 (PDCD4),
phosphatase and tensin homolog and B-cell lymphoma 2 (Bcl-2),
significantly increasing the sensitivity of tumor cells to
aromatase inhibitors (19,20). Concurrently, it was identified that
the expression of miR-21 was positively correlated with the
expression level of receptor tyrosine-protein kinase erbB-2
(HER2/neu). HER2/neu promotes the expression of miR-21 by
activating the mitogen-activated protein kinase extracellular
signal-related kinase 1/2 (MAPK ERK1/2) pathway, and the
upregulation of miR-21 inhibits the expression of its target gene,
PDCD4, a tumor metastasis suppressor protein (21). Therefore, the maintenance of
expression and function of various proteins, including enzymes,
depends on the specific regulation of miRNAs. If miRNA expression
is abnormal, it will disrupt the homeostasis of gene expression,
and lead to the abnormal expression of drug-associated genes, and
finally result in the occurrence of drug resistance. In the present
study, the human breast cancer MCF-7 cell line and two
drug-resistant strains, MCF-7C and MCF-7T, were studied. miRNA
expression was detected using the HiSeq 2500 sequencing system,
identifying miRNAs with differential expression, which provide a
basis for additional functional studies.
In the present study, two different drug-resistant
breast cancer cell lines were established using two different
methods of drug administration. Dose stepwise induction was used in
the establishment of the MCF-7C line. Following each treatment with
progressively increasing concentrations of 4-hydroxytamoxifen (OHT)
screening medium, the drug-containing medium was replaced with
drug-free complete medium, the apoptotic cells were removed, and
the viable cells were allowed to proliferate. This process provided
the appropriate time interval for tumor cells to activate relevant
physiological pathways, including the Erk and PI3K pathways
(10). Therefore, the tumor cells
gradually adapted to the stimulation of drugs, and then obtain drug
resistance. Drug-resistant cell lines established using gradual
increases in drug concentration are considered to have acquired
resistance during the processes of changing and remodeling of cell
proliferation, cell cycle, apoptosis and genetics, consequently
having acquired resistance in the process of screening and
culturing (22). Therefore, we
hypothesize that the MCF-7C line is a cell line with acquired
drug-resistance. The other drug-resistant cell line, MCF-7T, was
established by a temporal stepwise method, beginning with a high
concentration of OHT. The culture time gradually increased from 1
to 48 h, and then to 72 h. The selection process provided a time
interval that not only facilitated the repair of slightly damaged
cells, but also favored the removal of cells that were relatively
drug-sensitive, while cells that were inherently relatively
resistant were retained. It is considered that a temporal stepwise
method of inducing drug-resistant strains, beginning with high
concentrations, preserved the cells with drug-resistant
characteristics, namely the endogenous drug-resistant cells
(23). Therefore, we hypothesized
that the MCF-7T cell line was a combination of exogenous and
endogenous resistance characteristics.
Materials and methods
Reagents
The human breast cancer MCF-7 cell line was
purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA). Dulbecco's modified Eagle's medium (DMEM),
fetal bovine serum, trypsin and phosphate-buffered saline (PBS)
were all purchased from Hyclone; GE Healthcare Life Sciences
(Logan, UT, USA); OHT was purchased from Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany); EMD Millipore™
Immobilon™ Western Chemiluminescent HRP Substrate (ECL)
was purchased from EMD Millipore (Billerica, MA, USA), and the
rabbit anti-human ER (dilution 1:500; sc-543) and mouse anti-human
GAPDH (dilution 1:500; sc-293335) antibodies were purchased from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Establishment of the TAM-resistant
MCF-7C cell line using an OHT dose stepwise induction method
The MCF-7C cell line was established using OHT as
the screening drug and the two-step screening method based on an
OHT dose stepwise induction. This method was divided into two
phases: Adaptation and consolidation. In the adaptation phase,
MCF-7 cells were cultured to 70% confluence at 37°C in a humidified
atmosphere containing 5% CO2, then treated with 10 nM
OHT-containing DMEM for 72 h three times, with medium being removed
and re-introduced between each step, followed by 20, 50, 100, 200,
500 and 800 nM OHT dose screens. In the consolidation phase, the
cell line that was resistant to 800 nM would be cultured in 1000 nM
OHT medium at 37°C in a humidified atmosphere containing 5%
CO2 for 72 h and the medium was removed and
re-introduced ten times. PBS was used for immediate washing
following every change of medium. Following sequential treatment
and culturing for a period of 1 year, until the cells were
normalized in the selection culture medium (1 µM OHT), the MCF-7C
cell line was considered to be successfully established.
Establishment of the MCF-7T cell line
(temporal stepwise induction with high concentration of OHT)
The MCF-7T cell line was established by temporal
stepwise induction, beginning with a high concentration of OHT,
using the breast cancer MCF-7 cell line as the parental cell line
and OHT as the screening agent. Again, the method was divided into
two stages, adaptation and consolidation. MCF-7 cells were cultured
for 1 h in 1 µM OHT-containing DMEM three times, with medium being
removed and re-introduced between each step, in total at 37°C in a
humidified atmosphere containing 5% CO2. Then, culture
times of 2, 4, 12, 24 and 48 h were sequentially performed in 1 µM
OHT medium three times at 37°C. In the consolidation stage, the
cells obtained from the adaptation stage were cultured in 1 µM OHT
medium for 72 h and the 1 µM medium was removed and re-introduced
10 times. PBS was used for immediate washing following every medium
change. MCF-7T cell line was successfully established by continuous
culture with 1 µM OHT.
Expression of ER protein was detected
by western blot analysis
Proteins from MCF-7 cells and the drug-resistant
sub-clones were harvested using immunoprecipitation assay lysis
buffer (Beyotime Institute of Biotechnology, Haimen, China) and
protein samples were prepared through sonication at 20 KHz and the
concentration was determined by BCA method. Protein samples (50 µg)
were subjected to 8% SDS-PAGE, and the proteins were then
transferred from the gel to a polyvinylidene fluoride membrane, the
membrane was next blocked in TBS with Tween-20 with 5% skim milk
(BD Biosciences, Franklin Lakes, NJ, USA) at room temperature for 2
h, followed by incubation with the ER antibody (dilution 1:500) at
4°C overnight then with the mouse anti-rabbit secondary antibody
sc-2357 (dilution 1:500; Santa Cruz Biotechnology, Inc.) at room
temperature for 2 h. Bands were detecting using the ECL reagent,
developed with X-ray film and scanned using Gel Doc™ XR+
Imaging System (Bio-Rad Laboratories, Inc., Hercules, CA, USA) to
obtain image data. Finally, the expression level of the target
protein was obtained through AlphaEaseFC version 6.0
(ProteinSimple, San Jose, CA, USA) image analysis.
Total cellular RNA extraction
Cells in the log growth phase were harvested and
TRIzol® (Life Technologies; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) was used to isolate RNA at 4°C according to
the manufacturer's instructions. The separated RNA was dissolved in
diethyl pyrocarbonate water and diluted to 500 ng/µl. The
concentration and purity of RNA was quantified using a NanoDrop
2000 Ultraviolet Spectrophotometer (NanoDrop Technologies; Thermo
Fisher Scientific, Inc., Pittsburgh, PA, USA). Samples with an
optical density (OD)260/OD280 ratio of 1.8–2.0 were considered to
have met the requirements.
Library construction and
sequencing
Small RNA high-throughput sequencing was performed
by Beijing Biomarker Biotechnology Co., Ltd. (Beijing, China), and
the library was constructed and sequenced as follows: 1.5 µg RNA
was used as the initial amount of the RNA sample and the volume of
the sample was supplemented with water to make a total reaction
volume of 6 µl. Next, the library was constructed using the small
RNA Sample Prep kit (New England Biolabs, Ltd., Beijing, China),
according to the manufacturer's instructions. Subsequent to the
construction of the library, the concentration of the samples was
determined by Qubit 2.0 (Life Technologies; Thermo Fisher
Scientific, Inc.) and the samples within the library were diluted
to a concentration of 1 ng/µl. The insert size was then examined
using the Agilent 2100 Bioanalyzer (Agilent Technologies, Inc.,
Santa Clara, CA, USA). High-throughput sequencing of the library
was performed using the HiSeq 2500 library with sequencing read
length single-end (SE) of 50 nt at Biomarker Biotechnology Co.,
Ltd.
miRNA analysis and identification
As miRNA transcription initiation sites are
primarily located in the gene spacer, intron and reverse complement
sections of the coding sequence, the precursors have a marked
hairpin structure that is formed by the cleavage of the
endoribonuclease Dicer/Dicer-like enzyme (24). On the basis of the distribution of
length of reads on the precursor sequence (based on the
characteristics of the miRNA, mature, star, loop sections) and the
energy information of the precursor structure, which is based on
the sequence of reads (RNA-fold), miRDeep2 software version 2.0.0.8
(Max Delbrück Center for Molecular Medicine, Berlin, Germany) was
used to identify known and novel miRNAs.
Differential expression profiling of
miRNA
miRDeep2 software was used to identify known miRNAs
and predict novel miRNAs by alignment to the reference genome
GRCh37 (http://grch37.ensembl.org/Homo_sapiens/Info/Index).
The expression of miRNAs in each sample was analyzed statistically
using EBseq R package version 1.19.0 (Bioconductor; https://www.bioconductor.org/help/) and the
expression was normalized using a Transcript Per Million algorithm
(25). In the process of differential
expression miRNA detection, |log2 fold change (FC)| ≥1
and false discovery rate (FDR) ≤0.01 were used as the screening
criteria. The FC represents the ratio of expression between two
samples (groups). The P-value of the original hypothesis may be
expressed as the probability of non-discrepancy. As the
differential expression analysis of miRNAs is an independent
statistical hypothesis test for a large number of miRNA expression,
there will be a false-positive issue. Therefore, in the process of
analysis, the Benjamini-Hochberg correction method was used to
calculate the P-value, and the FDR was used as the key indicator of
differential expression profiling of miRNA to identify the
differentially expressed genes (DEGs). Hierarchical cluster
analysis of DEGs was performed using TMeV version 4.9.0 (GitHub,
Inc., San Francisco, CA, USA).
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical differences between groups in western blot analysis
were assessed using analysis of variance with a post hoc least
significant difference test on SPSS version 22 (IBM Corp., Armonk,
NY, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
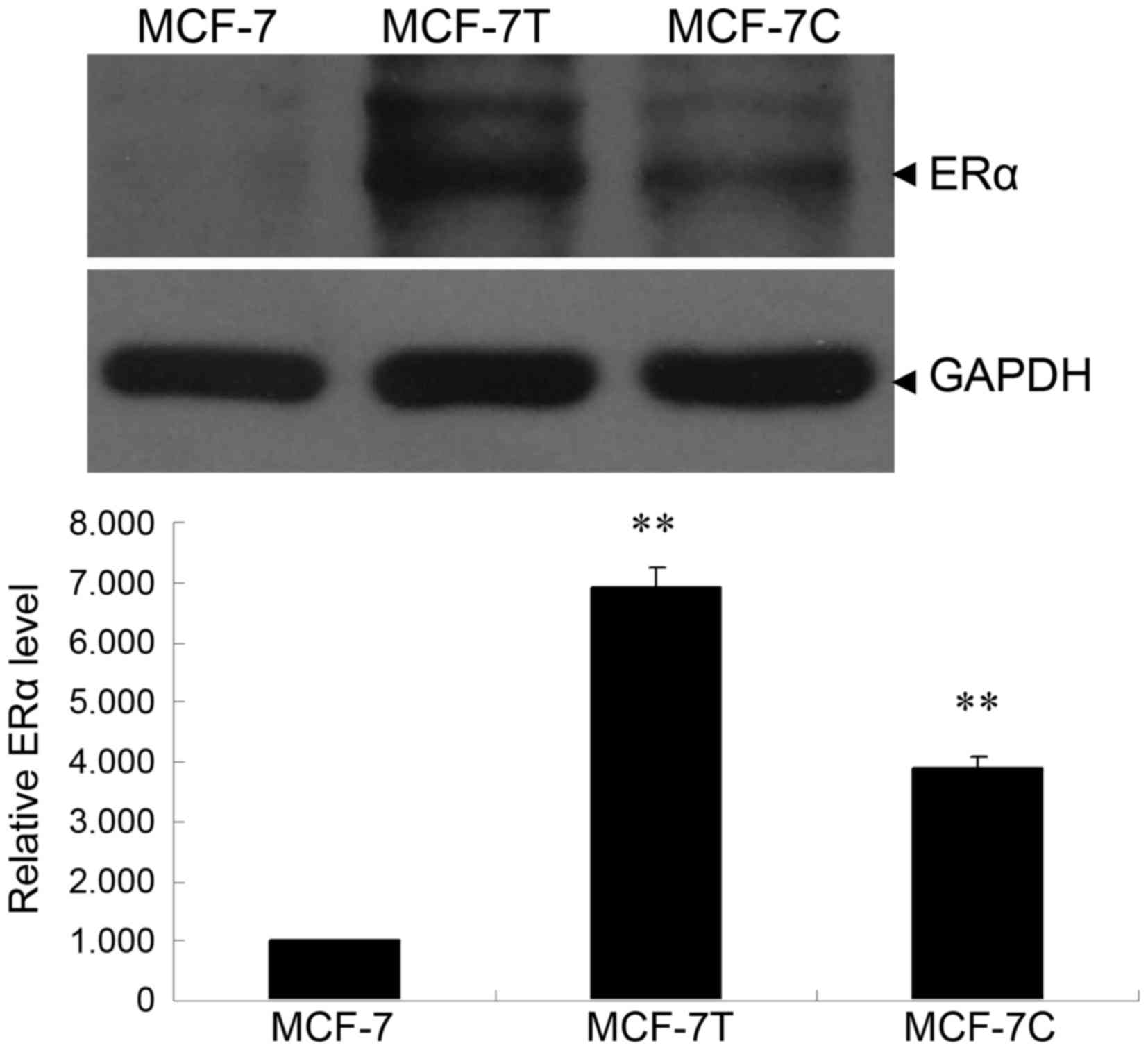
Western blotting detects ERα protein
in MCF-7, MCF-7C and MCF-7T cells
The protein expression of ERα was detected by
western blotting. The expression of ERα in MCF-7C and MCF-7T cells
was significantly increased, compared with MCF-7 cells (Fig. 1; P<0.01).
MiRNA differential expression
analysis
In the present study, the expression levels of
miRNAs in MCF-7 cells and TAM-resistant breast cancer cell lines
were assessed using HiSeq 2500 sequencing technology. The results
indicated that 1,646 miRNAs were detected in the 3 cell samples,
including 1,376 known miRNAs and 270 predicted miRNAs. In the
process of differential expression miRNA detection,
|log2 FC| ≥1 and FDR ≤0.01 was used as the screening
criteria. The results indicated that there were 118 miRNAs that
exhibited significant differences in expression between MCF-7C and
MCF-7 cells (P<0.05); among them, there were 67 upregulated and
51 downregulated miRNAs. Upregulated miRNAs in MCF-7C included
let-7c, let-7e, miR-21 and miR-486, whereas downregulated miRNAs
included miR-146a, miR-148a and miR-342. There were 42 miRNAs that
exhibited significant differences in expression between MCF-7T and
MCF-7 (P<0.05); among them there were 23 upregulated and 19
downregulated miRNAs. Upregulated miRNAs in MCF-7T included let-7i,
miR-145, miR-199b and miR-486, whereas downregulated miRNAs
included miR-146a and miR-27a. Compared with the drug-resistant
MCF-7T cell line, 126 miRNAs were expressed at significantly
different levels in MCF-7C cells (P<0.05); among them there were
76 upregulated and 50 downregulated miRNAs. Upregulated miRNAs in
MCF-7C included let-7c-3p and let-7e-5p, whereas the downregulated
miRNAs included miR-145-5p, miR-148a-5p and miR-199b. The number of
DEGs is summarized in Table I.
 | Table I.Differentially expressed miRNA. |
Table I.
Differentially expressed miRNA.
| DEG group | Total DEG | Upregulated | Downregulated |
|---|
| MCF-7C vs.
MCF-7 | 118 | 67 | 51 |
| MCF-7T vs.
MCF-7 | 42 | 23 | 19 |
| MCF-7C vs.
MCF-7T | 126 | 76 | 50 |
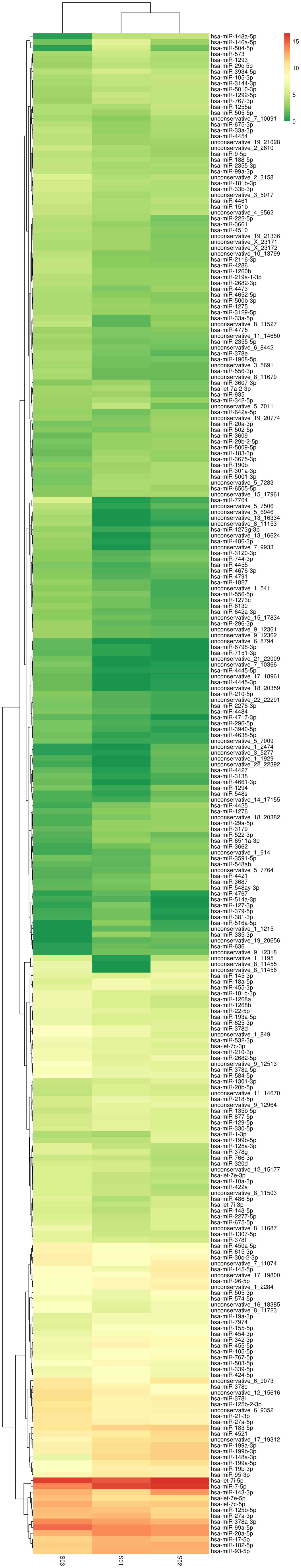
The differentially expressed miRNAs exhibiting the
same or similar expression behavior were clustered by hierarchical
cluster analysis. The cluster results of differentially expressed
miRNAs are presented in Fig. 2.
Certain differentially expressed miRNAs in the two drug-resistant
breast cancer cell lines are summarized in Table II.
| Table II.Expression of certain miRNAs in two
drug-resistant breast cancer cell lines. |
Table II.
Expression of certain miRNAs in two
drug-resistant breast cancer cell lines.
|
| MCF-7C vs.
MCF-7 | MCF-7T vs.
MCF-7 |
|---|
|
|
|
|
|---|
| DEG | log2FC | P-value | Regulated | log2FC | P-value | Regulated |
|---|
| Let-7c | 1.1522 | 0 | Up | – | – | – |
| Let-7e | 1.3271 | 0 | Up | – | – | – |
| Let-7i | 1.0847 | 0 | Up | 1.2287 | 0 | Up |
| miR-145-5p | – | – | – | 2.7938 | 0 | Up |
| miR-486-5p | 1.9927 | 0 | Up | 1.3708 | 0 | Up |
| miR-21-3p | 1.1983 | 0 | Up | – | – | – |
| miR-146a-5p | −4.2962 | 0 | Down | −3.0789 | 0 | Down |
| miR-148a-5p | −6.1588 | 0 | Down | – | – | – |
| miR-27a | – | – | – | −1.0310 | 0 | Down |
| miR-199b | −1.4658 | 0 | Down | 2.1652 | 0 | Up |
Discussion
miRNAs have become the focus of current oncology
studies. A previous study have identified that miRNA may regulate
gene expression, and affect the development, metabolism and other
physiological processes of disease (11). microRNAs are short non-coding RNA
molecules that regulate gene expression at post-transcriptional
levels by directly targeting the 3′-UTR of target mRNAs. These
molecules participate in the regulation of the signaling pathways
and genes involved in tumorigenesis and progression, thereby
affecting cellular processes such as differentiation, migration and
apoptosis (26). A large number of
studies have indicated that miRNAs may regulate the expression of
oncogenes or tumor suppressor genes, and serve the roles of
oncogenes or tumor suppressor genes (27). miRNAs are aberrantly expressed in a
variety of cancer cells, including those of breast cancer (28). Previously, studies have indicated that
miRNAs are not only closely associated with the occurrence and
development of tumors, but also serve a notable role in the
development of drug resistance (29).
It is speculated that the differential expression of miRNA may also
be involved in the regulation of TAM resistance in breast cancer.
At present, little is known about miRNAs in TAM-resistant breast
cancer. miRNA detection methods include miRNA microarrays and small
RNA sequencing. Compared with microRNA microarrays, microRNA
sequencing has a number of advantages, including increased data
throughput capacity, higher sensitivity and accuracy, wider dynamic
detection range, more abundant content of bioinformatics analysis
and the ability to detect unknown microRNAs (30).
The let-7 miRNA family was the second miRNA to be
identified (31), and is one of the
most widely-studied miRNAs. The let-7 family is highly conserved in
terms of sequence and functions, and its expression in tumor cells
is usually reduced (32). Let-7 may
regulate the epithelial-mesenchymal transition (EMT) of tumor cells
(33). Low let-7 expression is
usually characterized by mesenchymal cells. High expression of
let-7 is a characteristic of epithelial cells. The expression of
let-7 is closely associated with the malignancy of tumors. As a
tumor suppressor gene, it serves an important role in cell growth,
differentiation and apoptosis, primarily through the regulation of
proto-oncogenes or transcription factors (34). For example, a decrease in let-7
expression in breast cancer stem cells results in an overexpression
of the RAS proto-oncogenes (35).
Estrogen receptor signaling may induce the expression of let-7a,
while downregulating the expression of its target genes, v-myc
avian myelocytomatosis viral oncogene homolog MYC and K-Ras,
inhibits tumor cell proliferation (36). Chen et al (20) identified that the expression of ERα-36
(a 36-kDa novel isoform of ER-α66) was downregulated by let-7 in
mammary glands, which decreased the phosphorylation of ERK and RAC
serine/threonine-protein kinase (Akt), and affected the MAPK/ERK
and phosphoinositide 3-kinase (PI3K)/Akt signaling pathways. In
TAM-resistant breast cancer cells, increasing the expression of
let-7 may reduce the expression of ER-α36, thereby enhancing the
sensitivity of tumor cells to drugs (37). However, the high expression of let-7
in the MCF-7C and MCF-7T TAM-resistant cell lines is not consistent
with the previous study (37). The
high expression of let-7 may be explained by high ERα protein
expression; however, this mechanism in tumor cell resistance
requires additional study and verification.
It has been demonstrated that miR-21 serves as a
proto-oncogene and inhibits the expression of the pro-apoptotic
gene Bcl-2, thus affecting tumor growth (19). In addition, miR-21 may regulate the
metastasis and infiltration of cells by regulating a variety of
tumor metastasis suppressor genes such as PDCD4 and tropomyosin 1α
(38). The role of miR-21 in tumor
drug resistance has attracted attention. Blower et al
(39) demonstrated that the
expression of miR-21 was associated with chemotherapeutic efficacy,
and that reducing miR-21 levels improved the efficacy of multiple
drugs, indicating that miR-21 expression is associated with drug
resistance in breast cancer cells. miR-21 is highly expressed in
adriamycin-resistant breast cancer cells, and the upregulation or
downregulation of miR-21 may affect the sensitivity of breast
cancer cells to doxorubicin (40).
The results of the present study indicated that miR-21 expression
in TAM-resistant cells was significantly increased. This is
consistent with the findings of previous study, which demonstrated
that estradiol inhibits miR-21 expression and estrogen antagonists
may increase miR-21 expression, indicating that miR-21 and ER may
interact with each other (19).
Therefore, decreasing the expression of miR-21 may be a novel
method of improving the treatment of patients with breast cancer.
Lee et al (41) and Ota et
al (42) suggested that there was
a significant positive association between miR-21 expression and
HER-2 status. As a commonly used marker for breast cancer, HER-2 is
believed to promote tumor metastasis. Huang et al (21) demonstrated that HER-2 may increase the
expression of miR-21 through the MAPK signaling pathway, promoting
cell invasion. As ER and HER-2 are important indicators of
endocrine therapy for breast cancer, indicating that miR-21 could
be used as a predictor of endocrine therapy.
Previous studies demonstrated that miR-486 is
downregulated in the majority of tumors (43–45). Zhang
et al (46) identified that
miR-486 was significantly downregulated in breast cancer tissues
and cell lines. The overexpression of miR-486 may significantly
inhibit the proliferation of breast cancer cells, induce
G0/G1 arrest and promote cell apoptosis.
Additionally, it was confirmed that the overexpression of
proto-oncogene serine-threonine-protein kinase (PIM-1) attenuates
the inhibitory effect of miR-486 on breast cancer cells (43). It was deduced that miR-486 may serve
an anti-proliferative role by directly downregulating the
expression of PIM-1 (46). Therefore,
the interaction between the miR-486/PIM-1 axis may contribute to
the understanding of the mechanism of drug resistance in breast
cancer.
It has been identified that the overexpression of
miR-146a in the highly metastatic human breast cancer MDA-MB-231
cell line significantly downregulated interleukin-1
receptor-associated kinase 1 and tumor necrosis factor receptor
associated factor 6 expression, and negatively regulated nuclear
factor-κB (NF-κB) activity, significantly reducing the invasive and
migratory capacity of the cells (47). These data indicate that miR-146a is a
negative regulator of NF-κB activity in human breast cancer cells,
and the low miR-146a expression exhibited in the two drug-resistant
cell lines in the present study indicating that the modulation of
miR-146a levels may have potential therapeutic significance by
inhibiting breast cancer metastasis and drug resistance.
Insulin receptor substrate-1 (IRS-1) and
insulin-like growth factor I receptor (IGF-IR) induce multiple
signaling cascades such as PI3K/AKT and MAPK/ERK, and control tumor
formation and progression (48).
miR-148a may inhibit the proliferation of breast cancer cells by
regulating IRS-1 and IGF-IR. The upregulation of miR-148a may
specifically inhibit the expression of IGF-IR/PKM2. miR-148a may
inhibit tumor metastasis and invasion by the regulation of specific
target genes, including hematopoietic-related PBX interacting
proteins, matrix metalloproteinase 7, ubiquitin-specific protease
4, HOX transcriptional antisense RNA and S-phase kinase-associated
protein 1. miR-148a may inhibit Bcl-2-induced tumor cell apoptosis
(49). In the present study, when the
expression of miR-148a in TAM-resistant cell lines was
downregulated, the tumor inhibition in metastasis and invasion was
weakened and the level of drug resistance was increased.
The results of sequencing in the present study
indicated that miR-342 expression was significantly decreased in
the TAM-resistant cell lines, which is consistent with the results
of a previous study (50). Cittelly
et al (50) identified that
the inhibition of miR-342 expression incurred resistance to TAM in
the breast cancer MCF-7 cell line. Conversely, by increasing the
expression of miR-342, the rate of resistance development to
endocrine therapy may be decreased and rates of apoptosis are
increased in tumor cells (50);
therefore, the tumor cells become sensitive to the drug treatment.
miR-342 is aberrantly expressed in a variety of tumor cell models
and the expression of miR-342 is associated with the expression of
its host gene, Ena/vasodilator-stimulated phosphoprotein-like
(EVL). It was identified that miR-342 was highly expressed in
ERα+ breast cancer cells and decreased in ERα negative
cells (51). However, EVL expression
was significantly lower in ERα+ breast cancer cells that
failed to respond to TAM treatment, compared to TAM-sensitive
ERα+ breast cancer cells (50), so it was hypothesized that TAM-induced
tumor cell resistance may be induced by inhibiting miR-342
expression. miR-342 may serve an important role in promoting cell
apoptosis. miR-342 may be involved in the regulation of the
mitotic-like POLO-like kinase signaling pathway and cell
cycle-associated signaling pathway-associated proteins, such as
cyclin B1 and tumor protein P53 (50). It has also been demonstrated that
miR-342 may affect chemosensitivity by modulating histone
methylation (52). Therefore, miR-342
may affect the sensitivity of MCF-7 cells to TAM, indicating that
miR-342 may serve as a molecular marker for predicting TAM
sensitivity in ER+ breast cancer, and may be a potential
target for endocrine adjuvant therapy.
In addition, the results of the sequencing in the
present study indicated that miR-27a was downregulated in the
TAM-resistant cell lines. Feng et al (53) identified that miR-27a could bind to
the 3′-UTR of the adenosine 5-triphosphate binding cassette
subfamily B member 1 gene and inhibit its expression, and therefore
may directly regulate the expression of the multidrug resistance
protein P-glycoprotein. Concurrently, the overexpression of miR-27a
in leukemia cells may increase chemosensitivity in leukemia cells.
We hypothesized that inducible resistance as a result of low
miR-27a expression in breast cancer cells may be associated with
this mechanism.
Expression of miR-145 was upregulated in the
TAM-resistant cell lines in the present study. Nam et al
(54) suggested that miR-145 may
target the upstream regulatory factors IGF-1R and signal transducer
and activator of transcription 1 (STAT1) in glycolysis, thereby
affecting cell metabolism and the expression of the Janus
kinase/STAT signaling pathway, consequently regulating EMT and
tumor metastasis.
The drug-resistant MCF-7C and MCF-7T cell lines
exhibited different miRNA expression profiles, indicating that
these drug-resistant cell lines have different molecular mechanisms
of drug resistance. As they were established using different modes
of drug administration, we hypothesize that TAM administration in
the treatment of patients with breast cancer has an effect on the
formation of drug resistance. Silencing of miR-199b-5p due to
epigenetic changes, including promoter CpG island methylation,
activates the JAG1-mediated Notch1 signaling cascade, leading to
the development of acquired resistance in ovarian cancer (55). The expression of miR-199b-5p was
downregulated in the MCF-7C cell line and upregulated in the MCF-7T
cell line, indicating that MCF-7C may be a cell line that acquired
drug-resistance by a dose stepwise induction method, which was
consistent with a previous study (55). These results indicate that miR-199b-5p
may be a novel therapeutic target for breast cancer therapy.
In the present study, drug resistance in MCF-7C
cells was established by dose stepwise induction and in MCF-7T
cells by a temporal stepwise method. The mechanism of drug
resistance exhibited by MCF-7C cells may differ from that of MCF-7T
cells. MCF-7C may be an acquired drug resistant cell line, and
MCF-7T may be a combination of acquired resistance cells and
endogenous resistant cells. A clearer and more detailed drug
resistance mechanism requires further characterization. However, at
present the two cell lines may only be distinguished by clinical
concepts; there is a lack of specific criteria to determine whether
in vitro-established drug-resistant cell lines exhibit
endogenous or acquired resistance. The establishment of
drug-resistant cancer cell lines is designed to provide the correct
resources for overcoming clinical drug resistance to breast cancer.
Therefore, the specific association between endogenous and acquired
resistance requires additional study. The establishment of
appropriate drug-resistant cancer cell lines will provide useful
information for elucidating the mechanisms of tumor drug
resistance, and thus methods for clinical treatment.
The expression of ER in MCF-7C and MCF-7T cells was
significantly increased compared with that in the MCF-7 cells,
indicating that the acquisition of TAM resistance in tumor cells
may be independent of ER loss. On the basis of the results of the
present and previous studies, we hypothesize that miR-21, miR-146a,
miR-148a, miR-34a and miR-27a serve important roles in TAM
resistance in breast cancer, and may be potential targets for
breast cancer treatment. The roles of miR-486, miR-505, miR-29 and
miR-145 require additional study. In the present study, the
differentially expressed miRNAs associated with drug resistance in
breast cancer cells were obtained by RNA sequencing technology,
providing data for future studies and a basis for the
identification of the mechanism of drug resistance and reversal of
tumor drug resistance.
Acknowledgements
The present study was supported by grants from
National Natural Science Foundation of China (grant nos. 81272372
and 30873044). The Project-sponsored by Scientific Research
Foundation for Returned Overseas Chinese Scholars, State Education
Ministry.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rao X, Di Leva G, Li M, Fang F, Devlin C,
Hartman-Frey C, Burow ME, Ivan M, Croce CM and Nephew KP:
MicroRNA-221/222 confers breast cancer fulvestrant resistance by
regulating multiple signaling pathways. Oncogene. 30:1082–1097.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wolmark N, Mamounas EP, Baehner FL, Butler
SM, Tang G, Jamshidian F, Sing AP, Shak S and Paik S: Prognostic
impact of the combination of recurrence score and quantitative
estrogen receptor expression (ESR1) on predicting late distant
recurrence risk in estrogen receptor-positive breast cancer after 5
years of Tamoxifen: Results from NRG oncology/national surgical
adjuvant breast and bowel project B-28 and B-14. J Clin Oncol.
34:2350–2358. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ingle JN: Postmenopausal women with
hormone receptor-positive breast cancer: Balancing benefit and
toxicity from aromatase inhibitors. Breast. 22 Suppl 2:S180–S183.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Manavalan TT, Teng Y, Appana SN, Datta S,
Kalbfleisch TS, Li Y and Klinge CM: Differential expression of
microRNA expression in tamoxifen-sensitive MCF-7 versus
tamoxifen-resistant LY2 human breast cancer cells. Cancer Lett.
313:26–43. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Arpino G, Wiechmann L, Osborne CK and
Schiff R: Crosstalk between the estrogen receptor and the HER
tyrosine kinase receptor family: Molecular mechanism and clinical
implications for endocrine therapy resistance. Endocr Rev.
29:217–233. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ciupek A, Rechoum Y, Gu G, Gelsomino L,
Beyer AR, Brusco L, Covington KR, Tsimelzon A and Fuqua SA:
Androgen receptor promotes tamoxifen agonist activity by activation
of EGFR in ERalpha-positive breast cancer. Breast Cancer Res Treat.
154:225–237. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Karam M, Bièche I, Legay C, Vacher S,
Auclair C and Ricort JM: Protein kinase D1 regulates ERα-positive
breast cancer cell growth response to 17β-estradiol and contributes
to poor prognosis in patients. J Cell Mol Med. 18:2536–2552. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Musgrove EA and Sutherland RL: Biological
determinants of endocrine resistance in breast cancer. Nat Rev
Cancer. 9:631–643. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ma L, Teruya-Feldstein J and Weinberg RA:
Tumour invasion and metastasis initiated by microRNA-10b in breast
cancer. Nature. 449:682–688. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cancer Genome Atlas Network, .
Comprehensive molecular portraits of human breast tumours. Nature.
490:61–70. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Iorio MV, Ferracin M, Liu CG, Veronese A,
Spizzo R, Sabbioni S, Magri E, Pedriali M, Fabbri M, Campiglio M,
et al: MicroRNA gene expression deregulation in human breast
cancer. Cancer Res. 65:7065–7070. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kondo N, Toyama T, Sugiura H, Fujii Y and
Yamashita H: miR-206 expression is down-regulated in estrogen
receptor alpha-positive human breast cancer. Cancer Res.
68:5004–5008. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Adams BD, Furneaux H and White BA: The
micro-ribonucleic acid (miRNA) miR-206 targets the human estrogen
receptor-alpha (ERalpha) and represses ERalpha messenger RNA and
protein expression in breast cancer cell lines. Mol Endocrinol.
21:1132–1147. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Miller TE, Ghoshal K, Ramaswamy B, Roy S,
Datta J, Shapiro CL, Jacob S and Majumder S: MicroRNA-221/222
confers tamoxifen resistance in breast cancer by targeting
p27Kip1. J Biol Chem. 283:29897–29903. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yuan Y, Qin L, Liu D, Wu RC, Mussi P, Zhou
S, Songyang Z and Xu J: Genetic screening reveals an essential role
of p27kip1 in restriction of breast cancer progression.
Cancer Res. 67:8032–8042. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wickramasinghe NS, Manavalan TT, Dougherty
SM, Riggs KA, Li Y and Klinge CM: Estradiol downregulates miR-21
expression and increases miR-21 target gene expression in MCF-7
breast cancer cells. Nucleic Acids Res. 37:2584–2595. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen Z, Yuan YC, Wang Y, Liu Z, Chan HJ
and Chen S: Down-regulation of programmed cell death 4 (PDCD4) is
associated with aromatase inhibitor resistance and a poor prognosis
in estrogen receptor-positive breast cancer. Breast Cancer Res
Treat. 152:29–39. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang TH, Wu F, Loeb GB, Hsu R,
Heidersbach A, Brincat A, Horiuchi D, Lebbink RJ, Mo YY, Goga A and
McManus MT: Up-regulation of miR-21 by HER2/neu signaling promotes
cell invasion. J Biol Chem. 284:18515–18524. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Valeri N, Gasparini P, Braconi C, Paone A,
Lovat F, Fabbri M, Sumani KM, Alder H, Amadori D, Patel T, et al:
MicroRNA-21 induces resistance to 5-fluorouracil by down-regulating
human DNA MutS homolog 2 (hMSH2). Proc Natl Acad Sci USA. 107:pp.
21098–21103. 2010; View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang LY and Trujillo JM: Biological
characterization of multidrug-resistant human colon carcinoma
sublines induced/selected by two methods. Cancer Res. 50:3218–3225.
1990.PubMed/NCBI
|
|
24
|
Ha M and Kim VN: Regulation of microRNA
biogenesis. Nat Rev Mol Cell Biol. 15:509–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fahlgren N, Howell MD, Kasschau KD,
Chapman EJ, Sullivan CM, Cumbie JS, Givan SA, Law TF, Grant SR,
Dangl JL and Carrington JC: High-throughput sequencing of
Arabidopsis microRNAs: Evidence for frequent birth and death of
MIRNA genes. PLoS One. 2:e2192007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hammond SM: RNAi, microRNAs, and human
disease. Cancer Chemother Pharmacol. 58 Suppl 1:S63–S68. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bajan S and Hutvagner G: Regulation of
miRNA processing and miRNA mediated gene repression in cancer.
Microrna. 3:10–17. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nygaard S, Jacobsen A, Lindow M, Eriksen
J, Balslev E, Flyger H, Tolstrup N, Møller S, Krogh A and Litman T:
Identification and analysis of miRNAs in human breast cancer and
teratoma samples using deep sequencing. BMC Med Genomics. 2:352009.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yahya SM and Elsayed GH: A summary for
molecular regulations of miRNAs in breast cancer. Clin Biochem.
48:388–396. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rai G, Rai R, Saeidian AH and Rai M:
Microarray to deep sequencing: Transcriptome and miRNA profiling to
elucidate molecular pathways in systemic lupus erythematosus.
Immunol Res. 64:14–24. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Reinhart BJ, Slack FJ, Basson M,
Pasquinelli AE, Bettinger JC, Rougvie AE, Horvitz HR and Ruvkun G:
The 21-nucleotide let-7 RNA regulates developmental timing in
Caenorhabditis elegans. Nature. 403:901–906. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dahiya N, Sherman-Baust CA, Wang TL,
Davidson B, Shih IeM, Zhang Y, Wood W III, Becker KG and Morin PJ:
MicroRNA expression and identification of putative miRNA targets in
ovarian cancer. PLoS One. 3:e24362008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jin B, Wang W, Meng XX, Du G, Li J, Zhang
SZ, Zhou BH and Fu ZH: Let-7 inhibits self-renewal of
hepatocellular cancer stem-like cells through regulating the
epithelial-mesenchymal transition and the Wnt signaling pathway.
BMC Cancer. 16:8632016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shell S, Park SM, Radjabi AR, Schickel R,
Kistner EO, Jewell DA, Feig C, Lengyel E and Peter ME: Let-7
expression defines two differentiation stages of cancer. Proc Natl
Acad Sci USA. 104:pp. 11400–11405. 2007; View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sun X, Fan C, Hu LJ, Du N, Xu CW and Ren
H: Role of let-7 in maintaining characteristics of breast cancer
stem cells. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 28:789–792.
2012.(In Chinese). PubMed/NCBI
|
|
36
|
Barekati Z, Radpour R, Lu Q, Bitzer J,
Zheng H, Toniolo P, Lenner P and Zhong XY: Methylation signature of
lymph node metastases in breast cancer patients. BMC Cancer.
12:2442012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao Y, Deng C, Lu W, Xiao J, Ma D, Guo M,
Recker RR, Gatalica Z, Wang Z and Xiao GG: let-7 microRNAs induce
tamoxifen sensitivity by downregulation of estrogen receptor α
signaling in breast cancer. Mol Med. 17:1233–1241. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Qi L, Bart J, Tan LP, Platteel I, Sluis
Tv, Huitema S, Harms G, Fu L, Hollema H and Berg Av: Expression of
miR-21 and its targets (PTEN, PDCD4, TM1) in flat epithelial atypia
of the breast in relation to ductal carcinoma in situ and invasive
carcinoma. BMC Cancer. 9:1632009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Blower PE, Chung JH, Verducci JS, Lin S,
Park JK, Dai Z, Liu CG, Schmittgen TD, Reinhold WC, Croce CM, et
al: MicroRNAs modulate the chemosensitivity of tumor cells. Mol
Cancer Ther. 7:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang ZX, Lu BB, Wang H, Cheng ZX and Yin
YM: MicroRNA-21 modulates chemosensitivity of breast cancer cells
to doxorubicin by targeting PTEN. Arch Med Res. 42:281–290. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lee JA, Lee HY, Lee ES, Kim I and Bae JW:
Prognostic implications of microRNA-21 overexpression in invasive
ductal carcinomas of the breast. J Breast Cancer. 14:269–275. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ota D, Mimori K, Yokobori T, Iwatsuki M,
Kataoka A, Masuda N, Ishii H, Ohno S and Mori M: Identification of
recurrence-related microRNAs in the bone marrow of breast cancer
patients. Int J Oncol. 38:955–962. 2011.PubMed/NCBI
|
|
43
|
Tian F, Shen Y, Chen Z, Li R, Lu J and Ge
Q: Aberrant miR-181b-5p and miR-486-5p expression in serum and
tissue of non-small cell lung cancer. Gene. 591:338–343. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Shindo Y, Hazama S, Nakamura Y, Inoue Y,
Kanekiyo S, Suzuki N, Takenouchi H, Tsunedomi R, Nakajima M, Ueno
T, et al: miR-196b, miR-378a and miR-486 are predictive biomarkers
for the efficacy of vaccine treatment in colorectal cancer. Oncol
Lett. 14:1355–1362. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ren C, Chen H, Han C, Fu D, Zhou L, Jin G,
Wang F, Wang D, Chen Y, Ma L, et al: miR-486-5p expression pattern
in esophageal squamous cell carcinoma, gastric cancer and its
prognostic value. Oncotarget. 7:15840–15853. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang G, Liu Z, Cui G, Wang X and Yang Z:
MicroRNA-486-5p targeting PIM-1 suppresses cell proliferation in
breast cancer cells. Tumour Biol. 35:11137–11145. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rusca N and Monticelli S: MiR-146a in
immunity and disease. Mol Biol Int. 2011:4373012011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Madhala-Levy D, Williams VC, Hughes SM,
Reshef R and Halevy O: Cooperation between Shh and IGF-I in
promoting myogenic proliferation and differentiation via the
MAPK/ERK and PI3K/Akt pathways requires Smo activity. J Cell
Physiol. 227:1455–1464. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Li Y, Deng X, Zeng X and Peng X: The role
of mir-148a in cancer. J Cancer. 7:1233–1241. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cittelly DM, Das PM, Spoelstra NS,
Edgerton SM, Richer JK, Thor AD and Jones FE: Downregulation of
miR-342 is associated with tamoxifen resistant breast tumors. Mol
Cancer. 9:3172010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Farmer P, Bonnefoi H, Becette V,
Tubiana-Hulin M, Fumoleau P, Larsimont D, Macgrogan G, Bergh J,
Cameron D, Goldstein D, et al: Identification of molecular apocrine
breast tumours by microarray analysis. Oncogene. 24:4660–4671.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Jordan MA and Wilson L: Microtubules as a
target for anticancer drugs. Nat Rev Cancer. 4:253–265. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Feng D: Screening and functional studies
of miRNAs associated with drug-resistance and relapse in leukemia.
Journal. 2011.
|
|
54
|
Nam S, Long X, Kwon C, Kim S and Nephew
KP: An integrative analysis of cellular contexts, miRNAs and mRNAs
reveals network clusters associated with antiestrogen-resistant
breast cancer cells. BMC Genomics. 13:7322012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Liu MX, Siu MK, Liu SS, Yam JW, Ngan HY
and Chan DW: Epigenetic silencing of microRNA-199b-5p is associated
with acquired chemoresistance via activation of JAG1-Notch1
signaling in ovarian cancer. Oncotarget. 5:944–958. 2014.
View Article : Google Scholar : PubMed/NCBI
|